
Abstract
An evaluation of 30 naturally available candidate-plus trees (CPTs) of Pongamia pinnata was carried out to elucidate the genetic variability and relationship of pod and seed traits and progeny growth traits to select the best planting material with higher productivity. Significant variations were observed in pod and seed characteristics, oil content, and during the progeny trial studies. The highest values were found in 100-pod weight (410.4 gm) and 100-seed weight (195.92 gm) and the phenotypic and genotypic variance was found maximum in 100 pod weight (4364.71 and 4289.93) and 100 seed weight (813.8 and 768.93), respectively. Estimates of the phenotypic coefficient of variance were higher than the genotypic coefficient of variance for all the pod and seed traits and progeny growth traits, which depict the predominant role of the environment. Approximately 99 % of broad-sense heritability was revealed in oil content, followed by 100-pod weight (98.28 %), and canopy growth (96.93 %). Pod thickness shows the highest genetic advance of 56.61 %, followed by pod width (53.43 %) and canopy growth (49.3 %). Pod and seed traits have proven a positive correlation between each other and with progeny growth traits, except a few negative values. Hierarchical clustering by Ward’s minimum variance cluster analysis showed phylogeographical patterns of genetic diversity. K-means clustering revealed that trees from different geographical regions were grouped together in a cluster where as trees from the same geographical area are placed in a different cluster. Seven Pongamia ecotypes (GRP-8, 9, 13, 14, 18, 28, and 29) are found superior in traits, namely 100-pod weight, 100-seed weight, oil content, germination rate, plant height, canopy growth, and collar diameter. The traits 100-pod weight and 100-seed weight are highly correlated with the progeny growth traits and even have higher heritability and genetic advantages.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
India is the 11th largest economy in the world, fourth in terms of purchasing power and fifth largest energy consumer in the world, importing around 80 % of its total petroleum oil and lubricants (POL) (Indian Foreign Trade 2012). Fluctuating global prices, depletion of oil reserves, and environmental pollution are the major concerns associated with the use of fossil fuels. To sustain a clean environment and to meet future demands, India has to be committed to conduct research on alternative energy resources, such as utilization of biodiesel and bioethanol replacing the petrol and petro-diesel for transportation. Biodiesel plants have created tremendous interest all over the world for the use of seed oil as a commercial source of biodiesel. India has suitable climatic conditions and has much waste or barren land for cultivation of biodiesel plant species, mainly Pongamia pinnata, which is known to grow in these conditions. It has also been identified as a promising plant resource for the production of biodiesel. The challenging task as of today is to screen the naturally available wild P. pinnata plant resources for the best plant material with high productivity.
Pongamia pinnata (L.) Pierre is a fast-growing, glabrous, deciduous tree, commonly known as Indian beech, pongam tree, or karanja tree. It is a member of the subfamily Papilionideae, more specifically the tribe Millettieae. It is indigenous to the Indian subcontinent and South East Asia, and has been successfully introduced to humid and tropical regions of the world (Scott et al. 2008). It is tolerant of a wide range of abiotic stresses like drought, frost, heat, and salinity. It grows fast and matures after 4–7 years, yielding fruits that are flat, elliptical, and long; each fruit contains one or two kidney-shaped brownish red kernels. The oil content in the kernel is about 30–40 % and the fatty acid composition has been reported to be oleic (44 %), linoleic acid (17.4 %), erucic (15.9 %), palmitic (7.2 %), linolenic (5.5 %), stearic (3.3 %), 11-eicosenoic (3.4 %), behenic (2.5 %) and eicosanoic (0.78 %) acids (Bala et al. 2010). Pongamia seed oil, which has similar properties to that of petro-diesel, has gained importance as a biodiesel, which is a fast-emerging, viable alternative to fossil fuel.
Pod and seed traits are important in evaluating trees of unknown genetic potential, especially a tree like Pongamia, where little work has been done in terms of progeny evaluation. Yield stabilization of this tree is expected to be seen only after 7–8 years of establishment, and so it would take a minimum of 10 years to identify superior germplasm in terms of seed yield. Genetic variation in seed characteristics, growth, branching pattern, and yield can be of great potential in tree improvement programmes, particularly for the selection of genotypes having more oil content and yield.
Hence in our present investigation, the CPTs of Pongamia are evaluated by phenotypic and genotypic analysis of variability, broad sense heritability, genetic advance, and progeny trial to identify the best planting material. Similar studies have been carried out by several researchers across India for the selection of suitable quality plants for the respective ideotypes for afforestation programme. (Kaushik et al. 2007; Sunil et al. 2009; Divakara et al. 2010; Divakara and Rameshwar 2011; Rao et al. 2011; Deebe et al. 2011 and Pavithra et al. 2013).
Materials and methods
A wild germplasm exploration survey was conducted to identify the elite germplasm with high-yielding CPTs of Pongamia from different agro-climatic conditions in North Karnataka (Hyderabad Karnataka), India as given in Table 1. The survey covers an area of 45,582.88 km2 and the region has approximately 1.5–2 lakh hectares of waste or barren land. Collection of seeds and stem cuttings from CPTs was carried out over a two-year period from May to June 2011–2012 and 2012–2013. The selection criteria for CPTs was made by phenotypic assessment of characteristics as advocated by Cornelius (1994) and Kaushik et al. (2007).
A total of 76 Pongamia trees were identified as disease free, healthy, and high yielding plants. All plants were 15 years or older and out of these, 30 CPTs were selected based on their morphological and qualitative traits (e.g., yield potential, branching pattern, number of flowers per raceme, crown spread, girth, and disease resistance). A total of 300 healthy pods and seeds (hundred in each replication) were evaluated for their length, width, and thickness using a vernier caliper. The 100-pod weight and 100-seed weights were evaluated, using a scientific weighing scale in a randomized manner. Seed characteristics were analyzed after separation from pods, and the seeds were then cleaned and stored in muslin bags at ambient conditions. All seed lots were dried under ambient temperature and humidity to attain a constant weight. The oil content (percent) in seeds was estimated by soxhlet method using n-hexane as a solvent. The progeny trial study was conducted in the nursery of Biodiesel Technology Park, Gulbarga University campus. The germplasm accessions were raised under a randomized block design with three replications and planted at a spacing of 3 × 3 m, and the growth readings were recorded for germination percentage or germination rate (GR), seedling height or plant height (PH), collar diameter (CD), canopy growth (CG), and number of branches per plant (NBP) as suggested by Mukta et al. (2009), Divakara et al. (2010), NOVOD (2010), Deebe et al. (2011), and Rao et al. (2011). The canopy growth was measured in both north–south and east–west directions of the crown spread of the plant and its average was expressed in cm.
Statistical analysis
The raw data of all the parameters with replications was compiled and analyzed using analysis of variance (ANOVA) to calculate the mean, standard error of mean, variance, and coefficient of variation using statistical software SPSS (version 21). The correlation coefficient was analyzed to determine the relationship between pod and seed characteristics and progeny traits, both at the phenotypic and genotypic level. Phenotypic and genotypic variance and phenotypic and genotypic coefficient of variation were performed as suggested by different researchers (Baenziger et al. 2004; Pliura et al. 2007; Yoshida et al. 2007). Broad sense heritability was estimated by the variations in between and within the accessions by calculating the ratio of genotypic variance to the phenotypic variance (Falconer and MacKay 1996; Allard 1999).
\( {{\sigma }}^{2} {\text{g}} \) is genotypic variance = (variance between accessions-variance within accessions)/n
\( {{\sigma }}^{2} {\text{p}} \) is phenotypic variance = [(variance between accessions-variance within accessions)/n] + variance within accessions.
where PCV is the phenotypic coefficient of variation, GCV is the genotypic coefficient of variation, n is the number of replicates per treatment and X is the grand mean of each pod and seed trait and progeny trait respectively. Genetic advance (GA) and genetic advance as percent of mean were estimated in accordance with Johanson et al. (1955) as:GA = K·h 2 B· \( \sqrt {\sigma^{2} p} \); Genetic advance (as percent of mean) = (GA/X) × 100
The association and variation in-between and within the pod and seed parameters was observed by hierarchical euclidean cluster analysis by divisive method and non-hierarchical (K-means) by an agglomerative method for initially collected Pongamia accessions. The shortest Euclidean distances in the close neighborhood was calculated by Ward’s method (Ward 1963) and a dendrogram was constructed to prove the relationship within and between the population (Sokal and Rohlf 2003).
Results
Variability in pod and seed traits
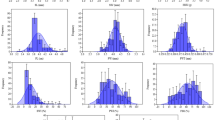
Analysis of variance of pod and seed traits revealed a significant variation among all the wild accessions studied for all the characters at P < 0.05. Variability studies revealed that the accession GRP-29 has recorded maximum values for pod width (2.96 cm), 100-pod weight (410.4 gm), 100-seed weight (195.92 gm), and oil content (43.53 %). The accession GRP-18 has shown maximum values for pod thickness (1.74 cm), seed length (2.71 cm), and seed width (1.74). The accession GRP-22 has higher values in pod length (5.97 cm) and pod width (2.96 cm). The accessions GRP-8, GRP-14, GRP-18, and GRP-29 showed highly significant values for 100-pod weight and 100-seed weight compared to other accessions.
Accessions having pod lengths greater than 5 cm was recorded in GRP-5, GRP-6, GRP-8, GRP-10, GRP-15, GRP-16, GRP-18, and GRP-22. Pod width of greater than 2.6 cm was recorded in GRP-4, GRP-5, GRP-21, GRP-22, GRP-28, GRP-29, and GRP-30. Seed length and seed width was recorded highest in GRP-2, GRP-8, GRP-12, GRP-18 and GRP-22. Oil content of more than 38 % was observed in accessions GRP-29, GRP-28, GRP-18, GRP-16, GRP-13, GRP-8, and GRP-27. Overall, 19 accessions have an oil content of more than 35 %. Accessions GRP-2, GRP-3, GRP-4, GRP-21, and GRP-25 have minimum values for pod and seed traits (e.g., 100PW and 100SW as shown in Table 2).
Genetic association and diversity among pod and seed traits
Genetic association and variability was evident from the study of phenotypic coefficient of variation (PCV), genotypic coefficient of variation (GCV), broad sense heritability (h2B) and genetic advance (GA). The magnitude of PCV was greater than the corresponding GCV for all of the pod and seed characteristics, indicating a predominant role of environmental factors. The phenotypic and genotypic coefficients of variance were closer to each other for all traits. The phenotypic variance (\( \sigma^{2} p \)) and genotypic variance (\( \sigma^{2} g \)) as higher for 100PW (4364.71 and 4289.93) and 100SW (813.8 and 768.93) respectively, moderate for germination rate, plant height, canopy growth, and oil content and low for the remaining traits.
Pod width and seed thickness have exhibited much higher value for both GCV and PCV in comparison to pod and seed traits and the canopy growth (CG) was observed the highest with a PCV value of 24.69 and GCV values of 24.31 in plant growth studies. Broad sense heritability was high in general and exceeded 70 % for all the pod and seed traits studied and the highest value of 99 % was observed in OC followed by 100PW (98.28 %), 100SW (94.48 %), PW (92.29 %) as given in Table 3.
The characteristics 100PW, 100SW, OC, GR, PH, CG, and CD showed a similar degree of variation in PCV and GCV, which may be due to maternal or environmental factors (Gutterman 2000). Among the progeny trial, h2B was highest in CG (96.93 %) followed by GR (96.1 4 %), CD (95.91 %), and PH (95.7 %). Genetic advance as percent of mean was observed highest in PT (56.61 %) followed by PW (53.43 %) and CG (49.3 %) and other traits and least was recorded in OC (16.54 %).
The correlation coefficient of pod and seed traits and progeny growth traits shows the contribution of each component to the yield and an indirect effect in association with the other components. The correlation coefficient (along the horizontal and vertical axises) between PL, PW, SL, ST, PSR, OC, GR, and PH were prominent and are considered as the main components in tree breeding programs. The association of pod, seed, and progeny growth traits are significantly correlated with each other, except for few negative values in both phenotypic and genotypic levels as in Table 4.
Euclidean clusters and cluster mean values, obtained by K-means non-hierarchical clustering for pod and seed characters of 30 accessions of Pongamia, are placed in five clusters, as depicted in Table 5. The maximum number of accessions are in cluster V (12 accessions); they have shown good mean values with respect to seed thickness, pod seed ratio, and number of branches. Cluster III with minimum accessions (3 accessions) has reliable mean values in almost all the traits studied except ST and CG. The dendrogram obtained from hierarchical Euclidian cluster analysis, according to Ward’s method, is shown in Fig. 1. The length of horizontal branches between clusters proved a high degree of dissimilarity between clusters. The maximum turn of members, apart from the super cluster containing all the accessions are populated among 30 accessions. The Pongamia ecotypes GRP-23 and GRP-29 coincide at a distance of 25 with 23 accessions between their values.

Dendrogram constructed by Ward method of cluster analysis on the Euclidean distances by hierarchical clustering of growth and reproductive phenology in progeny trial of selected Pongamia germplasm. The location of accession has been listed in Table 2
Progeny trial characters
Significant variations are found between and within the growth of 30 CPTs of Pongamia from the initial seed sources. Seven accessions have a germination rate of more than 80 % and only four accessions have a germination rate less than 70 %. Similar results was recorded in PH, CD, CG, and NBP in accession GRP-21, GRP-22, and GRP-29. A few low-ranking accessions are GRP-1, GRP-7, and GRP-26 from the initial trials; medium-ranking accessions are GRP-6, GRP-18, GRP-24, GRP-25, and GRP-28 (see Table 6). The lowest plant height was recorded in GRP-1 (57.76 cm) and the highest collar diameter is recorded in GPR-22 (1.12 cm). The collar diameter was observed to be more than 1.1 cm in four accessions and only one accession (GRP-1) has a value less than 0.9 cm. The highest number of branches per plant the range of only 5.7–5.9 branches in GRP-8, GRP-11, GRP-12, GRP-13, GRP-14 and GRP-19 and the highest of 7.7 branches in GRP-18. Most accessions have more than 5.8–6.9 branches with an intermediate value of 6.5.
The accessions with greater values in pod and seed traits (GRP-8, 9, 13, 14, 18, 28, and GRP-29) have shown a good germination rate of more than 70 %. Except for GRP-13, plant height ranged between 70–86 cm, where the accession GRP-29 showed high values for germination rate, plant height, and moderate values for collar diameter, canopy growth and number of branches per plant which indicates direct relationship between the seed size and oil content to germination rate and plant height.
The accessions from Raichur (GRP-10, 11, 12, and GRP-13) and Bellary (GRP-19, 20, 21, and GRP-22) having 100SW ranging from 131 to 177 gm and oil content ranging from 32 to 38 % when grown in Gulbarga region showed good germination rate, plant height, collar diameter, canopy growth and number of branches per plant depicts the role of the environment plays moderate role in the growth of the progenies.
The accessions (GRP-14, 15, 16, 17, and GRP-18) collected from Bidar district shows good values in all pod and seed traits and even in progeny growth studies. While Bidar has the highest rainfall and is at a higher altitude in North Karnataka region (Table 1), the Pongamia accessions showed a constant performance during progeny studies. The Pongamia ecotypes GRP-8, 14, and 18 are grouped with GRP-29 (superior quality based on high 100 seed-weight and oil content) in the non-hierarchical Euclidean cluster (Table 5). The dendrogram (Fig. 1) depicts the relevance of similar characters among these accessions, which are from different geographical regions.
Discussion
Pongamia is undoubtedly the most prominent non-edible oil-yielding plant species for combating the global fuel crisis through the production of biodiesel. In an agricultural country like India, having much barren/waste land and suitable climatic conditions for cultivation of biofuel plant species like Pongamia could produce feedstock of approximately 12–18 tons of seeds per year per hectare, which could produce 3000–4000 l of biodiesel per hectare at the least productive level.
But Pongamia has not yet been explored in terms of identifying elite quality plants through morphological and reproductive markers, which could be the beginning stage of breeding elite genotypes in relation to oil content and productivity. It is a highly cross-pollinated species and exhibits a wide spectrum of variation in pod and seed traits. The growth of Pongamia wild accessions under diverse conditions causes morphological, biochemical, and genetic adaptation because multiple factors may stimulate and maintain variations in the pod and seed size. The larger seeds may be chosen because they produce bigger and more vigorous seedlings with a better chance of natural selection as compared to small seeds (Kaushik et al. 2007; Kesari et al. 2008).
Variability in pod and seed characteristics
Pod and seed morphological characteristics of the wild accessions are considered to be the first step in ascertaining genetic variability of the population. Identification of better genotypes or superior phenotypes having better yield and oil content is the initial step in any tree improvement strategy. On the other hand, selection based on the variability in pod and seed characteristics alone is not a sure step in the selection of high production. Thus, it is imperative to follow up with the selection based on growth in the initial years for faster operation of breeding programmes especially in tree crops. Several researchers have studied variations in morphological (pod and seed traits), progeny growth for the identification of high quality planting material in different geographic regions across India (the data is represented in Table 7).
Variations in seed oil content are more widely reported in a wide variety of trees (O’Neill et al. 2003). Similar variations in morphological characteristics were evident in our present study, namely attributes that are a viable selection alternative at a very early stage (collection of germplasm) from the base seed material. Variation in growth of Pongamia ecotypes was clearly evident from the present study: pod width, pod thickness, and seed thickness were recorded with the highest values when compared to the other components as given in Table 7.
All the parameters exhibited a considerable amount of variability in the nursery as well in progeny growth study. The association between seed size and germination efficiency in Pongamia has been reported by Manonmani et al. (1996). Environmental factors cause the phenotypic and genotypic variations in plants to evolve a new plant type, hence the seeds good characteristics may be selected for producing better progenies in addition to 100 pod weight and 100 seed weight (Jaishankar et al. 2014).
The ability to estimate heritability is important because it makes it possible to rank the importance of each trait in a crossbreeding program, which depends on the type and extent of genetic variability. The study of GCV and heritability will give an idea for selection of candidate plus trees, as the level of heritability may be controlled by additive gene action in the expression of the characteristics. The marginal differences between PCV and GCV of all the characteristics studied suggest high heritability of variation among the characters and high values of genetic advance. These are indicative of additive genes that are actively involved in the expression of various polygenic traits and low values of non-additive gene action. The correlation coefficient analyses the relationship of variables but cannot quantify the effect of genotypes on dependent variables.
The phenotypic and genotypic correlation of pod and seed characteristics offers an opportunity to select phenotypes based on these traits. The genetic correlations are more reliable estimates for examining the relationship between characteristics. This offers an opportunity to select phenotypes with good quantitative characteristics resulting in improvement programs (Mahmood et al. 2003). In our present investigation, the correlation coefficient studies revealed interesting relationships between pod and seed characteristics to initial growth parameters. The phenotypic coefficient of correlation was higher than the corresponding genotypic coefficient of correlation in most of the parameters, indicating the genetic association among the characters. Correlated quantitative traits are of major interest in any tree-improvement programme, as the improvement of one character may cause simultaneous changes in other characters.
Genetic diversity
Analysis of genetic diversity in germplasm collections can facilitate consistent classification and recognition of elite accessions. Assessment in the level and pattern of genetic diversity can be helpful in crop breeding for analysis of inherited variability in cultivars and identification of diverse parental combinations. Hierarchical clustering of pod and seed characteristics and progeny growth traits, using Ward’s Minimum Cluster Analysis method, indicates a high degree of genetic diversity. The dendrogram indicates the relative participation of different constituent traits to the total diversity and quantity divergence between populations. Non-hierarchical (K means) clustering by is done to understand the trend of evolution and to choose genetically diverse parents in order to obtain the desirable recombination as suggested by Tams et al. (2006). Similarly in our present study, Pongamia ecotypes GRP-8, 14, 18 and GRP-29 are grouped in same cluster (cluster II, Table 5) and in non-hierarchical clustering (Fig. 1), which indicates these accessions may have similar quality characteristics, where GRP-29 has high values for 100-PW, 100-SW, and oil content and moderate values for progeny growth studies. The maximum distances indicate extensive genetic differences between the trees of these groups. These attributes are helpful for the selection of parents for novel hybrids with increased characteristics like increased growth, oil content, and yield.
The present study also reveals the details of the candidate plus trees necessary in the tree improvement program via the traits like 100-pod weight and 100-seed weight, which were highly correlated with the progeny growth traits. Hence, the identification of elite quality Pongamia ecotypes (GRP-8, 9, 13, 14, 28, and 29) are found superior in traits, namely 100PW, 100SW, OC, GR, PH, CG, and CD.
The North Karnataka region has about 1.5 lakh hectares of waste or barren land, where the plantation of Pongamia plants could produce 400–500 ml of biodiesel even by conventional methods. This could fulfil the need of biodiesel in the state and also generate employment in the Hyderabad Karnataka region. It could also increase the forest cover in the region and help to provide a clean environment. Hence, plantation of Pongamia trees may be helpful for massive afforestation programs in the semi-arid conditions of North Karnataka. Also, the present study helps as a pointer for taking a molecular biology and genetics approach to characterize genomic studies.
References
Allard RW (1999) Principles of plant breeding, 2nd edn. Wiley, New York, p 264
Baenziger SP, Gregory S, McMaster R, Wilhelm WW, Weiss A, Hays CJ (2004) Putting genes into genetic coefficients. Field Crops Res 90:133–143
Bala M, Nag TN, Kumar S, Vyas M, Kumar A, Bhogal NS (2010) Proximate composition and fatty acid profile of Pongamia pinnata, a potential biodiesel crop. J Am Oil Chem Soc. doi:10.1007/s11746-010-1699-2
Cornelius J (1994) The effectiveness of plus tree selection for yield. For Ecol Manag 67(1):23–34
Deebe PS, Rout GR, Das S, Aparajita S, Mahapatra AK (2011) Genotypic variability and correlation studies in pod and seed characteristics of Pongamia pinnata (L.) Pierre in Orissa, India. Int J For Res. doi:10.1155/2011/728985
Divakara BN, Rameshwar D (2011) Variability and divergence in Pongamia pinnata for further use in tree improvement. J For Res 22(2):193–200
Divakara BN, Alur AS, Tripati S (2010) Genetic variability and relationship of pod and seed traits in Pongamia pinnata (L.) Pierre, a potential agroforestry tree. Int J Plant Prod 4(2):1735–8013
Falconer DS, MacKay TFC (1996) Introduction to quantitative genetics. Chapman and Hall, London
Gutterman Y (2000) Maternal effects on seeds during development. In: Fenner M (ed) Seeds: the ecology of regeneration in plant communities, 2nd edn. CABI Publishing, Wallingford, pp 59–84
Indian foreign trade: 2011–2012. RBI monthly bulletin, March 2012
Jaishankar I, Sankaran M, Singh DR, Damodaran V (2014) Genetic variability and divergence studies in pod and seed traits of Pongamia pinnata (L.) Pierre, accessions in Bay islands. J For Res 25(2):351–358
Johanson HW, Robinson HF, Comstock RE (1955) Estimate of genetic and environmental variability in soybeans. Agron J 47:314–318
Kaushik N, Kumar S, Kumar K, Beniwal RS, Kaushik N, Roy S (2007) Genetic variability and association studies in pod and seed traits in Pongamia pinnata (L.) Pierre in Haryana India. Genet Resour Crop Evol 54:1827–1832
Kesari V, Krishnamachari A, Rangan L (2008) Systematic characterization and seed oil analysis in candidate plus trees of biodiesel plant, Pongamia pinnata. Ann Appl Biol 152:397–404
Mahmood K, Marcar NE, Naqvi MH, Arnold RJ, Crawford DF, Iqbal S, Aken KM (2003) Genetic variation in Eucalyptus camaldulensis Dehnh. for growth and stem straightness in a provenance–family trial on saltland in Pakistan. For Ecol Manag 176:405–416
Manonmani V, Vanagamudi K, Vinaya Rai RS (1996) Effect of seed size on seed germination and vigour in Pongamia pinnata. J Trop For Sci 9:1–5
Mukta N, Murthy IYLN, Sripal P (2009) Variability assessment in Pongamia pinnata (L.) Pierre germplasm for biodiesel traits. Ind Crops Prod 29:536–540
NOVOD (2010) 5th R&D report on tree borne oilseeds. In: NOVOD Board India. http://www.novodboard.com/5th%20R&D-Report.pdf. Accessed on 2 June 2010
O’Neill CM, Gill S, Hobbs D, Morgan C, Bancroft I (2003) Natural variation for seed oil composition in Arabidopsis thaliana. Phytochemistry 64:1077–1090
Pavithra HR, Gowda B, Prasanna KT, Shivanna MB (2013) Pod and seed traits in candidate plus trees of Pongamia pinnata (L.) Pierre from Southern Peninsular India in relation to provenance variation and genetic variability. J Crop Sci Biotechnol 16(2):131–142
Pliura A, Zhang SY, MacKay J, Bousquet J (2007) Genotypic variation in wood density and growth traits of poplar hybrids at four clonal trials. For Ecol Manag 238:92–106
Rao GR, Shanker AK, Srinivas I, Korwar GR, Venkateswarlu B (2011) Diversity and variability in seed characters and growth of Pongamia pinnata (L.) Pierre accessions. Trees 25:725–734
Scott PT, Pregel L, Chen N, Hadler JS, Djordjevic MA, Gresshoff PM (2008) Pongamia pinnata: an untapped resource for the biofuels industry of the future. Bioenergy Res 1:2–11
Sokal RR, Rohlf FJ. 2003. Biometry: the principles and practice of statistics in biological research. 8th Printing. Freeman W.H. and Comp, San Francisco
Sunil N, Kumar V, Sivaraj N, Lavanya C, Prasad RBN, Rao BVSK, Varaprasad KS (2009) Variability and divergence in Pongamia pinnata (L.) Pierre germplasm- a candidate tree for biodiesel. GCB Bioenergy 1:382–391
Tams SH, Bauer E, Oettler G, Melchinger AE, Scho¨n CC (2006) Prospects for hybrid breeding in winter triticale: II. Relationship between parental genetic distance and specific combining ability. Plant Breed 125:331–336
Ward JH (1963) Hierarchical grouping to optimize an objective function. J Am Stat Assoc 58:236–244
Yoshida H, Takeshi H, Katsura K, Tatsuhiko S (2007) A model explaining genotypic and environmental variation in leaf area development of rice based on biomass growth and leaf N accumulation. Field Crops Res 102:228–238
Acknowledgments
The authors thank UGC, Government of India, New Delhi, the funding agency for project UGC-MRP No. F. No-39-258/2010 (SF) dated 28.12.2010 and also to Karnataka State Biofuel Development Board (KSBDB) Bengaluru for the establishment of a Biodiesel unit in Gulbarga University, Gulbarga.
Author information
Authors and Affiliations
Corresponding author
Additional information
Project funding: This work was supported by the project UGC-MRP No. F. No-39-258/2010 (SF).
The online version is available at http://www.springerlink.com
Corresponding editor: Hu Yanbo
Rights and permissions
About this article

Cite this article
Patil, V.K., Naik, G.R. Variability in pod and seed traits of Pongamia pinnata Pierre ecotypes in North Karnataka, India. J. For. Res. 27, 557–567 (2016). https://doi.org/10.1007/s11676-015-0191-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-015-0191-0