
Abstract
Understanding forest ecosystem evapotranspiration (ET) is crucial for water-limited environments, particularly those that lack adequate quantified data such as the lower Heihe River basin of northwest China which is primarily dominated by Tamarix ramosissima Ledeb. and Populus euphratica Oliv. forests. Accordingly, we selected the growing season for 2 years (2012 and 2014) of two such forests under similar meteorological conditions to compare ET using the eddy covariance (EC) technique. During the growing seasons, daily ET of T. ramosissima ranged from 0.3 to 8.0 mm day−1 with a mean of 3.6 mm day−1, and daily ET of P. euphratica ranged from 0.9 to 7.9 mm day−1 with a mean of 4.6 mm day−1 for a total of 548 and 707 mm, respectively. The significantly higher ET of the P. euphratica stand was directly linked to high soil evaporation rates under sufficient water availability from irrigation. When the soil evaporation was disregarded, water use was comparable to two contrasting riparian forests, a P. euphratica forest with a total transpiration of 465 mm and a T. ramosissima forest with 473 mm. Regression analysis demonstrated that climate factors accounted for at least 80% of ET variation in both forest types. In conclusion, water use of the riparian forests was low and comparable in this arid region, that suggest the long-term plant adaptation to the local climate and conditions of water availability.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Evapotranspiration (ET) is a major component of catchment water balances. Because ET in most large basin ecosystems is limited by rainfall, riparian corridors are hot spots for atmospheric water loss for large arid- and semi-arid basins (Sala et al. 1996; Smith et al. 1998). In the lower Heihe River basin (HRB), northwestern China, riparian forests are dominated by phreatophytes, such as Populus euphratica Oliv. tree species and Tamarix ramosissima Ledeb. shrub species (Si et al. 2007). Phreatophytes are deep-rooted plants that obtain their water from the water table or the soil above it. Over the past several decades, the eco-environment of riparian forests has markedly deteriorated to the extent that riparian forests have decreased in size and their ecological functions have declined (Cheng et al. 2014; Feng and Cheng 1998; Si et al. 2007). In order to restore riparian forests in the lower HRB, the Ecological Water Conveyance Project (EWCP) was initiated in 2000 by the government of China to recharge water back into the lower HRB (Guo et al. 2008). Although P. euphratica forests were maintained after this water conveyance program was put into practice (from 295.69 km2 in 1982 to 294.30 km2 in 2010), T. ramosissima forests have expanded in area (from 1003.71 km2 in 1982 to 1529.56 km2 in 2010). Accordingly, it is important to quantify water consumption by these two forest types in order to effectively utilize their limited water resources.
Although P. euphratica is an important structural tree species of riparian forests, only a limited number of ET studies have been conducted in this region. Studies mainly focused on a tree-level scale (Si et al. 2007; Zhang et al. 2006) and rarely on a stand-level scale (Hou et al. 2010). Annual transpiration measured through sap flow was 214.9 mm (Si et al. 2007), less than the total ET (447.0 mm) estimated by the Bowen ratio energy balance (BREB) method at the same site (Hou et al. 2010). In addition, T. ramosissima is a native phreatophytic that originates from arid regions in Central Asia, and has widely invaded numerous western states (Di Tomaso 1998; Hatler and Hart 2009; Shafroth et al. 2005). Generally, T. ramosissima is recognized as a prolific water user. Data taken from flux towers (Cleverly et al. 2002, 2006; Devitt et al. 1998) and large area estimates using remote sensing techniques (Bawazir et al. 2009; Dennison et al. 2009; Nagler et al. 2009) have yielded moderate water use rates (from 800 to 1450 mm year−1). In contrast to the western U.S., ET from T. ramosissima stands was approximately 500 mm year−1 in the lower Tarim River basin (Yuan et al. 2014) and only 280 mm year−1 in the lower HRB in northwestern China (Si et al. 2005). It may therefore be deduced that ET from T. ramosissima stands is lower in its native regions than in regions it has invaded, but it remains unclear whether T. ramosissima stands have higher ET rates than other native species (e.g., P. euphratica).
In most previously published studies, the eddy covariance (EC) method has been widely used to estimate ET using direct measurements of latent heat fluxes (Sun et al. 2008). The objectives of this study were to determine the magnitude of ET for P. euphratica and T. ramosissima stands in the lower HRB, and compared them to other regions around the world. For this purpose, we gathered nearly 2 years of EC observations from two stands in the lower HRB. In addition, we concurrently observed meteorological variables and groundwater tables in the same plot used to determine environmental controls of ET.
Materials and methods
Study site
The climate in this region is extremely arid with a mean annual precipitation of only 37.4 mm, of which > 75% occurs between June and August. Annual pan evaporation is 3390.3 mm, which is greater than precipitation by a factor of 90. Mean annual temperatures in July and January are 27.0 and − 11.7 °C, respectively, as recorded at the Ejin meteorological station between 1959 and 2011 (Yu et al. 2013b). Mean values ± standard deviation (SD) of meteorological conditions for 2012 and 2014 are provided in Table 1. Given that both the highest rainfalls and temperatures occur concurrently in this region, most rainfall is depleted by evaporation before it infiltrates into the soil. Therefore, the effect of rainfall on the water balance is minimal. In addition to rainfall, groundwater recharged by runoff from the Heihe River sustains both local residents and regional ecosystems (Si et al. 2014).
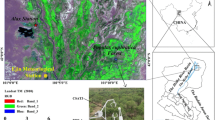
We selected T. ramosissima and P. euphratica stands, two typical forest types in arid regions, to conduct EC measurements. The T. ramosissima stand (42°01′N, 101°03′E; 926 m a.s.l.) is situated in a north–south direction in the Alxa Desert Ecohydrology Experimental Research Station, Chinese Academy of Sciences. The P. euphratica stand (41°59′N, 101°10′E; 920 m AMSL) is located at the P. euphratica Forest National Nature Refuge at Qidaoqiao (Fig. 1). We initially conducted a comprehensive quadrat sampling of vegetation in 2010 for both forest types (Table 2). Leaf area index (LAI) of canopies was measured using a plant canopy analyzer (LAI-2200, LI-COR Biosciences, Inc., Lincoln, NE, USA) with one wand for both above- and below-canopy measurements under diffused light conditions during sunrise and sunset. This was carried out approximately once every month throughout the growing season in both years. The understory was composed of shallow-rooted grasses, predominantly Sophora alopecuroides L., Karelinia caspica (Pall.) Less., and Achnatherum splendens (Trin.) Nevski. Soil characteristics from the T. ramosissima (Yu et al. 2013a) and P. euphratica stands were determined (Gao et al. 2016b).

Map of the Heihe River basin and the locations of the T. ramosissima and P. euphratica stands
Flux and meteorological measurements
Momentum flux, sensible heat (H, W m−2), and latent heat (LE, W m−2) were measured using a three-dimensional sonic anemometer (CSAT3, Campbell Scientific, Inc., North Logan, UT, USA) and an open path CO2/H2O analyzer (LI-7500A, LI-COR Biosciences, Inc., USA). In addition, meteorological variables were recorded, including net radiation (R n , W m−2), air temperature (T a , °C), and relative humidity (RH, %) in conjunction with online-processed 0.5 h mean horizontal wind speeds (U, m s−1) and CO2/H2O flux rates using the CR3000 datalogger (Campbell Scientific Inc., USA). R n was determined using a four-component net radiometer (CNR4, Kipp & Zonen, Delft, the Netherlands), while T a and RH were measured using a relative humidity and temperature sensor probe (HMP45C, Campbell Scientific, Inc, USA). The vapor pressure deficit (VPD, kPa) was calculated based on T a and RH (Campbell and Norman 1998). Soil heat flux (G, W m−2) was measured using a heat flux plate (HFP01, Hukseflux Thermal Sensors B.V., Delft, the Netherlands) at a 5 cm depth dispersed 0.5 m belowground horizontally. We conducted T. ramosissima stand flux and meteorological measurements from May 2011 to November 2012 at 3 m above-ground level. Subsequently, this method was applied to a 20 m high flux tower to measure P. euphratica stand flux and meteorological measurements from June 2013 to the present (Gao et al. 2016b). To compare, we selected the entire 2012 and 2014 growing seasons (i.e., from May to September) under similar meteorological conditions for analysis. In addition, two wells 30 m apart from the EC tower were installed and groundwater depth (GD, m) measured at 5 day intervals during the growing seasons, which provided results of a maximum of 2.65 m for the T. ramosissima stand and 2.25 m for the P. euphratica stand. Water was introduced via the Heihe River EWCP (set up to provide local herdsmen a means to irrigate their pastures) to the P. euphratica stand after September 16, 2014.
Data processing and gap filling
Raw CSAT3 and CO2/H2O flux data were processed using EddyPro software (LI-COR Biosciences, Inc., USA), which includes features such as double coordinate rotation, block average detrending, time lag compensation via covariance maximization (with defaults), and density fluctuation compensation via the use/convert features to the mixing ratio (Burba et al. 2012). Statistical analysis of the raw time series data was carried out, including spike count/removal, amplitude resolution, dropouts, absolute limits, and skewness and kurtosis, according to Vickers and Mahrt (1997). Quality checks/flagging were conducted according to the CarboEurope standard (Foken et al. 2004). After conducting a detailed postproduction on data, H and LE were calculated to obtain a 0.5 h average. We screened 0.5 h turbulent fluxes and meteorological variables to remove spurious data points caused by sensor malfunctions, rain events, and sensor maintenance.
The energy balance was analyzed from the EC data that include R n, LE, and H and G calculated from the average of the two heat flux plate measurements. We disregarded canopy storage heat and photosynthetic energy consumption due to the sparseness of canopy cover. The relationship between the half-hourly data of net radiation subtracted by the ground heat flux (R n − G) and the half-hourly data of the total turbulent flux (LE + H) indicated that EC underestimated turbulent fluxes by 22% (slope = 0.78; R 2 = 0.91) and 21% (slope = 0.79; R 2 = 0.80) for the T. ramosissima and P. euphratica stands, respectively (Fig. 2). LE data was also removed from the energy unbalance. In total, 34 and 19% of the original LE data was rejected during data quality processing of the T. ramosissima and P. euphratica stands, respectively.

Energy balance closures of eddy covariance (EC) for T. ramosissima (a) and P. euphratica (b) during 2012 and 2014, respectively. The 1:1 line (dotted line) and linear regression (solid line) between Rn − G and LE + H is shown
Gap filling was carried out when total ET had to be calculated by filling shorter gaps (i.e., < 2 h) in the data through linear interpolation, and longer gaps in LE flux using energy balance analysis. If EC measured LE was greater than R n + G − H or null, we then forced LE to close according to the energy balance analysis (Yuan et al. 2014), that is, LE = R n + G − H, otherwise LE was retained. ET (mm h−1) from LE was calculated hourly according to Yuan et al. (2014), the actual ET (mm h−1) from combined data with EC measured ET (ET EC, mm h−1), and the estimated ET from the energy balance analysis (ET EB, mm h−1) (Fig. 3). When hourly ET was negative, it was set to zero. Daily ET (mm day−1) was the sum of hourly ET during a single day. In addition, the daily reference ET (ET 0 , mm day−1) was calculated from the FAO Penman–Monteith equation (Allen et al. 1998) on a daily scale. R n and G data were used to calculate daily ET 0 derived from observations from flux tower measurements, and other daily data from the Ejin meteorological station. The crop coefficient (K c = ET/ET 0) was then determined.

Hourly variation in measured evapotranspiration from eddy covariance (EC) (ET EC, mm h−1), estimated evapotranspiration derived from energy balance analysis (ET EB, mm h−1), and actual evapotranspiration calculated from the combination of the two (ET, mm h−1) for the T. ramosissima stand from June 20 to June 23, 2012
Data analysis
Basic statistical features of mean values and SD for meteorological variables were analyzed and significant differences tested using one-way ANOVA at P = 0.05. Meteorological data collected from the site were used, including R n, T a, RH, U, and VPD, to explain seasonal ET variability by ANOVA as well as correlation and regression analyses provided in the SPSS Statistics 19 software package (IBM, USA). Finally, Origin 8.0 software was used to generate drawings (OriginLab Corp., Northampton, MA, USA).
Results
Comparisons of environmental variables
Limited precipitation, low RH as well as high T a and VPD were the main climatic variables used for the study area. Precipitation was 32.7 mm in 2012 and 17.2 mm in 2014. Two extreme precipitation events, 8.3 and 8.9 mm on June 6 and July 24, respectively, could account for the high precipitation in 2012. At P = 0.05, population means of N, T a, VPD, and U were not significantly different for either study year with the exception of RH from the meteorological station (Table 1).
For the T. ramosissima stand, daily R n increased during the spring and progressed into the summer before progressively decreasing during the autumn from 3.0 to 21.0 MJ m−2 day−1 with an average of 12.4 MJ m−2 day−1. Moreover, G was coincident with R n (Fig. 4a). Daily T a remained positive at an average of 18.5 °C throughout the growing seasons, particularly in August when maximum daily T a reached 31.6 °C. Seasonal variation in RH changed following R n and T a with an average of 31.3% (Fig. 4b) and was particularly low from March to May compared to other months (20.2 vs. 35.9%, P = 0.01), which may be attributable to the high U observed during this period (2.1 vs. 1.4 m s−1, P = 0.05) (Fig. 4c). Mean VPD was 1.8 kPa, and seasonal variation in VPD was closely correlated to R n and T a (Fig. 4c).

Daily variation in meteorological variables, including net radiation (R n, W m−2), soil heat flux (G, W m−2), air temperature (T a, °C), relative humidity (RH, %), vapor pressure deficit (VPD, kPa), and wind speed (U, m s−1) for T. ramosissima (a–c) and P. euphratica (d–f) during 2012 and 2014 growing seasons
For the P. euphratica stand, R n was significantly higher with an average of 14.1 MJ m−2 day−1 compared to the T. ramosissima stand (P = 0.01) during the growing seasons, and G coincided with R n (Fig. 4d). T a and RH, with an average of 19.0 °C and 29.1%, was not significantly different from the T. ramosissima stand (P = 0.50 and P = 0.07, respectively) (Fig. 4e). Undifferentiated T a and RH resulting from VPD was also not significantly different (1.8 kPa, P = 0.75) to the T. ramosissima stand (Fig. 4f). Height (20 m) of measured U results was significantly higher (3.4 vs. 1.6 m s−1, P = 0.01) compared to the T. ramosissima stand.
Comparison of evapotranspiration between the two stands
Energy balance closures (LE + H)/(Rn − G) were 79 and 78% in 2012 and 2014, respectively (Fig. 2). During the dormant season, daily ET was generally less than 0.5 mm day−1. It is noteworthy that ET remained weak throughout March and April but ET 0 remained relatively high (Fig. 5a), from which T a, VPD, and U increased rapidly (Fig. 4b, c). For the growing season, daily ET increased during the spring and progressed into the summer before progressively decreasing in the autumn from 0.3 to 8.0 mm day−1 with an average of 3.6 mm day−1 (Fig. 5a). However, there was pronounced lower ET value in June that was consistent with the lower U for the same period, although it rose sharply afterwards in conjunction with the high VPD in August with a maximum of 7.7 mm day−1. In contrast, ET 0 was high during the late spring and autumn, indicating a low ET to ET 0 ratio (K c: from 0.09 to 0.22) during these periods. In comparison, K c remained high (0.36–0.86) during the summer and early autumn with an average of 0.46 in 2012. This phenomenon indicated that soil evaporation was weak because plant transpiration could not have occurred in March and April. Monthly ET increased from 34.1 mm in May to 180.1 mm in July and decreased to 93.1 mm in September for a total of 548 mm in 2012 (Table 3).

Seasonal patterns in daily reference evapotranspiration (ET 0, mm day−1) and evapotranspiration (ET, mm day−1) for T. ramosissima (a) and P. euphratica (b) in 2012 and 2014. The 5-day average value of ET (black solid line) is shown
For the P. euphratica stand, ET increased earlier and ended later than for the T. ramosissima stand, after which it was consistent with ET 0 (Fig. 5b). Daily ET was less than 1 mm prior to March (T a < 0 °C) during the dormant season (Fig. 5b). However, ET increased rapidly with a peak of 3 mm day−1 after March, at which time T a, VPD and U increased rapidly (Fig. 4e and f), and ET 0 was relatively high (Fig. 5b). During the growing season, daily ET ranged from 0.9 to 7.9 mm with an average of 4.6 mm day−1 (Fig. 5b), and monthly ET increased from 131.3 mm in May to 170.0 mm in July and decreased to 98.5 mm in September for a total of 707 mm in 2014 (Table 3).
Comparison of variables associated with evapotranspiration
For the T. ramosissima stand, daily ET was positive and significantly correlated to R n, T a, and VPD and negative and significantly correlated to RH, but not significantly correlated to U. Due to the high correlations between VPD and RH, the latter was removed during stepwise regression analysis, for which the results revealed that climate variables accounted for at least 81% of ET variation. For the P. euphratica stand, daily ET also positive and significantly correlated to R n, T a, and VPD and negative and significantly correlated to RH (U being the exception), and climate factors, including R n, T a, and VPD, accounted for at least 80% of ET variation (Table 4).
Discussion
Comparison of evapotranspiration between the two stands
Energy balance closures for the two stands (Fig. 2) were comparable with FLUXNET sites (Wilson et al. 2002) as well as with other EC measurements, especially in the arid region of northwestern China (Hao et al. 2007; Yuan et al. 2014). This suggests that direct ET measurements using the EC method were reliable. The total ET of T. ramosissima and P. euphratica during the growing season has been estimated at tree-level (Si et al. 2007; Zhang et al. 2006) and stand-level scales (Hou et al. 2010; Si et al. 2005; Wang et al. 2014). The BREB estimated ET was approximately 248 mm with an average of 1.6 mm day−1 for T. ramosissima (Si et al. 2005) and 447 mm with an average of 3.2 mm day−1 for the P. euphratica stand (Hou et al. 2010). In addition, annual transpiration measured through sap flow was 214.9 mm for P. euphratica (Si et al. 2007). The daily ET rate during the summer months (June–August) was estimated to be 0.63–0.73 mm for T. ramosissima and 1.89–2.33 mm for P. euphratica using the water table fluctuation (WTF) method (Wang et al. 2014). A comparison of results from this study and those of the aforementioned studies indicate that the ET rates estimated using the EC method for both stands were higher than values determined through sap flow, BREB, and WTF methods.
Previous studies (Cleverly et al. 2006; Devitt et al. 2011) have shown that climate factors, vegetation parameters, soil properties, and water table depths are principal factors controlling ET variation in phreatophytic communities. Climate variables, soil types, plant community composition, and water table depth for the two stands were similar for both years (Tables 1, 2). Thus, their different ET values were likely caused by differing soil moisture levels induced by irrigation and further different soil evaporative conditions (Fig. 5). Before 2013, the P. euphratica stand had no access to irrigation and groundwater depth may have been 4 m. After 2013, the irrigation was applied in autumn. Thus, we deduced that the significantly higher ET found in the P. euphratica stand was directly related to the high soil evaporation rate at this site. Reasons for this are discussed below.
Comparison of evapotranspiration between the study sites to other regions
Due to its wide distribution, ET from Tamarix species, an introduced, stress-tolerant shrub from Central Asia, has been extensively investigated throughout the world. The ET results from the T. ramosissima stand using EC were similar to results in the lower Tarim River basin, China, with a mean ET of 500 mm a−1 and an average ET of 3.85 mm day−1 (Yuan et al. 2014). A comparison of results obtained from this study and one in the lower Tarim River (Yuan et al. 2014) showed that T. ramosissima growth in both did not experience water stress. Thus, results from both studies may represent potential ET for native T. ramosissima stands in Central Asia. It is noteworthy that the slightly higher ET found in our study (compared to the results of Yuan et al. 2014) may be attributed to the relatively higher LAI (1.97 vs. 1.15, respectively) and the lower groundwater depth (2.65 vs. 6.62 m, respectively).
However, total ET obtained from EC flux tower measurements (Cleverly et al. 2002, 2006; Devitt et al. 1998; Westenburg et al. 2006) in combination with remote sensing data (Bawazir et al. 2009; Nagler et al. 2005, 2009; Taghvaeian et al. 2014) yielded a broad range from 800 to 1450 mm year−1 in the western United States. It may therefore be concluded that observed ET under arid conditions in native regions was clearly low compared to invasive T. ramosissima stands in non-native regions. These differences could be attributable to two specific reasons. First, soil evaporation in T. ramosissima stands in arid regions could be either weak or negligible (Yuan et al. 2014). Second, although LAI can have a deterministic impact on total ET (Dahm et al. 2002; Sala et al. 1996; Yuan et al. 2014), values from the study and those of Yuan et al. (2014) were lower than those in the middle Rio Grande district (2.5 and 3.5 for flooding and non-flooding, respectively) (Cleverly et al. 2006; Nagler et al. 2005) and the lower Colorado River (2.54) (Nagler et al. 2009), both in the western U.S.
Recent studies have shown that water use by individual T. ramosissima trees is comparable to the native species that it typically replaces such as cottonwood and willow (Glenn and Nagler 2005; Nagler et al. 2003). However, in these cases tree-level soil evaporation had not been taken into account. Although soil evaporation was weak for the T. ramosissima stand as reported by Yuan et al. (2014), this was not the case for the P. euphratica stand due to the high ET observed during the dormant season (Fig. 5b). Based on the simplified Shuttleworth–Wallace (SSW) model, the ratio of canopy transpiration to ET ranged from 0.53 to 0.66 (Gao et al. 2016a). Thus, we estimated that transpiration was 465 mm for the P. euphratica stand which was comparable to 473 mm for the T. ramosissima stand when soil evaporation was disregarded (a total of 77 mm a−1 with an average of 0.5 mm day−1 determined by measuring ET during the dormant season in conjunction with results from Yuan et al. (2014). For greater accuracy, further research is required to partition ET into plant transpiration and soil evaporation.
Factors and variables associated with evapotranspiration
ET rates responded to GD, atmospheric surface layer conditions, stomatal coupling in conjunction with atmospheric humidity levels, energy balances, and LAI (Cleverly et al. 2006). In desert riparian regions, given the limited rainfall, groundwater provides an ample water supply to riparian forests (Si et al. 2014), permitting greater annual ET rates than that provided by rainfall alone (Table 3). Some studies have shown that growth and eco-physiological parameters of riparian forests were less severely affected by GD within a depth of 6 m in arid regions in China (Chen et al. 2006; Feng et al. 2012; Hao et al. 2009). Measured ET of T. ramosissima and P. euphratica stands using the EC method in the lower HRB was comparable to results in the Tarim River basin (Yuan et al. 2014). In contrast, the clear differences in ET in native and invasive regions indicate that climate factors and water availability, which both impact plant eco-physiological characteristics, were the primary controllers of water use for these species.
Regression analysis demonstrate that climate and soil water variables accounting for at least 80% of ET variation in the two riparian forests investigated (Table 4). In addition, plant eco-physiological characteristics, particularly LAI, determined spatial patterns of ET for riparian phreatophytes. Moreover, LAI in combination with plant physiological parameters such as stomatal conductance, would impact seasonal variation in ET (Hatler and Hart 2009). Desert riparian species adapted to arid climates may use of deep soil water and groundwater at the same time. In contrast, greater amounts of rainfall and flooding may supply sufficient available water in shallow soil layers of the arid southwestern U.S. This suggests that plant water use is the result of long-term adaption to local climates and water availability.
Conclusions
During the growing season, ET of the T. ramosissima stand was 548 mm a−1 with a mean of 3.6 mm day−1, similar to that of native forests in Central Asia (500 mm a−1) but clearly lower than that of invasive regions in the western U.S. (from 800 to 1450 mm a −1). ET of the P. euphratica stand was 707 mm with a mean of 4.6 mm day−1, which was directly linked to high soil evaporation rates under sufficient water availability induced by irrigation. When soil evaporation was disregarded, total transpiration was 473 and 465 mm for the P. euphratica and T. ramosissima stands, respectively, indicating comparable water use of the two contrasting riparian forests. Furthermore, regression analysis demonstrated that climate factors accounted for at least 80% of ET variation for both stands. Thus, our results show that plant water use is the result of long-term adaptation to local climates and water availability.
References
Allen R, Pereira LS, Raes D, Smith M (1998) Crop evapotranspiration-guidelines for computing crop requirements. In: FAO irrigation and drainage paper 56. FAO, Rome
Bawazir AS, Samani Z, Bleiweiss M, Skaggs R, Schmugge T (2009) Using ASTER satellite data to calculate riparian evapotranspiration in the Middle Rio Grande, New Mexico. Int J Remote Sens 30:5593–5603
Burba G, Schmidt A, Scott RL, Nakai T, Kathilankal J, Fratini G, Hanson C, Law B, McDermitt DK, Eckles R, Furtaw M, Velgersdyk M (2012) Calculating CO2 and H2O eddy covariance fluxes from an enclosed gas analyzer using an instantaneous mixing ratio. Glob Change Biol 18:385–399
Campbell GS, Norman JM (1998) An introduction to environmental biophysics. Springer-Verlag New York Inc, New York
Chen YN, Wang Q, Li W, Ruan X, Chen Y, Zhang L (2006) Rational groundwater table indicated by the eco-physiological parameters of the vegetation: a case study of ecological restoration in the lower reaches of the Tarim River. Chin Sci Bull 51:8–15
Cheng GD, Li X, Zhao WZ, Xu ZM, Feng Q, Xiao SC, Xiao HL (2014) Integrated study of the water-ecosystem-economy in the Heihe River basin. Natl Sci Rev 1:413–428
Cleverly JR, Dahm CN, Thibault JR, Gilroy DJ, Allred Coonrod JE (2002) Seasonal estimates of actual evapo-transpiration from Tamarix ramosissima stands using three-dimensional eddy covariance. J Arid Environ 52:181–197
Cleverly JR, Dahm CN, Thibault JR, McDonnell DE, Allred Coonrod JE (2006) Riparian ecohydrology: regulation of water flux from the ground to the atmosphere in the Middle Rio Grande, New Mexico. Hydrol Process 20:3207–3225
Dahm CN, Cleverly JR, Allred Coonrod JE, Thibault JR, McDonnell DE, Gilroy DJ (2002) Evapotranspiration at the land/water interface in a semi-arid drainage basin. Freshw Biol 47:831–843
Dennison PE, Nagler PL, Hultine KR, Glenn EP, Ehleringer JR (2009) Remote monitoring of tamarisk defoliation and evapotranspiration following saltcedar leaf beetle attack. Remote Sens Environ 113:1462–1472
Devitt DA, Sala A, Smith SD, Cleverly J, Shaulis LK, Hammett R (1998) Bowen ratio estimates of evapotranspiration for Tamarix ramosissima stands on the Virgin River in southern Nevada. Water Resour Res 34:2407–2414
Devitt DA, Fenstermaker LF, Young MH, Conrad B, Baghzouz M, Bird BM (2011) Evapotranspiration of mixed shrub communities in phreatophytic zones of the Great Basin region of Nevada (USA). Ecohydrology 4:807–822
Di Tomaso JM (1998) Impact, biology, and ecology of Saltcedar (Tamarix spp.) in the southwestern United States. Weed Technol 12:326–336
Feng Q, Cheng GD (1998) Current situation, problems and rational utilization of water resources in arid north-western China. J Arid Environ 40:373–382
Feng Q, Peng JZ, Li J, Xi HY, Si JH (2012) Using the concept of ecological groundwater level to evaluate shallow groundwater resources in hyperarid desert regions. J Arid Land 4:378–389
Foken T, Gockede M, Mauder M, Mahrt L, Amiro B, Munger W (2004) Post-field data quality control. In: Xuhui L, William M, Beverly L (eds) Handbook of micrometeorology: a guide for surface flux measurements. Kluwer Academic Publishers, New York, pp 181–203
Gao GL, Zhang XY, Yu TF, Liu B (2016a) Comparison of three evapotranspiration models with eddy covariance measurements for a Populus euphratica Oliv. forest in an arid region of northwestern China. J Arid Land 8:146–156
Gao GL, Zhang XY, Yu TF (2016b) Evapotranspiration of a Populus euphratica forest during the growing season in an extremely arid region of northwest China using the shuttleworth-wallace model. J For Res 27:879–887
Glenn EP, Nagler PL (2005) Comparative ecophysiology of Tamarix ramosissima and native trees in western U.S. riparian zones. J Arid Environ 61:419–446
Guo QL, Feng Q, Li JL (2008) Environmental changes after ecological water conveyance in the lower reaches of Heihe River, northwest China. Environ Geol 58:1387–1396
Hao YB, Wang Y, Huang X, Cui X, Zhou X, Wang S, Niu H, Jiang G (2007) Seasonal and interannual variation in water vapor and energy exchange over a typical steppe in Inner Mongolia, China. Agric For Meteorol 146:57–69
Hao XM, Li WH, Huang X, Zhu C, Ma J (2009) Assessment of the groundwater threshold of desert riparian forest vegetation along the middle and lower reaches of the Tarim River, China. Hydrol Process 24:178–186
Hatler WL, Hart CR (2009) Water loss and salvage in saltcedar (Tamarix spp.) stands on the Pecos River, Texas. Invasive Plant Sci Manag 2:309–317
Hou LG, Xiao HL, Si JH, Xiao SC, Zhou MX, Yang YG (2010) Evapotranspiration and crop coefficient of Populus euphratica Oliv forest during the growing season in the extreme arid region northwest China. Agric Water Manag 97:351–356
Nagler PL, Glenn EP, Lewis Thompson T (2003) Comparison of transpiration rates among saltcedar, cottonwood and willow trees by sap flow and canopy temperature methods. Agric For Meteorol 116:73–89
Nagler P, Scott R, Westenburg C, Cleverly J, Glenn E, Huete A (2005) Evapotranspiration on western U.S. rivers estimated using the Enhanced Vegetation Index from MODIS and data from eddy covariance and Bowen ratio flux towers. Remote Sens Environ 97:337–351
Nagler PL, Morino K, Didan K, Erker J, Osterberg J, Hultine KR, Glenn EP (2009) Wide-area estimates of saltcedar (Tamarix spp.) evapotranspiration on the lower Colorado River measured by heat balance and remote sensing methods. Ecohydrology 2:18–33
Sala A, Smith SD, Devitt DA (1996) Water use by Tamarix ramosissima and associated phreatophytes in a Mojave Desert floodplain. Ecol Appl 6:888–898
Shafroth PB, Cleverly JR, Dudley TL, Taylor JP, van Riper C III, Weeks EP, Stuart JN (2005) Control of Tamarix in the Western United States: implications for water salvage, wildlife use, and riparian restoration. Environ Manag 35:231–246
Si JH, Feng Q, Zhang XY, Liu W, Su YH, Zhang YW (2005) Growing season evapotranspiration from Tamarix ramosissima stands under extreme arid conditions in northwest China. Environ Geol 48:861–870
Si JH, Feng Q, Zhang XY, Chang ZQ, Su YH, Xi HY (2007) Sap flow of Populus euphratica in a desert riparian forest in an extreme arid region during the growing season. J Integr Plant Biol 49:425–436
Si JH, Feng Q, Cao SK, Yu TF, Zhao CY (2014) Water use sources of desert riparian Populus euphratica forests. Environ Monit Assess 186:5469–5477
Smith SD, Devitt DA, Sala A, Cleverly JR, Busch DE (1998) Water relations of riparian plants from warm desert regions. Wetlands 18:687–696
Sun G, Noormets A, Chen J, McNulty SG (2008) Evapotranspiration estimates from eddy covariance towers and hydrologic modeling in managed forests in Northern Wisconsin, USA. Agric For Meteorol 148:257–267
Taghvaeian S, Neale CMU, Osterberg J, Sritharan SI, Watts DR (2014) Water use and stream-aquifer-phreatophyte interaction along a Tamarisk-dominated segment of the lower Colorado River. Remote Sens Terr Water Cycle, Geophysical Monograph 206, pp 95–113
Vickers D, Mahrt L (1997) Quality control and flux sampling problems for tower and aircraft data. J Atmos Ocean Technol 14:512–526
Wang P, Grinevsky SO, Pozdniakov SP, Yu JJ, Dautova DS, Min L, Du C, Zhang Y (2014) Application of the water table fluctuation method for estimating evapotranspiration at two phreatophyte-dominated sites under hyper-arid environments. J Hydrol 519:2289–2300
Westenburg CL, Harper DP, DeMeo GA (2006) Evapotranspiration by phreatophytes along the lower Colorado River at Havasu National Wildlife Refuge, Arizona. In: Geological survey scientific investigations report 2006-5043. Henderson
Wilson K, Goldstein A, Falge E, Aubinet M, Baldocchi D, Berbigier P, Bernhofer C, Ceulemans R, Dolman H, Field C, Grelle A, Ibrom A, Law BE, Kowalski A, Meyers T, Moncrieff J, Monson R, Oechel W, Tenhunen J, Valentini R, Verma S (2002) Energy balance closure at FLUXNET sites. Agric For Meteorol 113:223–243
Yu TF, Feng Q, Si JH, Xi HY, Li W (2013a) Patterns, magnitude, and controlling factors of hydraulic redistribution of soil water by Tamarix ramosissima roots. J Arid Land 5:396–407
Yu TF, Feng Q, Si JH, Xi HY, Li ZX, Chen AF (2013b) Hydraulic redistribution of soil water by roots of two desert riparian phreatophytes in northwest China’s extremely arid region. Plant Soil 372:297–308
Yuan GF, Zhang P, Shao MA, Luo Y, Zhu XC (2014) Energy and water exchanges over a riparian Tamarix spp. stand in the lower Tarim River basin under a hyper-arid climate. Agric For Meteorol 194:144–154
Zhang XY, Kang ES, Si JH, Zhou MX (2006) Stem sap flow of individual plant of Populus euphratica and its conversion to forest water consumption. Scientia Silvae Sinicae 42:28–32
Acknowledgements
The authors would like to thank two anonymous reviewers for their constructive and valuable assessment and comments, which helped to improve this article.
Author information
Authors and Affiliations
Corresponding author
Additional information
Project funding: This study was supported by the National Natural Science Foundation of China (41401033, 31370466, and 41271037), the China Postdoctoral Science Foundation (2014M560819) and the National Key Research and Development Program of China (2016YFC0501002).
The online version is available at http://www.springerlink.com
Corresponding editor: Hu Yanbo.
Rights and permissions
About this article

Cite this article
Yu, T., Feng, Q., Si, J. et al. Comparable water use of two contrasting riparian forests in the lower Heihe River basin, Northwest China. J. For. Res. 29, 1215–1224 (2018). https://doi.org/10.1007/s11676-017-0540-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-017-0540-2