
Abstract
Purpose
Bats belong to different guilds and thus differ in the environmental resources that they exploit. They also evolved to have a very rich diversity of roosting and feeding habits. Therefore, it enables them to harbor different species of parasites that could be zoonotic and may pose public health importance. The current study aimed to identify the ectoparasites of bats, under Family Streblidae, in different caves of Unisan, Quezon, Philippines.
Methods
Four caves were sampled in the study namely, Malusak, Mahangin, De Lamesa, and Bonifacio. Two mist nets for each cave were set up to capture bats. The bats were carefully observed for the presence of ectoparasites using forceps and cotton balls.
Results
Eight bat species belonging to four families, namely Hipposideridae, Vespertilionidae, Rhinolophidae, and Pteropodidae, were recovered from the study. Results showed that 130 out of 202 (64.36%) bats were infested with streblids with an overall mean intensity of 1 ectoparasite/bat. Out of four streblid species, Brachytarsina werneri, Brachytarsina amboinensis, and Raymondia pseudopagodarum were classified as stenoxenous. High specificity index was recorded for R. pseudopagodarum (STD = 3) and low specificity index for Brachytarsina species.
Conclusion
These results can be used by the local government in providing necessary actions that may lead to public awareness as these bats may also play a role in the transmission of zoonotic parasites in the area.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Bats (Mammalia: Chiroptera) are known to have a very rich diversity in terms of behavioral, roosting, and feeding habits. One of the locations they occupy is caves, where thousands of individuals can inhabit [1]. This behavior makes them a natural reservoir of many ectoparasitic arthropods [2, 3]. Ectoparasites of bats have complex assemblages and can be a potential vector-borne that transmit numerous diseases to both wildlife and domestic animals [4]. There have been studies done on other parts of the region such as in Mount Makiling, Luzon Island, and in Mindoro Island Philippines. In Mount Makiling, four bat species were recorded specifically, Cynopterus brachyotis (Muller), Ptenochirus jagori (Peters), Rousettus amplexicaudatus (Geoffroy), and Rhinolophus arcuatus (Peters) [5]. In Mindoro Island, there are seven species of Nycteribiid flies, five species of Streblid flies, and one species of Ischnopsyllid flea. Furthermore, Raymondia pagodarum is a new record of Streblid species found in Mindoro [6].
Bats flies (Streblidae) are one of the most common blood-sucking and obligate ectoparasites of bats. As hematophagous arthropods, they may harbor different bacteria, viruses, and other parasites that are in the blood of their host. Streblids are also known to be highly host-specific and live most of their lives on their host [3, 7, 8]. Being highly host-specific and hematophagous arthropods, streblids may pose as a public health important arthropod and they could also play an important role in zoonotic transmission.
Aroon et al. [9] stated that despite the documented species of bats, there is still a lack of data about parasites associated with bats in tropical regions, particularly in Southeast Asian countries. There are 26 recently recorded bat species in the Philippines, wherein 17 of which are endemic to the country. These bat species were commonly found roosting in caves [10]. Philippines are known to have at least 1500 known caves, where most of the bats are found roosting; however, only four are under the protection of the National Integrated Protected Areas System (NIPAS) act (Tanalgo and Hughes, 2018). Most of the caves in Unisan, Quezon, Philippines are being used by the community in performing their religious rituals like getting water, praying in the cave, and mass gathering inside the cave. These practices could expose the community to the bats and to its numerous parasites that were naturally harbored by the bats. Thus, the current study provided new data on the prevalence and host specificity of streblid ectoparasites among bats that are of public health importance.
Materials and Methods
Sampling
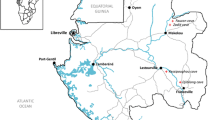
Four caves are included in this study: Malusak (122.058943, 13.844378), Mahangin (122.058766, 13.84415), De Lamesa (122.058756, 13.844639), and Bonifacio (122.027466, 13.898252) (Fig. 1). The sampling of bats was conducted from September to October 2018.

source: QGIA, 2018)
Location of sampled caves in Unisan, Quezon. (Author: Paulo Obico;
Two mist nets measuring 3 m × 2.5 m were positioned in the possible entry and exit points of every cave to capture bats. The preparation and collection were from 1800 to 0500 h, and the setup was observed every 3 h to ensure if there were already bats captured [11]. To avoid stress to the bats, captured bats were placed in a separate cloth bag for the collection and observation of samples. Individual bats were carefully handled and examined after collection. Flashlights were used to search for the ectoparasites in different body parts including the central and dorsal parts of the ears and pelage, toes, and wing ridges. Visible ectoparasites were collected using forceps and placed in a microcentrifuge tube containing 70% ethanol to ensure that the specimen will remain dehydrated and the body parts will still be intact [12]. Each sample was labeled and recorded accordingly. Aside from visible ectoparasites, there are also very small ones that are not visible to the naked eye. These were collected by brushing the whole body of the bats with cotton wools soaked in 70% ethanol and secured the pieces of cotton in a ziplock bag with a proper label [13].
As soon as the collection of samples was done, the bats were immediately released with no harm done. Non-toxic colored paint was used to mark their uropatagium on hindfoot before releasing to know that the bats have been captured. The identification for bat species was based on the key presented by Ingle and Heaney [14]. Meanwhile, ectoparasites were initially identified through different journals and were verified by an entomologist, Mr. Ace Kevin Amarga of the University of the Philippines, Los Banos.
Data Analysis
The prevalence and mean intensity of ectoparasites present in bats were calculated using the formula by Bush et al. [15]. Meanwhile, the confidence interval for prevalence and mean intensity was computed using Quantitative Parasitology 3.0 (http://www.zoologia.hu/qp/qp.html). The collected data were analyzed if it is within the acceptable parameters to ensure that they could be numerically analyzed. Only those bat fly samples with greater than five bat individuals of any species and 20 bat flies of the same species were included in the analysis. To assess the host specificity, the final ratio of bat fly species abundance across host species must also be identified. To compute the ratio, the individual bat flies per host species were divided by the total number of bat fly individuals collected. Furthermore, to account for the sampling errors that can be due to sampling contamination, a set of parameters was employed for the ratio. A criterion of 0.05 of the ratios was considered reasonable. All species that have a ratio less than 0.05 were dismissed to decrease the likelihood of spurious results [16].
Host specificity was calculated using the index of host specificity (1) by Poulin and Mouillot [17]:
where s is the number of host species used by a parasite, the double summation is over the set {i = 1,…..s; j = 1,….s}, and ωij is the taxonomic distinctness between host species i and j. The number of taxonomic steps required to reach a node common to both can also be computed using the following formula (2):
where ϖ is simply the average taxonomic distinctness or STD's is the number of host species used by a parasite. Note that the variance in STD can only be computed when a parasite exploits three or more host species (it always equals zero with two species).
Results
Prevalence and Mean Intensity of Streblidae Ectoparasites in Bats
Eight species of bats from four families were captured in the study. A total of 130 out of 202 bats (64.36%) were found positive with ectoparasites under Family Streblidae. The bat flies recovered were Raymondia pseudopagodarum, Brachytarsina werneri, Megastrebla parvior, and Brachytarsina amboinensis.
Host Specificity of the Parasites
The abundance of Streblid bat flies on each sampled bat species is presented in Table 1. It showed that Brachytarsina werneri was found infesting all the sampled bat species. Raymondia pseudopagodarum was found infecting two species of bats, Hipposideros ater and Rhinolophus arcuatus, which belongs to Family Hipposideridae and Family Rhinolophidae, respectively. Meanwhile, Megastrebla parvior was found to infest only one individual of bat species, Rousettus amplexicaudatus, which makes it difficult to be classified to any of the host–parasite associations.
Bat species that are lesser than the standard sample size parameters were dismissed for the computation of host specificity, since it may lead to spurious results [16]. With this, there are only 133 individual bats infected with specific bat flies assessed for host specificity (out of an original total of 148) (Table 1).
The categories of the bat flies species can also be found in Table 2. Raymondia pseudopagodarum, Brachytarsina werneri, and Brachytarsina amboinensis are categorized as stenoxenous, since they are host specific to two host species. It is also noted that there are few host species for the bat flies, which gives a high specificity index for R. pseudopagodarum (STD = 3) and low specificity index for B. werneri and B. amboinensis (STD = 1.667).
Discussion
The current study presented the prevalence of streblid bat flies recovered from bats in caves of Unisan, Quezon, and its association with its host. The prevalence of ectoparasite in the current study is seemingly high (64.36%) in comparison with the past studies. In Ghana, 159 out of 480 bats (33.13%) captured were found infested with ectoparasites [18]. In Central Panama, a study revealed that there is a prevalence of between 6.8% and 69.3% among 3, 456 individual captured bats [7]. In all mentioned studies including the current study, it was noted that it is warm during the collection of the samples. Woods [2] described that there is a higher prevalence of parasites during the summer season. Furthermore, a low prevalence of parasites was observed during the rainy season with a low temperature in the study of Aroon et al. [9] and Nartey [18].
Another factor that may have influenced the prevalence of ectoparasites in the current study is to where the captured bats roost, which is in caves. Caves are considered to be more permanent structures compared to other living spaces of bats. Since the sampling sites of the current study are caves, then the captured bats are more likely to harbor heavy loads of different species of parasites [19]. Hiller et al. [7] added that caves are a good site to host a stable population of bats. With this, a higher prevalence and intensity of ectoparasite infestation can be recorded in this area.
Meanwhile, Table 1 shows that the mean intensity among bats ranges from 1.00 to 2.53 ectoparasites per bats. The intensity of ectoparasite infection among the captured bats does not significantly far from each other, since most of the sampled bats are already sub-adults to adults. This means that these bats are already active hunting for foods and they might also be in their reproductive stage. Nartey et al. [18] stated that the age and reproductive stage of bats have a role in harboring parasites, since adult bats are active than the juveniles; while those who are in their lactating stage (reproductive female adults) can be immunosuppressed and can easily be infested with parasites.
Among the four ectoparasites recovered, Brachytarsina werneri is the most prevalent, infecting all bat species captured. This bat fly is also found to be the most prevalent parasite in Northeastern Thailand with 47.83% and mean intensity of 2 parasites/bat [9]. The presence of ectoparasite fauna in bats was also recorded in Mindoro Island, Philippines. The recorded streblid ectoparasites, that were the same in the current study, were Megastrebla parvior, Brachytarsina amboinensis, Brachytarsina werneri, and Raymondia pseudopagodarum [6]. It is interesting to note that in the study of Alvarez et al. [6], M. parvior, B. amboinensis, and R. pseudopagodarum were found infesting Myotis minimus, Hipposideros diadema, and Hipposideros bicolor, respectively, which are different from the current study. Meanwhile, Brachytarsina werneri was found infesting the same species of bats (Rhinolophus arcuatus).
In terms of host specificity, the current study reveals that streblid bat flies are generally highly host-specific. This result is congruent to previous studies in French Guiana [3], Paraguay [20], and the Lamanai area of Belize [13]. Furthermore, results revealed that Raymondia pseudopagodarum, Brachytarsina werneri, and Brachytarsina amboinensis are categorized as stenoxenous as it is host-specific to two different bat species. In the study of Maa [21], R. pseudopagodarum and B. amboinensis are classified as oligoxenous. However, it was also found that it infests the same bat species as in the current study. R. pseudopagodarum was found infesting Rhinolophus and Hipposideros, while B. amboinensis was found in Myotis and Minioptera. These bats are known as gregarious bats, which are found roosting in deep caves just like in the current study. Hiller et al. [7] added that roosting behavior of bats like using permanent roost (e.g., caves) increases the chance of parasite transmission from one bat to the other. The difference in the categorization of the bat flies of the current study to the previous studies could be due to the smaller sample size of the study. However, since all the samples that did not qualify for the parameters set are dismissed, the chance of having spurious results is already lessened. The results also showed high and low STD values, which are also possible when there are few host species known, as supported by the study of Poulin and Mouillot [17].
Summary and Conclusion
The current study presents the first record of Streblid ectoparasites in the caves of Unisan, Quezon, Philippines, and therefore, all the recovered ectoparasites are new records in this province. Raymondia pseudopagodarum, Brachytarsinawerneri, and B. amboinensis are the recovered parasites that are categorized as stenoxenous. Specificity index was also considered in the study which revealed that P. pseudopagodarum has high specificity index while Brachytarsina species have a low specificity index. With this, it is recommended to further study bats harbor parasites (endo and ectoparasites) that are transmissible to humans. Bigger sample size is also recommended to get a better analysis of the host specificity of the different parasites among bats.
Availability of Data and Material
All data generated or analysed during this study are included in this published article.
Code Availability
Not applicable.
References
Kunz TH, De Torrez EB, Bauer D, Lobova T, Fleming TH (2011) Ecosystem services provided by bats. Ann NY Acad Sci 1223(1):1–38. https://doi.org/10.1111/j.1749-6632.2011.06004.x
Woods S (2012) Geographic distribution and composition of the parasite assemblage of the insectivorous bat, Miniopterus natalensis (Chiroptera: Miniopteridae), in South Africa. J Zool.
Graciolli G, Guerrero R, Catzeflis F (2019) Streblid bat flies (Diptera) and other ectoparasites on bats (Mammalia: Chiroptera) from French Guiana. Biota Neotrop 19(4):e20180724. https://doi.org/10.1590/1676-0611-BN-2018-0724
Dick CW, Dittmar K (2014) Chapter 6 Parasitic bat Flies (Diptera: Streblidae and Nycteribiidae) host specificity and potential as vectors. In Sven Klimpel and Heinz Mehlhorn, bats (Chiroptera) as vectors of diseases and parasites: facts and myths, p 131–151.
Alvarez JDV, Lit IL Jr, Alviola PA (2015) Bat flies (Diptera: Nycteribiidae) from Makiling, Luzon Island: New host and distribution records, with a checklist of species found in the Philippines. Museum of Natural History, University of the Philippines, Los Baños College. Laguna, Philippines. 4031.
Alvarez JDV, Lit IL Jr, Alviola PA, Cosico EA, Eres EG (2016) A contribution to the ectoparasite fauna of bats (Mammalia: Chiroptera) in Mindoro Island, Philippines: I. Blood sucking Diptera (Nycteribiidae, Streblidae) and Siphonaptera (Ischnopsyllidae). Int J Tropical Insect Sci. 36(4):188–194. https://doi.org/10.1017/S1742758416000187
Hiller T, Brändel SD, Honner B, Page RA, Tschapka M (2019) Parasitization of bats by bat flies (Streblidae) in fragmented habitats. Biotropica. https://doi.org/10.1111/btp.12757
Szentiványi T, Christe P, Glaizot O (2019) Bat flies and their microparasites: current knowledge and distribution. Front Vet Sci 6:115. https://doi.org/10.3389/fvets.2019.00115
Aroon SJ, Hill JG III, Artchawakom T, Pinmongkholgul S, Kupittayanant S, Thanee N (2015) Ectoparasites associated with Bats in Tropical Forest of Northeastern Thailand. J Agricult Technol 11(8):1781–1792
Luczon AU, Ampo SAMM, Rono JGA, Duya MRM, Ong PO, Fontanilla IKC (2019) DNA barcodes reveal high genetic diversity in philippine fruit bats. Philipp J Sci 148(S1):133–140
Amarga AS, Alviola PA, Lit IL Jr, Yap SA (2017) Checklist of ectoparasitic arthropods among cave-dwelling bats from Marinduque Island Philippines. Checklist J Biodivers Data 13(1):2029. https://doi.org/10.15560/13.1.2029
ter Hofstede HM, Fenton MB, Whitaker JO Jr (2004) Host and host specificity of Bat flies (Diptera: Streblidae and Nycteribiidae) on Neotropical Bats (Chiroptera). J Zool 82:616–626
Dick CK, Patterson BD (2006) Eleven bat flies—obligate ectoparasite of bats. Springer, New York
Ingle NR, Heaney LR (1992) Fieldiana: a key to bats of the Philippine Islands. Field Museum of Natural History, Chicago
Bush AO, Lafferty KD, Lotz JM, Shostak AW (1997) Parasitology meets ecology on its own terms: Margolis et al. revisited. J Parasitol 83(4):575–583. https://doi.org/10.2307/3284227
Stamper E (2012) Host specificity of ecuadorian bat flies (Diptera: Streblidae). Retrieved from Western Kentucky University Top Scholar: Honors College Capstone Experience/Thesis Projects: http://digitalcommons.wku.edu/stu_hon_theses/358
Poulin R, Mouillot D (2002) Parasite specialization from a phylogenetic perspective: a new index of host specificity. Parasitology. https://doi.org/10.1017/S0031182003002993
Nartey N (2015) Common parasites of fruit-eating bats in Southern Ghana. Mphil Zoology Degree. University of Ghana http://ugspace.ug.edu.gh
Patterson BD, Dick CW, Dittmar K (2007) Roosting habits of bats affect their parasitism by bat flies (Diptera: Streblidae). J Trop Ecol 23:177–189. https://doi.org/10.1017/S0266467406003816
Dick CW, Hettinger D (2005) A faunal survey of streblid flies (Diptera: Streblidae) associated with bats in Paraguay. J Parasitol. https://doi.org/10.1645/GE-536R.1
Maa T (1971) Revision of the Australian bat flies (Diptera: Streblidae and Nycteribiidae). In: Pacific insects monograph, p 1–118.
Acknowledgments
The researchers would like to acknowledge Department of Environment and Natural Resources Regional Office No. IV-A for the endorsement of wildlife gratuitous permit and assistance from the Mathematics and Natural Sciences Department of Southern Luzon State University.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
MCO: Methodology, Investigation, Resources, Data Curation MPVG Methodology, Investigation, Resources, Data Curation EVS: Conceptualization, Methodology, Writing-Review & Editing, Validation, Formal Analysis, Supervision, Project Administration KP: Validation, Formal Analysis, Resources, Data Curation, Writing-Original Draft, Writing-Review & Editing, Visualization, Supervision, Project Administration.
Corresponding author
Ethics declarations
Conflict of Interest
None declared.
Ethics Approval
Wildlife Gratuitous Permit No. R4A-WGP-2018-QUE-024 was endorsed by the Department of Environment and Natural Resources Regional Office No. IV-A.
Consent to Participate
Not applicable 2.
Consent for Publication
All the authors whose names appear on the submission, have contributed to the study and approved for its publication.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
*
Rights and permissions
About this article

Cite this article
Obdianela, M.C.N., Guanlao, M.P.V.V., Samaniego, E.V.E. et al. Prevalence and Host Specificity of Bat Flies (Streblidae) from Selected Caves in Unisan, Quezon, Philippines. Acta Parasit. 66, 983–988 (2021). https://doi.org/10.1007/s11686-021-00373-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11686-021-00373-9