
Abstract
The stoichiometry of carbon, nitrogen and phosphorous in plants can reflect the interactions between plants and their environment. The interplay between plant nutrients, climatic factors, and soil properties and the underlying regulatory mechanisms are pillars of ecology but remain underexplored. In this study of plant C–N–P stoichiometry and nutrient resorption in Castanopsis hystrix groves in three cities (Guangzhou, Zhongshan, and Lechang) that represent an urban–rural gradient in Guangdong Province, South China, we explored potential relationships among NO2 concentrations, diameter at breast height (DBH), and resident human population. Mean annual temperature, mean annual precipitation, insolation duration per year, and the human resident population differed significantly among the three cities. Soil C-N-P was always highest in suburban Lechang, and the concentration of NO2 was highest in urban Guanghzou (55.33 ± 0.67 μg m–3) and positively correlated with the resident population and leaf N:P. Our findings suggest that C–N–P stoichiometry of C. hystrix was better explained by NO2 than by soil C–N–P stoichiometry and that nutrient resorption was better explained by leaf nutrients and DBH than by NO2 and soil stoichiometry. Our study supports the hypothesis that rapid urbanization influences NO2 concentrations and microclimate, which may jointly change the stoichiometry of plant nutrients in the forest ecosystems.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Ecological stoichiometry has been described as the bridge linking species with ecosystems (Elser et al. 1996; Zhang et al. 2019). In terrestrial ecosystems, the variations in the carbon (C), nitrogen (N) and phosphorous (P) stoichiometry in different plant organs can be used as reliable indicators of ecological function (Elser et al. 2000; Xie et al. 2020). For example, C-N-P stoichiometry can be used to predict plant growth, nutrient cycling and nutrient limitation in forest communities (Sterner and Elser 2002; Elser et al. 2007). Plant N:P ratios can help us understand the responses of vegetation to nutrient supply, phytometabolic processes, and community assembly (Vitousek 1982; Güsewell 2004). At the global scale, patterns of N and P are closely related with climatic factors such as temperature and environmental variables such as latitude (Reich and Oleksyn 2004). Numerous studies have found that human activities (e.g., urbanization, deforestation, release of fossil fuels) influence nutrient cycles, such as carbon from CO2 and nitrogen from NO2 (Jonasson et al. 1999; Lamsal et al. 2013). As a major source of N deposition influencing nutrient cycle of plant, NO2 is mainly derived from fossil energy combustion processes like transportation; thus, NO2 concentration is highly correlated with the resident human population due to increased industrialization caused by population growth (Lamsal et al. 2013). C:N:P ratios (the stoichiometry of C, N and P) are of great importance to understand nutrient cycles and biotic feedbacks in both marine and terrestrial ecosystems worldwide (McGroddy et al. 2004; Sardans et al. 2012; Bell et al. 2014). However, the mechanisms responsible for ecological stoichiometry and nutrient resorption of plant organs and soil in a specific region along an urban–rural gradient remain unclear, but are key to understanding plant responses in an era defined by rapid climate evolution and urbanization.
Soil nutrients are not only essential for plant growth, but also influence the expression of physiological and ecological functions as well (Nye 1960; Schoenholtz et al. 2000) and can be used to assess nutrient limitation in terrestrial ecosystems. Globally, the C:N:P atomic ratio of soils has been shown to be roughly 186:13:1 (Cleveland and Liptzin 2007). This general estimated ratio of nutrient allocation provides a background value for specific investigations. However, C:P and N:P ratios are highly variable across space, climatic zone, soil depth and type (Millard and Grelet 2010), and weathering stage (Tian et al. 2010). Ecological stoichiometry can explain the state of plant nutrient resorption in ecosystem dynamics and functioning (Sterner and Elser 2002; Fan et al. 2015; Zechmeister-Boltenstern et al 2015). Plant nutrient resorption, the process by which nutrients from aged tissues are mobilized to other tissues, prolongs nutrient retention by the plant, improves the utilization efficiency of the nutrients, and may reduce plant dependency on soil nutrients (Vergutz et al. 2012; Zhang et al. 2015). Multiple studies have found that a large portion of the nutrient requirements of perennial plants are obtained through resorption (Aerts 1996; Reed et al. 2012).
Castanopsis hystrix, one of the most important evergreen tree species in South China, is primarily distributed throughout the provinces of Fujian, southern Hunan, Guangdong, Guangxi, and southwestern Guizhou, as well as in southeastern Tibet (Li et al. 2007). C. hystrix is a dominant landscape species often grown in commercial stands for luxury furniture (Li 1996; Li et al. 2007). In this study, we focused on C. hystrix groves in three Chinese subtropical forests along urban–rural gradients to answer two fundamental questions: (1) How do NO2 concentrations correlate with the C–N–P stoichiometry in the plant? (2) How is the C-N-P stoichiometry in different organs related to soil and climatic factors? The answers will improve our understanding of nutrient cycling in C. hystrix plantations in subtropical regions, South China. Determining the C-N-P status in different organs and soil of C. hystrix forests will help us understand nutrient cycling processes in the plantations in South China and design appropriate management strategies.
Materials and methods
Study site
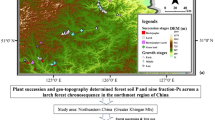
We selected five sampling locations in three subtropical forests along the urban–rural gradients in Guangdong Province, South China (Fig. 1). Specifically, a mixed population of C. hystrix and S. superba (G1), a monoculture of C. hystrix (G2), and a mixed population of C. hystrix and Acacia mangium (G3) of Longdong Forest Farm in Guangzhou city (highly urbanized); a mixed population of C. hystrix and S. superba (Z1) of Wugui Mountain in Zhongshan city (moderately urbanized); and a monoculture of C. hystrix (L1) of Longshan Forest Farm in Lechang city (less urbanized). Stand age for the sampled populations were relatively similar: 18 years in Lechang and Zhongshan and 20 years in Guangzhou according to records from earlier field investigations. All stands were fertilized and managed for 2 years before being allowed grow naturally, and all soil types were classified as red soil with similar acid pH (4.47, 4.58 and 4.86 in the city of Guangzhou, Zhongshan and Lechang, respectively). All sites have a subtropical monsoon climate with an annual mean precipitation between 1336–1777 mm and a mean annual temperature between 18.7–23.4 °C.

Research areas and sites of the Castanopsis hystrix plantations sampled in Guangdong Province, South China. Red triangles represent the Longdong Forest Farm in Guangzhou, Wugui Mountain in Zhongshan, and Longshan Forest Farm in Lechang
Sample collection and data processing
The plots in Guangzhou city, Zhongshan city and Lechang city were 20 m × 30 m, 20 m × 20 m and 30 m × 40 m, respectively. At each site, three replicate plots with similar topography and minimal human interference were established. The position and direction of slope for these plots were the same, and C. hystrix individuals of similar age were selected. We measured the diameter at breast height (DBH > 2 cm), tree height, and height under branches (HUB). Three representative trees were selected to sample leaves and branches for each forest plot from five places. Samples were taken from healthy pest-free foliage and branches from the upper and outer portions of the crown. Three litter samples of fresh and undecomposed material were randomly collected from the soil surface to better understand nutrient resorption in the forest site. Three soil cores (6 cm in diameter) in each stand were taken 0–20 cm depth after removing the surface litter.
Regional data of mean annual temperature (MAT), mean annual precipitation (MAP), insolation duration per year, and resident human population (RP) were compiled from China’s National Bureau of Statistics (http://www.stats.gov.cn/) for the years of 2015–2017. Simultaneously, regional NO2 concentrations were retrieved from the Municipal Eco-Environment Bureau of Guangzhou (http://www.gzepb.gov.cn/), Zhongshan (http://www.zsepb.gov.cn) and Shaoguan (http://epb.sg.gov.cn/) during the same period.
C, N, and P analyses
Branches and leaves were dried at 105 °C for 15 min and then at 75 °C for 72 h to constant mass (Zheng et al. 2010). Soil samples were sieved (2 mm mesh) and air-dried. Leaf and soil C were measured via external heating of potassium dichromate (Kalembasa and Jenkinson 1973). Leaf N and P were measured using H2SO4 + H2O2 digestion, then distillation titration for N and vanadium molybdenum yellow colorimetry for P (Parkinson and Allen 1975). Total soil N and P were measured by the micro-Kjeldahl distillation titration method and the NaOH melting molybdenum-antimony anticolorimtric method, respectively (Parkinson and Allen 1975).
Data analyses
Nutrient resorption efficiency (NuRE) of N and P were calculated as:
where MLCF (0.780) is the mass loss correction factor for evergreen woody angiosperms (Vergutz et al. 2012), and Nle and Nli represent the concentrations of leaf and litter, respectively. SPSS 19.0 (IBM, Armonk, NY, USA) was used for one-way analyses of variance (ANOVAs) to compare the variation of C, N, P stoichiometry and NuRE of plant-soil. Origin 2017 (OriginLab, Northampton, Massachusetts, USA) was used to draw graphs. R-studio (Public Benefit Corporation, Boston, Massachusetts, USA) was used for hierarchical partitioning analysis (HP), which was used to measure the explanatory value of independent variables.
Results
Variation of regional climatic and social factors
NO2 concentrations in Guangzhou city (55.33 ± 0.67 μg m–3; mean ± SE) were higher than in Zhongshan city (46.00 ± 1.53 μg m–3) and Lechang city (50.67 ± 0.88 μg m–3), based on public data from 2015 to 2017. Climatic factors of MAP, MAT, insolation duration per year and resident population (RP) were significantly different across the three localities (p < 0.05) (Table 1).
Soil C-N-P stoichiometry in C. hystrix plantations
Significant differences were detected in soil C–N–P stoichiometry at the 0–20 cm depth among the three forests. Soil C-N-P was highest in Lechang and lowest in Zhongshan, while soil C:N, C:P and N:P were lowest in Lechang and highest in Zhongshan. Soil N concentration was not significantly different between Guangzhou and Zhongshan (Table 2).
Plant C-N-P stoichiometry in C. hystrix plantations
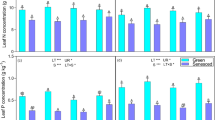
N and P concentrations and C:N, C:P and N:P ratios varied among branch, leaf, and litter samples, but did not differ significant among the cities (Fig. 2b–f). Plant C concentration and C:N were higher in Zhongshan than in Lechang and Guangzhou (Fig. 2a, d). Plant N and N:P were higher in Guangzhou than in Lechang and Zhongshan (Fig. 2b, f). Plant P concentrations were higher and C:P were lower in Lechang compared to Guangzhou (Fig. 2c, e).

Plant C-N-P stoichiometry in branch, leaf, and litter samples from plantations of Castanopsis hystrix at three sites in South China. See Notes with Table 2 for stand types and urbanization level. Different letters above bars for the same sample type (n = 3) indicate a significant difference among stand types (n = 5) based on a one-way ANOVA (p = 0.05)
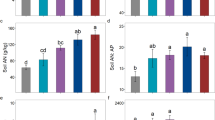
Regression and HP analysis (Table 3) showed that most plant stoichiometric measures could be explained by a model including DBH, NO2, and soil stoichiometry. Furthermore, of the measures that could be explained by our model, DBH and NO2 explained the majority of the variance, with the exception of leaf C concentration, where soil stoichiometry had the strongest predictive power. Branch C, branch C:P and leaf N:P increased linearly with NO2 (Fig. 3a, f, g). Branch/leaf P and C:P decreased linearly with NO2 (Fig. 3b, c) and with DBH (Fig. 3d, e).

Relationships between NO2 (a–c, g, f) and DBH (d, e) with plant stoichiometry from plantations of Castanopsis hystrix at three sites in South China. See Notes with Table 2 for stand types and urbanization level
Characteristics of nutrient resorption
Litter N concentrations were higher in Guangzhou than in Zhongshan and Lechang, while litter C:N was higher in Zhongshan than in Lechang and Guangzhou (Fig. 2b, d). NRE (N nutrient resorption efficiency) and PRE (P nutrient resorption efficiency) differed significantly among the three sites (Fig. 4). With the exception of litter N:P, the full model that included DBH, NO2 and leaf and soil stoichiometries explained the majority of the variance in litter stoichiometry and NuRE (Table 4). DBH and leaf stoichiometry were the most predictive independent variables for all dependent variables except for litter N:P, for which the model had poor predictive power. Soil stoichiometry was not the most predictive variable. Litter C, N and P concentrations were negatively correlated with DBH, leaf N concentration, and DBH, respectively (Fig. 5a–c). Litter and leaf C:N were also negatively correlated (Fig. 5d). NRE was positively correlated with leaf N concentration (Fig. 5e), and PRE was positively correlated with DBH (Fig. 5f).

Variation in nutrient resorption efficiency (NuRE) in Castanopsis hystrix plantations in Guangzhou, Zhongshan, and Lechang, which differed in their urbanization levels. See Notes with Table 2 for stand types

The relationships from plantations of Castanopsis hystrix at three sites in South China between DBH and litter stoichiometry (a, c), litter and leaf stoichiometries (b, d), and nutrient resorption and leaf stoichiometry and DBH (e, f). See Notes with Table 2 for stand types and urbanization level
Discussion
C-N-P stoichiometry and interactions in soil
Soil characteristics are influenced by the diversity of soil microbes, spatial heterogeneity, and structural complexity. Although the global C:N:P ratios (186:13:1) of soils may be stable (Cleveland and Liptzin 2007), local climate and soil formation affect the stoichiometry in soils (Tian et al. 2010), creating regional and local heterogeneity. Furthermore, the same plant species in different stand types may have different nutrient concentrations. Here we found that C:N ranged from 22.09 to 28.97, C:P from 81.87 to 297.48, and N:P from 3.67 to 10.49, values that differed significantly from previous means of 14.4, 136 and 9.3 in China’s soil (Tian et al. 2010). These differences may be caused by differences in tree species and climate zones. Soil C-N-P concentrations were highest in Lechang and lowest in Zhongshan, which may be due to latitudinal differences (Sardans et al. 2011), vegetation age (Bui and Henderson 2013), and understory vegetation diversity (Small and McCarthy 2005). Hence, soil stoichiometry was likely influenced by complex factors (Huang et al. 2016).
Our study shows that soil stoichiometry played a minor role in the C–N–P content in branches and leaves, and NO2 was often the most predictive variable. However, many studies have found that plant nutrients are sensitive to soil nutrients (Schreeg et al. 2014; Tang et al. 2018). Although plant and soil nutrient concentrations are usually correlated, at least in some organs (He et al. 2016; Erinle et al. 2020), the relationship maybe confounded by air pollution and other climatic factors that influence forest nutrient cycling. Nutrient cycling has been shown to be a complicated process involving plant-soil interactions (Zechmeister-Boltenstern et al. 2015).
Ecological stoichiometry of plant organs and relationships with environmental factors
N concentrations and N:P ratios in branches and leaves were higher in Guangzhou than in Zhongshan and Lechang, but P concentration in branches was lower in Guangzhou. P concentrations in branches and leaves were significantly negatively correlated with NO2. N:P ratios of branches and leaves were significantly positively correlated with NO2, indicating that NO2 can reflect the status of plant nutrient P. Global change, N deposition and anthropogenic activities are primary factors affecting N:P ratios in leaves (Güsewell 2004; Sardans et al. 2016). A regional study showed a positive correlation between MAT and C:P and N:P in branches of 803 plant species in China (Zhang et al. 2018b). In our study of the three sites, mean leaf N–P ranged from 0.75 to 0.89 g kg–1, and N:P from 18.17 to 31.11. Leaf P concentration was lower than found in a meta-analysis of leaf P in terrestrial species in China (1.21 g kg–1) (Han et al. 2005). In addition, leaf N concentrations were lower than the mean for 753 Chinese terrestrial species (18.6 g kg–1; Han et al. 2005), except for the mixed population of C. hystrix and A. mangium from Longdong Forest Farm in Guangzhou city (25.62 g kg–1). However, the leaf N:P ratio was higher than in the Han et al. (2005) study (14.4).
N:P ratios in plants are influenced by species composition, community, plant age, etc. (Güsewell 2004). In our study, in leaves, C:N ranged from 28.99.28 to 33.03 and C:P from 571.10 to 1007.40. We also found that leaf C:N in this study was lower than the global mean for broadleaf forests (35.1 ± 3.7), but C:P (922.3 ± 77.3) was within the global range (McGroddy et al. 2004). N:P ratios of plant above 16 suggest P limitation; those below 14 indicate N limitation (Meuleman 1996; Wang et al. 2018). The N:P ratios of leaves collected from the C. hystrix populations were greater than 16 across all sites, suggesting that P limitation is widespread in the region. Our results show that NO2 correlates with plant N content and significantly influences P content and N:P ratios. Therefore, NO2 concentration may be used as an indicator of plant N and P status.
Nutrient resorption and influencing factors
Atmospheric NO2 concentrations often exhibit variations across diverse cities, as in Guangdong Province in South China. Our study confirmed that NO2 was highest in Guangzhou and increased significantly with population growth. Plants distribute nutrients to leaves for growth (Sardans and Peñuelas 2013) and often use the nutrients stored in branches when nutrients are limited (Heineman et al. 2016; Yan et al. 2016). Litter P content was lower in our study compared to the global mean (0.85 g kg–1) (Kang et al. 2010). In contrast, N content and N:P ratios were both higher than the global means (10.9 g kg–1 and 18.3, respectively) (Kang et al. 2010). Litter composition and stoichiometry are important factors in litter decomposition. Specifically, N content and C:N of litter strongly affects decomposition (Leitner et al. 2012). Usually, net N release from litter decomposition begins when litter C:N is less than 40 (Parton et al. 2007). Litter decomposition rate is negatively correlated with C:N and C:P; lower C:N and C:P typically correspond to faster decomposition rates (Enríquez et al. 1993). MAP and NO2 are key factors influencing litter N content, and litter N content follows a linear relationship with MAP (Liu et al. 2006; Yuan and Chen 2009b). Old leaves and branches may be more sensitive than young ones to N content (Di et al. 2018), and here, N and P were higher in leaves than in branches and litter.
Foliar N content was significantly positive correlated with NRE (p = 0.042), but PRE was not correlated with leaf P content. However, some studies have found the opposite trend, with P concentrations in leaves positively correlated with PRE, and a lack of correlation between NRE and N content in a Metasequoia glyptostrobodies forest (Zhang et al. 2018a). Here, we did not find any significant correlations between either NRE or PRE and MAP. However, a contrasting report suggested that NRE increases, while PRE decreases, with MAP at a global scale (Yuan and Chen 2009a).
Conclusions
Plant N:P in branches and leaves were positively correlated with NO2 in C. hystrix populations. Variance in nutrient resorption was better explained by leaf nutrients and DBH than by NO2 and soil ecological stoichiometry. Our results highlight that NO2 concentration decreased along the urban–rural gradient exemplified by Guangzhou–Zhongshan–Lechang in Guangdong Province, South China. The effect of NO2 concentration was greater than soil stoichiometry on plant nutrient cycling.
References
Aerts R (1996) Nutrient resorption from senescing leaves of perennials: Are there general patterns? J Ecol 84:597–608
Bell C, Carrillo Y, Boot CM, Rocca JD, Pendall E, Wallenstein MD (2014) Rhizosphere stoichiometry: are C:N: P ratios of plants, soils, and enzymes conserved at the plant species-level? New Phytol 201:505–517
Bui EN, Henderson BL (2013) C:N: P stoichiometry in Australian soils with respect to vegetation and environmental factors. Plant Soil 373:553–568
Cleveland CC, Liptzin D (2007) C:N: P stoichiometry in soil: is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 85:235–252
Di YB, Fu XL, Wang HM, Li WH, Wang SL (2018) N concentration of old leaves and twigs is more sensitive to stand density than that of young ones in Chinese fir plantations: a case study in subtropical China. Journal of Forestry Research 29:163–169
Elser JJ, Bracken MWS, Cleland EE, Gruner DS, Harpole WS, Hillebrand H, Ngai JT, Seabloom EW, Shurin JB, Smith JE (2007) Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol Lett 10:1135–1142
Elser JJ, Dobberfuhl DR, MacKay NA, Schampel JH (1996) Organism size, life history, and N: P stoichiometry. Bioscience 46:674–684
Elser JJ, Sterner RW, Gorokhova E, Fagan WF, Markow TA, Cotner JB, Harris on JF, Hobbie SE, Odell GM, Weider LW, (2000) Biological stoichiometry from genes to ecosystems. Ecol Lett 3:540–550
Enríquez S, Duarte CM, Sand-Jensen K (1993) Patterns is decomposition rates among photosynthetic organism: the importance of detritus C:N: P content. Oecologia 94:457–471
Erinle KO, Marschner P (2020) Phosphorus and nitrogen in the soil interface between two plant residues differing in C/nutrient ratio: a short-term laboratory incubation study. Soil Ecology Letters 2:188–194
Fan HB, Wu JP, Liu WF, Yuan YH, Hu L, Cai QK (2015) Linkages of plant and soil C:N: P stoichiometry and their relationships to forest growth in subtropical plantations. Plant Soil 392:127–138
Güsewell S (2004) N: P ratios in terrestrial plants: variation and functional significance. New Phytol 164:243–266
Han WX, Fang JY, Guo DL, Zhang Y (2005) Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytol 168:377–385
He XJ, Hou EQ, Liu Y, Wen DZ (2016) Altitudinal patterns and controls of plant and soil nutrient concentrations and stoichiometry in subtropical China. Scientific Reports 6:24261
Heineman KD, Turner BL, Dalling JW (2016) Variation in wood nutrients along a tropical soil fertility gradient. New Phytol 211:440–454
Huang ZQ, Liu B, Davis M, Sardans J, Peñuelas J, Billings S (2016) Long-term nitrogen deposition linked to reduced water use efficiency in forests with low phosphorus availability. New Phytol 210:431–442
Jonasson S, Michelsen A, Schmidt IK (1999) Coupling of nutrient cycling and carbon dynamics in the Arctic, integration of soil microbial and plant processes. Appl Soil Ecol 11:135–146
Kalembasa SJ, Jenkinson DS (1973) A comparative study of titrimetric and gravimetric methods for the determination of organic carbon in soil. J Sci Food Agric 24:1085–1090
Kang HZ, Xin ZJ, Berg B, Burgess PJ, Liu QL, Liu ZC, Li ZH, Liu CJ (2010) Global pattern of leaf litter nitrogen and phosphorus in woody plants. Ann For Sci 67:811–811
Lamsal LN, Martin RV, Parrish DD, Krotkov NA (2013) Scaling relationship for NO2 pollution and urban population size: a satellite perspective. Environ Sci Technol 47:7855–7861
Leitner S, Wanek W, Wild B, Haemmerle I, Kohl L, Keiblinger KM, Boltenstern SZ, Richter A (2012) Influence of litter chemistry and stoichiometry on glucan depolymerization during decomposition of beech (Fagus sylvatica L.) litter. Soil Biol Biochem 50:174–187
Li J, Ge XJ, Cao HL, Ye WH (2007) Chloroplast DNA diversity in Castanopsis hystrix populations in south China. For Ecol Manage 243:94–101
Li JQ (1996) The origin and distribution of the family Fageceae. Journal of Systematics and Evolution 34:3763–3796
Liu CJ, Berg B, Kutsch W, Westman CJ, Ilvesniemi H, Shen XH, Shen GR, Chen XB (2006) Leaf litter nitrogen concentration as related to climatic factors in Eurasian forests. Glob Ecol Biogeogr 15:438–444
McGroddy ME, Daufresne T, Hedin LO (2004) Scaling of C:N: P stoichiometry in forests worldwide: implications of terrestrial Redfield-type ratios. Ecology 85:2390–2401
Meuleman WKAFM (1996) The vegetation N: P ratio: a new tool to detect the nature of nutrient limitation. The Journal of Applied Ecology 33:1441–1450
Millard P, Grelet GA (2010) Nitrogen storage and remobilization by trees: ecophysiological relevance in a changing world. Tree Physiol 9:1083–1095
Nye PH (1960) Organic matter and nutrient cycles under moist tropical forest. Plant Soil 13:333–346
Parkinson JA, Allen SE (1975) A wet oxidation procedure suitable for the determination of nitrogen and mineral nutrients in biological material. Commun Soil Sci Plant Anal 6:1–11
Parton W, Silver WL, Burke IC, Grassens L, Harmon ME, Currie WS, King JY, Adair EC, Brandt LA, Hart SC, Fasth B (2007) Global-scale similarities in nitrogen release patterns during long-term decomposition. Science 315:361–364
Reed SC, Townsend AR, Davidson EA, Cleveland CC (2012) Stoichiometric patterns in foliar nutrient resorption across multiple scales. New Phytol 196:173–180
Reich PB, Oleksyn J (2004) Global patterns of plant leaf N and P in relation to temperature and latitude. Proceeding of the National Academy Sciences 101:11001–11006
Sardans J, Alonso R, Janssens IA, Carnicer J, Vereseglou S, Rilliget MC, Martinea MF, Sanders TGM, Peñuelas J (2016) Foliar and soil concentrations and stoichiometry of nitrogen and phosphorous across European Pinus sylvestris forests: relationships with climate, N deposition and tree growth. Funct Ecol 30:676–689
Sardans J, Peñuelas J (2013) Tree growth changes with climate and forest type are associated with relative allocation of nutrients, especially phosphorus, to leaves and wood. Glob Ecol Biogeogr 22:494–507
Sardans J, Rivas-Ubach A, Peñuelas J (2011) Factors affecting nutrient concentration and stoichiometry of forest trees in Catalonia (NE Spain). For Ecol Manage 262:2024–2034
Sardans J, Rivas-Ubach A, Peñuelas J (2012) The C:N: P stoichiometry of organisms and ecosystems in a changing world: a review and perspectives. Perspectives in Plant Ecology, Evolution and Systematics 14:33–47
Schoenholtz SH, Miegroet HV, Burger JA (2000) A review of chemical and physical properties as indicators of forest soil quality: challenges and opportunities. For Ecol Manage 138:335–356
Schreeg LA, Santiago LS, Wright SJ, Turner BL (2014) Stem, root, and older leaf N: P ratios are more responsive indicators of soil nutrient availability than new foliage. Ecology 95:2062–2068
Small CJ, McCarthy BC (2005) Relationship of understory diversity to soil nitrogen, topographic variation, and stand age in an eastern oak forest, USA. For Ecol Manage 217:229–243
Sterner RW, Elser JJ (2002) Ecological stoichiometry: the biology of elements from molecules to the biosphere. Princeton University Press, Princeton, p 439
Tang ZY, Xu WT, Zhou GY, Bai YF, Li JX, Tang XL, Chen DM, Liu Q, Ma WH, Xiong GM, He HL, He NP, Guo YP, Guo Q, Zhu JL, Han WX, Hu HF, Fang JY, Xie ZQ (2018) Patterns of plant carbon, nitrogen, and phosphorus concentration in relation to productivity in China’s terrestrial ecosystems. Proc Natl Acad Sci 115:4033–4038
Tian HQ, Chen GS, Zhang C, Melillo JM, Hall CAS (2010) Pattern and variation of C:N: P ratios in China’s soils: a synthesis of observational data. Biogeochemistry 98:139–151
Vergutz L, Manzoni S, Porporato A, Novais RF, Jackson RB (2012) Global resorption efficiencies and concentrations of carbon and nutrients in leaves of terrestrial plants. Ecol Monogr 82:205–220
Vitousek P (1982) Nutrient cycling and nutrient use efficiency. Am Nat 119:553–572
Wang N, Fu FZ, Wang BT, Wang RJ (2018) Carbon, nitrogen and phosphorus stoichiometry in Pinus tabulaeformis forest ecosystems in warm temperate Shanxi province, North China. Journal of Forestry Research 29:1665–1673
Xie J, Yan QL, Yuan JF, Li R, Lü XT, Liu SL, Zhu JJ (2020) Temporal effects of thinning on the leaf C:N: P stoichiometry of regenerated broadleaved trees in larch plantations. Forests 11:54
Yan ZB, Li P, Chen YH, Han WX, Fang JY (2016) Nutrient allocation strategies of woody plants: an approach from the scaling of nitrogen and phosphorus between twig stems and leaves. Scientific Reports 6:20099
Yuan ZY, Chen HYH (2009a) Global-scale patterns of nutrient resorption associated with latitude, temperature and precipitation. Glob Ecol Biogeogr 18:11–18
Yuan ZY, Chen HYH (2009b) Global trends in senesced-leaf nitrogen and phosphorus. Glob Ecol Biogeogr 18:532–542
Zechmeister-Boltenstern S, Keiblinger KM, Mooshammer M, Peñuelas J, Richter A, Sardans J, Wolfgang W (2015) The application of ecological stoichiometry to plant-microbial-soil organic matter transformations. Ecol Monogr 85:133–155
Zhang H, Wang JN, Wang JY, Guo ZW, Wang GG, Zeng DH, Wu TG (2018a) Tree stoichiometry and nutrient resorption along a chronosequence of Metasequoia glyptostroboides forests in coastal China. For Ecol Manage 430:445–450
Zhang JH, He NP, Liu CC, Xu L, Chen Z, Li Y, Wang RM, Yu GR, Sun W, Xiao CW, Chen HYH, Reich PB (2019) Variation and evolution of C: N ratio among different organs enable plants to adapt to N-limited environments. Glob Change Biol 26:2534–2543
Zhang JH, Tang ZY, Luo YK, Chi XL, Chen YH, Fang JY, Shen HH (2015) Resorption efficiency of leaf nutrients in woody plants on Mt. Dongling of Beijing, North China. Journal of Plant Ecology 8:530–538
Zhang JH, Zhao N, Liu CC, Yang H, Li ML, Yu GR, Wilcox K, Yu Q, He NP (2018b) C:N: P stoichiometry in China’s forests: from organs to ecosystems. Funct Ecol 32:50–60
Zheng YH, Xu XB, Simmons M, Zhang C, Gao FJ, Li ZJ (2010) Responses of physiological parameters, grain yield, and grain quality to foliar application of potassium nitrate in two contrasting winter wheat cultivars under salinity stress. J Plant Nutr Soil Sci 173:444–452
Acknowledgements
We appreciate Mr. Xu Wang and Xinhua Luo for their help with fieldwork, three anonymous reviewers for their valuable comments, and Stephen Murphy and Elizabeth Tokarz at Yale University for their assistance with English language and grammatical editing of the manuscript.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Project funding: The work was supported by the National Key Research and Development Program of China (2017YFC0505606), National Natural Science Foundation of China (31570594, 31600449), and Forestry Science and Technology Innovation Project of Guangdong Province (2021–2023).
The online version is available at http://www.springerlink.com.
Corresponding editor: Tao Xu.
Rights and permissions
About this article

Cite this article
Li, F., Sun, B., Shi, Z. et al. Changes in ecological stoichiometry and nutrient resorption in Castanopsis hystrix plantations along an urbanization gradient in the lower subtropics. J. For. Res. 32, 2323–2331 (2021). https://doi.org/10.1007/s11676-021-01293-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-021-01293-0