
Abstract
Understanding the influence of environmental variables on the spatial distribution of ecological communities is essential to predict the response of vegetation to various environmental drivers. Ecological theory suggests that multiple environmental factors shape local species assemblages and should influence the various components of community structure and composition in different ways. This study aimed to classify Pinus wallichiana dominated forests in the Swat Hindukush range mountains to understand the relative influence of multiple environmental filters on its composition and structure. These forests represent the most typical of the species distribution in northern Pakistan and were not subjected to any phytosociological study. For this purpose, thirty forest stands, spanning a wide range of physical habitats were sampled using 10 × 10 m plots and the importance value index was calculated. The floristic and environmental data were subjected to Ward’s agglomerative cluster analysis for objective classification and ordinated with NMS ordination for pattern description and testing the vegetation–environmental relationships. Three floristically and ecologically distinct communities were recognized along the topographic gradient (elevation, r = 0.377; slope, r = 0.5548) coupled with soil physical (clay, r = 0.2782; silt, r = 0.3225) and chemical properties (pH, r = 0.4975; lime, r = 3982). An elevation gradient of 100 m separated the low (Pinus wallichiana–Quercus dilatata community) and middle elevations forest stands (P. wallichiana pure population) from the highland population type (Pinus wallichiana–Cedrus deodara community). The floristics and structure of these forest types respond directly or indirectly to topographic and soil variables which were evidenced from the floristic composition, species richness, and community physiognomy. These characteristics of the communities changed from heterogenous, dense stands to sparsely dispersed conifers, broadleaved-evergreen, and deciduous vegetation types along the environmental gradients. We concluded that several factors explained the spatial pattern. Thus, we predict that elevation coupled with soil physical and chemical properties could be considered if the ultimate goal is the restoration and conservation of these forests. We also recommend broad-scale phytosociological study on P. wallichiana forests including multiple sampling from diverse locations to define the floristic diversity and to better understand its ecological requirements.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Pinus wallichiana is one of the three naturally distributed pines in Pakistan and besides its relict distribution at Baluchistan, it mainly appears in Himalaya, Karakoram and Hindukush mountains from eastern Afghanistan across northern Pakistan and India to Yunnan in southwest China (Yadav et al. 1997; Ahmed et al. 2006). This species is a major component of the middle to high elevation and grows at elevations from 1800 to 3400 m (rarely as low as 1200 m), varying between 5 and 18 m in height (Farjon 2013). It covers a substantial area and is one of the characteristic elements of both dry and moist temperate forests in Pakistan (Khan et al. 2014). P. wallichiana usually grows on deep moist soils (Yadav et al. 1997) but also grows well on fertile, well drained sandy-clay to sandy-clay-loamy soils (Sheikh 1993).
One site supporting these natural forests is District Swat of the Hindukush range mountains (HRM) which has been the focus of various organizations and tourists due to the high timber quality, associated wildlife, and abundant non-timber forest products (NTFPs) (Baig et al. 2008). It forms both pure and mixed stands depending on large-scale ecological gradients (Wahab 2011). These stands have not been classified phytosociologically and their correlations with environmental parameters are not yet explored. Elucidation of such relationships is fundamental to understanding plant species composition and structure in a particular habitat, landscape, and region (e.g. Barbour et al. 1987; Mucina 1997; Burke 2001). Generally, plant communities are dynamic entities undergoing continuous changes in response to environmental factors and intrinsic dynamics within the community (e.g. Ma et al. 2012; Ren et al. 2013).
Communities and habitats in transition zones are potentially affected by climate change and thus are often the first communities to shift, especially in areas providing marginal conditions for plant growth. This phenomenon prevails in the Swat mountains which is an ecotone between the moist and dry temperate areas of Pakistan (Champion et al. 1965). These mountains form part of the Great Hindukush escarpment, an extension of the Hindu Raj and Himalaya lies in the Sino-Japanese phyto-geographical region that extended to Afghanistan and the Karakorum Range of Gilgit-Baltistan (Ahmed et al. 2006). Climatic, topographic and soil factors are believed to be the overriding factors determining vegetation patterns (Siddiqui et al. 2010; Ahmed et al. 2011). The variety of habitats, edaphic conditions and its transitional position between moist and dry parts make the Swat Hindukush range mountains an ideal study area to test whether these variables are the driving factors of vegetation structure? Steep climatic gradients, and high-elevation habitats can form an important buffer for climate-induced changes (Chapin et al. 1998; Zhang and Zhang 2007). These forests are being degraded by deforestation, overgrazing and clearing of land for terrace cultivation (Hussain et al. 1997; Sher et al. 2010; Ahmed et al. 2010; Shaheen et al. 2011), thus documenting existing plant communities and understanding their relationships with the environment is critical. Although an early inventory of plant species was compiled for the study area (Stewart 1967; Beg and Khan1984; Ali 2011; Rahman 2012; Ilyas et al. 2012), vegetation parameters were never quantified except by Siddiqui et al. (2016), Ilyas et al. (2015), and Rahman et al. (2017). These authors focused on understory vegetation, leaving woody vegetation to be either ignored or studied sporadically. Some studies of other issues such as ethnobotany and species niche modeling were also undertaken (Ahmed et al. 2010; Sher et al. 2010; Qasim et al. 2012; Akhtar et al. 2013, 2014, 2015). However, no comprehensive work has been done on the classification of woody vegetation and its correlations with environmental variables although the area has both dry and moist forest thickets with a variety of vegetation types. Thus, this study was aimed to document the existing plant communities of P. wallichiana populations and to investigate major ecological and environmental gradients responsible for its distribution in Swat Hindukush mountains. This study will help to understand to predict the development of P. wallichiana plant communities and their potential determinants because of its significant ecosystem value as a reference across the northern areas of Pakistan and adjoining countries.
Materials and methods
Study area
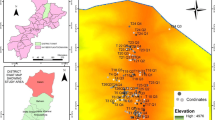
Our study sites were located in two contrasting climatic zones, i.e., moist and dry temperate forests, in the Swat Hindukush mountains of northern Pakistan (Fig. 1). These sampling sites were frequently distributed in the subalpine zone just above the Pinus roxburghii and admixed with dry and moist temperate evergreen forests in the Sino-Japanese phytogeographical region of Pakistan (Champion et al. 1965). The total area of the district is 5337 km2 (lat. 34°34′–35°55′N, long. 72°08′–72°50′E (Shinwari et al. 2003). The main geological formations are Hornblenditic and Schistose rocks (Zeb 1970) characterized by felsic zones. Low elevation portions of the study area were rich in alluvial soils (Hussain and Illahi 1991) and supported cereal crops, vegetable and fruit orchards (Ilyas et al. 2015).

Locations of forest stands sampled in the Hindukush mountains ranges of district Swat, Pakistan
The climate is under the strong influence of elevation gradient and generally falls in the temperate zone and a humid subtropical climate under the Köppen climate classification (Cfa) with four distinct seasons. Cold winters with heavy snowfall accumulation in the form of glaciers are the characteristic feature of the area at high elevations, where summers are mild and short (Sher et al. 2010). Annual precipitation varies from 693 to 897 mm and temperature ranges from 13.4 to 21.7 °C (data source PMD; http://www.pmd.gov.pk). November and April are the driest and wettest months, respectively, with 15 mm and 93 mm of precipitation at high elevations, respectively, and generally varying 112 mm between the driest and wettest month in the entire region. In January, the average temperature is 1.5 °C and July is the hottest month with an average temperature of 24.1 °C (Ali et al. 2014). Due to variable topography and climate, much of the area is covered by shrubland (Dodonea viscosa, Indigofera gerardiana and Parrotiopsis jacquemontiana) or broadleaved evergreen (Quercus baloot, Q. Incana, Olea ferruginea) and broadleaved deciduous (Acacia modesta, Ficus palmata, Alnus nitida) forests. Introduced Eucalyptus lanceolata and Populous nigra dominate where they occur along roads and rivers (Ilyas et al. 2013). Other vegetation includes, coniferous, meadows, wetlands and ruderal and gullies vegetation communities developed by ice melting.
Sampling and data collection
We sampled vegetation dominated by P. wallichiana in its natural zone of occurrence in both moist and dry temperate forests during the peak growing vegetation period in 2014–2016. Thirty forest stands were sampled between 1200 and 3000 m a.s.l., at sites with varied topographic and soil characters, and starting around the transition zones of subtropical pine forests (P. roxburghii) in the region. At each stand, we sampled 10 plots measuring 10 × 10 m for a total of 300 plots. We also sampled a 200 m straight transect for overstory and 5 × 5 m for understory (Uprety et al. 2014). We attempted to identify each tree species with the help of a senior taxonomist in the field and we recorded frequency, density, diameter, and height. Frequency and density for understory species were also recorded and were used as species abundance data to describe community types. Within each forest stand we recorded elevation, slope gradient, and aspect by use of GPS, clinometer and magnetic compass. Three topsoil samples of 1 kg were taken at three different positions to a depth of 15 cm (or less, if the surface soil horizon was shallow) using a soil bucket agar. The three samples were stored in polyethylene bags and bulked into one composite sample. The three soil sampling spots were taken from the edge of the study plots so as not to disturb plot vegetation as recommended by Woch et al. (2016).
pH values were measured in situ directly by making a suspension of distilled water and soil (1:5 ratio) in the field (pH meter PH119) following Khan et al. (2014). All the soil samples were brought to the Agriculture Research Station (ARS) Mingora North and were sieved (2 mm mesh) and dried at 105 °C prior to physiochemical analysis. Soil texture properties, viz. sand, silt, and clay, were quantified by a combination of sieving and sedimentation. Soil organic matter (SOM) content was determined by the Potassium dichromate (K2Cr2O7) method while a modified semi-micro Kjeldahl procedure was adopted for estimation of total nitrogen (Lu 2000). Micro diffusion and Olsen methods were used to determine the available nitrogen and phosphorus (AP) respectively, whereas, available potassium (AK) was estimated following the method of Bray and Kurtz (1945).
Data analysis
Frequency, density and basal area for different tree species were calculated and sorted according to Mueller-Dombois and Ellenberg (1974). Relative values were calculated by dividing individual values for frequency, density and basal area by the sum of frequencies, densities and basal areas of all species in a plot, then multiplying by 100. Importance value index (IVI) for each species was calculated as the sum of the relative frequency, density and basal area values (Cottam and Curtis 1956). We used importance values in the analysis as it discloses the ecological importance of a species and gives more information than any other single ecological attribute (Brown and Curtis 1952). Data matrices of tree species IVI and the corresponding environmental variables were prepared in Microsoft Excel and were used in the quantitative multivariate analyses using PC-ORD software version 5.10 (McCune and Mefford 2005). In the assignment of slope and aspect, 4 and 8 grades were set and assigned with figures respectively as suggested by Khan et al. (2013) for better interpretation. We used Ward’s Agglomerative cluster analysis (WACA) for objective classification of forest vegetation (McCune and Grace 2002). We used the quantitative Sorensen (Bray–Curtis) distance measure, an effective distance measure for ecological community analysis (McCune and Grace 2002) and flexible beta (β = − 0.25) as a linkage method (Lance and Williams 1967) which is often compatible with Sorensen distance and is space-conserving (Legendre and Legendre 1998). Community type was declared based on the first two species with highest average importance values in the groups following Chai and Wang (2016). The phytosociological parameters of vegetation and associated environmental variables in groups identified by cluster analysis were subjected to one-way analysis of variance (ANOVA) to compare means. The significant differences (p < 0.05) among different parameters in vegetation groups (community types) and habitat properties were further examined using a Tukey’s HSD post hoc test for multiple comparisons using SPSS 18.0. The variability of vegetation parameters and topographic variables between groups were tested with Box–Whisker diagram following Kavgaci et al. (2013). Non-metric Multinational Scaling (NMS) ordination was applied to identify important ecological gradients determining plant species distribution and to verify the classification of the forest vegetation as the two results are often complementary (Atkinson and Treitz 2012; Khan et al. 2014). We preferred the use of NMS-ordination as it is a superior ordination method because (1) it is not based on the assumption of multivariate normality, and (2) it is robust to problems caused by large numbers of zero truncation (Minchin 1987). We employed the low stress-value in the analysis as lower values of stress indicate a more reliable ordination (Benítez-Malvido et al. 2014). Intra- and inter-set correlations among ordination scores and environmental factors were performed with Pearson’s r (Peterson and McCune 2001).
Results
Classification of vegetation
We recorded 105 plant species of 90 genera and 50 families from both overstory and understory in the 300 sampling plots. Dicotyledonous angiosperms were the most abundant category with 90 species (Table 1). Of these species, Gymnospermae included 5 species of 2 families, and Pteridophyta included 3 species of 3 families. The plant species in this region mainly represented 50 families, including Asteraceae and Rosaceae which were rich species families with twelve species each. Lamiaceae was the second most rich family with 9 species, whereas, the remaining families had 4 or less species each.
Our 30 forest stands were comprised of 12 tree species that we classified into three ecologically distinct vegetation types using Ward’s agglomerative cluster analysis (Fig. 2). Dominant and co-occurring species of these populations types with their importance values (IV) are listed in Table 2, and the corresponding topographic and soil variables are listed in Table 3.

Classification of 30 sample stands dominated by Pinus wallichiana into three groups (vegetation types) using Ward’s Agglomerative clustering procedure. Groups were extracted at 70% information and arranged from top (Group-I) to bottom (Group-III) on the Dendrogram
The first division of the clustering dendrogram generated a group of 11 forest stands dominated by P. wallichiana (IV = 76 ± 2.7%) and Quercus dilatata (IV = 6.4 ± 2.5%). This group was both floristically and environmentally heterogeneous and consisted of 12 trees and 66 understory species. P. smithiana, A. pindrow, and Q. incana were co-occurring species with importance values ranging from 4.3 to 3.1%. T. fauna, C. deodara, A. indica, J. regia, P. cornuta, P. pashia and P. duthiei contributed < 5% of importance value in this group (Table 2). The understory flora included 66 species, of which Fragaria nubicola was abundant, while Dryopteris stewartii was a frequently occurring species. Rabdosia rugosa, Viola canescens, Viburnum grandiflorum, Indigofera heterantha, Rubia cordifolia, Arisaema flavum, Bistorta amplexicaulis and Berberis lyceum were in the frequency range from 20 to 40%. The remaining 56 species were rare with average frequency ranging from 0.4 to 20%.
The second group generated a homogenous cluster and we considered this to be a monospecific community of P. wallichiana (IV = 95%). Each co-dominant species contributed < 2% of the importance value (Table 2). This group constituted a single cohort stand structure and generally formed mature stands in the region. In comparison to groups 1 and 3, this group was floristically rich in terms of understory species (80 species). Fragaria nubicola (70%) and Dryopteris stewertii (52%) were frequent species in the understory. The small number of species was occasional and occurred at a frequency ranging from 22% to 38%, whereas 69 species recorded in this group occurred rarely.
The third group was dominated by P. wallichiana (IV = 52.8%) and C. deodara (IV = 24.5%) whereas, P. smithiana, Q. dilatata and A. pindrow locally join the population with importance values ranging from 6.3% to 8.7%. This community was represented by mature stands of a multi-cohort structure. The community was relatively poor with 28 understory species, of which F. nubicola (69%) was abundant and Rumex dentatus (41%) was frequent. V. canescens (29%) and I. heterantha (24%) were occasional species. Remaining 24 species were rare with average frequency ranging from 0.4 to 20%.
Environmental characteristics of the vegetation
The environmental variables topographic, edaphic and soil variables varied widely (Table 3). Forest stands mainly were located in the middle of the forest vegetation belt (2144–2361 m a.s.l.) with moderate (26°) to steep (32°) slope inclination and south to east aspect. Clay and sand particles, and pH progressively increased with increasing elevation. However, silt, organic matter and inorganic ions of soil exhibited a reverse trend. Vegetation group I occurred at low elevation on steep slopes with fertile soils containing high concentrations of N, P, K, and organic matter. The soil type was loamy and acidic.
Aspect varied greatly among stands of group I, i.e., north, north-west, west, north-east, and south-east. Group II stands were mostly located at mid elevation (average 2257 m), with steep slopes (32°) and south, east, and west aspects. Among soil nutrients, organic matter, N, P, and K+ contents were medium, while the concentration of lime was high. The soil type was loamy-sand with medium pH. Stands of group III were located at high elevation (2361 m) with comparatively gentle slopes (26°) on north-west aspects. In comparison to groups I and II, organic matter and the concentration of essential nutrients (N, P, and K+) were lower with slightly acidic soils.
Univariate analysis of variance (ANOVA)
Environmental variables corresponding to the three identified vegetation groups were analyzed using one-way ANOVA (Table 4). Among the topographic variables, slope gradient differed significantly (Tukey HSD test, F = 10.89; p < 0.05) by group but elevation and slope aspect did not. Edaphic variables organic matter (%), soil nitrogen (%), lime (%) and phosphorus (mg kg−1) did not vary by group but soil pH did.
Correlation among the environmental variables
The intra-set correlation among different environmental factors exhibited significant relationships on various probability levels (Table 4). Among the 12 variables, elevation showed a negative relationship (r = − 0.419; p < 0.05) with slope whereas; aspect, lime and potassium contents were positively correlated. pH showed a positive relationship with aspect and clay content of the soil. Strong negative relationship of silt was also observed with sand, pH, and lime (%) respectively.
The sand particles showed a strong negative relationship with clay (r = − 0.553; p < 0.05) and silt particles (r = − 0.901; p < 0.05). This relationship was also observed in phosphorus contents with sand contents and soil pH. Lime, nitrogen and potassium content showed a positive correlation with soil organic matter, while nitrogen content of the soil was positively correlated with potassium.
Ordination and gradient analysis
To detect the significance of individual environmental factors, Non-metric multidimensional scaling produced a 2-dimensional solution (Fig. 3) with a stress value of 11.70 (p < 0.01) based on 250 iterations with randomized data and a final instability of 0.01019. The ordination explained 79% of the variance in the original dissimilarity matrix, with axes 1 and 2 accounting for 51% and 28% of the total. In the NMS-ordination, three groups identified by cluster analysis were superimposed into the graph configuration quite neatly and could be distinguished along both axes 1 and 2. P. wallichiana–Q. dilatata (Group-I) dominated forests appears at the upper middle part of the axes and P. wallichiana (Group-II) pure population forest, located at the extreme top of the 2-D NMS-ordination along axis 1 and 2. P. wallichiana–C. deodara (Group-III) dominated forests plot on the lower middle part of the graph configuration.

Non-metric multidirectional scaling ordination (NMS) showing the pattern of the 30 stands dominated by P. wallichiana along axes 1 and 2. Groups i.e., Pinus–Quercus; Pinus (pure community) and Pinus–Cedrus community obtained from cluster analyses were superimposed in the ordination space
In the NMS ordination diagram of the forest stand dominated by P. wallichiana, a clear gradient of the floristic variations in the communities were evident along axes 1 and 2 (Fig. 3). The ordination axes 1 and 2 is positively correlated with the elevation (r = 0.3778; p < 0.05), however, slope (r = 0.5548; p < 0.01) and pH (r = 0.49750; p < 0.01) were correlated strongly with ordination axis 2 only (Table 5). Aspect exhibited a significant relation with both the axes whereas, significant relationships of soil clay (r = 0.2782; p < 0.05) and silt (r = 0.32251; p < 0.05) contents with axes 2 were also observed (Table 5).
The soil variables organic matter, nitrogen, phosphorus, and potassium did not show any significant relationships. However, among the edaphic factors, pH and lime (%) affected vegetation in the study area (Table 5).
Discussion
The quantitative clustering and ordination approach in the present study clearly describe the distribution pattern of the tree vegetation composition. Although the vegetation types were dominated by P. wallichiana, they showed substantial structural and floristic heterogeneity and complex relationship with the local environment. The three community types were dominated by P. wallichiana and were distributed at elevations from 2144 to 2361 m a.s.l., a middle elevation in the study area. The species rich Pinus–Quercus community grew best at low elevations, whereas, the pure population was located at medium elevation (2257 m) on steep slopes (32°). The second species rich Pinus–Cedrus community occurred at high elevation (2361 m) and on moderate (26°) slopes. Many studies have reported similar results in classifying the coniferous forests in Himalayan, Hindukush and Karakoram ranges of northern Pakistan. Akbar et al. (2017) described the coniferous forests of Gilgit-Baltistan and concluded that P. wallichiana dominated pure and mixed forest stands at low and mid elevation zones on medium to steep slopes (26°–33°). Ahmed et al. (2010) studied P. wallichiana from different climatic zones of Pakistan at elevations from 1950 to 2700 m and slopes of 23°–45° gradient. They concluded that Betula utilis and P. wallichiana prefer moist areas of dry temperate zones. Ahmed et al. (2006) recorded P. wallichiana in pure stands at elevations of 2770 m and 3100 m on south exposures in Naltar and Takht-e-Suleiman area of Baluchistan Province.
The results of cluster analysis have generally suggested a vertical zonal distribution of P. wallichiana dominated vegetation types in the Swat Hindukush range mountains. However, the vertical zonal spectrum is incomplete due to the occurrence of sampling forests only in middle elevation range. Forest area in this region is comparatively small and mainly composed of coniferous mixed forests (Champion et al. 1965; Ahmed et al. 2006).
The ordination results were complementary to classification and ecologically sound as the floristic variations in communities dominated by P. wallichiana showed a clear gradient of environmental variables along the axes 1 and 2 (Fig. 2). Among the environmental variables elevation (r = 0.3778; p < 0.05) and aspect showed significant positive correlation with both of the axis, whereas, slope showed statistical strong correlation (r = 0.5548; p < 0.01) with axis 2 (Table 5). These results indicated that topographical variables have strong influence on vegetation, because the change in physiography leads to change in temperature and humidity gradient (Zhang et al. 2006). Several authors have discussed the prominent role of elevation, aspect and slope in vegetation development and distribution in mountains (e.g. Fisher and Fuel 2004; Kabrick et al. 2004; Jonathan and Givnish 2007; Mahdavi et al. 2013; Khan et al. 2014), and therefore, it is significant in vegetation restoration in such microhabitats.
The results of NMS-ordination also suggest that soil textural and chemical properties i.e. clay (r = 0.278), silt (r = 0.323) and pH (r = 0.49750) have significantly positive correlations with ecological groups of the species (Table 5). Our findings are in line with several studies (e.g. Badano et al. 2005; Small and McCarthy 2005; Mataji et al. 2009), that soil texture also greatly influences the formation of plant communities. Additionally, pH (r = 0.4975) and lime (r = 0.398) are important in defining the ecological groups and species richness in the present study as it plays an important role in regulating photosynthesis, and other physiological processes (Xu et al. 2006; Zhang et al. 2012; Sekulova et al. 2013). The availability of essential nutrients like potassium, phosphorus, organic matter and nitrogen etc., remained inactive in the floristic variation of the P. wallichiana dominated forest of the study area. Although the role of these factors in the distribution of plant species is described by several researchers in their studies (e.g., Enright et al. 2005; Zare Chahoki et al. 2007) as these factors make easy the transformation of water and nutrients and thus increase soil fertility (e.g. Biggelow and Canham 2002; Amorin and Batalha 2007).
Conclusions
Three different tree communities dominated by P. wallichiana were identified and elevation-coupled soil textural and physiochemical properties were strongly correlated with vegetation distribution and must be considered if the goal is the restoration and conservation of these forests. This study provided a better understanding of the mechanisms driving vegetation distribution in the similar or other vegetation types at adjoining high mountain ranges in Pakistan, India, and Afghanistan. We recommend a more comprehensive study at both local and regional scales to expose the vegetation–environment relationships of these forests by considering several other biotic and abiotic factors for better understanding.
References
Ahmed M, Husain T, Sheikh AH, Hussain SS, Siddiqui MF (2006) Phytosociology and structure of Himalayan Forests from different climatic zones of Pakistan. Pak J Bot 38:361–383
Ahmed M, Nazim K, Siddiqui MF, Wahab M, Khan N, Khan MU, Hussain SS (2010) Community description of Deodar forests from Himalayan range of Pakistan. Pak J Bot 42(5):3091–3102
Ahmed M, Shaukat SS, Siddiqui MF (2011) A multivariate analysis of the vegetation of Cedrus deodara (Roxb.) G. Don f. forests in Hindukush and Himalayan ranges of Pakistan: evaluating the structure and dynamics. Turk J Bot 35:419–438
Akbar M, Ahmed M, Hussain F, Siddiqui MF, Raza G, Hyder S, Abbas N (2017) Relationship of forest vegetation and environmental gradients (Edaphic, topographic and soil nutrients) from some frosted areas of Himalayan, Hindu Kush and Karakoram ranges of Gilgit baltsitan, Pakistan—(a multivariate approach). Pak J Bot 49(SI):255–272
Akhtar N, Rashid A, Murad W, Bergmeier E (2013) Diversity and use of ethno-medicinal plants in the region of Swat, North Pakistan. J Ethnobiol Ethnomed 9(25):1–13
Ali K (2011) The conservation issues of medicinal plants of the Swat valley, with special reference to the tree flora. PhD dissertation, School of Biological Sciences, University of Reading, UK
Ali K, Ahmad H, Khan N, Jury S (2014) Future of Abies pindrow in Swat district, northern Pakistan. J For Res 25:211–214
Ali K, Khan N, Rahman IU, Ahmad H, Jury S (2015) Multivariate analysis and vegetation mapping of a biodiversity hotspot in the Hindu Kush Mountains. Int J Adv Res 3(6):990–1006
Amorin PK, Batalha MA (2007) Soil vegetation relationship in hyper seasonal cerrado and wet grassland in Emas National Park (Central Brazil). Acta Oecol 32:319–327
Atkinson DM, Treitz P (2012) Arctic ecological classifications derived from vegetation community and satellite spectral data. Remote Sens 4:3948–3971
Badano EI, Cavieres LA, Molinga-Montenegro MA, Quiroz CL (2005) Slope aspect influences association pattern in the Mediterranean natural of central Chile. J Arid Environ 62:93–108
Baig MB, Ahmad S, Khan N, Ahmad I, Straquadine GS (2008) The history of social forestry in Pakistan: an overview. Int J Soc For 1(2):167–183
Barbour MG, Burk JH, Pitts WD (1987) Terrestrial plant ecology. The Benjamin/Cummings Publishing Company, Inc, New York
Beg AR, Khan MH (1984) Some more plant communities and the future of dry oak forest zone in Swat valley. Pak J For 34(1):25–35
Benítez-Malvido J, Gallardo-Vásquez JC, Alvarez-añorve MY, Avila-Cabadilla LD (2014) Influence of matrix type on tree community assemblages along tropical dry forest edges. Am J Bot 101(5):820–829
Biggelow SW, Canham CD (2002) Community organization of tree species along soil gradients in a north-eastern USA forests. J Ecol 90(1):188–200
Bray RH, Kurtz LT (1945) Determination of total, organic and available forms of phosphorus in soils. Soil Sci 59:39–45
Brown RJ, Curtis JJ (1952) The upland conifer–hardwood communities of southern Wisconsin. Ecol Monogr 22:217–234
Burke A (2001) Classification and ordination of plant communities of the Naukluft Mountains, Namibia. J Veg Sci 12(1):53–60
Chai Z, Wang D (2016) A comparison of species composition and community assemblage of secondary forests between the birch and pine–oak belts in the mid-altitude zone of the Qinling Mountains, China. Peer J 4(1–2):e1900. https://doi.org/10.7717/peSerj
Champion H, Seth SK, Khattak GM (1965) Forest types of Pakistan. Pakistan Forest Research Institute, Peshawar, Bulletin No. 7
Chapin FS III, Sala OE, Burke IC, Grime JP, Hooper DU, Lauenroth WK, Lombard A, Mooney HA, Mosier AR, Naeem S, Pacala SW, Roy J, Steffen WL, Tilman D (1998) Ecosystem consequences of changing biodiversity. Bioscience 1:45–52
Cottam G, Curtis JT (1956) The use of distance measures in phytosociological sampling. Ecology 37(3):451–460
Enright NJ, Miller BP, Akhter R (2005) Desert vegetation and vegetation–environment relationships in Kirthar National Park, Sindh, Pakistan. J Arid Environ 61:397–418
Farjon A (2013) Pinus wallichiana var. wallichiana. The IUCN red list of threatened species 2013: T191650A1991477. http://dx.doi.org/10.2305/IUCN.UK.2013-1.RLTS.T191650A1991477.en. Downloaded on 23 Oct (2015)
Fisher MA, Fuel PZ (2004) Changes in forest vegetation and arbuscular mycorrhizae along a steep elevation gradient in Arizona. For Ecol Manag 200:293–311
Hussain F, Illahi I (1991) Ecology and vegetation of lesser Himalayan Pakistan. Botany Department, University of Peshawar, Khyber Pakhtunkhwa, p 187
Hussain F, Ilyas M, Takatsuki S (1997) Plant communities of Girbanr hills, Swat district, Northwestern Pakistan. Ecol Rev 23(4):247–260
Ilyas M, Shinwari ZK, Qureshi R (2012) Vegetation composition and threats to the montane temperate forest ecosystem of Qalagai Hills, Swat, Khyber Pakhtunkhwa, Pakistan. Pak J Bot 44:113–122
Ilyas M, Qureshi R, Arshad M, Mirza SN (2013) A preliminary checklist of the vascular flora of Kabal Valley, Swat, Pakistan. Pak J Bot 45(2):605–615
Ilyas M, Qureshi R, Akhtar N, Munir M, Zia-Ul-Haq (2015) Vegetation analysis of Kabal valley, district Swat, Pakistan using multivariate approach. Pak J Bot 47(SI):77–86
Jonathan CD, Givnish TJ (2007) Gradient analysis of reversed tree lines and grasslands of the Valles Caldera, New Mexico. J Veg Sci 18:43–54
Kabrick JM, Shifley SR, Jensen RG, Larsen DR, Grabner JK (2004) Oak forest composition, site quality, and dynamics in relation to site factors in the southeastern Missouri Ozarks. Gen Tech Rep SRS-73. U.S. Department of Agriculture. Forest Service, Southern Res Sta, pp 94–101
Kavgaci A, Sevgi O, Tecimen HB, Yilmaz OY, Carus S, Dündar T (2013) Classification and ordination of Pinus nigra dominated forests at Alacam mountains (NW Anatolia-Turkey). Eur J For Sci 1(1):38–50
Khan N, Shaukat SS, Ahmed M, Siddiqui FM (2013) Vegetation–environment relationships in the forests of Chitral District Hindukush range of Pakistan. J For Res 24(2):205–216
Khan N, Ali K, Shaukat SS (2014) Phytosociology, structure and dynamics of Pinus roxburghii associations from Northern Pakistan. J For Res 25(3):511–521
Lance GN, Williams WT (1967) A general theory of classification sorting strategies. II. 525 Clustering strategies. Comput J 10:271–277
Legendre P, Legendre L (1998) Numerical ecology, 2nd edn. Elsevier, Amsterdam, p 853
Lu RK (2000) Method of soil agricultural chemistry analysis. Chinese Agricultural Science and Technology Press, Beijing, pp 362–370
Ma M, Zhou X, Ma Z, Du G (2012) Composition of the soil seed bank and vegetation changes after wetland drying and soil salinization on the Tibetan Plateau. Ecol Eng 44:18–24
Mahdavi P, Akhani H, Van der Maarel E (2013) Species diversity and life-form pattern in steppe vegetation along a 300 m altitudinal gradient in the Alborz Mountains. Iran Folia Geobotanica 48:7–22
Mataji M, Amiri Z, Gh Asri Y (2009) Vegetation analysis based on plant associations and soil properties in natural forests. Iran J For Pop Res 17(1):85–98
McCune BJ, Grace B (2002) Analysis of ecological communities. MjM Software, Gleneden Beach. ISBN 0-9721290-0-6
McCune B, Mefford MJ (2005) Multivariate analysis of ecological data (PCORD Version 5.10 Mj-M Software) Gleneden Beach, United State of America, Oregon
Minchin PR (1987) An evaluation of the relative robustness of techniques for ecological ordination. Vegetatio 69:89–107
Mucina L (1997) Conspectus of classes of European vegetation. Folia Geobot 32:117–172
Mueller-Dombois D, Ellenberg H (1974) Aims and methods of vegetation ecology. Wiley, New York, p 547
Peterson EB, McCune B (2001) Diversity and succession of epiphytic macro lichen communities in low-elevation managed conifer forests in western Oregon. J Veg Sci 12:511–524
Qasim M, Hubacek K, Termansen M, Fleskens L (2012) Modelling land use change across elevation gradients in district Swat, Pakistan. Reg Environ Change 12(4):567–581
Rahman MU (2012) Wild plants of swat Pakistan. Department of Botany, GJPGC Saidu sharif Swat. Alam graphics and printers, Matta swat
Rahman IU, Khan N, Ali K (2017) Classification and ordination of understory vegetation using multivariate techniques in the Pinus wallichiana forests of Swat Valley, northern Pakistan. Sci Nat 104(3–4):24
Ren GH, Deng B, Shang ZH, Hou BY, Long RJ (2013) Plant communities and soil variations along a successional gradient in an alpine wetland on the Qinghai-Tibetan Plateau. Ecol Eng 61:110–116
Sekulova L, Hajek M, Vıt Syrovatka (2013) Vegetation–environment relationships in alpine mires of the West Carpathians and the Alps. J Veg Sci 24:1118–1128
Shaheen H, Khan SM, Harper DM, Ullah Z, Quresh RA (2011) Species diversity, community structure, and distribution patterns in Western Himalayan Alpine Pastures of Kashmir, Pakistan. Mt Res Dev 31(2):153–159
Sheikh MI (1993) Trees of Pakistan. Pictorial Printing (Pvt) Ltd, Islamabad
Sher H, Alyemeni MN, Faridullah (2010) Cultivation and domestication study of high value medicinal plant species (its economic potential and linkages with commercialization). Afr J Agric Res 5:2462–2470
Shinwari ZK, Khan AS, Nakaike T (2003) Medicinal and other useful plants of District Swat. Al Aziz Communications, Peshawar. ISBN 969-8283-21-8
Siddiqui MF, Ahmed M, Khan N, Hussain SS, Khan IA (2010) A quantitative description of moist temperate conifer forests of Himalayan region of Pakistan and Azad Kashmir. Int J Biotechnol 7:175–185
Siddiqui MF, Arsalan A, Ahmed M, Hussain MI, Iqbal J, Wahab M (2016) Assessment of understorey vegetation of Malam Jabba Forest, Kpk after cleanup operation using multivariate techniques. Jurnal Teknologi 78:9–17
Small CJ, McCarthy BC (2005) Relationship of understory diversity to soil nitrogen, topographic variation, and stand age in an eastern oak forest, USA. For Ecol Manag 217(2):229–243
Stewart RR (1967) Check list of the plants of south Northwest Pakistan. Pak J For 45:457–528
Uprety Y, Asselin H, Bergeron Y, Mazerolle MJ (2014) White pine (Pinus strobus L.) regeneration dynamics at the species natural limits of continuous distribution. New For 45(131):1–18. https://doi.org/10.1007/s11056-013-9396-2
Wahab M (2011) Population dynamics and dendrochronological potential of pine tree species from districts Dir. PhD thesis, Federal Urdu university of Arts, Sciences and Technology, Karachi
Woch MW, Kapusta P, Stefanowicz AM (2016) Variation in dry grassland communities along a heavy metals gradient. Ecotoxicology 25:80–90
Xu L, Liu H, Chu X, Su K (2006) Desert vegetation patterns at the northern foot of Tianshan Mountains: the role of soil conditions. Flora 201(1):44–50
Yadav RR, Park WK, Battacharyyar A (1997) Climate and Growth relationship in blue Pine (Pinus wallichiana) from the western Himalaya India. Korean J Ecol 2:95–102
Zare Chahoki MA, Jafari M, Azarnivand H (2007) Relationships between species diversity and environmental factors of Poshtkouh rangelands in Yazd. Iran J Paj Saz 21(1):192–199
Zeb A (1970) The geology of the Shah Deri Kabal area, Swat. J Himal Earth Sci 5:96–111
Zhang JT, Zhang F (2007) Diversity and composition of plant functional groups in mountain forests of the Lishan Nature Reserve, North China. Bot Stud 48:339–348
Zhang XP, Wang MB, Xiao Y (2006) Quantitative classification and ordination of forest communities in Pangquangou National Nature Reserve. Acta Ecol Sin 26(3):754–761
Zhang HY, Qian YB, Wu ZN, Wang ZC (2012) Vegetation–environment relationships between northern slope of Karlik Mountain and Naomaohu Basin, East Tianshan Mountains. Chin Geogr Sci 22(3):288–301
Acknowledgements
The authors are thankful to the Director of Agriculture Research Station (ARS) Mingora North for providing laboratory facility for soil analysis. We are also grateful to Higher Education Commission of Pakistan for funding the research project.
Author information
Authors and Affiliations
Corresponding author
Additional information
Project funding: This study was supported by Higher Education Commission of Pakistan.
The online version is available at http://www.springerlink.com
Corresponding editor: Hu Yanbo.
Rights and permissions
About this article

Cite this article
Rahman, I.U., Khan, N., Ali, K. et al. Vegetation–environment relationship in Pinus wallichiana forests of the Swat Hindukush range of Pakistan. J. For. Res. 31, 185–195 (2020). https://doi.org/10.1007/s11676-018-0864-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-018-0864-6