
Abstract
Brain development during childhood and early adolescence is characterized by global changes in brain architecture. Neuroimaging studies have revealed overall decreases in cortical thickness (CT) and increases in fractional anisotropy (FA). Furthermore, previous studies have shown that certain cortical regions display coordinated growth during development. However, there is significant heterogeneity in the timing and speed of these developmental transformations, and it is still unclear whether white and grey matter changes are co-localized. In this multimodal neuroimaging study, we investigated the relationship between grey and white matter developmental changes and asynchronous maturation within brain regions in 249 normally developing children between the ages 7–14. We used structural magnetic resonance imaging (MRI) and diffusion tensor imaging (DTI) to analyze CT and FA, respectively, as well as their covariance across development. Consistent with previous studies, we observed overall cortical thinning with age, which was accompanied by increased FA. We then compared the coordinated development of grey and white matter as indexed by covariance measures. Covariance between grey matter regions and the microstructure of white matter tracts connecting those regions were highly similar, suggesting that coordinated changes in the cortex were mirrored by coordinated changes in their respective tracts. Examining within-brain divergent trajectories, we found significant structural decoupling (decreased covariance) between several brain regions and tracts in the 9- to 11-year-old group, particularly involving the forceps minor and the regions that it connects to. We argue that this decoupling could reflect a developmental pattern within the prefrontal region in 9- and 11-year-old children, possibly related to the significant changes in cognitive control observed at this age.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Neuroimaging studies have provided a great deal of information about normal brain development. However, the complex interplay between grey and white matter development is still poorly understood (Tamnes et al. 2010). From a neuroimaging perspective, two main issues remain unsolved: 1) how changes in white matter microstructure are related to changes in grey matter macrostructure; and 2) how homogeneous the interplay between these two maturational processes is within the brain. Structural neuroimaging studies have shown that brain cortical thickness (CT) increases in early childhood, followed by cortical thinning during puberty (Giedd 2004; Gogtay et al. 2004; Shaw et al. 2008; Sowell et al. 2003). In addition, diffusion tensor imaging (DTI) studies have revealed an increase in fractional anisotropy (FA) during childhood and adolescence (Lebel and Beaulieu 2011; Lebel et al. 2008; Peters et al. 2012; Schmithorst and Yuan 2010; Tamnes et al. 2010; Westlye et al. 2009). Furthermore, white matter changes are not synchronous across the whole brain, with fronto-temporal connections developing at a later stage (Lebel et al. 2008), even going into early adulthood (Bava et al. 2010).
The microstructural correlates of grey matter macrostructural changes are unknown. Cortical thinning could result from the pruning of synaptic processes (Tau and Peterson 2010), intracortical myelination (Tomáš Paus 2010; T Paus et al. 2001), or from a variety of changes in the microstructure bundle, including increased axon diameter or packing density (Jones et al. 2013; Tomáš Paus 2010; Takahashi et al. 2002). Furthermore, the timing and speed of these cortical changes in specific brain regions seem to be carefully orchestrated. It has recently been proposed that studying the structural covariance pattern of CT could reveal novel underlying developmental processes and that these patterns could be used as signatures of brain development (Alexander-Bloch et al. 2013a).
Several structural and functional brain changes occur during the critical periods between late childhood, puberty and adolescence (Blakemore 2012; Giedd and Rapoport 2010; Yurgelun-Todd 2007). An accurate understanding of healthy developmental brain trajectories during this period could provide the foundation for hypotheses regarding the neural bases of developmentally-based psychopathologies, such as schizophrenia and bipolar disorder (Marsh et al. 2008; Tau and Peterson 2010). Thus, studying the neural developmental signatures embedded in different patterns of structural covariance across childhood could help to elucidate the distributed neuroanatomical abnormalities observed in neuropsychiatric disorders in older individuals (Paus et al. 2008; Raznahan et al. 2011).
It has been shown that synchronized structural or/and functional changes occur in the human cortex over development (A. Alexander-Bloch et al. 2013a; Mechelli et al. 2005; Raznahan et al. 2011; Zielinski et al. 2010). In addition, a partial correlation has been found between cortical thickness and diffusion connection in adulthood (Gong et al. 2012). It is still unknown whether white matter tract development follows a similar coordinated development, which would manifest as covariance in fractional anisotropy between tracts. Furthermore, we do not know whether these possible synchronous maturational changes mirrors the harmonic changes in cortical thickness in childhood. Thereby, we present a cross-sectional MRI study exploring the brain structures of 249 children and adolescents between 7 and 14 years of age. Our first goal was to investigate the overall age-related CT and FA patterns by focusing on how FA changes are related to CT changes in the whole sample. We hypothesized that tracts of regions showing coordinated decreases in CT would also demonstrate coordinated increases in FA, indicating synchronous maturational changes in the soma and axons of the relevant neurons. Second, we subdivided our sample into 3 groups: 7–8 years olds, 9–11 years olds, and 12–14 years olds. Then, using a structural covariance analysis of both FA and CT, we investigated whether certain brain regions demonstrated time-specific developmental divergence in their coordinated grey and white matter maturation relative to the rest of the brain during these periods.
Methods
Participants
A total of 249 normally developing children and adolescents (7 to 14 years of age, 139 males) participated in this study. The sample was obtained from a large community school-based study (Salum et al. 2014) from Brazil, a middle-income country that is usually under-represented in scientific studies (Tomlinson et al. 2009). Children with any DSM-IV mental health disorders – as confirmed using the Diagnostic and Well-Being Assessment (DAWBA) (Goodman et al. 2000) – were excluded. The study methodology is outlined in the Fig S1 available in the supplementary material.
To study the coordinated development across childhood, the sample was divided into three age groups based on Shaw et al. (2008) (7–8, 9–11, and 12–14 years old). Each group was composed of 83 subjects and was balanced by gender. The subjects’ estimated IQs were determined using the Wechsler Intelligence Scale for Children (blocks and vocabulary), 3rd edition (WISC-III) (Wechsler 2002), following Brazilian norms (Figueiredo 2001). The demographics of the three groups are displayed in Table 1.
MRI acquisition
MRI scanning was performed at two centers using a 1.5-T MRI system (Signa HD at the Porto Alegre Center and Signa HDX at the Sao Paulo Center; General Electric, Milwaukee, WI, USA). A total of 114 subjects were scanned in Porto Alegre, and 135 were scanned in Sao Paulo. The scanning site was included as a covariate of no interest in all our analyses.
All children underwent a high-resolution axial T1 sequence (SPGR sequence) [NEX = 1, FOV = 24.0 × 18.0 cm, flip angle = 15 degrees, TE = in phase (4.2 ms), TR = 10.91 ms, matrix size = 256 × 192, and slice thickness = 1.2 mm with a maximum of 160 slices]. An axial brain MR-DTI sequence was also acquired (spin-echo diffusion weighted EPI, b = 800 s/mm2, 15 non collinear directions, TE = 99 ms, TR = 11,600 ms, flip angle = 90 degrees, matrix size = 128 × 128, NEX = 2, FOV = 24.0 cm, slice thickness = 3 mm/0 mm gap, yielding 47 slices). All children participated in playful training sessions for 30–60 min before the scan. All scans were visually inspected for quality control purposes.
Cortical thickness and diffusion tensor imaging preprocessing analyses
CT processing involved cortical reconstruction and volumetric segmentation using the FreeSurfer image analysis suite (version 5.1), which is available at http://surfer.nmr.mgh.harvard.edu/. This method uses both intensity and continuity information from the entire three-dimensional MR volume in the segmentation and deformation procedures to produce CT representations (Fischl and Dale 2000). Data were smoothed (FWHM = 10mm) (Bernal-Rusiel et al. 2010) and the statistical results were obtained using SurfStat toolbox (http://www.math.mcgill.ca/keith/surfstat/) applying a P < 0.05 threshold to find the clusters.
The diffusion tensor data were processed using FSL platform software (version 4.1.9) (Smith et al. 2006). Eddy current correction was applied to correct distortions through an affine transformation, with what each individual diffusion weighted image is aligned to the non-diffusion image. The tool also corrects for global subject head motion, since the affine transformation applies rotation and translation to each diffusion weighted image. Non-brain voxels were deleted, and FA maps were generated using the FDT tool. Tract based spatial statistics (TBSS) registration and tract skeletonization process was performed according to Smith et al. (2006). Briefly, each subject’s FA map was aligned to identify the most representative FA map of the sample, and this reference image was used as a target to non-linearly register all FA images. The entire aligned dataset was subjected to affine transformation into MNI152 standard space (1 mm3). The average of the aligned FA images was merged into a single 4D mean FA image (the group image). A mean FA skeleton was then derived from all aligned FA images, and the most relevant tracts from the spatially normalized FA map of each subject were projected onto this skeleton using a threshold of 0.2 to prevent the inclusion of non-skeleton voxels.
Movement is often a significant source of data bias in neuroimaging studies of children (Power et al. 2012). There are many methods to avoid and explore the effects of movement on DTI data (Theys et al. 2014; Yendiki et al. 2014). We used avscale FSL tool (http://fsl.fmrib.ox.ac.uk/fsl/fslwiki/FLIRT/UserGuide) to extract the mean relative displacement values (for each subject) from transformation matrix generated by Eddy Current Correction tool. Mean displacement was then regressed out in further FA analyses.
Defining regions of interest (ROIs)
Identification of DTI tracts was based on the Johns Hopkins University (JHU) atlas of white matter tractography (Hua et al. 2008; Mori et al. 2008). For each ROI tract, we obtained the mean FA value for each subject. For cortical regions, the Desikan-Killiany cortical parcellation atlas was used (Desikan et al. 2006) to obtain the mean thickness for each region and for each subject. In this study, we addressed the relationship between developmental changes in cortical regions and the tracts that connect to them. Therefore, it was necessary to define pairs of cortical regions and tracts that corresponded to one another. In other words, volume-based ROIs were deemed “connected” if they shared voxels.
First, we needed to identify the cortical regions where the soma or the ending synapses of the neurons making up the core of the tract were located. One difficulty is that the tracts terminating in one of the Desikan-Killiany regions are not usually define. As Fig. 1 shows, we used the following method to overcome this problem. We then allocated cortical regions from the Desikan-Killiany parcellation to each tract from the JHU atlas that terminated in them. After we continued the process of allocating a ‘target’ cortical region to each tract using the following two principles: 1) Cortical regions that received connections from more than one tract were allocated to one tract only – namely, the one with the largest area (proportion of connections) terminating on them (touching each specific region). Cortical target regions for a specific tract were built from the union of these allocated regions. We used this approach of allocating regions only to a single tract in order to avoid over-estimating the covariance between FA of tracts and CT of regions being driven by cortical regions contributing as target for more than one tract. However, it is unlikely that this reflects the wiring of the cortex. 2) A tract might fully end on one Desikan-Killiany cortical region but also send a few connections to another. We assumed that the first region was more related to the tract than the second, and therefore results from the first region should be more representative of changes in the neurons or synapses of that tract. Although both regions were included in the new ROI (union), the thickness was calculated using a weighted average of the CT of all the included regions for each tract, weighting them according to the proportion of the tracts that reached them. 3) When tracts had no major contribution to a cortical region, we did not initially assign any region to them. Then, we concentrated on the second largest contribution. After that, we built the equivalent volume for each of the tracts and the weighted thickness. In this way, we created 20 new target ROIs composed of the union of the ROIs from the Desikan-Killiany atlas corresponding to the 20 tracts identified by the tractography atlas (JHU) (Tables S1 and S2 available as supplementary material). These new ROIs consisted of nine pairs of inter-hemispheric homotopic tracts and regions (namely each tract and each region has the correspondent in the left and in the right hemisphere), as well as the forceps major and forceps minor.

Definition of cortical ROIs related to each tract. Thickness of cortical regions related to a specific tract were based on a weighted average of the regions in which the tract in a normative atlas terminated. For example, the thickness of target region A, which corresponds to tract A (T), corresponds to the average of R1-R5 weighted according to the number of connections finishing in each region (0.2 x CT R1 + 0.3 x CT R2 + 0.1 x CT R3 + 0.3 x CT R4 + 0.1 x CT R5). Note that each tract can have more than one region related to it, but a region would be allocated to one tract only. This implies that there is no overlap in the projections to different tracts of cortical regions. A total of nine pairs of tracts have an homotopic pair (in the contralateral hemisphere); the exceptions being forceps minor and major
Statistical analyses
-
(i)
Whole-brain analyses
-
a)
Cortical thickness: A between-subject general linear model was fitted at each surface vertex for age-related CT differences, controlling for the site of acquisition and gender. Random field theory (RFT) (Hayasaka et al. 2004) was used for cluster-wise correction for multiple comparisons with a significance level of P < 0.05 (corrected).
-
b)
FA: Specific contrast matrices were created using the FEAT tool to perform statistical analyses using a general linear model for all voxels included in the group skeleton and examining for the effects of age and controlling for the site of acquisition and gender. This statistical analysis was performed using the Randomise tool. Monte Carlo permutation-based inferences were made on unsmoothed statistical maps by applying 10,000 permutations, and cluster-like structures were enhanced using the threshold-free cluster enhancement (TFCE) algorithm (Smith and Nichols 2009). Statistical significance was defined as P < 0.05 (FWE corrected).
-
c)
Global brain changes and age: we calculated the correlation between the mean CT (regressed out the site effect) across the ROIs and age across subjects and then, the same for FA across tracts and age.
-
(ii)
Covariance analyses
Previous studies have shown that cortical thickness of certain pairs of brain regions covary across subjects, and that this pattern of coordinated growth might correspond to joint developmental changes (Alexander-Bloch et al. 2013b). We here explored whether the covariance observed between a cortical region A and B across subjects might also mirror covariance in the FA values of their respective tracts (tracts connecting A and tracts connecting B). We thus calculated the covariance in cortical thickness between all possible pairs of the twenty included CT target regions, and in a separate analysis, the twenty tracts. Correlations values were then z-Fisher normalized, and analyzed in the following way:
-
a)
It is well known that homotopic regions tend to covary in their volume and functional activity (Mechelli et al. 2005; Zielinski et al. 2010). What is still unknown is whether homotopic tracts covary in their FA. We thus performed a covariance between the inter-hemispheric homotopic and non-homotopic regions for cortical thickness regions, and separately, for white matter tracts. We compared homotopic and non-homotopic groups by calculating a t-test statistic between them, which was then compared to a null distribution of t-test statistics obtained from random permutations of the homologous labels among pairs of regions. Note that for the homotopic analyses, we considered nine pairs of tracts and target regions, excluding the forceps minor and major.
-
b)
We then explored whether high covariance in pairs of cortical regions were mirrored by high covariance between their respective tracts. Thus, we examined the similarity between the covariance of each pair of CT ROIs and the FA covariance between their corresponding white matter tracts. Statistical significance was determined by building a null distribution of correlation values where the ROIs had been randomly permuted (i.e., the correlation between the covariance of pairs of tracts and randomly chosen ROIs, without taking into account that tracts are related to the ROIs where they terminate).
-
c)
Finally, we performed an exploratory analysis of the effect of age as defined for the 3 previously mentioned groups using the covariance matrices. In other words, we explored whether covariance between regions or tracts varied in the different age groups, suggesting a change in the coordinated development of the brain at that point. We tested for differences between groups (between 7 and 8; 9 and 11; and 12 and 14 years of age) in FA and CT covariance using ANOVA, as well as by building a null model by randomly permuting the subjects into the three age groups. The uncorrected level of significance was set at P < 0.01 (two-tailed).
Results
Structural changes and age effects
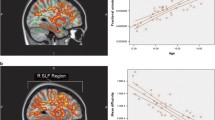
We first compared changes in CT with patient age. In agreement with the previously published literature (Shaw et al. 2008; Sowell et al. 2003; Sowell et al. 2004), voxel level analyses showed that several brain regions showed a negative relationship between CT and age (P < 0.05, cluster-corrected RFT; Fig. S2a available as supplementary material). These were primarily bilateral posterior regions (occipito-parieto-temporal cortices), but also included medial prefrontal areas. No positive relationship between CT and age was observed.
Similar results were observed when considering a single-value summarizing “average cortical brain thickness” and age. Here, the CT values of vertices within the ROIs were averaged, and mean CT across all ROIs was calculated. As shown in Fig. S2b available as supplementary material, there was a clear thinning of the cortex with age (R = −0.56, P < 10−4, permutation test).
We then explored the relationship between FA values, TBSS-identified tracts and age. In contrast with CT, but in agreement with the literature (Lebel and Beaulieu 2011; Lebel et al. 2008; Westlye et al. 2009), several clusters of voxels in different tracts showed increased FA with age (P < 0.05, FWE correction), and no voxels showed a negative relationship (Fig. S3a, available as supplementary material). A whole-brain analysis using mean FA across the tracts revealed a significant increase in FA with age (R = 0.33, P < 10−4, permutation test; Fig. S3b, available as supplementary material). Thus, just as the cortex presents anatomically distributed thinning with age, the white matter tracts show an increase in their fractional anisotropy.
Structural covariance between the ROIs in the whole group
We then focused on the covariance in brain structures among the children. As has been previously reported (Mechelli et al. 2005), pairs of homotopic (i.e., anatomically contralateral correspondent tracts) regions showed higher covariance in CT compared with non-homotopical pairs (P < 10−4, permutation test, Fig. 2a). This previously known pattern of grey matter architecture was also mirrored in our novel analogous analysis of the white matter tracts, showing that the FA covariance between pairs of homotopic tracts was higher than for non-homotopic pairs (P < 10−4, permutation test; Fig. 2b).

Symmetrical lateral organization of cortical areas and tracts. Pairs of inter-hemispheric homotopic regions (a), and, in a separate analysis, of tracts (b). Both analyses showed stronger covariance of CT and FA in homotopic pairs than non-homotopic regions (both P < 10−4 based on a permutation test)
We then investigated the relationship between the covariance of the tracts and the corresponding covariance of the cortical regions that they connected to (i.e., whether coordinated changes in the cortex were mirrored by coordinated changes in their respective tracts). There was a clear relationship between the FA covariance of the tracts and the CT covariance of the target regions connected by the tracts (R = 0.41, P < 10−4, permutation test; Fig. 3a). In other words, the coordinated morphology of grey matter closely followed the coordinated structure of white matter tracts, even during this period of great change. We also tested this relationship regressing out the effect of age in each measure. The same behavior persisted with higher covariance (R = 0.51, P < 10−4, permutation test; Fig S4 – support information). To better visualize covariance matrices tract by tract (or target region by target region), plots are available respectively of FA covariance (Fig 3b) and CT covariance (Fig 3c).

Cortical thickness covariance between regions is followed closely by FA tract covariance of their respective tract projections. (a) Scatter plot showing relationship between CT covariance and FA covariance (R = 0.41, P < 10−4, permutation test). Heatmaps detailing regional variations of the relationship of (b) FA covariance and (c) CT covariance. antTHR – anterior thalamic radiation, CS corticospinal tract, CG - cingulum (cingulate gyrus), CGH – cingulum (hippocampus), FMj – forceps major, FMn – forceps minor, infFO - inferior fronto-occipital fasciculus, infLF – inferior longitudinal fasciculus supLF R –superior longitudinal fasciculus, UF - uncinated fasciculus. supLFtemp L –superior longitudinal fasciculus
Age-related changes in structural covariance
Finally, we compared the structural covariance (CT and FA) across the three age groups (7–8; 9–11; 12–14 years old). There were no differences between the 3 groups when considering the mean value of the correlations across pairs of ROIs and tracts (P = 0.38 and P = 0.68, respectively, permutation test). As such, there was no evidence suggesting that CT in the different ROIs or FA in the different tracts increased or decreased in global symmetry over time.
Exploratory analyses revealed a few between-group differences that emerged when investigating the correlation between specific pairs of regions, and in a separate analysis, of tracts. The correlations in CT for 8 pairs of target regions significantly changed across the groups. Similarly, the correlation between FA values for 2 pairs of tracts changed significantly with age (P < 0.01, two-tailed, permutation test, uncorrected; Fig. 4a). Plots of CT covariance matrices are available in Fig S5 (support information). Further analysis revealed that all these differences but one were due to a dip in the covariance in the 9- to 11-year-old group. For both modalities, the forceps minor (or the connecting ROI) was the most frequently involved tract/region in these pairs, which changed significantly with time. This dip in the covariance was observed in the CT correlations with the forceps major, left inferior fronto-occipital fasciculus and right superior longitudinal fasciculus target regions, as well as in the FA correlations with the right cingulum (cingulate gyrus) and left inferior fronto-occipital fasciculus at P < 0.01 (two-tailed, permutation test, Fig. 4b)). Plots of FA covariance matrices are available in Fig S6 (support information). The only increase was observed in CT covariance observed was for the right anterior thalamic radiation/left uncinate fasciculus pair).

Developmental changes of the cortical and white matter structural organization. (a) Eight pairs of CT target regions showed age-related changes in their covariance. Seven pairs showed a marked dip in the 9-to 11-year-old group (P < 0.01, two-tailed, permutation test); the forceps minor was involved in three cases. (b) Two pairs of tracts showed age-related changes in FA covariance (P < 0.01, two-tailed, permutation test). Forceps minor was one of the involved tracts in both pairs. antTHR R – right anterior thalamic radiation, CG R – right cingulum (cingulate gyrus), FMj – forceps major, FMn – forceps minor, infFO L – left inferior fronto-occipital fasciculus, supLF R – right superior longitudinal fasciculus, supLFtemp L – left superior longitudinal fasciculus, UF L – left uncinated fasciculus
Discussion
Here, we present a multimodal neuroimaging study of healthy children 7 to 14 years of age, focusing on the structural properties of grey and white matter in the developing brain and the relationships between them. Consistent with previous studies, we showed that age is negatively related to CT during this period (Gogtay et al. 2004; Sowell et al. 2004), whereas fractional anisotropy is positively correlated with age (Lebel et al. 2008; Westlye et al. 2009). Other well-known characteristics of the organization of the adult brain are maintained across these age groups, including a higher similarity in CT covariance across homotopic regions compared to non-homotopic regions. Echoing this cortical organization, we also here show that pairs of homotopic tracts are more similar than non-homotopic pairs of tracts. Furthermore, we demonstrate that the coordinated morphology of cortical brain areas is closely mirrored by the coordinated microstructure of the tracts to which they connect. Overall, the relationship between grey and white matter as indexed by these two neuroimaging markers remained relatively similar across different developmental stages. This finding supports the premise that the maturational mechanisms driving these transformations in white and grey matter are closely linked.
White matter was represented using FA, which is primarily affected by axonal membranes and secondarily by myelin (Beaulieu in Johansen-Berg and Behrens (2014)). With this in mind, our data reinforce the idea that the transition from childhood to adolescence is a dynamic critical period and that CT reflects a maturational process related to changes in axon microstructure. Microstructural changes in white matter continue throughout adolescence and are associated with corresponding age-related changes in cortical gray matter regions (Giorgio et al. 2008), and it has been suggested that sustained grey matter loss during adolescence, beginning near the age of puberty, is critical for adult development, perhaps representing cortical synaptic pruning (Schmithorst and Yuan 2010). Furthermore, the onset of pubertal characteristics is thought to be an important trigger for the pruning of frontal regions, beginning around age 9 (Peper et al. 2009). Our findings are consistent with a close relationship between grey and white matter developmental trajectories. However, the similarities between CT and FA covariances were not absolute (CT and FA, R = 0.41). Thus, certain developmental processes could be specific to grey or white matter, just as direct fiber connections only partially reflect the correlations found in the CT (Gong et al. 2012). It is worth highlighting that after regressing out the age (Fig. S4, available as supplementary material) the same behavior persisted and actually the covariance between CT and FA became higher (R = 0.51). This leads us to consider the possibility that covariance might be driven by the maturational phase rather than by the age-effect. Therefore it is recommended to pursue this kind of combined analyses in future studies.
Our findings revealed some heterogeneity in the coordinated development of specific regions, with a decrease in the covariance of CT and FA in the 9- to 11-year-old group. Such results should be seen as being part of an exploratory analysis. The observed dip was detected in seven pairs of cortical regions and two tracts which displayed a decoupling (lower correlation). Thus, we could infer that they are undergoing changes (increased or decreased) at this specific age group. Though in the context of multiple comparisons we cannot rule out that developmental results could be due to chance, the clustering of these findings suggests a biological effect, which might indicate a divergence in the maturational trajectory of these specific pair of regions or tracts at this age. In addition, although the regions overlapped, differences in CT were more apparent in the regions undergoing these changes. This finding may be due to the differential sensitivities of each method, as TBSS only analyzes voxels within a predefined skeleton, which could increase its reliability at the expense of sensitivity, or the specific DTI acquisition sequence used that provides a relatively low resolution. A more biologically interesting hypothesis would be that the cortex (soma and dendrites) is more responsive to developmental plasticity than the white matter tracts (axons). Further studies might shed light on this possibility.
Sowell et al. (2003) and Gogtay et al. (2004) demonstrated that the frontal and temporal lobes are the last brain regions to mature, which is consistent with our findings showing that the occipital territories connected by the forceps major matured earlier than the temporal and frontal territories connected by other tracts involved in these pairs of divergent trajectories (forceps minor, right inferior longitudinal fasciculus, left superior longitudinal fasciculus (temporal part)). Moreover, the forceps minor and the cortical regions that it connects were the most common group of regions/tracts showing divergent developmental trajectories. The forceps minor connects the lateral and medial surfaces of the frontal lobes (Abe et al. 2004), Brodmann area 10 and frontal pole cortex (FPC) (Chao et al. 2009; Gutman et al. 2009; Hua et al. 2009; Park et al. 2008), and this region plays a pivotal role in prefrontal function (Ramnani and Owen 2004; Tsujimoto et al. 2010; Wallis 2010). Within the frontal lobe, the FPC (Badre and D’Esposito 2009; Gogtay et al. 2004) begins its maturation process earlier, at approximately the same time as the premotor cortices (between 9 and 10 years of age); however, it continues developing throughout adolescence, increasing its susceptibility to developmental abnormalities (Dumontheil et al. 2008). Consistent with these findings, a dip in the covariance involving the FPC between 9 and 11 years of age might reflect the structural decoupling of this region from an otherwise homogenous maturation network.
With respect to brain function, the implications of these changes in developmental trajectory are unclear. It has been suggested that 10-year-old children differ significantly from their younger peers in terms of their executive abilities (Tomáš Paus (2005). We hypothesize that the period between 9 and 11 years of age could be a key brain developmental phase involving changes in microstructural coordination related to cognitive control (represented by the FPC), and to some extent, language maturation (represented by temporal regions).
The current study has several limitations. To confirm the white matter changes identified using FA, one could consider using another indirect measure of axonal caliber/diameter and/or myelin content, such as radial and/or axial diffusivity or magnetization transfer measures. Our findings are based on a cross-sectional design, which is arguably not the most appropriate design for a developmental investigation. We are currently conducting a follow-up study to better investigate the developmental trajectories of various brain circuits and cortices. We also used a DTI acquisition with 15 directions that did not permit the performance of reliable tractography, leading us to use a less fine parcellation scheme. Concerning the calculation of subject’s head movement, mean displacement was calculated only using DTI images, through the transformation matrix derived from eddy current correction which is subject to criticism (Jones and Cercignani 2010) (i.e. inaccuracies in motion estimation due to the use of affine transformation to simultaneously correct for eddy current distortion and motion). The lack of B0 and gradwarp correction can also reduce the accuracy of co-registration between DTI and structural images. Moreover, our characterization of maturity in children was based solely on age, without considering other methods of ascertaining development, such as sexual maturity ratings. Despite these limitations, these results contribute to our understanding of the complex dynamics of structural covariance during the normal transition from childhood to adolescence, a period when changes in brain architecture appear to drive the main characteristics of cognitive maturation toward adulthood.
References
Abe, O., Masutani, Y., Aoki, S., Yamasue, H., Yamada, H., Kasai, K., et al. (2004). Topography of the human corpus callosum using diffusion tensor tractography. J Comp Assist Tomogr, 28(4), 533–539.
Alexander-Bloch, A., Raznahan, A., Bullmore, E., & Giedd, J. (2013a). The convergence of maturational change and structural covariance in human cortical networks. The Journal of Neuroscience, 33(7), 2889–2899. doi:10.1523/JNEUROSCI.3554-12.2013.
Alexander-Bloch, A. F., Vértes, P. E., Stidd, R., Lalonde, F., Clasen, L., Rapoport, J., et al. (2013b). The anatomical distance of functional connections predicts brain network topology in health and schizophrenia. Cereb Cortex. doi:10.1093/cercor/bhr388.
Badre, D., & D’Esposito, M. (2009). Is the rostro-caudal axis of the frontal lobe hierarchical? Nature Reviews. Neuroscience, 10(9), 659–669.
Bava, S., Thayer, R., Jacobus, J., Ward, M., Jernigan, T. L., & Tapert, S. F. (2010). Longitudinal characterization of white matter maturation during adolescence. Brain Research, 1327, 38–46. doi:10.1016/j.brainres.2010.02.066.
Bernal-Rusiel, J. L., Atienza, M., & Cantero, J. L. (2010). Determining the optimal level of smoothing in cortical thickness analysis: a hierarchical approach based on sequential statistical thresholding. NeuroImage, 52(1), 158–171.
Blakemore, S. J. (2012). Imaging brain development: the adolescent brain. NeuroImage, 61(2), 397–406. doi:10.1016/j.neuroimage.2011.11.080.
Chao, Y. P., Cho, K. H., Yeh, C. H., Chou, K. H., Chen, J. H., & Lin, C. P. (2009). Probabilistic topography of human corpus callosum using cytoarchitectural parcellation and high angular resolution diffusion imaging tractography. Human Brain Mapping, 30(10), 3172–3187.
Desikan, R. S., Segonne, F., Fischl, B., Quinn, B. T., Dickerson, B. C., Blacker, D., et al. (2006). An automated labeling system for subdividing the human cerebral cortex on MRI scans into gyral based regions of interest. NeuroImage, 31(3), 968–980. doi:10.1016/j.neuroimage.2006.01.021.
Dumontheil, I., Burgess, P. W., & Blakemore, S. J. (2008). Development of rostral prefrontal cortex and cognitive and behavioural disorders. Developmental Medicine and Child Neurology, 50(3), 168–181.
Figueiredo, V. L. M. (2001). Uma adaptação brasileira do teste de inteligência WISC-III. (Doutorado), Universidade Federal de Brasília, Brasília
Fischl, B., & Dale, A. M. (2000). Measuring the thickness of the human cerebral cortex from magnetic resonance images. Proceedings of the National Academy of Sciences of the United States of America, 97(20), 11050–11055. doi:10.1073/pnas.200033797.
Giedd, J. N. (2004). Structural magnetic resonance imaging of the adolescent brain. Annals of the New York Academy of Sciences, 1021, 77–85. doi:10.1196/annals.1308.009.
Giedd, J. N., & Rapoport, J. L. (2010). Structural MRI of pediatric brain development: what have we learned and where are we going? Neuron, 67(5), 728–734. doi:10.1016/j.neuron.2010.08.040.
Giorgio, A., Watkins, K., Douaud, G., James, A., James, S., De Stefano, N., et al. (2008). Changes in white matter microstructure during adolescence. NeuroImage, 39(1), 52–61.
Gogtay, N., Giedd, J. N., Lusk, L., Hayashi, K. M., Greenstein, D., Vaituzis, A. C., et al. (2004). Dynamic mapping of human cortical development during childhood through early adulthood. Proceedings of the National Academy of Sciences of the United States of America, 101(21), 8174–8179. doi:10.1073/pnas.0402680101.
Gong, G., He, Y., Chen, Z. J., & Evans, A. C. (2012). Convergence and divergence of thickness correlations with diffusion connections across the human cerebral cortex. NeuroImage, 59(2), 1239–1248.
Goodman, R., Ford, T., Richards, H., Gatward, R., & Meltzer, H. (2000). The development and well-being assessment: description and initial validation of an integrated assessment of child and adolescent psychopathology. Journal of Child Psychology and Psychiatry, 41(5), 645–655.
Gutman, D. A., Holtzheimer, P. E., Behrens, T. E., Johansen-Berg, H., & Mayberg, H. S. (2009). A tractography analysis of two deep brain stimulation white matter targets for depression. Biological Psychiatry, 65(4), 276–282.
Hayasaka, S., Phan, K. L., Liberzon, I., Worsley, K. J., & Nichols, T. E. (2004). Nonstationary cluster-size inference with random field and permutation methods. NeuroImage, 22(2), 676–687.
Hua, K., Zhang, J., Wakana, S., Jiang, H., Li, X., Reich, D. S., et al. (2008). Tract probability maps in stereotaxic spaces: analyses of white matter anatomy and tract-specific quantification. NeuroImage, 39(1), 336–347.
Hua, K., Oishi, K., Zhang, J., Wakana, S., Yoshioka, T., Zhang, W., et al. (2009). Mapping of functional areas in the human cortex based on connectivity through association fibers. Cerebral Cortex, 19(8), 1889–1895.
Johansen-Berg, H., & Behrens, T. E. (2014). Diffusion MRI: From quantitative measurement to in-vivo neuroanatomy (Second ed.). Amsterdam: Academic Press.
Jones, D. K., & Cercignani, M. (2010). Twenty-five pitfalls in the analysis of diffusion MRI data. NMR in Biomedicine, 23(7), 803–820.
Jones, D. K., Knosche, T. R., & Turner, R. (2013). White matter integrity, fiber count, and other fallacies: the do’s and don’ts of diffusion MRI. NeuroImage, 73, 239–254. doi:10.1016/j.neuroimage.2012.06.081.
Lebel, C., & Beaulieu, C. (2011). Longitudinal development of human brain wiring continues from childhood into adulthood. The Journal of Neuroscience, 31(30), 10937–10947. doi:10.1523/JNEUROSCI.5302-10.2011.
Lebel, C., Walker, L., Leemans, A., Phillips, L., & Beaulieu, C. (2008). Microstructural maturation of the human brain from childhood to adulthood. NeuroImage, 40(3), 1044–1055. doi:10.1016/j.neuroimage.2007.12.053.
Marsh, R., Gerber, A. J., & Peterson, B. S. (2008). Neuroimaging studies of normal brain development and their relevance for understanding childhood neuropsychiatric disorders. Journal of the American Academy of Child and Adolescent Psychiatry, 47(11), 1233–1251.
Mechelli, A., Friston, K. J., Frackowiak, R. S., & Price, C. J. (2005). Structural covariance in the human cortex. The Journal of Neuroscience, 25(36), 8303–8310. doi:10.1523/JNEUROSCI.0357-05.2005.
Mori, S., Oishi, K., Jiang, H., Jiang, L., Li, X., Akhter, K., et al. (2008). Stereotaxic white matter atlas based on diffusion tensor imaging in an ICBM template. NeuroImage, 40(2), 570–582.
Park, H. J., Kim, J. J., Lee, S. K., Seok, J. H., Chun, J., Kim, D. I., et al. (2008). Corpus callosal connection mapping using cortical gray matter parcellation and DT-MRI. Human Brain Mapping, 29(5), 503–516.
Paus, T. (2005). Mapping brain maturation and cognitive development during adolescence. Trends in Cognitive Sciences, 9(2), 60–68.
Paus, T. (2010). Growth of white matter in the adolescent brain: myelin or axon? Brain and Cognition, 72(1), 26–35.
Paus, T., Collins, D., Evans, A., Leonard, G., Pike, B., & Zijdenbos, A. (2001). Maturation of white matter in the human brain: a review of magnetic resonance studies. Brain Research Bulletin, 54(3), 255–266.
Paus, T., Keshavan, M., & Giedd, J. N. (2008). Why do many psychiatric disorders emerge during adolescence? Nature Reviews. Neuroscience, 9(12), 947–957. doi:10.1038/nrn2513.
Peper, J. S., Schnack, H. G., Brouwer, R. M., Van Baal, G. C., Pjetri, E., Szekely, E., et al. (2009). Heritability of regional and global brain structure at the onset of puberty: a magnetic resonance imaging study in 9-year-old twin pairs. Human Brain Mapping, 30(7), 2184–2196. doi:10.1002/hbm.20660.
Peters, B. D., Szeszko, P. R., Radua, J., Ikuta, T., Gruner, P., DeRosse, P., et al. (2012). White matter development in adolescence: diffusion tensor imaging and meta-analytic results. Schizophrenia Bulletin, 38(6), 1308–1317. doi:10.1093/schbul/sbs054.
Power, J. D., Barnes, K. A., Snyder, A. Z., Schlaggar, B. L., & Petersen, S. E. (2012). Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. NeuroImage, 59(3), 2142–2154.
Ramnani, N., & Owen, A. M. (2004). Anterior prefrontal cortex: insights into function from anatomy and neuroimaging. Nature Reviews. Neuroscience, 5(3), 184–194.
Raznahan, A., Lerch, J. P., Lee, N., Greenstein, D., Wallace, G. L., Stockman, M., et al. (2011). Patterns of coordinated anatomical change in human cortical development: a longitudinal neuroimaging study of maturational coupling. Neuron, 72(5), 873–884. doi:10.1016/j.neuron.2011.09.028.
Salum, G. A., Gadelha, A., Pan, P. M., Moriyama, T. S., Graeff-Martins, A. S., Tamanaha, A. C., et al. (2014). High risk cohort study for psychiatric disorders in childhood: rationale, design, methods and preliminary results. Int J Method Psychiatr Res, 24(1), 58–73. doi:10.1002/mpr.1459.
Schmithorst, V. J., & Yuan, W. (2010). White matter development during adolescence as shown by diffusion MRI. Brain and Cognition, 72(1), 16–25. doi:10.1016/j.bandc.2009.06.005.
Shaw, P., Kabani, N. J., Lerch, J. P., Eckstrand, K., Lenroot, R., Gogtay, N., et al. (2008). Neurodevelopmental trajectories of the human cerebral cortex. The Journal of Neuroscience, 28(14), 3586–3594. doi:10.1523/JNEUROSCI.5309-07.2008.
Smith, S. M., & Nichols, T. E. (2009). Threshold-free cluster enhancement: addressing problems of smoothing, threshold dependence and localisation in cluster inference. NeuroImage, 44(1), 83–98. doi:10.1016/j.neuroimage.2008.03.061.
Smith, S. M., Jenkinson, M., Johansen-Berg, H., Rueckert, D., Nichols, T. E., Mackay, C. E., et al. (2006). Tract-based spatial statistics: voxelwise analysis of multi-subject diffusion data. NeuroImage, 31(4), 1487–1505. doi:10.1016/j.neuroimage.2006.02.024.
Sowell, E. R., Peterson, B. S., Thompson, P. M., Welcome, S. E., Henkenius, A. L., & Toga, A. W. (2003). Mapping cortical change across the human life span. Nature Neurosciscience, 6(3), 309–315. doi:10.1038/nn1008.
Sowell, E. R., Thompson, P. M., Leonard, C. M., Welcome, S. E., Kan, E., & Toga, A. W. (2004). Longitudinal mapping of cortical thickness and brain growth in normal children. The Journal of Neuroscience, 24(38), 8223–8231. doi:10.1523/JNEUROSCI.1798-04.2004.
Takahashi, M., Hackney, D. B., Zhang, G., Wehrli, S. L., Wright, A. C., O’Brien, W. T., et al. (2002). Magnetic resonance microimaging of intraaxonal water diffusion in live excised lamprey spinal cord. Proceedings of the National Academy of Sciences of the United States of America, 99(25), 16192–16196. doi:10.1073/pnas.252249999.
Tamnes, C. K., Ostby, Y., Fjell, A. M., Westlye, L. T., Due-Tonnessen, P., & Walhovd, K. B. (2010). Brain maturation in adolescence and young adulthood: regional age-related changes in cortical thickness and white matter volume and microstructure. Cerebral Cortex, 20(3), 534–548. doi:10.1093/cercor/bhp118.
Tau, G. Z., & Peterson, B. S. (2010). Normal development of brain circuits. Neuropsychopharmacology, 35(1), 147–168. doi:10.1038/npp.2009.115.
Theys, C., Wouters, J., & Ghesquiere, P. (2014). Diffusion tensor imaging and resting-state functional MRI-scanning in 5-and 6-year-old children: training protocol and motion assessment. PloS One, 9(4), e94019.
Tomlinson, M., Rudan, I., Saxena, S., Swartz, L., Tsai, A. C., & Patel, V. (2009). Setting priorities for global mental health research. Bulletin of the World Health Organization, 87(6), 438–446.
Tsujimoto, S., Genovesio, A., & Wise, S. P. (2010). Evaluating self-generated decisions in frontal pole cortex of monkeys. Nature Neuroscience, 13(1), 120–126.
Wallis, J. D. (2010). Polar exploration. Nature Neuroscience, 13(1), 7–8.
Wechsler, D. (2002). WISC-III: Escala de inteligência Wechsler Para crianças: Manual. São Paulo: Casa do Psicólogo.
Westlye, L. T., Walhovd, K. B., Dale, A. M., Bjørnerud, A., Due-Tønnessen, P., Engvig, A., et al. (2009). Life-span changes of the human brain white matter: diffusion tensor imaging (DTI) and volumetry. Cerebral Cortex, 2055–2068.
Yendiki, A., Koldewyn, K., Kakunoori, S., Kanwisher, N., & Fischl, B. (2014). Spurious group differences due to head motion in a diffusion MRI study. NeuroImage, 88, 79–90.
Yurgelun-Todd, D. (2007). Emotional and cognitive changes during adolescence. Current Opinion in Neurobiology, 17(2), 251–257.
Zielinski, B. A., Gennatas, E. D., Zhou, J., & Seeley, W. W. (2010). Network-level structural covariance in the developing brain. Proceedings of the National Academy of Sciences of the United States of America, 107(42), 18191–18196. doi:10.1073/pnas.1003109107.
Acknowledgments
The opinions, hypotheses, conclusions, and recommendations of this study are the responsibilities of the authors and are not necessarily representative of the opinions of the funding agencies. The authors are grateful to the CAPES Foundation for the fellowship (Moura, L.M. 17930/12-0), the Sao Paulo Research Foundation–FAPESP (Sato, J.R. grant nos. 2013/ 10498-6 and 2013/00506-1 and Jackowski, A.P. grant no. 2013/08531-5), and CNPq, Brazil for funding this research. This study is from the National Institutes of Science and Technology for Developmental Psychiatry of Children and Adolescents (INPD) and is supported by CNPq (573974/2008-0) and FAPESP (2008/ 57896-8).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Ethics statement
The ethics committee at the University of Sao Paulo (Sao Paulo, Brazil) approved the study procedures (IORG0004884, 1138/08) and have therefore been performed in accordance with ethical standards laid down in the 1964 Declaration of Helsinki and its later amendments. Written informed consent was obtained from all parents, and all children provided verbal (or if possible, written) assent.
Rights and permissions
About this article

Cite this article
Moura, L.M., Crossley, N.A., Zugman, A. et al. Coordinated brain development: exploring the synchrony between changes in grey and white matter during childhood maturation. Brain Imaging and Behavior 11, 808–817 (2017). https://doi.org/10.1007/s11682-016-9555-0
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11682-016-9555-0