
Abstract
We studied forest vegetation at Lata–Tolma–Phagti, a protected area and part of Nanda Devi Biosphere Reserve in the western Himalaya. We analyzed community composition, population structure, regeneration patterns, and projected development of future compositional patterns. We sampled ten 10 × 10 m quadrat for tree species in each of 30 forest stands. We sampled shrubs in ten 5 × 5 m quadrat, and herbs in twenty 1 × 1 m quadrat within each forest stand. We recorded 248 plant species from 8 forest communities. Broadly the demographic profiles at study sites exhibited progressive structures suggesting long term persistence of the communities/species. Density–diameter distribution revealed greater proportions of seedlings and a significant decline (P < 0.05) in the proportion of trees in older age/size classes. Of the 23 recorded tree species, 13.0 % showed good, 52.2 % fair, 26.1 % poor and 8.7 % no regeneration. Differences in regeneration by species are indicative of future forest structure and dynamics. Assessment of changes in structure of forest types provides baseline data for development of priorities for conservation of other representative landscapes in the reserve as well as in the Himalaya.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Tree diversity, forest structure, tree growth, and forest turnover are important parameters for monitoring ecological processes in dynamic environments. Forests are characterized by their three main life stages called seedling (newly emerged plants), sapling (established plants between seedling and tree) and tree (plants undisturbed by micro-environmental conditions). The number (density) and type (richness) of trees define the structure and composition of forests (Shankar 2001; Mishra et al. 2003). Species richness patterns in relation to the environment need to be understood before drawing conclusions on the effect of biodiversity on ecosystem processes. The number of trees at seedling, sapling and tree age classes reveals population structure and their establishment at seedling and sapling level represent regeneration status (Baduni and Sharma 2001; Bhandari 2003).
The nature of forest communities depends on the ecological characteristics of sites, species diversity and regeneration status of species (Criddle et al. 2003; Todaria et al. 2010). The tree strata (i.e. seedling, sapling and tree layers that define the population structure of a forest) are affected by micro-environmental factors that vary seasonally (Khumbongmayum et al. 2006; Kharkwal 2009). Environmental variation within a small geographical area makes elevational gradients ideal for investigating several ecological and biogeographical hypotheses (Korner 1998). It is necessary to understand species richness, population structure, germination and establishment of seedlings and saplings across seasons and elevations for maintenance of forest structures (Khumbongmayum et al. 2006; Rao 1988). Absence of tree seedlings and saplings indicates poor regeneration, while their presence indicates successful regeneration (Saxena and Singh 1984).
Forest composition, community structure and diversity patterns are important ecological attributes often correlated with environmental and anthropogenic variables (Gairola et al. 2008; Ahmad et al. 2010). Numbers of seedlings, saplings and young trees are influenced by biotic and abiotic factors (Boring et al. 1981; Aksamit and Irving 1984).
The impediments in recruitment establishment, destruction of forest composition and its diversity result in changes in structural and functional aspect s of a forest ecosystem (Macdonald et al. 1989; Dhar et al. 1997). The protected area (PA) network in the Indian Himalayan region comprises of seven biosphere reserves, 25 national parks and 98 wildlife sanctuaries occupying 9.90 % area of the Indian Himalaya. Voluminous databases on existing biodiversity, especially on regeneration potential in most PAs in the Himalaya are not available. Without significant biological information, it is difficult to develop conservation plans for efficient management of PAs (Dhar et al. 1997). The present investigation in Lata–Tolma–Phagti (LTP), part of Nanda Devi Biosphere Reserve (NDBR) in the Himalaya focuses on (1) describing plant diversity (structural and compositional); (2) assessment of regeneration status with reference to seasonal recruitment pattern and, (3) predicting potential changes to forest structure/composition in future.
Materials and methods
Study area and site selection
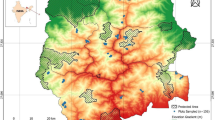
The NDBR, represents the west Himalayan highland (2b) province of the biogeographic zone-Himalaya and lies between 30°06′ and 31°04′ north latitude and 79°13′ and 80°17′ east longitude, covering 6407.03 km2 (core zone 712.12 km2; buffer zone 5148.57 km2; transition zone 546.34 km2). The site experiences three seasons, rainy (mid June–mid September), prolonged winter (late September–April) and summer (May–mid June). Average annual rainfall in the reserve ranges from 1500 to 2000 mm, of which more than half occurs during the rainy season (Adhikari et al. 1991; Rawal and Pangtey 1994) suggesting a strong monsoonic influence. September–November months are the driest of all. One representative site (i.e., Lata–Tolma–Phagti: LTP in Garhwal region) in the buffer zone of NDBR was selected as the intensive study site (Fig. 1). Surveys were conducted during 2010–2012 at this site.

Location of study site Lata–Tolma–Phagti (bottom right panel) in Nanda Devi Biosphere Reserve (Uttarakhand in Western Himalaya) (bottom left panel), India
Sampling and data collection
Thirty forest stands (plots) at LTP site were sampled in three vertical transects named Lata (Lata village–Lata Kharak, 2350–3900 m), Tolma (Suraithoda–Jhandidhar, 2400–3850 m), and Phagti (Pagrasu–Dhunadhar, 2350–3900 m). Details of locations (altitude, latitude and longitude) were recorded using hand-held global positioning system [GPS (Garmin model-12)]. Standard phytosociological methods were used to obtain quadrat data (Greig-Smith 1957; Misra 1968; Kershaw 1973; Muller-Dombois and Ellenberg 1974; Dhar et al. 1997). Trees, saplings and seedlings were enumerated in ten randomly placed (10 × 10 m) quadrat. Shrubs were counted in ten (5 × 5 m) quadrat and herbs were counted in twenty (1 × 1 m) quadrat in each stand. In general, in each quadrat circumference at breast height (CBH at 1.37 m from the ground) was recorded for all tree species. Based on this information, individuals were classified as tree >31 cm; sapling 11–30 cm; seedling <11 cm CBH. Individuals of tree species were grouped into six arbitrary CBH classes (A: <10; B: 11–30; C: 31–60; D: 61–120; E: >121 cm). The total number of individuals in each of these classes was recorded for each species in individual stands, and stand information was pooled to represent communities. Size classes A and B represented seedlings and saplings, respectively. Other classes (C–E) represented tree classes. Relatively density of species in a particular size class was calculated as a percentage of total number of individuals in all size classes. Species diversity index was computed using the Shannon-Wiener information function (Shannon and Weiner 1963).
Community structure and regeneration patterns
Seasonal investigation (Negi 1995) on population structure and regeneration pattern of all tree species was carried out during summer (May–June), rainy season (mid July–August) and winter (November–December) in 2010 and 2011. The 30 forest stands were grouped into eight representative forest communities distributed between 2450 and 3586 m a.s.l. for studying detailed population structure and to predict future compositional changes in parent communities.
Regeneration status of species was determined based on population size of seedlings and saplings (Khan et al. 1987; Shankar 2001; Bhuyan et al. 2003): good regeneration, if seedlings > saplings > trees; fair regeneration, if seedlings > or ≤ saplings ≤ trees; poor regeneration, if the species survives only in sapling stage, but seedlings are absent (saplings may be <, > or =trees). If a species is present only in tree form it is considered as not regenerating, while species having no trees but only seedlings is considered a ‘new’ species.
Results
Floristic diversity and demographic patterns
Community types, site representation and species importance (IVI) are listed in Table 1. In total, 248 plant species were recorded at LTP site in NDBR. Of these, most (72.6 %, 181 spp.) were herbs. Shrubs (44 spp.) constituted 18.1 % and trees (23 spp.) 9.3 % (Fig. 2). Taxonomically, of the total 248 species, 87 % were angiosperms, 3.2 % were gymnosperms and 9.9 % were pteridophytes.

Proportional distribution of species richness across different tree strata and life forms
Proportional distribution of individuals in three age classes (tree, sapling, and seedling) and life forms (tree, shrubs, and herbs) in the study area and representative sites is depicted in Fig. 2. LTP site was represented by 23 tree species. The overall population structure at site LTP is shown in Fig. 3. LTP site had more seedlings and significantly fewer (P < 0.05) individuals in older age classes. Girth class C (31–60 cm) showed lower establishment of trees as compared to girth class D (61–120 cm). The demographic profiles for different forests exhibited more or less similar patterns (Fig. 4).

Mean density–diameter distribution of trees in LTP (Lata–Tolma–Phagti) site. (A <10; B 11–30; C 31–60; D 61–120; E >121 cm)

Density–diameter distribution of trees in different forest communities in the LTP site. (A <10; B 11–30; C 31–60; D 61–120; E >121 cm)
Community diversity and distribution pattern
Quantitative ecological parameters for different forest communities are presented in Table 2. Considering the forest composition, Pinus wallichiana community (18 spp.) followed by B. utilis (10 spp.) and Abies spectabilis and A. pindrow (9 spp. each) exhibited relatively higher tree species richness. Lowest tree species richness was recorded in the Mixed Taxus wallichiana–A. pindrow and Cedrus deodara communities (4 spp. each). Maximum species richness of saplings (11 spp.) and seedlings (16 spp.) was also recorded in the P. wallichiana community.
Tree density ranged between 599 and 1211 indiv ha−1, with maximum density in Mixed T. wallichiana–A. pindrow community (1211 indiv ha−1) followed by A. caesium (960 indiv ha−1) and B. utilis (856 indiv ha−1). Sapling density ranged from 70 (Mixed J. regia–P. cornuta community) to 951 indiv ha−1 (A. pindrow community). Seedling density ranged from 470 indiv ha−1 (C. deodara community) to 1665 indiv ha−1 (A. pindrow community).
Tree diversity ranged from 0.63 (C. deodara community) to 1.61 (Mixed J. regia–-P. cornuta community). Sapling diversity was greatest in the A. spectabilis community (1.36) and lowest in the Mixed J. regia–P. cornuta and A. caesium–P. cornuta mixed community (0.76 each). P. wallichiana community (1.79) showed maximum diversity of seedlings and lowest seedling diversity was recorded in the Mixed A. caesium–P. cornuta community (0.35).
Regeneration status and seasonal behavior
Of the 23 tree species recorded at site LTP, 13.0 % showed good, 52.2 % fair, 26.1 % poor, 8.7 % no regeneration (Table 3). ANOVA revealed uneven variation in density values across seasons (Fig. 5). Starting from summer (2010), a significant (P < 0.05) increase in the number of seedlings was observed with the onset of rainy season (2010). Afterward, the seedling density gradually decreased in winter (2010) and summer (2011), and increased significantly in the next rainy season (2011). In 2010, average seedling density was about 315 indiv ha−1 in summer and reached 519 indiv ha−1 in rainy season. In winter 2010, gradual decrease in seedling density (410 indiv ha−1) was recorded. Similar trends were observed in 2011. A linear regression line showed gradual but non-significant (P > 0.05) increase in seedling density across years.

Average seasonal recruitment pattern in entire forest communities
Discussion
Compositional diversity
LTP site with over 248 plant species was low in species richness and less representative than other sites in NDBR (Rawat 2013). This can be attributed to the existence and dominance of coniferous forest communities (Singh and Singh 1992). In general, LTP site has 63 % coverage of conifer-dominated forests and the remaining 37 % includes broadleaf forests. These observations are comparable to those of previous studies (Joshi 2002; Joshi and Samant 2004). The explanations for this are lies in the fact that LTP site has more xeric (dry) climatic conditions. Further, the steep slopes at LTP site do not allow water retention in the soil, thereby giving rise to dry conditions that are more suitable for growth of coniferous communities (Joshi 2002). In general, coniferous communities are broadly reported to be species poor as compared to broadleaf communities (Singh and Singh 1992).
Mean tree density (599–1211 indiv ha−1) was comparable to values (320–1670 indiv ha−1) reported in earlier studies pertaining to low and high altitude forests of the west Himalaya (Bankoti et al. 1992; Joshi 2002; Joshi and Samant 2004; Gairola et al. 2008; Garkoti 2008). Tree densities corresponded closely with values recorded from same region one decade earlier (Joshi 2002). Mean seedling density (470–1665 indiv ha−1) was also comparable to that reported for earlier studies (Bankoti et al. 1992; Joshi 2002; Joshi and Samant 2004). The range of mean sapling density (40–951 indiv ha−1) falls within the lower range (40–6667 indiv ha−1) reported earlier (Bankoti et al. 1992; Joshi 2002; Joshi and Samant 2004; Gairola et al. 2008).
Current state of tree species richness (4–18 species) falls within the lower range reported earlier (24–42 species) from high elevation forests of the region. However, diversity values (0.63–1.61) are within the reported range (0.45–3.29) from the region (Bankoti et al. 1992; Adhikari et al. 1991; Joshi 2002; Gairola 2005; Gairola et al. 2008). The trends of species richness and diversity indicate forests at mid to higher elevations are more diverse than the lower altitude forests in the west Himalaya.
Current demographic profiles
The size class distribution gives a demographic profile of a stand which can indicate future prospects for target communities (Gairola 2005). In general, the forest communities showed greater accumulation of individual in seedlings and a significant decline towards sapling and tree size classes. This structure revealed that the conversion from seedling to sapling was not proportional due to greater seedling mortality during severe winters. Similar conclusions have been drawn by other workers (Khumbongmayum et al. 2006). Further, large scale extraction of biomass, particularly of selected species, has also been reported to cause structural changes in plant communities in the region and elsewhere (Spurr and Barnes 1980; Singh et al. 1996, 1997; Cairns and Moen 2004; Shrestha et al. 2007). Such disturbances exert profound effects on forest development by altering stand structure and release growing space, making the latter available for other species to occupy (Oliver and Larson 1990; Mishra et al. 2003, 2004; Gairola et al. 2008).
The lower density of the higher girth classes of trees in the region, as compared to intermediate or lower girth classes, can be attributed to the relatively high mortality of large canopy trees (Goff and West 1975; Lorimer et al. 2001). However, the lower density of the intermediate diameter class (31–60 cm) as compared to lower and higher girth classes can be explained as the result of selective felling of this girth class. Moreover, it can be interpreted as a result of low tree mortality and lower removal rates across the lower and higher girth classes (Saxena et al. 1984; West et al. 1981).
Seasonal regeneration pattern
Greater numbers of seedlings can be attributed to high seedling formation by dominant species (i.e., B. utilis, Taxus baccata subsp. wallichiana). The long-term survival of these seedlings was later confirmed by 2 years of seasonal investigation on seedling survival. If the trend toward high survival rates continues, the forest communities will have increased dominance of these species in future.
Germination of freshly dispersed seeds was high for most of the species during the rainy season. Lieberman and Li (1992) and Swaine et al. (1990) observed similar patterns in tropical dry forest at Pinkwae, Ghana. Adverse effects of soil moisture stress and unfavorable temperatures are typically responsible for reduction of seedling population during winter (Perira and Kozlowski 1977; Schulte and Marshall 1983; Khumbongmayum et al. 2006). The gradual decrease in recruitments in summer can be attributed to the anthropogenic pressure of lopping and grazing. Evolutionary history of grazing and environmental moisture or primary productivity interacts in determining species adaptations for tolerance or avoidance of herbivores and in community responses to grazing (Milchunas and Lauenroth 1993).
Expected changes in forests vis- a-vis representativeness
Following patterns across recruitment layers are noticeable for different communities:
-
Sapling layer Communities having high representation of dominant species: C. deodara, P. wallichiana, A. spectabilis, Mixed Taxus wallichiana–A. pindrow, B. utilis communities. Communities having high representation of co-dominant species: Mixed J. regia–P. cornurta, A. caesium–P. cornuta, A. pindrow communities.
-
Seedling layer Communities having high seedling representation of dominant species: C. deodara, Mixed J. regia–P. cornurta, P. wallichiana, A. spectabilis, Mixed Taxus wallichiana–A. pindrow, B. utilis communities. Communities having sufficient representation of both dominant and co-dominant species, and accompanied by high representation of other species as well: A. caesium–P. cornuta, and A. pindrow communities.
Therefore, based on above trends of seedlings and saplings various combinations and trends of communities can be suggested. For example, (i) the communities with greater representation of dominant species in both seedling and sapling stage would suggest further strengthening of dominant species; (ii) the communities with greater representation of both dominant and co-dominant in sapling and seedling layers would indicate the composition remains unchanged in future; (iii) the communities having greater proportion of seedling and saplings of co-dominant would indicate possible dominance of such species in future; (iv) the communities having greater representation of seedlings and saplings of the species other than the dominants and the co-dominants would indicate likely future changes in composition of target communities.
The demographic profiles of some of the dominant and some relatively less prominent tree species require attention. For example, in the case of three dominant species, B. utilis, P. wallichiana and Taxus baccata subsp. wallichiana in spite of their greater seedling numbers both were less prominently represented in sapling and tree layers. Certain relatively less prominent species like Malus baccata and Prunus persica, however, were represented only in tree layer suggesting that these species did not regenerate in the recent past and at present. Therefore, long-term persistence of such species is in question. Besides, Juglans regia, Picea smithiana, Pyrus pashia, Salix daphnoides and S. disperma were represented only in tree and seedling classes and require further monitoring.
Conclusions
The targeted sites in the study reflect richness and representativeness values of the reserve. The data sets available, and generated through this study, provide enough bases for establishing the LTP site in NDBR as potential sites for long-term ecological monitoring under various change scenarios. LTP site supported lower diversity of plant communities and species as compared to other sites in the reserve. Seasonal recruitment pattern showed successful establishment of seedlings that is indicative of possible changes in composition of communities in this site.
References
Adhikari BS, Rikhari HC, Rawat YS, Singh SP (1991) High altitude forests: composition diversity and profile structures in a part of Kumaun Himalaya. Trop Ecol 32:86–97
Ahmad I, Ahmad MSA, Hussain M, Ashraf M, Ashraf MY, Hameed M (2010) Spatiotemporal aspects of plant community structure in open scrub rangelands of submountainous Himalayan plateaus. Pak J Bot 42(5):3431–3440
Aksamit SE, Irving FD (1984) Prescribed burning for lowland black spruce regeneration in Northern Minnesota. Can J For Res 14:107–113
Baduni NP, Sharma CM (2001) Population structure and community analysis on different aspects of Sal savanna forest type in outer Garhwal Himalaya. Indian For 127(9):1001–1011
Bankoti NS, Rawal RS, Samant SS, Pangtey YPS (1992) Forest vegetation of inner hill ranges in Kumaun, Central Himalaya. Trop Ecol 33:41–53
Bhandari BS (2003) Blue pine (Pinus wallichiana) forest stands of Garhwal Himalaya: composition, population structure and diversity. J Trop For Sci 15(1):26–36
Bhuyan P, Khan ML, Tripathi RS (2003) Tree diversity and population structure in undisturbed and human-impacted stands of tropical wet evergreen forest in Arunachal Pradesh, Eastern Himalayas, India. Biodivers Conserv 12(8):1753–1773
Boring LR, Monk CD, Swank WT (1981) Early regeneration of a clear cut southern Appalachian forest. Ecology 62:1244–1253
Cairns DM, Moen J (2004) Herbivory influences tree lines. J Ecol 92:1019–1024
Criddle RS, Church JN, Smith BN, Hansen LD (2003) Fundamental causes of the global patterns of species range and richness. Russ J Plant Physl 50:192–199
Dhar U, Rawal RS, Samant SS (1997) Structural diversity and representativeness of forest vegetation in a protected area of Kumaun Himalaya, India: implications for conservation. Biodivers Conserv 6:1045–1062
Gairola S (2005) Assessment of diversity pattern in sub-alpine forests of west Himalaya: recruitment strategy, litterfall and nutrient return. Ph.D. thesis. India: HNB Garhwal University, Srinagar (Garhwal)
Gairola S, Rawal RS, Todaria NP (2008) Forest vegetation patterns along an altitudinal gradient in sub-alpine zone of west Himalaya, India. Afr J Plant Sci 2:42–48
Garkoti SC (2008) Estimates of biomass and primary productivity in a high-altitude maple forest of the west central Himalaya. Ecol Res 23:41–49
Goff FG, West DC (1975) Canopy-under storey interaction effects on forest population structure. For Sci 21:98–108
Greig-Smith P (1957) Quantitative plant ecology. Academic Press, New York, p 198
Joshi HC (2002) Assessment of habitat diversity, forest vegetation and human dependence in the buffer zone of Nanda Devi Biosphere Reserve of west Himalaya. Ph.D. theis. Nainital India: Kumaun University, p 535
Joshi HC, Samant SS (2004) Assessment of forest vegetation and conservation priorities of communities in part of Nanda Devi Biosphere Reserve, West Himalaya. Part I. Int J Sustain Dev World Ecol 11:326–336
Kershaw KA (1973) Quantitative and dynamic plant ecology, 2nd edn. Edward Arnold Limited, London, p 308
Khan ML, Rai JPN, Tripathi RS (1987) Population structure of some tree species in disturbed and protected sub-tropical forests of north-east India. Acta Oecol-Oec Appl 8(3):247–255
Kharkwal G (2009) Qualitative analysis of tree species in evergreen forests of Kumaun Himalaya, Uttarakhand, India. Afr J Plant Sci 3(3):49–52
Khumbongmayum AD, Khan ML, Tripathi RS (2006) Biodiversity conservation in sacred groves of Manipur, northeast India: population structure and regeneration status of woody species. Biodivers Conserv 15:2439–2456
Korner C (1998) A re-assessment of high elevation treeline positions and their explanation. Oecologia 115:445–459
Lieberman D, Li M (1992) Seedling recruitment patterns in a tropical dry forest in Ghana. J Veg Sci 3:375–382
Lorimer CG, Dahir SE, Nordhiem EV (2001) Tree mortality rates and longetivity in mature and old growth hemlock-hardwood forests. J Ecol 89:960–970
Macdonald IAW, Loope LL, Usher MB, Hamann O (1989) Wildlife conservation and the invasion of nature reserves by introduced species: a global perspective. In: Drake JA, Mooney HA, di Castri F, Groves RH, Kruger FJ, Rejmanek M, Williamson M (eds) Biological invasions: a global perspective. SCOPE, Wiley, pp 281–300
Milchunas DG, Lauenroth WK (1993) Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecol Monogr 63:327–366
Mishra BP, Tripathi RS, Tripathi OP, Pandey HN (2003) Effects of disturbance on the regeneration of four dominant and economically important woody species in a broad-leaved subtropical humid forest of Meghalaya, north east India. Curr Sci 84(11):1449–1453
Mishra BP, Tripathi OP, Tripathi RS, Pandey HN (2004) Effect of anthropogenic disturbance on plant diversity and community structure of a sacred grove in Meghalaya, northeast India. Biodivers Conserv 13:421–436
Misra R (1968) Ecological work book. Oxford & IBH Publishing Company, Calcutta, p 242
Muller-Dombois D, Ellenberg H (1974) Aims and methods of vegetation ecology. Wiley, New York, p 547
Negi SS (1995) Uttarakhand: land and people. MD Publications Pvt Ltd, Delhi, p 149
Oliver CD, Larson BC (1990) Forest Stand dynamics. Government document. McGraw Hill, Inc, New York, p 467
Perira JS, Kozlwski TT (1977) Water relations and drought resistance or young Pinus banksiana and Pinus resinosa plantation trees. Can J For Res 7:132–137
Rao PB (1988) Effects or environmental factors on germination and seedling growth in Quercus floribunda and Cupressus torulosa, tree species of central Himalaya. Ann Bot 61:531–540
Rawal RS, Pangtey YPS (1994) Distribution and structural-functional attributes of trees in the high altitude zone of central Himalaya, India. Vegetatio 112:29–34
Rawat B (2013) Changes in vegetation diversity and plant responses in Nanda Devi Biosphere Reserve over the last two decades. Ph.D. thesis. Kumaun University, Nainital, Uttarakhand, India, p 179
Saxena AK, Singh JS (1984) Tree population structure of certain Himalayan forests and implications concerning the future composition. Vegetatio 58:61–69
Saxena AK, Singh SP, Singh JS (1984) Population structure of forests of Kumaun Himalaya: implications for management. J Environ Manag 19:307–324
Schulte PJ, Marshall PE (1983) Growth and water relations of black locust and Pine seedlings exposed to controlled water-stress. Can J For Res 13:334–338
Shankar U (2001) A case study of high tree diversity in a sa1 (Shorea robusta)—dominated lowland forest of Eastern Himalaya: floristic composition, regeneration and conservation. Curr Sci 81:776–786
Shannon CZ, Weiner W (1963) The mathematical theory of communications. University of Illinois Press, Urbane
Shrestha BB, Ghimire B, Lekhak HD, Jha PK (2007) Regeneration of treeline Birch (Betual utilis D. Don) forest in a trans-Himalayan dry valley in central Nepal. Mt Res Dev 27:259–267
Singh JS, Singh SP (1992) Forests of the Himalaya. Structure, functioning and impact of man, Nainital, India: Gyanodaya Prakashan. Fine Art Press, Delhi, p 295
Singh SP, Adhikari BS, Garkoti SC, Rawat YS (1996) Structural and functional characteristics of the forest ecosystems around Nanda Devi Biosphere Reserve. In: Ramakrishnan PS, Purohit AN, Saxena KG, Rao KS, Maikhuri RK (eds) Conservation and management of biological resources in Himalaya. Oxford & IBH Publishing Co. Ltd, New Delhi, pp 413–432
Singh SP, Rawat YS, Garkoti SC (1997) Failure of brown oak (Quercus semecarpifolia) to regenerate in central Himalaya: a case of environmental semisurprise. Curr Sci 73:371–374
Spurr SH, Barnes BV (1980) Forest ecology. Wiley, New York, p 687
Swaine MD, Lieberman D, Hall JB (1990) Structure and dynamics in a tropical dry forest in Ghana. Vegetatio 88:31–51
Todaria NP, Pokhriyal P, Uniyal P, Chauhan DS (2010) Regeneration status of tree species in forest of Phakot and Pathri Rao watersheds in Garhwal Himalaya. Curr Sci 98(2):171–175
West DC, Shugart HH Jr, Ranny JW (1981) Population structure of forest over a large area. Forest Sci 27:701–710
Acknowledgments
Authors are grateful to the Director of the Institute for providing the necessary facilities to carry out this work. We are thankful to In-House Project-08 (2007–2012) for providing the financial facilities throughout the study period. Sincere thanks goes to DST SB/YS/LS-205/2013 and CSIR (09/560 (0015)/2011-EMRI), Govt. of India for the financial support. We also thank the local inhabitants for their generous help during extensive field visits.
Author information
Authors and Affiliations
Corresponding author
Additional information
Project funding: This study was supported by DST SB/YS/LS-205/2013 and CSIR (09/560 (0015)/2011-EMRI), Govt. of India.
The online version is available at http://www.springerlink.com
Corresponding editor: Zhu Hong
Rights and permissions
About this article

Cite this article
Rawat, B., Gairola, S., Chandra Sekar, K. et al. The present status and future prospects of forest vegetation in part of Nanda Devi Biosphere Reserve (a World Heritage Site), India. J. For. Res. 26, 897–904 (2015). https://doi.org/10.1007/s11676-015-0109-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11676-015-0109-x