
Abstract
Monochasma savatieri Franch. ex Maxim is a valuable medicinal plant that is currently threatened due to human over excavation. Therefore, an efficient plant micropropagation protocol for M. savatieri through seed germination and direct shoot regeneration is described. The effects of different seed germination treatments, explant source on shoot induction, plant growth regulators on shoot multiplication, and survival rate of regenerated plants were examined. The highest M. savatieri seed germination rate was over 90% on Murashige and Skoog (MS) medium supplemented with 0.1 mg L−1 6-benzylaminopurine (6-BA) and 0.01 mg L−1 α-naphthaleneacetic acid (NAA), with the survival rate reaching 86.0% one-mo after transplanting to soil. Direct shoot proliferation was achieved with two explant types, but the use of nodal stem explants gave best results in comparison to shoot tip explants regarding shoot multiplication frequency and average number of shoots obtained per explant. For nodal segments, medium supplemented with 0.5 mg L−1 6-BA and 0.2 mg L−1 NAA gave the highest multiplication frequency of 90.0%. Among the various treatments tested for shoot multiplication, the best response occurred on MS medium containing 0.5 mg L−1 6-BA and 0.5 mg L−1 gibberellin (GA), with the number of shoots per explant reaching 27.6 and the average height of 4.25 cm. To obtain thick, elongated, and good quality buds, 6-BA at 0.1 and 0.5 mg L−1 along with 0.4 mg L−1 GA should be used interchangeably. Four-mo after transplantation, the survival rates were significantly affected by the types and concentrations of plant growth regulators used during plant tissue culture. The most effective combination of plant growth regulators 0.2 mg L−1 was 6-BA and 0.5 g L−1 ABT (mixture composed of 20% α-naphthaleneacetic acid content and 30% indole-3-acetic acid as active ingredients) with a survival rate of 85.5%. The in vitro propagation protocol for M. savatieri reported here will be useful for the rapid and large-scale propagation of this species.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Monochasma savatieri Franch. ex Maxim is a perennial herb that belongs to the Scrophulariaceae family and only grows in the Zhejiang, Fujian, Jiangxi, and other Southern Provinces in China and Japan, on sunny hillsides and under Pinus massoniana Lamb. forests. It is a semi-parasitic plant with alternated double leaves, which are long round lanceolate to linear lanceolate, that grow to a length of 15 to 23 cm and are silvery white due to the presence of wooly trichomes (Zhang et al. 2015). This plant is used for treating a number of diseases in Traditional Chinese Medicine including the common cold, cough, and menoxenia, toothache (Fu et al. 2010; Shi et al. 2013). Recently, it has been reported that M. savatieri contains chemical constituents, such as glycoside, chlorogenic acid, alkaloids, saponins, and anthraquinone (Li et al. 2012; Zheng et al. 2012), which possess antimicrobial, anti-inflammatory, and antioxidant activities (Fu et al. 2008; Liu et al. 2013; Shi et al. 2013; Gao et al. 2017). At present, the pharmaceutical industry still relies on naturally occurring M. savatieri, of which the existing populations have decreased sharply due to the destruction of habitat and overexploitation (Zhang et al. 2015) which does not meet the market demand.
To strengthen its natural conservation, development, and utilization of plant resources, there is an urgent need for an efficient in vitro micropropagation and plant regeneration protocol. Micropropagation has the advantage of rapidly increasing the number of plants, as well as allowing for the maintenance of both an in vitro and in vivo germplasm collection and is a critical tool for basic and applied plant research (Chen et al. 2016a; Kulus 2020). Previous studies published on the tissue culture propagation of M. savatieri showed that adventitious shoots could be induced from stem and root explants, but there was no survival of regenerated plants up to several months after transplantation even with haustoria to obtain nutrients from a host plant (Yang 2009; Zhang et al. 2017; Wang and Qiu 2018a, b). In the present study, we established a rapid propagation protocol for M. savatieri through seed germination and direct shoot regeneration and report for the first time the successful plant survivial ex vitro without haustorial anatomy and capable of completing their life cycle. This study lays a foundation for large-scale in vitro propagation of M. savatieri which is of great significance for the commercial supply of this species.
Materials and Methods
Explant preparation and surface sterilization - Seed collection
In May 2016, mature uncracked M. savatieri capsules were collected on top of the hillside in Shuichuan village, Fenyi County in Jiangxi Province, China. According to the position on the branch, each capsule was divided into an upper, middle, and lower parts. All seeds from each part of the capsule were quickly and separately detached after collection. Gravity separation was used to float the wizened seeds away so the remaining seeds were thoroughly washed with a 0.1% (w/v) Tween and Carbendazim solution for 30 min. Various sterilization methods and times were tested to establish an aseptic culture system (Table 1). The surface-sterilized seeds were washed three and five times with sterile distilled water after being treated with 75% (v/v) alcohol and mercuric chloride (HgCl2), respectively, before placing on Murashige and Skoog (MS; Murashige and Skoog 1962) medium containing 0.1 mg L−1 6-BA and 0.01 mg L−1 NAA for germination (Yang 2009). After adding all media components, the pH was adjusted to 5.8 ± 0.1 (with 0.1 M HCl and/or 0.1 M NaOH) before the addition of 0.58% (w/v) agar (Dingguo Changsheng Biotechnology CO. LTD, China) and boiled to dissolve the agar. The medium was dispensed as 30-mL aliquots into 200-mL glass jars and sealed with plastic caps, prior to autoclaving at 121°C and 105 kpa for 17 min (autoclave SMS ASVE, Guangzhou, China). All cultures were incubated at 25 ± 2°C for an 8-h photoperiod with a density of 40 μmol m−2 s−1, which was provided by cool white fluorescent lamps (12w; Phillips, China).
Explant material preparation
In April 2017, stems were collected from plants at the same location stated above. Stem cuttings were placed into 0.1% (v/v) dishwashing liquid (diluted with water) for 2 min, after which the surface of the segment and leaf was finely brushed under tap water to remove the surface dirt. Stems were cleared of leaves, after which they were cut into 2-cm-long pieces. Because the stem surface is densely covered with glandular and cotton hairs to which dust and bacteria easily adhere, it is necessary to thoroughly sterilize each explant. Ten different sterilization procedures were tested, the details of which are presented in Table 2. Each procedure involved 18 to 21 pots with one explant per pot.
After freshly cutting small nodal segments (1 to 2 cm), sterile explants were inoculated on Murashige and Skoog (MS) (Murashige and Skoog 1962) medium supplemented with 2.5 mg L−1 6-BA, 0.2 mg L−1 NAA, and 3% (w/v) sucrose (Wang and Qiu 2018b), solidified with 0.2% (w/v) gelrite (Sigma-Aldrich, St. Louis, MO). All cultures were maintained at 25 ± 2°C and cultured under a 12-h photoperiod with a light intensity of 40 μmol m−2 s−1, which was provided by cool white fluorescent lamps (12w; Phillips, Beijing, China). After new shoots were induced, shoots were separated and transferred to fresh MS basal medium as described above at a 3-wk interval. Shoot tips and nodal stems from successive secondary cultures (3 to 4 cycles) were excised and used as a source of explants.
Shoot induction and multiplication
MS medium supplemented with various concentrations of 6-BA and NAA was used for shoot multiplication (Table 3). To compare the shoot induction ability of different explants, one to two cm long shoot tip and nodal stem segments were tested. The culture conditions were the same as those used for the shoot induction of field grown plants. Each treatment was replicated four times with 10 to 14 explants per replicated treatment. The time elapsed until buds breaking and the multiplication frequency (number shoots per explant) was recorded.
Focus was placed on nodal stem segments due to a higher in vitro multiplication frequency and number of shoots. Healthy in vitro shoots were excised, cut into nodal stem segments (1.5 to 2.0 cm long with two leaves), and placed on MS medium supplemented with various hormone combinations (Table 4). To quantify the optimal shoot induction protocol, the number of shoots per explant, micro-shoot length, and shoot characteristics and quality were recorded after three cycles of subculture at a three-wk interval. Each treatment was replicated four times (one bottle for each replicated treatment with 10 explants per bottle).
Rooting and acclimatization
To obtain hardened plantlets, isolated shoots (1.5 to 2.5 cm long) were excised from adventitious shoots and subcultured on MS medium supplemented with 0.1 mg L−1 6-BA and 0.04 mg L−1 NAA for 2 wk, then transferred to medium with different plant growth regulators (PGRs, see Table 5 for PGRs). For acclimatization, plantlets (subcultured on medium with PGRs in Table 5) were transferred to a climate-controlled greenhouse set to 27 ± 2°C, 60 to 70% relative humidity, and natural light and photoperiod (10 to 12 h). Initially, both rooted (aerial root) and unrooted plantlets were carefully removed from the medium and gently washed to remove any medium adhered to the base of the stem under running water. Subsequently, rooted plantlets were transferred to a tray with sterile mixed media (2 peat/2 rotten bark/2 carbonated husk/3 soil), in which the bark decayed more than one-y and husk was heated to carbonization in insufficient air. Unrooted plantlets were quickly dipping in rooting agent consisting of 0.2 mg L−1 6-BA and 0.5 mg L−1 ABT (mixture composed of 20% α-naphthaleneacetic acid content and 30% indole-3-acetic acid as active ingredients; Beijing ai-bidi Biotechonology CO. LTD, China) for 30 s before transplanting. Plantlets were covered using transparent plastic and maintained in an environment with semi-controlled light and temperature conditions. The presence of fungi was controlled with Carbendazim sprayings and after 20-d plastic mulch was opened gradually over a two-d period. Percent plant survival and plant morphology were recorded four-mo after transplantation.
Analysis
A single factor experimental design with three repetitions per treatment and four vessels per repetition was adopted for statistical analysis. The data were analyzed by one-way analysis of variance (ANOVA) and a Tukey’s multiple comparison test at a probability level of 0.05 using SPSS v.18.0 (IBM Chicago, IL).
Results
Seed germination
M. savatieri seed germination in vitro showed no signs of microbial contamination or effect of sterilization agent on germination rate (data not shown). Any differences in seed germination rate are therefore likely to be due to seed dormancy and capsule/seed position/maturation stage. Under all conditions, the germination rate of seeds treated with GA was slightly higher and the germination time was earlier than that of untreated seeds. Furthermore, the germination rate of seeds from the upper part of the capsule was reduced when compared with those from the lower part of the capsule (Table 1). The highest germination rate of M. savatieri seeds was over 90% on MS medium with 0.1 mg L−1 6-BA and 0.01 mg L−1 NAA under a 16-h photoperiod with a light intensity of 40 μmol m−2 s−1. The earliest germination started after seven-d and the germination rate was between 54.25 and 91.50%. Cotyledons emerged after two-wk, and seedlings with a height of 1.0 to 1.5 cm, but still unrooted, were obtained after three-wk (Fig. 1b, c). Complete seedlings were obtained after subcultured on MS medium without any PGRs, and the survival rate reached 86% 1 mo after transplantation (Fig. 1d, e). The detailed results are presented in Table 1 and Fig. 1.

In vitro Monochasma savatieri Franch. ex Maxim seed germination. (a) Seeds turning green after seven-d in vitro. (b) Cotyledons appeared after two-wk. (c) Unrooted seedlings obtained after three-wk. (d) Seedlings with roots after four-wk. (e) Four wk after transplantation of seedlings. Bars 1 cm.
Surface sterilization
Various treatments for explant sterilization were tested. Extending carbendazim and mercuric chloride treatment time resulted in reduced contamination and increased survival rate. The synergistic effect of a lower concentration of both penicillin and streptomycin improved effectiveness over the use of a single antibiotic at a high concentration. Among the ten different treatments tested, T9 (1000 mg L−1 carbendazim for 30 min, 75% ethanol for 20 min, 300 mg L−1 streptocycline for 10 min, 300 mg L−1 penicillin for 10 min, and 1000 mg·L−1 mercuric chloride for five-min) and T10 (1000 mg·L−1 carbendazim for 30 min, 300 mg L−1 streptocycline for 10 min, 300 mg L−1 penicillin for 10 min, and 1000 mg L−1 mercuric chloride for seven-min) were found to be the most effective, having a contamination percentage and survival rate of 16.67% and 55.56%, 27.78% and 66.67%, respectively. The detailed results are presented in Table 2.
Shoot induction and multiplication
Shoot and bud multiplication treatments were evaluated to improve micropropagation of M. savatieri. Approximately 10% of the shoot tips and 60% of the nodal stem explants produced new buds when cultured for four-wk on MS medium containing various combinations of 6-BA and NAA. After several cycles of cultivation, the new buds were divided into different explants (nodal stem and shoot tip), which were then used for shoot/bud multiplication experiments.
Out of these two different types of tissues, nodal stem explants gave better results compared to shoot tips in both shoot multiplication frequency and average number of shoots per explant. Shoot tip explants grown on medium containing 0.5 mg L−1 6-BA and 0.4 mg L−1 NAA required 18-d to bud break with a shoot multiplication frequency of 23.75%. By comparison, nodal segments cultured on medium supplemented with 0.5 mg L−1 6-BA and 0.2 mg L−1 NAA gave the highest multiplication frequency of 90.00% with only eight-d required to bud break. The reason for the bud break delay with shoot tips is because when it grew to a certain height (approximately five cm), and if the tip is dead or damaged, axillary buds will be induced, making the time needed for bud break longer than for nodal segments. Furthermore, when cultured on medium supplemented with 0.5 mg L−1 6-BA and 0.2 to 0.4 mg L−1 NAA, shoots induced from nodal stem explants were larger and stronger than those formed from shoot tip explants. The detailed results are presented in Table 3.
The mean number and quality of induced shoots were significantly affected by the concentration of PGRs in the medium (Table 4) where 0.5 mg L−1 6-BA induced more buds than 0.1 mg L−1 6-BA. A further increase of the 6-BA concentration inhibited bud induction with many buds becoming hyperhydric (Fig. 2a). When 0.2 to 0.4 mg L−1 GA was added to medium containing 0.5 mg L−1 6-BA, good quality buds were formed that elongated noticeably (Fig. 2b). However, when shoots lacking apical buds were subcultured for more than two cycles onto medium containing 0.5 mg L−1 6-BA, shoots became slender and fragile. To overcome this problem, continued sub-culturing on medium containing 0.1 mg L−1 6-BA induced thicker, elongated, and good quality buds. Aerial roots were induced when cultured on medium containing auxin, both NAA (1.0 mg L−1) and indoleacetic acid (IAA, 0.5 mg L−1) (Fig. 2c, d). When the culture period was extended to five-wk, shoot and leaves at the top became yellowish, and eventually, the whole plant died.

Multiple shoot architectures of Monochasma savatieri Franch. ex Maxim under different in vitro plant growth regulator treatments. (a) Slightly hyperhydric adventitious buds induced from the base of the stem when cultured on medium with 1.0 mg L−1 to 2.0 mg L−1 6-BA. (b) Strong and elongated adventitious buds induced on the medium supplemented with 0.5 mg L−1 6-BA and 0.4 mg L−1 GA. (c) Aerial roots, cluster, and slender buds induced on MS medium containing 0.5 mg L−1 6-BA and 1.0 mg L−1 NAA. (d) Low number of aerial roots and strong buds formed on medium with 0.5 mg L−1 6-BA and 0.5 mg L−1 IAA. Bars 1 cm. 6-BA 6-benzyladenine; GA gibberellin; NAA α-n acid; IAA indoleacetic acid.
Rooting and acclimatization
Plant survival was evaluated after transplanting to soil substrate. Based on previous experimental results, a low number (two to three) and frequency (5 to 10%) of roots or aerial roots were induced from regenerated plants on agar medium (Fig. 3c, d). When plantlets were directly transplanted to mixed media (2 peat/2 bark/2 husk/3 soil), the survival rate was 0% 30 d after transplantation. However, when apical buds (2-cm top segments containing apical buds) were first transferred to MS basal medium supplemented with 0.2 mg L−1 6-BA and different auxins (NAA and ABT; Table 5) for two-wk (Fig. 3b), they were transplanted to soil, and the survival rate was more than 90% 1 mo after transplantation in all treatments tested (Fig. 3e). With the extension of transplanting time (up to four-mo), survival rates decreased and were significantly affected by the types and concentrations of plant growth regulators used during plant tissue culture.

. In vitro Monochasma savatieri Franch. ex Maxim seedling production and acclimatization/planting. (a) Selection of in vitro seedlings. (b) Shoots cultured for 25 to 30 d were transferred for acclimatization. (c) Roots induced on medium were low in number (two to three roots per shoot) and frequency (5% to 10%). (d) Many aerial roots grew above the agar medium surface. (e) Seedlings without roots cultured in light substrate (1 carbonated husk/3 sawdust/2 rotten bark/3 peat/1 perlite) or mixed substrate with yellow soil (2 peat/2 rotten bark/2 carbonated husk/3 soil). (f) Seedlings grew into whole plants 30 d after transplantation and the plants grew to approximately 8 cm in height two-mo after transplantation. (g) In November, new buds sprouted at the base of tissue culture seedlings. (h) Plantlets flowered in the second y after transplantation. Bars 1 cm.
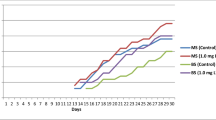
We found that applying the auxin ABT in tissue culture was more effective at promoting prolonged survival after transplantation than NAA. The survival rate decreased (45.5%) with use of an increasing concentration (0.4 to 1.0 mg L−1) of NAA, with ABT showing the opposite trend (Table 5). The most effective combination of plant growth regulators was 0.2 mg L−1 6-BA and 0.5 g L−1 ABT with a survival rate of 85.5% (Table 5).
Roots emerged at the cut ends of shoots, with a well-developed root system after four-wk of culture (Fig. 3e). Plant height reached around eight-cm approximately 2 mo after transplantation (Fig. 3f). Moreover, new buds sprouted from the plant base with the death of above ground parts four-mo after transplantation (Fig. 3g). In the spring of second year after transplantation, plants flowered both on light substrate (1 carbonated husk/3 sawdust/2 rotten bark/3 peat/1 perlite) and mixed substrate with soil (2 peat/2 rotten bark/2 carbonated husk/3 soil) (Fig. 3h).
Discussion
Seed germination
In vitro germination and growth of M. savatieri were established quickly and with relative ease. However, direct soil germination was not successful, although 30% of seeds did germinate, less than one percent of seedlings could grow into plants (data not shown). This is likely because it is a semi-parasitic plant at the seedling stage in its natural environment (Zhang et al. 2015; Zhang et al. 2017). To overcome this problem, whole plants were grown through aseptic seed culture with a germination rate of over 90%, which exceeded a previously reported rate by Yang (2009), who obtained a 67.5% and 86.9% germination rate via treatment with cold stratification and 200 mg L−1 GA soaking (48 h), respectively. This may also be related to the use of newly collected seeds in this study, which have an improved germinating rate.
We found that GA slightly promoted the germination of newly harvested seeds, similar to what Yang et al. (2009) found in M. savatieri and other medical plants (Ou et al. 2010; Chen et al. 2016b). This could be due to GA-mediated cell expansion which initiated the seed-to-seedling transition (Stamm et al. 2017). However, the germination rate of mature seeds stored at 4°C for more than two-wk decreased significantly, even reached 0%, and the reason for which remains unknown.
Explant type and PGR concentration affect in vitro shoot formation
The successful choice of explant is based on its physiological state, which determines whether it remains quiescent or starts actively dividing in vitro. A single non-zygotic plant cell has the unique ability to recover an entire organism through its totipotency. Plants produced by direct organogenesis show higher genetic fidelity than those regenerated by indirect organogenesis (with an intermediate callus stage), and the latter may lead to higher rates of somaclonal variation (Skała et al. 2015; Pérez-Alonso et al. 2018; Bhusare et al. 2018). In the Scrophulariaceae family, direct organogenesis of medicinal herbs has been previously reported (Jitendra et al. 2013; Martínez-Bonfil et al. 2011). To the best of our knowledge, this is the first report of an efficient in vitro propagation method of M. savatieri through direct organogenesis from nodal stem segments and apical meristems. We found the potential for in vitro shoot induction of nodal stem segments to be significantly higher than that of shoot tips. This may be due to shoot tips containing apical meristem that exerts strong apical dominance and inhibits bud development (Raja and Arockiasamy 2008). Similar results have also been reported in Morinda officinalis (Rubiaceae; Deng et al. 2015).
Cytokinins play a key role in the in vitro shoot induction of a number of plants within the Scrophulariaceae family, which includes some well-known medicinal herbs. For each species, a different combination of the choice of cytokinin and the type of explant yields the best results. In Digitalis purpurea L., for example, shoots were induced from nodal, leaf, and petiole segments at decreasing efficiency, with 6-BA giving higher induction than thidiazuron (TDZ) and KIN (Jitendra et al. 2013; Bhusare et al. 2018). In M. savatieri, the cytokinin TDZ gave the highest multiple shoot induction when nodal segments were used (Zhang et al. 2017). However, in Castilleja tenuiflora Benth., KIN was the most effective cytokinin in comparison to TDZ and 6-BA (Martínez-Bonfil et al. 2011). In the current study, the best shoot multiplication rates were obtained on MS medium supplemented with 6-BA (Table 2; Fig. 2), similar to what was found for Rosa canina L. as reported by Pahnekolayi et al. (2015). Furthermore, we found that the use of naphthaleneacetic acid (NAA) and IAA reduced the number of shoots, but that a combination of NAA and 6-BA positively influenced shoot hardening. Results opposite to this finding were obtained in the species Anthurium andreanum cv. Nitta., where addition of NAA to a medium that contained 6-BA effectively induced shoots (Islam et al. 2010). Although the combination of 6-BA and NAA inhibited shoot differentiation, it promoted shoot elongation in Anthurium andraeanum Lind. cv. Nicoya (Liendo and Mogollón 2009).
Ex vitro rooting
Zhang et al. (2017) observed that M. savatieri roots may be hydrophobic and succeeded in obtaining a high survival rate of transplanted plants by using vermiculite as a substitute for agar. Furthermore, when co-cultured with cuttings of hosts Kuhnia rosmarinifolia and Lonicera japonica, M. savatieri plantlets could survive for a few months (October to May) in the South China Botanical Garden (SCBG), Guangzhou, Guangdong province, possibly because Guangzhou is not its natural habitat. In the present study, the process was simplified, in which rooting in medium was eliminated, and the rootless tissue culture plantlets were directly transplanted into light substrate (1 carbonated husk/3 sawdust/2 rotten bark/3 peat/1 perlite) and mixed substrate with soil (2 peat/2 rotten bark/2 carbonated husk/3 soil). The survival rate of transplanting was more than 90% while controlling the appearance of fungi. Their ex vitro adaptation revealed the relative ease of acclimatization of M. savatieri plantlets to surviving in the environment of artificial control with the temperature that ranged from 25 to 32°C and 70% average daily relative humidity. Previous studies reported that M. savatieri is a semi-parasitic plant in terms of host range and haustorial connection and parasitizing up to 27 species from 25 families that constitute shrubs and herbs with multi-branched and shallow root systems. The maximum number of acceptable hosts was determined by genetics and tissue incompatibility (Zhang et al. 2015), with the host range depending on the relationship between geography of host distribution and ecology, particularly diffusion biology and environment factors (Joel et al. 2013). However, in this study, we found that plantlets could survive in Fenyi, Jiangxi province for the entire year, without parasitic host co-culture and were blooming in the second year after transplantation. This protocol avoids root induction in vitro where the agar was replaced by vermiculite media and reduces transplantation of regenerated plantlets with roots, thus eliminating the possibility of damaging roots and improving the survival of transplanted plantlets (Table 5). The survival rate after four-mo transplantation was higher (85.5%) than that reported by Zhang et al. (2017). The successful survival of plantlets without host shows that M. savatieri can also obtain all of the water and nutrition required to complete its life cycle under non-parasitic conditions via its own root system. This was also confirmed by the anatomical structure of mature haustoria with some of the fine rootlets terminating in haustoria (Zhang et al. 2015). Therefore, in the future, to optimize the cultivation process, an appropriate host could be selected to promote M. savatieri growth and yield. In summary, our study provides an effective method to micropropagate M. savatieri and shows that it is possible to do so without using any hosts for co-culture.
Change history
19 February 2021
A Correction to this paper has been published: https://doi.org/10.1007/s11627-020-10153-1
References
Bhusare BP, John CK, Bhatt VP, Nikam TD (2018) In vitro propagation of, Digitalis lanata, Ehrh. Through direct shoot regeneration- a source of cardiotonic glycosides. Ind Crop Prod 121:313–319. https://doi.org/10.1016/j.indcrop.2018.05.019
Chen QL, Wang HL, Wang ZF, Shan CG, Zhai ZX, Guo YH (2016a) Effects of cold stratification and exogenous gibberellic on seed germination and contents of endogenous gibberellins and abscisic acid in Cistanche deserticola Y. C. Ma. Plant Physiol J 45:270–272 (in Chinese)
Chen YL, Zhang YY, Cheng QW, Niu MY, Liang HZ, Yan HF, Zhang XH, Jaime A, Ma GH (2016b) Plant regeneration via direct and callus-mediated organogenesis from leaf explants of Chirita swinglei(Merr.) W. T. Wang. In Vitro Cell Dev Biol - Plant 52:521–529. https://doi.org/10.1007/s11627-016-9766-5
Deng ZC, Jin H, He H (2015) An efficient micropropagation system for Morinda officinalis How. (Rubiaceae), an endangered medicinal plant. J Agr Sci Tech 17:1609–1618
Fu G, Pang H, Wong Y (2008) Naturally occurring phenylethanoid glycosides: potential leads for new therapeutics. Curr Med Chem 15:2592–2613. https://doi.org/10.2174/092986708785908996
Fu JX, Wang LX, Wu J (2010) Inflammation Yinning particle treatment of acute upper respiratory tract infection clinical observation. Strait Pharm J 22:171–173 CNKI:SUN:HAIX. 0.2010–10-099 (in Chinese)
Gao H, Cui Y, Kang N, Liu X, Liu Y, Zou Y, Chen X (2017) Isoacteoside, a dihydroxyphenylethyl glycoside, exhibits anti-inflammatory effects through blocking toll-like receptor 4 dimerization. Br J Pharmacol 174:2880–2896. https://doi.org/10.1111/bph.13912
Islam SA, Dewan M, Mukul M, Hossain KF (2010) In vitro regeneration of Anthurium andreanum cv. Nitta Bangladesh J Agril Res 35:217–226. https://doi.org/10.3329/bjar.v35i2.5884
Jitendra GP, Mahendra LA, Kirti MK, Sayantan P, Vijay PB, Pola-varapu BKK, Tukaram DN (2013) In vitro propagation and production of cardiotonic glycosides in shoot cultures of Digitalis purpurea L. by elicitation and precursor feeding. Appl Microbiol Biotechnol 97:2379–2393. https://doi.org/10.1007/s00253-012-4489-y
Joel MD, Gressel J, Musselman JL (2013) Parasitic Orobanchaceae:parasitic mechanisms and control strategies. Springer, Berlin
Kulus D (2020) Influence of growth regulators on the development, quality, and physiological state of in vitro-propagated Lamprocapnos spectabilis (L.) Fukuhara. In Vitro Cell Dev Biol -- Plant https://doi.org/10.1007/s11627-020-10064-1
Li M, Shi MF, Liu YL, Xu QM, Yang SL (2012) Phenylethanoid glycosides from Monochasma savatieri and their anticomplement activity through the classical pathway. Plant Med 78:1381–1386. https://doi.org/10.1055/s-0032-1314982
Liendo M, Mogollón N (2009) Multiplicación clonal in vitro del anturio (Anthurium andraeanum Lind. cv. Nicoya). Bioagro 21:179–182 http://www.redalyc.org/articulo.oa?id=85714207005. Accessed 24 Mar 2020
Liu YL, He WJ, Mo L, Shi MF, Zhu YY, Pan S, Li XR, Xu QM, Yang SL (2013) Antimicrobial, anti-inflammatory activities and toxicology of phenylethanoid glycosides from Monochasma savatieri Franch. ex Maxim. J Environ Sci 149:431–437. https://doi.org/10.1016/j.jep.2013.06.042
Martínez-Bonfil BP, Salcedo-Morales G, López-Laredo AR, Ventura-Zapata E, Evangelista-Lozano S, Trejo-Tapia G (2011) Shoot regeneration and determination of iridoid levels in the medicinal plant Castilleja tenuiflora Benth. Plant Cell Tiss Org Cult 107:195–203. https://doi.org/10.1007/s11240-011-9970-2
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol Plant 15:473–497. https://doi.org/10.1111/j.1399-3054.1962.tb08052.x
Ou Y, Duan LD, Wang FY, Qian FM, Zheng X, Zhou SX, Yi JY (2010) Effect of stratification and GA3 joint treatment on the germination of Mosla chinensis Maxim. Seed. Med Plant 1:44–46 CNKI:SUN:MDPT.0.2010-11-018 (in Chinese)
Pahnekolayi MD, Tehranifar A, Samiei L, Shoor M (2015) Micropropagation of Rosa canina through axillary shoot proliferation. J Ornament Plant 4:45–51
Pérez-Alonso N, Martin R, Capote A, Pérez A, Hernández-Díazb E, Rojasa L, Quialad E, Angenone G, Garcia-Gonzalesf R, Chong-Pérez B (2018) Efficient direct shoot organogenesis, genetic stability and secondary metabolite production of micropropagated Digitalis purpurea L. Ind Crop Prod 116:259–266
Raja HD, Arockiasamy DI (2008) In vitro propagation of Mentha viridis L. from nodal and shoot tip explants. Plant Tiss Cult Biotech 18:1–6. https://doi.org/10.3329/ptcb.v18i1.3243
Shi M, He W, Liu Y, Li X, Yang S, Xu Q (2013) Protective effect of total phenylethanoid glycosides from Monochasma savatieri Franch on myocardial ischemia injury. Phytomedicine 20:1251–1255. https://doi.org/10.1016/j.phymed.2013.06.014
Skała E, Grąbkowska R, Sitarek P, Kuźma L, Błauż A, Wysokińska H (2015) Rhaponticum carthamoides regeneration through direct and indirect organogenesis, molecular profiles and secondary metabolite production. Plant Cell Tiss Org Cult 123:83–98. https://doi.org/10.1007/s11240-015-0816-1
Stamm P, Topham AT, Mukhtar NK, Jackson MD, Tomé DF, Beynon JL, Bassel GW (2017) The transcription factor ATHB5 affects GA-mediated plasticity in hypocotyl cell growth during seed germination. Plant Physiol 173:907–917. https://doi.org/10.1104/pp.16.01099
Wang Y, Qiu JM (2018a) Induction of disinfectant and hormone on explants of Monochasma savatieri Franch ex Maxim. Xiandai Hort 363:20–21 CNKI:SUN:JXYA.0.2018.11.009 (in Chinese)
Wang Y, Qiu JM (2018b) Tissue culture technigues of Monochasma savatieri Franch ex Maxim. Xiandai Hort 367:33. https://doi.org/10.14051/j.cnki.xdyy.2018.19.016 (in Chinese)
Yang YZ (2009) Plantlet regeneration of Monochasma savatieri Franch. ex Maxim. in vitro culture from inducing adventitious root. Plant Physiol Commun 7:693–694 (in Chinese)
Zhang M, Chen Y, Ouyang Y, Huang Z, Jaime A, Ma G (2015) The biology and haustorial anatomy of semi-parasitic Monochasma savatieri Franch. ex Maxim. Plant Growth Regul 75:473–481. https://doi.org/10.1007/s10725-014-0010-1
Zhang Y, Chen Y, Zhang X, Jaime A, Ma G (2017) Adventitious shoot induction from internode and root explants in a Semiparasitic herb Monochasma savatieri Franch ex Maxim. Plant Growth Regul 36:799–804. https://doi.org/10.1007/s00344-017-9681-y
Zheng W, Tan XQ, Guo LJ, Kong FF, Lu P, Ni DJ, Wang P (2012) Chemical constituents from Monochasma savatieri. Chin J Nat Med 10:102–104. https://doi.org/10.3724/sp.j.1009.2012.00102
Acknowledgments
The authors wish to thank the editor and reviewers for their helpful comments and suggestions.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Editor: Baochun Li
Rights and permissions
About this article

Cite this article
Li, F., Liu, S. & Zeng, M. An efficient micropropagation protocol for Monochasma savatieri Franch. ex Maxim through seed germination and direct shoot regeneration. In Vitro Cell.Dev.Biol.-Plant 57, 30–38 (2021). https://doi.org/10.1007/s11627-020-10103-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11627-020-10103-x