
Abstract
The traditional medicinal fungus, Lingzhi, has long been considered to be Ganoderma lucidum, but was recently described as G. lingzhi based on both morphological and molecular data. Months earlier than this description, the species was identified as G. sichuanense by another research group. A sequenced epitype for G. sichuanense was selected that, if accepted, meant that it became a synonym of, and an earlier name for, G. lingzhi. However, the holotype of G. sichuanense is well developed with distinct morphological characters, and its ITS sequence, the DNA barcode for fungal taxonomy, is available. The holotype of G. sichuanense does not possess the typical morphological characteristics of Lingzhi, and the designated epitype is not conspecific with the holotype from morphological, phylogenetic and ecological perspectives. Consequently, the designation of the epitype is considered to conflict with the regulations of the Code and should be rejected. As no other previously published species name matches Lingzhi, the correct name for this fungus is G. lingzhi.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Lingzhi, an economically important wood-decay polypore fungus, has been used in traditional Chinese medicine for more than 2000 years (Anonymous 1955). The successful production of Lingzhi fruitbodies in 1969 (Yu and Shen 2003) led to widespread cultivation in China and adjacent countries, stimulating scientific research on the identity and function of the medicinal metabolites. These metabolites include polysaccharides, triterpenes, lucidenic acids, adenosines, ergosterols, glucosamines and cerebrosides with medicinal functions in the prevention and treatment of diseases such as cancers, chronic bronchitis, diabetes mellitus, hyperlipidemia and hypertension, as well as improvement of immunity and anti-aging (Lin 2009). The products containing Lingzhi have an estimated annual value of US$2.5 billion in global trade (Li et al. 2013). Besides the economic importance and health benefits, cultivation and utilization of Lingzhi is also considered an essential part of Chinese traditional culture.
Nomenclatural history of Lingzhi
The first possible name applied to Lingzhi (Mannen-take or Reishi in Japanese) in East Asia was Boletus dimidiatus Thunb. based on specimens collected on the Japanese mainland around Tokyo and Nagasaki (Thunberg 1784). Fries (1838) renamed this Polyporus japonicus Fr. Because this name is not sanctioned, it is illegitimate because there was no obstacle to recombining Thunberg’s epithet dimidiatus into Polyporus, and Fries explicitly refers to Thunberg’s name. Later, Yasuda (1913) suggested that Fomes japonicus (Fr.) Sacc. (= P. japonicus) recorded from Japan actually represents F. lucidus (Curtis) Sacc. [= Polyporus lucidus (Curtis) Fr.]. Imazeki (1939) made intensive studies of specimens of the Ganoderma lucidum (Curtis) P. Karst. complex collected in Japan, and concluded that specimens of “G. lucidum (Curtis) P. Karst.” recorded after Yasuda (1913) included G. lucidum s.s., G. neojaponicum Imazeki and G. curtisii (Berk.) Murrill. Imazeki considered that Mannen-take represented G. lucidum, while specimens with almost black pilei and restricted to conifers were G. neojaponicum.
Likewise, for Chinese mycota, Patouillard (1907) identified Chinese Lingzhi collections from Guizhou Province as Ganoderma lucidum (Moncalvo and Ryvarden 1997). While G. lucidum was then broadly defined, more recently many species have been differentiated from the original concept. However, the name G. lucidum has been widely used for Lingzhi in later and even recent publications including studies of medicinal metabolites and functions (Kino et al. 1989; Bao et al. 2002; Sliva et al. 2002). Detailed morphological examinations (Pegler and Yao 1996) and phylogenetic analyses (Moncalvo et al. 1995; Smith and Sivasithamparam 2000; Hong and Jung 2004) have demonstrated that the application of the name G. lucidum for Lingzhi is no longer correct (Szedlay 2002; Wasser 2011). However, the nomenclatural consequences of these taxonomic results have been hotly debated.
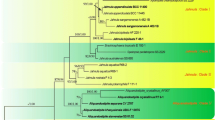
Cao et al. (2012) sampled both natural and cultivated Lingzhi specimens from China and Japan as well as specimens of Ganoderma species resembling Lingzhi, including G. curtisii (Berk.) Murrill, G. flexipes Pat., G. lucidum, G. multipileum Ding Hou, G. sichuanense J.D. Zhao & X.Q. Zhang and G. tropicum (Jungh.) Bres., and performed a phylogenetic analysis including a sequence from the holotype of G. sichuanense. They demonstrated that Lingzhi represented an independent lineage. Moreover, distinct morphological differences between G. lingzhi and these species were documented in Cao et al. (2012). Therefore, a new species, Ganoderma lingzhi Sheng H. Wu, Y. Cao & Y.C. Dai, was introduced for Lingzhi, a species with a wide natural distribution in temperate East Asia.
Months earlier than the description of Ganoderma lingzhi, Wang et al. (2012) proposed Ganoderma sichuanense as the correct name of Lingzhi. Ganoderma sichuanense was originally described based on two collections from Panzhihua Iron and Steel (Group) Company Limited in Dukou City (former name of Panzhihua City), southern Sichuan Province, southwestern China (Zhao et al. 1983). Wang et al. (2012) concluded that the two collections represented different species: the paratype (HMAS 43728, with basidiospore dimensions 8–8.5 × 6–6.5 μm) was identified as Ganoderma weberianum (Bres. & Henn. ex Sacc.) Steyaert, while the holotype (HMAS 42798) was considered to be conspecific with Lingzhi. However, Wang et al. (2012) could not obtain any DNA sequence from the holotype of G. sichuanense. Instead, for DNA analyses, they used four recently collected samples from Sichuan (HMAS 251145–251,148) that they considered to be morphologically similar, from an area close to but distinct from the type locality. These samples were considered by Wang et al. (2012) to be Lingzhi and to match G. sichuanense. To stabilize this concept, Yao et al. (2013) subsequently designated a sample recently collected from Huangqiao Village, Miyi County, Panzhihua, Sichuan Province (HMAS 252081) and bearing an ITS sequence identical to that of Lingzhi, as the epitype of G. sichuanense. They justified this with the argument that DNA in the holotype of G. sichuanense had not been well preserved. This action, were it to be accepted, meant that G. lingzhi became a later synonym of G. sichuanense.
Evidence to support Ganoderma lingzhi as the correct scientific binomial for Lingzhi
As stated by Zhou et al. (2015), the epitypification of G. sichuanense does not comply with the International Code of Nomenclature for algae, fungi, and plants (Melbourne Code). Art. 9.8 defines an epitype as “an interpretative type” selected when the holotype or other original material "is demonstrably ambiguous and cannot be critically identified for purposes of the precise application of the name to a taxon".
The holotype of G. sichuanense is well developed with clear morphological characters and its ITS region, the DNA barcode for fungal taxonomy, was successfully sequenced by Cao et al. (2012). The failure of Wang et al. (2012) and Yao et al. (2013) to amplify any sequence from the holotype was, in their statement, because of “the specimen’s condition”. However, a failed PCR can have more causes than damage of the DNA, so the interpretations of Yao et al. (2013) are vulnerable.
Furthermore, the epitype of G. sichuanense was inappropriately selected and could not be interpreted with certainty as conspecific with the holotype, because these collections differ in at least four aspects:
-
a)
According to phylogenetic analysis, the holotype (GenBank accession JQ781877, as sequenced by Cao et al. 2012) and the epitype (KC662402, as sequenced by Yao et al. 2013) occur in distinctly separated lineages (Yao et al. 2013). The two sequences are not identical and in our examination showed a difference of more than 8%.
-
b)
The basidiospore dimensions of the epitype [(9.0–)9.5–10.5(−11.0) × 6.5–8.0 μm] in Yao et al. (2013) are much larger than those of the holotype of G. sichuanense [(7–)7.4–9.2(−9.3) × (4.6–)5–6.6(−6.8)] in Cao et al. (2012; Table 2). Yao et al. (2013) stated that their measurements were from at least 30 basidiospores, and the quoted measurements from Cao et al. (2012) were also based on 30 basidiospores. Yao et al. (2013), following Wang et al. (2012), maintained that basidiospores from the holotype of G. sichuanense are 9–11.5 × 6.5–8 μm, and explained the discrepancy in basidiospore dimensions by stating that Cao et al. (2012) also included measurements from the paratype of G. sichuanense that had smaller basidiospores than the holotype. However, Cao et al. (2012) only borrowed the holotype, not the paratype, to study. Moreover, Cao et al. (2012)‘s measurements of the holotype are consistent with the basidiospore dimensions (7.4–9.5 × 5–7 μm) mentioned in the original publication on G. sichuanense by Zhao et al. (1983), where the four illustrated basidiospores from the holotype are all shorter than 10 μm. Hence, there is no evidence of confusion of basidiospore size between the holotype and paratype.
-
c)
Macromorphologically, the holotype has a much darker pore surface than the epitype (see photographs in Cao et al. 2012; Yao et al. 2013).
-
d)
The ecological environment of the epitype is distinctly different from that of the holotype of G. sichuanense: 80 km distant in a straight line and, more importantly, at an altitude of nearly 1000 m above that of the holotype. The epitype from Miyi occurred in a humid and temperate forest at an altitude of 1933 m as mentioned by Yao et al. (2013), representing a temperate species like Lingzhi, while the locality of the holotype of G. sichuanense (Dukou) was found at an altitude of 985 m with dry and nearly tropical vegetation (indicated by mango cultivation on the basis of our observation in this area). Another specimen of G. sichuanense (Cui 7691), with an identical ITS sequence to that of the holotype, was also collected from the subtropical to tropical zone in Guangdong (Cao et al. 2012). Yao et al. (2013) considered the sequence from the holotype “unusual” without further explanation. If this term was to indicate artifacts of DNA preparation, the 100% identity with the sequence from Cui 7691 is hardly explicable.
It is evident that the so-called epitype of G. sichuanense is different from the holotype of G. sichuanense, according to the above-mentioned phylogenetic, morphological and ecological perspectives. This means that the epitype represents G. lingzhi, but not G. sichuanense. The intention of Yao et al. (2013) in proposing the epitype actually serves to replace the holotype rather than add to it. Richter et al. (2015) agreed with Yao et al. (2013)‘s opinion, but did not provide any evidence or new data to support this epitype.
Other potential synonyms
Like Ganoderma sichuanense, several other previously described Chinese species of Ganoderma, such as G. atrum J.D. Zhao, L.W. Hsu & X.Q. Zhang, G. calidophilum J.D. Zhao, L.W. Hsu & X.Q. Zhang, G. chenghaiense J.D. Zhao, G. hainanense J.D. Zhao, L.W. Hsu & X.Q. Zhang, G. parviungulatum J.D. Zhao & X.Q. Zhang and G. simaoense J.D. Zhao, are more or less morphologically similar to G. lingzhi (Zhao et al. 1979, 1986; Zhao 1987, 1989). However, Cao et al. (2012) clearly concluded these species are not conspecific with G. lingzhi by type studies. Moreover, in the absence of molecular sequences from the holotypes of these species, their species circumscriptions are not fully clear and it is not possible to use any of these names for Lingzhi (Ryvarden 1995).
Considering Boletus dimidiatus and Polyporus japonicus, the original descriptions and drawings suggest that they represent a member of the G. lucidum complex. The name Ganoderma japonicum (Fr.) Sawada (= P. japonicus) has often been applied to a Chinese species with a dark pileus and distribution in subtropical areas (e.g. Teng 1964). However, it is most likely a misapplication, because specimens of G. japonicum from China and Japan are not conspecific. The existing sequences of G. japonicum were obtained from Chinese specimens (e.g., Wang and Yao 2005). While they differ from those of Lingzhi, this is not a valid reason to conclude that G. japonicum is distinct from Lingzhi. However, the authentic material of B. dimidiatus and P. japonicus deposited at the Museum of Evolution Herbarium (UPS: BOT: F-118347) was not available for loan and exists as drawings provided by Thunberg (1784), and we conclude that the rejection of the name Boletus dimidiatus is better in order to stabilize the scientific binomial for Lingzhi.
Conclusion
Ganoderma lingzhi is a temperate species widely distributed mainly in Fagaceae forests of East Asia. Using both morphological and phylogenetic evidence, this name unambiguously represents the economically important and widely cultivated Lingzhi species. In 2013, G. lingzhi was nominated as the fungus of the year in Mycology, an international journal issued by the Mycological Society of China (Yang and Feng 2013). Moreover, in addition to taxonomic mycologists (Hawksworth 2013; Li et al. 2015a, 2015b; Hennicke et al. 2016), the name G. lingzhi has also been accepted instead of G. lucidum by mycological chemists (Fatmawati et al. 2013; Gao et al. 2014; Li et al. 2014; Chen et al. 2015; Yan et al. 2015; Wang et al. 2016; Zhang et al. 2017), while to the best of our knowledge the name G. sichuanense has never been applied to these kinds of chemical studies.
Ganoderma sichuanense is a subtropical-tropical species from South China, and bears morphological features distinct from those of G. lingzhi (Cao et al. 2012), whereas the so-called epitype of G. sichuanense designated by Yao et al. (2013) represents G. lingzhi and is inadequate as an epitype. In order to stop the widespread misapplication of epitypification in fungi, Ariyawansa et al. (2014) proposed strict criteria for meaningful epitypification, such as multilocus-based phylogenetic analyses, which are not met in the epitypification of Yao et al. (2013). Wang et al. (2012) and Cao et al. (2012) presented different basidiospore measurements from the holotype of G. sichuanense, and the latter corresponds with the protologue. The single available DNA sequence from this specimen clusters in a clade comprising G. weberianum and G. sichuanense. The present inconsistent findings indicate that the species concept of G. sichuanense is ambiguous. There is urgent need to stabilize nomenclature for the economically important Lingzhi species. Currently, the only unambiguous name for the widely cultivated Lingzhi is G. lingzhi. A formal proposal to conserve Ganoderma lingzhi over G. sichuanense and Boletus dimidiatus will be prepared for solving the scientific binomial for the widely cultivated medicinal fungus Lingzhi thoroughly.
References
Anonymous (1955) Shen Nong materia medica, 102-200 A.D. In: Han E (ed) Reprinted. People’s Hygiene Press, Beijing (in Chinese)
Ariyawansa HA, Hawksworth DL, Hyde KD et al (2014) Epitypification and neotypification: guidelines with appropriate and inappropriate examples. Fungal Divers 69:57–91
Bao XF, Wang XS, Dong Q, Fang JN, Li XY (2002) Structural features of immunologically active polysaccharides from Ganoderma lucidum. Phytochemistry 59:175–181
Cao Y, Wu SH, Dai YC (2012) Species clarification of the prize medicinal Ganoderma mushroom “Lingzhi”. Fungal Divers 56:49–62
Chen H, Yang HL, Liu GQ (2015) Biosynthesis and fermentation control of triterpenoids from Ganoderma lingzhi. Mycosystema 34:1–9 (in Chinese)
Fatmawati S, Kondo R, Shimizu K (2013) Structure–activity relationships of lanostane-type triterpenoids from Ganoderma lingzhi as α-glucosidase inhibitors. Bioorg Med Chem Lett 23:5900–5903
Fries E (1838) Epicrisis systematis mycologici seu synopsis Hymenomycetum. Typographia Academica, Uppsala
Gao YJ, Yuan B, Yang WJ, Fang Y, Ma N, Hu QH (2014) Preparation of Ganoderma lingzhi spore oil microcapsules. Mycosystema 33:483–492 (in Chinese)
Hawksworth DL (2013) What is the Chinese Lingzhi Ganoderma? IMA Fungus 4(2):47
Hennicke F, Cheikh-Ali Z, Liebisch T, Maciá-Vicente JG, Bode HB, Piepenbring M (2016) Distinguishing commercially grown Ganoderma lucidum from Ganoderma lingzhi from Europe and East Asia on the basis of morphology, molecular phylogeny, and triterpenic acid profiles. Phytochemistry 127:29–37
Hong SG, Jung HS (2004) Phylogenetic analysis of Ganoderma based on nearly complete mitochondrial small-subunit ribosomal DNA sequences. Mycologia 96:742–755
Imazeki R (1939) Studies on Ganoderma of Nippon. Bull Tokyo Sci Mus 1:29–52 (in Japanese)
Kino K, Yamashita A, Yamaoka K, Watanabe J, Tanaka S, Ko K, Shimizu K, Tsunoo H (1989) Isolation and characterization of a new immunomodulatory protein, ling zhi-8 (LZ-8), from Ganoderma lucidum. J Biol Chem 264:472–478
Li J, Zhang J, Chen H, Chen XQ, Lan L, Liu C (2013) Complete mitochondrial genome of the medicinal mushroom Ganoderma lucidum. PLoS ONE 8:e72038
Li KK, Zhang CM, Gu HD, Jin Y, Qu JJ (2014) Optimization of Ganoderma lingzhi S3 fermentation medium and inhibition of the fermented liquid against Fusarium oxysporum. Mycosystema 33:401–410 (in Chinese)
Li TH, Hu HP, Deng WQ, Wu SH, Wang DM, Tsering T (2015a) Ganoderma leucocontextum, a new member of the G. lucidum complex from southwestern China. Mycoscience 56:81–85
Li Y, Li TH, Yang ZL, Bau T, Dai YC (2015b) Atlas of Chinese macrofungal resources. Central China Farmer's Publishing House, Zhengzhou
Lin ZB (2009) Lingzhi: from mystery to science. Peking University Medical Press, Beijing
Moncalvo JM, Ryvarden L (1997) A nomenclatural study of the Ganodermataceae Donk. Synopsis Fungorum 11:1–114
Moncalvo JM, Wang HF, Hseu RS (1995) Gene phylogeny of the Ganoderma lucidum complex based on ribosomal DNA sequences. Comparison with traditional taxonomic characters. Mycol Res 99:1489–1499
Patouillard N (1907) Champignons du Kouy-tcheou. Monde d be Pl Seri II 9:31
Pegler DN, Yao YJ (1996) Oriental species of Ganoderma section Ganoderma. In: Wasser SP (ed) Botany and mycology for the next millenium: collection of scientific articles devoted to the 70th Anniversary of Academician Sytnik KM. Kholodny NG Institute of Botany, National Academy of Sciences of Ukraine, Kyiv, pp 336–347
Richter C, Wittstein K, Kirk PM, Stadler M (2015) An assessment of the taxonomy and chemotaxonomy of Ganoderma. Fungal Divers 71:1–15
Ryvarden L (1995) Can we trust morphology in Ganoderma? In: Buchanan PK, Hseu RS, Moncalvo JM (eds) Ganoderma: systematics, phytophathology and pharmacology. Proceedings of contributed symposium 59A, B. 5th international mycological congress, Vancouver, August 14–21, vol 1994. National Taiwan University, Taipei, pp 19–24
Sliva D, Labarrere C, Slivova V, Sedlak M, Lloyd FP Jr, Ho NWY (2002) Ganoderma lucidum suppresses motility of highly invasive breast and prostate cancer cells. Biochem Biophys Res Commun 298:603–612
Smith BJ, Sivasithamparam K (2000) Internal transcribed spacer ribosomal DNA sequence of five species of Ganoderma from Australia. Mycol Res 104:943–951
Szedlay G (2002) Is the widely used medicinal fungus the Ganoderma lucidum (Fr.) Karst. sensu stricto? (A short review). Acta Microbiol Immunol Hung 49:235–243
Teng SC (1964) Fungi of China. Academia Sinica, Beijing (in Chinese)
Thunberg CP (1784) Flora Japonica. Reprinted. Oriole, New York
Wang DM, Yao YJ (2005) Intrastrain internal transcribed spacer heterogeneity in Ganoderma species. Can J Microbiol 51:113–121
Wang XC, Xi RJ, Li Y, Wang DM, Yao YJ (2012) The species identity of the widely cultivated Ganoderma, ‘G. lucidum’ (Ling-zhi), in China. PLoS ONE 7:e40857
Wang XL, Ding ZY, Liu GQ, Yang H, Zhou GY (2016) Improved production and antitumor properties of triterpene acids from submerged culture of Ganoderma lingzhi. Molecules 21:1395
Wasser SP (2011) Current findings, future trends, and unsolved problems in studies of medicinal mushrooms. Appl Microbiol Biotechnol 89:1323–1332
Yan YM, Wang XL, Luo Q, Jiang LP, Yang CP, Hou B, Zuo ZL, Chen YB, Cheng YX (2015) Metabolites from the mushroom Ganoderma lingzhi as stimulators of neural stem cell proliferation. Phytochemistry 114:155–162
Yang ZL, Feng B (2013) What is the Chinese “Lingzhi”? – a taxonomic minireview. Mycology 4:1–4
Yao YJ, Wang XC, Wang B (2013) Epitypification of Ganoderma sichuanense J.D. Zhao & X.Q. Zhang (Ganodermataceae). Taxon 62:1025–1031
Yasuda A (1913) Kinrui-Zakki 17. Bot Mag Tokyo 27:23–24 (in Japanese)
Yu YN, Shen MZ (2003) The history of Lingzhi (Ganoderma spp.) cultivation. Mycosystema 22(Supp l):3–9 (in Chinese)
Zhang DH, Li N, Yu X, Zhao P, Li T, Xu JW (2017) Overexpression of the homologous lanosterol synthase gene in ganoderic acid biosynthesis in Ganoderma lingzhi. Phytochemistry 134:46–53
Zhao JD (1987) Studies on the taxonomy of Ganodermataceae in China 10. Subgen. Ganoderma sect. Phaeonema. Acta Mycol Sin 7:205–211
Zhao JD (1989) Studies on the taxonomy of Ganodermataceae in China 11. Subgen. Ganoderma sect. Ganoderma. Acta Mycol Sin 8:25–34 (in Chinese)
Zhao JD, Xu LW, Zhang XQ (1979) Taxonomic studies on the subfamily Ganodermoideae of China. Acta Microbiol Sin 19:265–279 (in Chinese)
Zhao JD, Xu LW, Zhang XQ (1983) Taxonomic studies on the family Ganodermataceae of China 2. Acta Mycol Sin 2:159–167
Zhao JD, Zhang XQ, Xu LW (1986) Studies on the taxonomy of Ganodermataceae in China 4. Acta Mycol Sin 5:86–93 (in Chinese)
Zhou LW, Cao Y, Wu SH, Vlasák J, Li DW, Li MJ, Dai YC (2015) Global diversity of the Ganoderma lucidum complex (Ganodermataceae, Polyporales) inferred from morphology and multilocus phylogeny. Phytochemistry 114:7–15
Acknowledgement
LWZ is grateful to the Youth Innovation Promotion Association CAS (No. 2017240) for financial support.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Section Editor: Marc Stadler
Rights and permissions
About this article

Cite this article
Dai, YC., Zhou, LW., Hattori, T. et al. Ganoderma lingzhi (Polyporales, Basidiomycota): the scientific binomial for the widely cultivated medicinal fungus Lingzhi. Mycol Progress 16, 1051–1055 (2017). https://doi.org/10.1007/s11557-017-1347-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11557-017-1347-4