
Abstract
Plants have different strategies to avoid selfing and buffer its negative consequences on plant fitness. One strategy is the arrangement of petals and the disposition of the reproductive structures (RS) inside the flowers, allowing the development of different pollination mechanisms. In Calceolaria L. species two possible floral phenotypes can be found: short RS protected by the upper corolla lip (nototribic flowers) and long RS resting in the lower corolla lip (sternotribic flowers), the latter being hypothesized to favor selfing.
We selected 13 Calceolaria taxa and characterized their floral phenotype as nototribic or sternotribic, measured RS length and herkogamy, and performed hand-pollination treatments to determine the number of seeds produced by self- and cross-pollination to test whether floral phenotype influences inbreeding. GLMs analysis was performed to determine the differences between the sizes of RS and both floral phenotypes, and LMM was performed to evaluate the relationship between the RS and inbreeding with both floral phenotypes.
We found a relationship between stamen length and herkogamy in both floral phenotypes, where sternotribic flowers have a higher stamen length and lower herkogamy, whereas the opposite occurred in taxa with nototribic morphology. Stamen length significantly influences the inbreeding with sternotribic flowers having a higher inbreeding depression by geitonogamous self-pollination than nototribic ones.
Our results suggest that plants may evolve different reproductive mechanisms to cope with pollination unreliability. Thus, floral phenotype may favor the development of geitonogamy selfing, which may explain the two floral phenotypes present in this specialized oil-secreting genus.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The evolution of specialized floral traits has been viewed as a reflection of natural selection favoring cross-pollination (Fenster and Martén-Rodríguez, 2007). Flowers play a crucial role in attracting pollinators, potentially leading to reproductive isolation in order to attract those pollinators with specific traits that are able to efficiently transfer the pollen. Some of these floral specializations restrict the access to rewards, as well as the size of pollinators, as determined by the size and shape of the flowers (Pauw, 2006; Muchhala, 2007). Another floral specialization is related to the attachment location of the pollen grain on the pollinator’s body, either sternotribic (i.e., ventral placement) or nototribic (i.e., dorsal placement) (Westerkamp & Claßen-Bockhoff, 2007; Rani & Raju, 2020).
While pollinator specialization can confer an evolutionary advantage through efficient pollen transfer, it may also be disadvantageous when pollinators are scarce or ephemeral (Marten-Rodríguez & Fenster, 2010). Thus, specialized plants are more vulnerable to fluctuations in pollinator availability than generalists (Morgan & Wilson, 2005). Consequently, specialized plants have evolved alternative reproductive mechanisms to mitigate this scenario (Bond, 1994). One of these mechanisms involves autonomous selfing, which provides reproductive assurance in self-compatible lineages when pollinators are unreliable (Eckert et al., 2006; Opedal et al., 2016 ). Depending on the degree of pollinator unreliability, three modes of autonomous selfing may be favored: prior selfing (i.e., before anthesis, with spatial and developmental overlap of female and male functions), competing (i.e., before the arrival of outcrossing pollen), and delayed selfing (i.e., at the end of the flower’s lifespan) (Lloyd & Schoen, 1992). Of these modes, delayed selfing is advantageous because it does not cause pollen or seed discounting (Schoen & Brown, 1991; Lloyd & Schoen, 1992; Schoen and Lloyd, 1992; but see Vaughton & Ramsey, 2010).
Despite the advantage that reproductive assurance mechanism could have on plant fertilization, several studies have shown that selfing can result in inbreeding depression (Takebayashi & Morrell, 2001; Goldberg et al., 2010; Siopa et al., 2020; Igic & Busch, 2013 as cited in Zhang et al., 2021) due to increased homozygosity in subsequent generations, leading to a reduction in the survival and fertility of selfing offspring (Goodwillie et al., 2005; Charlesworth & Willis, 2009). Consequently, various adaptations have emerged to avoid inbreeding depression (Charlesworth & Willis, 2009) and mitigate pollen-stigma interference (Webb & Lloyd, 1986; Dai & Galloway, 2011), including temporal/spatial separation of reproductive organs, gynoecium/androecium arrangement, or parts of the perianth (Fetscher, 2001; Barrett, 2002). Among these adaptations, herkogamy, defined as the spatial separation between anthers and stigmas, is an important trait for reducing autonomous pollination (Opedal, 2018). On the other hand, floral arrangements can also limit pollen deposition by orienting the anthers and styles in a way that prevents pollen from being deposited on the same plant’s stigmatic surface. Thus, plants rely on vectors to ensure adequate pollen transfer and promote outcrossing. Given that there is wide variation in self- and cross-pollination rates between plant species (Belaoussoff & Shore, 1995; Goodwillie et al., 2005), finding models to explore how floral phenotypes relate to self-pollination in specialized plant species may be crucial to understanding reproduction in specialized systems.
Calceolaria L. (Calceolariaceae) is a genus with interesting characteristics that make it a good model for studying its phenotypic arrangements. This genus belongs to one of eleven plant families in which the flowers secrete oil instead of nectar as a floral reward (Carneiro & Machado, 2023; Possobom & Machado, 2017). Flowers consist of fused petals that form two lips, a smaller upper lip, and a larger lower lip. Both are almost completely closed, the upper lip generally protecting the reproductive structures (stamen and pistil) and the lower lip containing the oil-secreting glands called elaiophore (Ehrhart, 2000). Additionally, two types of corolla arrangements can be observed based on the positioning of reproductive structures, which determines the pollen deposition zone on the pollinators’ body (i.e., dorsal deposition (over the head) or ventral part of the abdomen). In some species, both stamen and stigma rest on the lower lip (sternotribic flower), while in others, both structures are protected by the upper lip (nototribic flower: Sérsic, 1991, 2004; Fig. 1). These differences in stamen position not only determine the place where pollen is deposited, but also the reproductive success of the species. In sternotribic flowers, pollen is deposited in a less optimal location (i.e., the ventral part of the pollinator), making it vulnerable to grooming and reducing the likelihood of accurate pollen transfer to conspecific stigmas (Thorp, 2000). In contrast, nototribic flowers, pollen is deposited in optimal areas, allowing for more precise pollen delivery, favoring pollination efficiency (Wiesenborn, 2019). Therefore, understanding how these adaptations may affect breeding systems can be helpful in elucidating how species cope with changing environmental scenarios, especially those with a high dependence on vectors for pollination.

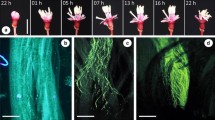
(A) Calceolaria photograph we use to measure the reproductive structures and herkogamy and (B) example illustration of floral phenotypes. Red arrows are indicating the position of reproductive structures in each floral type (style and stamen)
*Note: illustrations were taken from Ehrhart, C. Die Gattung Calceolaria (Scrophulariaceae) in Chile. Biblioth. Bot. 2000, 153, 1–283
Although the genus Calceolaria has both self-incompatible and self-compatible species, it is a highly specialized group of plants that depends entirely on pollinators for its reproduction, being exclusively pollinated by solitary oil-collecting bees of the genera Centris (Hymenoptera: Apidae: Centridini) and Chalepogenus (Hymenoptera: Apidae: Tapinotaspidini), which would be considered generalist pollinators because they visit different plant species for pollen (Molau, 1988; Sérsic, 2004). It has been suggested that floral specialization facilitates outcrossing. However, under certain circumstances (e.g., higher herkogamy), self-compatible species cannot achieve pollination autonomously. Instead, they rely on geitonogamy, where fertilization occurs through pollen from a flower on the same plant, facilitated by an animal vector (Murúa et al., 2014, 2020; Cisterna & Murúa, 2018).
In the present study, we examined differences in the breeding system among Calceolaria taxa. Specifically, we conducted a pollination experiment to compare between geitonogamy (i.e., self-pollination within the same plant) and xenogamy (i.e., cross-pollination between different plants). Additionally, we evaluated whether reproductive structure arrangements and herkogamy are associated with breeding systems and whether there is an inbreeding effect. Furthermore, we examined whether these factors correlate with floral phenotypes (i.e., sternotribic and nototribic). Given that it is known that sternotribic flowers deposit pollen in less optimal areas on the pollinator’s body than nototribic flowers, our expectation is that plant species with sternotribic flowers will produce more seeds through geitonogamy than through xenogamy. Consequently, we expect a higher level of inbreeding depression in sternotribic flowers compared to nototribic flowers.
Materials and Methods
Reproductive Structure Measurements and Phenotype Classification
Thirteen Calceolaria taxa, including a subspecies, were selected during the spring-summer season of 2016–2018 from different locations throughout Chile (Table 1). To characterize the floral phenotype, each species was classified as sternotribic or nototribic depending on the arrangement of their reproductive structures (i.e., together covered by the upper lobe or resting in the lower lobe) as described by Sersic 2004 (Fig. 1B). To measure the size of the reproductive structures, three open flowers per individual (n = 30–50 plants per taxa) were cut, opened at the side so that the reproductive structures were visible while maintaining the shape of the flower, and immediately photographed using a Panasonic camera model DMC-LZ20 (Fig. 1A). Afterwards, the following traits were measured: (a) style length (mm): measured from the superior part of the ovary to the stigma surface, (b) stamen length (mm): measured from the base of the filament to the anther, and (c) herkogamy (mm): distance between the two reproductive structures (Fig. 1A). All measurements were made using ImageJ 1.46r software (available online at http://rsb.info.nih.gov/ij/).
Plant Breeding Assessment
Breeding experiments were carried out in the field (eleven taxa) and in the greenhouse (two taxa). In the field, twenty to fifty individuals per taxa were selected, four flower buds per plant were emasculated (n = 80–200 flower buds per taxa), covered with tulle mesh and randomly assigned to the following hand-pollination treatment: (a) self-pollination: emasculated bud pollinated with pollen from the same individual and different flowers, and (b) cross-pollination: emasculated bud pollinated with pollen from a plant donor located at least 1 m away. After hand-pollination, each bud was covered with a tulle mesh to prevent pollinators from visiting until fruiting. For the taxa growing in the greenhouse, we follow the protocol presented by Murúa et al. (2017), where twenty-five seeds from forty capsules were collected at the study site, with 1000 seeds per taxa. The growing plants were transplanted after three weeks, and then sixty of them were selected and subjected to the same treatments used in the field. When the fruit was ripened, the seeds were extracted and quantified under a magnifying glass in the laboratory.To estimate inbreeding depression (ẟ), we use the offspring data as follows:
A positive value of inbreeding depression (ẟ) indicates that self-pollination has a detrimental effect on the offspring, whereas a negative value indicates the opposite.
Statistical Analysis
To determine the difference in the size of the reproductive structures between the two floral phenotypes, a Kruskal-Wallis test was performed given the non-normal distribution of the data set. Subsequently a Dunn test with Bonferroni correction was conducted to identify the difference between taxa. To visualize the difference in the reproductive structures’ length and herkogamy between the two floral phenotypes, a principal component analysis (PCA) was conducted, including all Calceolaria taxa. GLM was performed with a Gaussian distribution, the measurements of reproductive structures as response variable, and floral phenotype (nototribic/sternotribic) and taxa as factors. The Akaike information criterion (AIC) with small sample correction was used to select the model that better explains our data and to test for multicollinearity in our model, the variance inflation factor (VIF) was used, with values under 3.5 for all the factors in our model (Zuur et al., 2010). In addition, a null model was generated to determine if there was a relationship with the evaluated characteristics in our model, or if it was random. We tested different models including all the variables (i.e., taxa, style length, stamen length and herkogamy) as factors. In our case, the better model excludes style length from the analysis (supplementary information). Therefore, we only report the values of the model that best explains our data.
To determine differences in geitonogamous selfing and cross-pollination between the two floral phenotypes and between taxa, a Kruskal-Wallis test was performed due to the non-normal distribution of our data. Subsequently a Dunn test with Bonferroni correction was conducted to identify the difference between taxa. To determine the relationship between floral phenotype and breeding system, we fitted a Linear mixed model (LMM) with the inbreeding depression (ẟ) per taxa as the response variable, floral phenotype, and the morphological characters (i.e., with stamen length, style length and herkogamy) as factors. As the sample size of the reproductive measurements exceeded that of the breeding experiment, a random sample of the plant individuals was selected in R for this analysis. Also, as we do not currently have a complete phylogeny of the genus Calceolaria (which includes all the species studied) we control for the species effect by including it as a random effect in the model. In order to ensure that the inbreeding depression followed a normal distribution, it was logarithmically transformed. Models were fitted to aGaussian distribution and used the floral phenotype sternotribic as 0 and nototribic as 1. LMMs were first run with all interactions, subsequently, to avoid over-parameterization of the analysis, a further model was run excluding interactions that were not significant (supplementary information). All the analyses were performed in R studio software version 1.1.453 (Core Team 2024).
Results
Of the thirteen taxa, six have a sternotribic floral phenotype, and seven have a nototribic one. According to the size of their reproductive structures, the sternotribic taxa showed, on average, a significantly longer stamen and style than those belonging to nototribic flowers (style length: d.f. = 1, chi-square = 174.63, P < 0.001; stamen length: d.f. = 1, chi-square = 163.59, P < 0.001; Fig. 2A and B). While the herkogamy, no significant differences were observed (herkogamy: d.f. = 1, chi-square = 0.58, P = 0.44; Fig. 2C). We also found significant differences between taxa in their reproductive structures (style length: d.f. = 12, chi-square = 440.53, P < 0.001; stamen length: d.f. = 12, chi-square = 425.66, P < 0.001; herkogamy: d.f. = 12, chi-square = 182.76, P < 0.001). On average, C. latifolia Benth has the longest style length and stamen length, and C. filicaulis Clos was the taxon with the shortest reproductive structures (Fig. 2A and B). In terms of herkogamy, C. tripartite Ruiz and Pav. had the lowest distance between reproductive structures, while C. purpurea Graham had the highest one (Fig. 2C).

Violin plots with mean morphological traits’ measurement (i.e., style length, stamen length and herkogamy) of two morphological floral phenotypes (i.e., nototribic and sternotribic) of the thirteen Calceolaria taxa (ARA = C. arachnoidea, COR = C. corymbosa, FIL = C. filicaulis, FILUX = C. filicaulis subsp. luxurians, INT = C. integrifolia, LANI = C. lanigera, LATI = C. latifolia, PET = C. petiolaris, POL = C. polifolia, PUR = C. purpurea, SEG = C. seguetti, THY = C. thyrsiflora, TRI = C. tripartita). Letters over bars indicate statistically significant differences among species. Dunn multiple comparisons with P-value adjusted with Bonferroni method
Principal components analysis (PCA) of the reproductive structure measurements yielded two eigenvectors, which collectively accounted for 95.26% of the variation observed in the data set (Table 2). The first eigenvector (PC1) was found to be strongly associated with the style length and stamen length measurements, accounting for 63.8% of the observed variation. The second eigenvector (PC2) was found to be related to herkogamy, accounting for 31.4% of the observed variation (Fig. 3).

Biplot of the first principal component (PC1) and the second principal component (PC2) showing the variation of reproductive structures (ST: style length and STA: stamen length) and herkogamy (H) between two floral phenotypes (sternotribic and nototribic) of thirteen Calceolaria’s taxa
GLM analysis showed a correlation between the herkogamy and the stamen length, the floral phenotype and the taxa (stamen length: d.f. = 1, t-value = 4.74, P < 0.001, floral phenotype: d.f. = 1, t-value = 3.36, P < 0.001; taxa: d.f. = 12, t-value = -2.19, P < 0.05) showing thatthose taxa with higher stamen length is more likely to exhibit a lower herkogamy and vice versa.
Breeding Assessment
All taxa were able to develop seeds by cross-pollination, but three of them were not able to develop seeds by self-pollination (C. filicaulis, C. filicaulis subsp. luxurians (Witasek) C. Ehrh. and C. purpurea; Fig. 4A and B). We did not find a significant difference between self-pollination and floral phenotypes (d.f. = 1, chi-square = -1.39, P = 0.23), however significant differences were observed between taxa (d.f. = 12, chi-square = 121.25, P < 0.001). In relation to cross-pollination no significant differences were found between floral phenotypes (d.f.=1, chi-square = 1.02, P = 0.31), but significant differences were found between taxa (d.f. = 12, chi-square = 53.33, P < 0.001). C. purpurea was the taxa with the lowest number of seeds per fruit in cross-fertilization, while C. tripartita and C. filicaulis subsp. luxurians were the taxa with the highest number of seeds self- and cross-pollination, respectively (Fig. 4A and B). Regarding the inbreeding depression (ẟ) no significant differences were observed between floral phenotypes (d.f. = 1, chi-square = 1.82, P = 0.17). However, we found significant differences between taxa (d.f. = 12, chi-square = 111.73, P < 0.001; Fig. 4C).

Mean seeds per pollination treatments andinbreeding depression (ẟ) estimation of thirteen Calceolaria taxa. Values are given as mean ± standard error. Letters over bars indicate statistically significant differences among species. Dunn multiple comparisons with P-value adjusted with Bonferroni method
The LMM analysis showed the stamen length significantly associated with the inbreeding depression (ẟ) (Table 3). This model suggests that flowers with a longer stamen length may have a higher inbreeding depression effect on their progeny.
Discussion
From an evolutionary perspective, it has been suggested that plants evolve to reduce pollen limitation (Lloyd, 1974; Lloyd & Schoen, 1992), where floral phenotype may have some implications for pollen reception (Knight et al., 2005). For instance, in the floral arrangement of the gynoecium/androecium, as in the genus Calceolaria, the floral phenotype may present adaptations for pollen deposition in optimal places on the pollinator’s body, increasing removal and the likelihood of pollen deposition in conspecifics (Westerkamp & Claßen-Bockhoff, 2007; Wiesenborn, 2019). The latter was recently documented in a study of pollination efficiency in Adenophora maximowiziana Makino (Funamoto, 2019), where it was found that nocturnal moths were the most efficient pollinators, which would be explained by the area where pollen is carried (i.e., ventral side). Accordingly, it has been suggested that these adaptations are key innovations that facilitated the diversification in some angiosperms (Westerkamp & Claßen-Bockhoff, 2007), including Calceolaria (Cosacov et al., 2009).
In our case, of the thirteen Calceolaria taxa studied, eight have nototribic flowers, and five have sternotribic flowers. We found a relationship between stamen length and herkogamy with the floral phenotypes (nototribic/ sternotribic). Accordingly, taxa with a sternotribic morphology tend to have higher stamen length and a lower herkogamy, whereas nototribic taxa show the opposite pattern- lower stamen length and a higher herkogamy. The length of the stamens has a significant impact on inbreeding depression. Sternotribic flowers, which produce fewer seeds through geitonogamy, exhibit higher inbreeding depression compared to nototribic flowers. Although these results partially contradicted our expectations, they could be explained by differences in the reproductive assurance mechanism followed by these floral phenotypes (i.e., nototribic and sternotribic).
Pollinators develop morphological adaptations that fit the morphology of the flowers they pollinate. These adaptations enable them to optimize pollination (Armbruster & Muchhala, 2009; Poblete Palacios et al., 2019). For instance, Anemopaegma album has a spatial arrangement where the anthers are hidden beneath the upper lip, resulting in pollen deposition on the dorsal surface of the bees (i.e., nototribic pollination). The floral architecture of this plant species minimizes pollen-stigma interference through herkogamy, where pollinators initially contact the pollen. Even if a pollinator touches the stigma of the same plant during departure, it occurs away from the receptive area (positioned adaxially) (Guimarães et al., 2015). Moreover, Müller (1996) conducted a study on thirteen bee species in Central Europe that have specialized pollen collection apparatus on their faces. These bees extract pollen from nototribic flowers of the families Lamiaceae and Scrophulariaceae through one of three methods: by rubbing their heads against the anthers, by buzzing the pollen among the transformed hairs, or by a combination of these two modes. Nototribic pollination has been described as a strategy to reduce the pollen loss due to grooming (Westerkamp & Claßen-Bockhoff, 2007), and is commonly observed in bees (Neal et al., 1998; Westerkamp & Claßen-Bockhoff, 2007; El Otra et al.,2016). Our results demonstrated no significant correlation between self-pollination, cross-pollination, and inbreeding depression with floral phenotypes (i.e., nototribic and sternotribic). This indicates that the spatial arrangement of morphological structures in Calceolaria taxa is not a primary factor influencing the fecundity of these taxa. Instead, the size of these structures appears to be a crucial determinant of reproductive success. Except for Calceolaria tripartita, nototribic flowers generally produce a lower number of seeds through geitonogamy than through xenogamy. Additionally, they present a lower stamen length and higher herkogamy, possibly to facilitate xenogamy and thereby avoid or reduce self-pollination (Cosacov et al., 2009).
On the other hand, in a comparative study of two sympatric species Angostura bracteate and Conchocarpus macrophyllus, specific floral arrangements were observed. These include secondary pollen presentation and very narrow corolla tubes with recessed anthers, respectively. As a result, sternotribic pollen transfer occurs, allowing for accurate pollination by lepidopterans. The authors also pointed out that herkogamy and the behavior of the pollinators both play crucial roles in the pollination of these species (El Ottra et al., 2016). Furthermore, in Adenophora maximowicziana a study demonstrated that its specific floral morphology, characterized by a pendant, bell-shaped corolla and a narrow corolla tube opening, likely restricts moth movements during visits. This restriction influences pollination efficacy, as pollen is deposited onto the ventral part of the moth’s bodies (i.e., sternotribic pollination) (Funamoto, 2019). Notably, A. maximowicziana is visited by both diurnal and nocturnal pollinators. Nevertheless, diurnal pollinators, including bumblebees, bees, and hoverflies, have been observed to reduce successful pollen transfer in this species (Castellanos et al., 2003; Thomson and Wilson, 2008; as cited in Funamoto, 2019). In contrast, our Calceolaria taxa are pollinated exclusively by bees. The deposition of pollen in a less optimal location may result in a reduction in the likelihood of accurate pollination (Thorp, 2000), and may also lead to an increased risk of inbreeding depression. Our results demonstrate an inbreeding effect in sternotribic flowers, as they produce fewer seeds through self-pollination. Surprisingly, this suggests that these taxa may be more pollinator-dependent than we expected, relying on xenogamy (cross-pollination) rather than geitonogamy (self-pollination) to achieve successful seed production. However, further research is required to elucidate this relationship.
While these modes of pollination may have an impact on the breeding systems of plants, information about their impact on inbreeding systems remains scarce. Our study revealed that all Calceolaria taxa are susceptible to the effects of self-pollination, showing higher values of inbreeding depression. LMM analysis showed a negative correlation with the floral phenotypes and the style length (supplementary information), and a significant association with the stamen length. For instance, C. latifolia (sternotribic flower) and C.tripartita (nototribic flower) showed lower values of inbreeding depression and had the highest self-pollination mean of all the taxa analyzed (Fig. 4A and C). Previous research using microsatellites in other species of the genus revealed that self-compatible species did not present inbreeding parameters (Fis index) significantly different from those of self-incompatible species (Murúa et al., 2020). These findings would reinforce the idea that a set of mechanisms may be interacting to circumvent the impacts of inbreeding in these species. In a study of Trichostema species, pollen deposition on the pollinator’s body (nototribic) and herkogamy function as mechanisms to reduce inbreeding depression (promoting cross-pollination). Nevertheless, the authors suggest that the capacity for self-pollination may be maintained due to unreliable pollinators (Spira, 1980). Additionally, Calceolaria latifolia is a self-compatible species that has been observed capable of self-fertilization (Murúa et al., 2020). Calceolaria tripartita is an invasive herb naturalized in Chile. This taxon has the highest self-pollination mean of all the nototribic taxa analyzed (Fig. 4A). Like many alien species, its selfing ability favors invasion success and establishment of new environments, allowing the taxon to reproduce (van Kleunen et al., 2015).
Conclusions
In our study, we investigated whether reproductive structure arrangement and herkogamy were associated with the breeding systems in Calcolaria’s taxa, and if those factors were correlated with floral phenotype (i.e., sternotribic and nototribic). We expected that those Calceolaria’s taxa with sternotribic flowers would produce more seeds by geitonogamy than nototribic ones. A correlation was identified between stamen length and herkogamy with the floral phenotypes. Plants with sternotribic flowers presented a higher stamen length and lower herkogamy. Additionally, a relationship was observed between stamen length and inbreeding depression, indicating thatthose flowers with longer stamen length are more affected by inbreeding depression. Among the species with a lower inbreeding depression is C. tripartita (nototribic), an invasive species that its capacity for self-fertilization contributes to its successful invasion and establishment in new environments. Thus, plants implement different reproductive assurance mechanisms under unreliable scenarios. Therefore, it is important to conduct further studies that consider other factors (e.g., flower phenology, pollen quality, insect behavior, among others) that may influence the relationship between flower phenotype, pollen extraction, and deposition, and reproductive success and compatibility system, to fully understand the evolution and maintenance of this important specialized genus.
Data Availability
The data that support the findings of this study are available from the Dryad Digital Repository: https://doi.org/10.5061/dryad.k3j9kd5bf.
References
Armbruster, W. S., & Muchhala, N. (2009). Associations between floral specialization and species diversity: Cause, effect, or correlation? Evolutionary Ecology, 23, 159–179.
Barrett, S. (2002). The evolution of plant sexual diversity. Nature Reviews Genetics, 3(4), 274–284.
Belaoussoff, S., & Shore, J. S. (1995). Floral correlates and fitness consequences of mating-system variation in Turnera ulmifolia. Evolution, 49(3), 545–556.
Bond, W. J. (1994). Do mutualisms matter? Assessing the impact of pollinator and disperser disruption on plant extinction. Philosophical Transactions of the Royal Society of London Series B: Biological Sciences, 344(1307), 83–90.
Carneiro, L. T., & Machado, I. C. (2023). Oil flowers and related oil-collecting bees: A 50-year timeline of knowledge and future directions. Arthropod-Plant Interactions, 17, 543–562.
Charlesworth, D., & Willis, J. H. (2009). The genetics of inbreeding depression. Nature Reviews Genetics, 10, 783–796.
Cisterna, J., & Murúa, M. (2018). Morfología floral y sistemas reproductivos contrastantes en dos subespecies de Calceolaria corymbosa en Chile central. Gayana Botanica, 75(1), 544–548.
Cosacov, A., Sérsic, A. N., Sosa, V., De-Nova, J. A., Nylinder, S., & Cocucci, A. A. (2009). New insights into the phylogenetic relationships, character evolution, and phytogeographic patterns of Calceolaria (Calceolariaceae). American Journal of Botany, 96, 2240–2255.
Dai, C., & Galloway, L. F. (2011). Do dichogamy and herkogamy reduce sexual interference in a self-incompatible species? Functional Ecology, 25, 271–278.
Eckert, C. G., Samis, K. E., & Dart, S. (2006). Reproductive assurance and the evolution of uniparental reproduction in flowering plants. In L. D. Harder, & S. C. H. Barrett (Eds.), Ecology and Evolution of Flowers (pp. 183–203). Oxford University Press.
Ehrhart, C. (2000). Die Gattung Calceolaria (Scrophulariaceae) in Chile. Bibliotheca Botanica, 153, 1–283.
El Otra, J. H. L., Pirani, J. R., & Pansarin, E. R. (2016). Floral biology and pollination of two sympatric species of Galipeinae (Galipeeae, Rutaceae) endemic to the Brazilian Atlantic Forest. Flora, 221, 107–116.
Fenster, C. B., & Marten-Rodriguez, S. (2007). Reproductive assurance and the evolution of pollination specialization. International Journal of Plant Sciences, 168, 215–228.
Fetscher, A. E. (2001). Resolution of male-female conflict in a hermaphroditic flower. Proceedings of the Royal Society of London Series B: Biological Sciences, 268(1466), 525–529.
Funamoto, D. (2019). Precise sternotribic pollination by settling moths in Adenophora maximowicziana (Campanulaceae). International Journal of Plant Sciences, 180(3), 000–000.
Goldberg, E. E., Kohn, J. R., Lande, R., Robertson, K. A., & Smith S. A. (2010). Species selection maintains self-incompatibility. Science, 330, 493–495.
Goodwillie, C., Kalisz, S., & Eckert, C. G. (2005). The evolutionary enigma of mixed mating systems in plants: Occurrence, theoretical explanations, and empirical evidence. Annual Review Ecology Evolution and Systematics, 36, 47–79.
Guimarães, E., Nogueira, A., Dias Netto, C. G., & Rodrigues Machado, S. (2015). Pollination of Anemopaegma album (Bignoniaeae) with focus on floral nectar as the mediator of interactions with mutualistic and antagonistic bees. The International Journal of Plant Reproductive Biology, 7(2), 177–188.
Igic, B., & Busch, J. W. (2013). Is self-fertilization an evolutionary dead end? New Phytologist, 198, 386–397.
Knight, T. M., Steets, J. A., Vamosi, S. J., Mazer, S. J., Burd, M., Campbell, D. R., Dudash, M. R., et al. (2005). Pollen limitation of plant reproduction: Pattern and process. Annual Review Ecology Evolution and Systematics, 36, 467–497.
Lloyd, D. G. (1974). ‘Theoretical sex ratios of dioecious and gynodioecious angiosperms’, Heredity, 32(1), pp 11–34.
Lloyd, D. G., & Schoen, D. J. (1992). Self-fertilization and cross-fertilization in plants. I. Functional dimensions. International Journal of Plant Sciences, 153, 358–369.
Martén-Rodrìguez, S., & Fenster, C. B. (2010). Pollen limitation and reproductive assurance in Antillean Gesnerieae: A specialist vs. generalist comparison. Ecology, 91(1), 155–165.
Molau, U. (1988). Scrophulariaceae-part I. Calceolarieae. Flora Neotropica, 47, 1–326.
Morgan, M. T., & Wilson, W. G. (2005). Self-fertilization and the escape from pollen-limitation in variable pollination environments. Evolution, 59, 1143–1148.
Muchhala, N. (2007). Adaptative trade-off in floral morphology mediates specialization for flowers pollinated by bats and hummingbirds. American Naturalist, 169, 494–504.
Müller, A. (1996). Convergent evolution of morphological specializations in central European bee and honey wasp species as an adaptation to the uptake of pollen from nototribic flowers (Hymenoptera, Apoidea and Masaridae). Biological Journal of the Linnean Society, 57, 235–252.
Murúa, M., Cisterna, J., & Rosende, B. (2014). Pollination ecology and breeding system of two Calceolaria species in Chile. Revista Chilena De Historia Natural, 87, 7.
Murúa, M., Espíndola, A., Gonzalez, A., & Medel, R. (2017). Assessment of reproductive barriers in two sympatric oil-rewarding Calceolaria. Evolutionary Ecology, 31, 421–434.
Murúa, M., Espíndola, A., & Pérez, F. (2020). Self-Compatibility Not Associated with morphological or genetic diversity reduction in oil-rewarding Calceolaria Species. Plants, 9(10), 1377.
Neal, P. R., Dafni, A., & Giurfa, M. (1998). Floral symmetry and its role in plant-pollinator systems: Terminology, distribution, and hypotheses. Annual Review of Ecology Evolution and Systematics, 29, 345–373.
Opedal, Ø. H. (2018). Herkogamy, a principal functional trait of plant reproductive biology. International Journal of Plant Sciences, 179(9), 677–687.
Opedal, Ø. H., Albertsen, E., Armbruster, W. S., Pérez-Barrales, R., Falahati-Anbaran, M., & Pélabon, C. (2016). Evolutionary consequences of ecological factors: Pollinator reliability predicts mating-system traits of a perennial plant. Ecology Letters, 19, 1486–1495.
Pauw, A. (2006). Floral syndromes accurately predict pollination by a specialized oil-collecting bee (Rediviva peringueyi, Melittidae) in a guild of South African orchids (Coryciinae). American Journal of Botany, 93(6), 917–926.
Poblete Palacios, J. A., Soteras, F., & Cocucci, A. A. (2019). Mechanical fit between flower and pollinators in relation to realized precision and accuracy in the hummingbird-pollinated Dolichandra cynanchoides. Biological Journal of the Linnean Society, 126(4), 655–665.
Possobom, C. C. F., & Machado, S. R. (2017). Elaiophores in three neotropical Malpighiaceae species: A comparative study. Plant Systematics and Evolution, 304(1), 15–32.
Rani, R. S., & Raju, A. J. S. (2020). Reproductive ecology of nutritionally important perennial climber Canavalia gladiata (Fabaceae: Faboideae). Phytologia Balcanica, 26(1), 43–55.
SchoenDJ, & BrownAH (1991). Intraspecific variation in population gene diversity and effective population size correlates with the mating system in plants. Proceedings of the National Academy of Sciences of America, 88, 4494–4497.
Schoen, D. J., & Lloyd, D. G. (1992). Self and cross-fertilization in plants III methods for studying modes and functional aspects of self-fertilization. International Journal of Plant Science, 153, 381–393.
Sérsic, A. N. (1991). Observaciones Sobre El mecanismo floral de Calceolaria (Vol. ,21Kurtziana, 21, pp. 153–164). Scrophulariaceae)’.
Sérsic, A. N. (2004). ‘Pollination biology in the genus Calceolaria L. (Calceolariaceae)’, Stapfia, 82, pp 122.
Siopa, C., Dias, M. C., Castro, M., Loureiro, J., & Castro, S. (2020). Is selfing a reproductive assurance promoting polyploid establishment? Reduced fitness, leaky self-incompatibility and lower inbreeding depression in neotetraploids. American Journal of Botany, 107(3), 526–538.
Spira, T. P. (1980). Floral parameters, breeding system and pollinator type in Trichostema (Labiatae). American Journal of Botany, 67(3), 278–284.
Takebayashi, N., & Morrell, P. L. (2001). Is self-fertilization an evolutionary dead end? Revisiting an old hypothesis with genetic theories and a macroevolutionary approach. American Journal of Botany, 88, 1143–1150.
Thorp, R. W. (2000). The collection of pollen by bees. In A. Dafni, M. Hesse, & E. Pacini (Eds.), Pollen and pollination (pp. 211–223). Springer.
Van Kleunen, M., Dawson, W., & Maurel, N. (2015). Characteristics of successful alien plants. Molecular Ecology, 24, 1954–1968.
Vaughton, G., & Ramsey, M. (2010). Pollinator-mediated selfing erodes the flexibility of the best-of-both-worlds mating strategy in Bulbine vagans. Functional Ecology, 24, 374–381.
Webb, C. J., & Lloyd, D. G. (1986). The avoidance of interference between the presentation of pollen and stigmas in Angiosperms II. Herkogamy. New Zealand Journal of Botany, 24(1), 163–178.
Westerkamp, C., & Claßen-Bockhoff, R. (2007). Bilabiate flowers: The ultimate response to bees? Annals of Botany, 100, 361–374.
Wiesenborn, W. D. (2019). Pollination of Scutellaria mexicana (Lamiaceae) in southern Nevada by Anthophora bees. Western North American Naturalist, 79(2), 195–203.
Zhang, W., Hu, Y. F., He, X., Zhou, W., & Shao, J. W. (2021). Evolution of autonomous selfing in marginal habitats: Spatiotemporal variation in the floral traits of the distylous Primula wannanensis. Frontiers in Plant Science, 12, 781281.
Zuur, A. F., Leno, E. N., & Elphick, C. S. (2010). A protocol for data exploration to avoid common statistical problems. Methods in Ecology and Evolution, 1, 3–14.
Acknowledgements
The authors thank Pontificia Universidad Católica de Chile for providing access to laboratory facilities. The authors also thank Isidora Sepúlveda for field assistance. All laboratory and field work were founded by Fondecyt 1170377, Agencia Nacional de Investigación y Desarrollo (ANID).
Funding
All laboratory and field work were founded by Fondecyt 1170377, Agencia Nacional de Investigación y Desarrollo (ANID).
Author information
Authors and Affiliations
Contributions
MM conceived and designed the experiments and carried out the reproductive experiments, MJR analyzed the data. MM, CE, and MJR wrote the first draft of the manuscript, MM and MJR substantially contributed to the revisions and prepared the final version of the article. All authors have read and agreed to the published version of the manuscript.
Corresponding author
Ethics declarations
Ethical Approval
Not applicable.
Consent for Publication
Not applicable.
Competing Interests
The authors declare no competing interests.
Conflict of Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Consent to Participate
Not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Ramírez, M.J., Escanilla-Jaramillo, C. & Murúa, M.M. A Specialized Combination: The Relationship between Reproductive Structure Arrangement and Breeding Systems in oil-rewarding Calceolaria Species (Calceolariaceae). Evol Biol (2024). https://doi.org/10.1007/s11692-024-09639-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11692-024-09639-2