
Abstract
The nutritional status, the soil chemical attributes prior to vine killing, and the classified yield of tubers may characterise soil–plant interactions with nitrogen (N) sidedressing for potato cultivars. Two experiments with potato cultivars Agata and Atlantic were carried out with the aim of evaluating the effects of N sources (urea, ammonium sulfate, and calcium nitrate) and N application rates (70, 140, and 210 kg ha−1) as sidedressing, plus a zero-N control treatment, in southern Brazil. Discrete variations in nutrients in diagnostic leaves and in soil chemical attributes in the Agata experiment were not reflected in the maximum average increase of 69.0% (14.2 Mg ha−1) in total tuber yield obtained with the calculated rate for maximum technical efficiency of 152 kg N ha−1 as ammonium sulfate or calcium nitrate. These N sources did not differ in agronomic efficiency (AE). For the Atlantic cultivar, urea was practically inert in terms of soil and plant attributes, and the ammonium sulfate addition acidified the soil, increased its Al3+ concentration in the ridges, reduced the foliar Ca and Mg concentrations, and did not increase the marketable tuber yield. Calcium nitrate increased foliar N, Ca, and Mg concentrations, increasing marketable tuber yields by 26.6% (7.3 Mg ha−1) with 116.7% higher AE than other N sources. In subtropical acidic inceptisols with potentially available Al3+, 153 kg N ha−1 ammonium sulfate or calcium nitrate for the cultivar Agata and 152 kg N ha−1 as calcium nitrate for the cultivar Atlantic are the best options for sidedressing fertilisation.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
With a global production of 359 million tons of tubers in 2020, potato (Solanum tuberosum L.) is the third most important food crop in the world (FAOSTAT 2023) and could become one of the most advantageous in terms of food security when maximum cereal crop yields are close to their limit (Dongyu 2022). However, under climate change, global warming could harm potato crops in regions at higher latitudes, although it should favour potato cultivation during the winter season in subtropical regions (Hijmans 2003; Bender and Sentelhas 2020). Potato production has stood out in tropical and subtropical regions, including Brazil (Haverkort and Struik 2015; Lizana et al. 2020; FAOSTAT 2023). In these environments, improvements in crop management, such as the application of nitrogen (N), are more important than advances in genetic parameters (Kleinwechter et al. 2016).
Nitrogen is the second nutrient most commonly taken up by the main potato cultivars planted in Brazil, mostly (> 80%) during the tuberization phase (Fernandes et al. 2011, 2021; Silva et al. 2020). Thus, it is a key nutrient that increases tuber yields in regions with subtropical climates in the Northern Hemisphere (Zotarelli et al. 2014; Makani et al. 2020) and tropical Brazilian environments (Fontes et al. 2010; Souza et al. 2019; Fernandes et al. 2021). In addition, supplying N in the most demanding crop development stage has been the best strategy for obtaining higher tuber yields avoiding N losses (Zotarelli et al. 2014; Rens et al. 2016; Souza et al. 2019; Fernandes et al. 2022). In Brazil, the Agata and Atlantic cultivars are the main potato cultivars produced for use in fresh markets and as chips (or crisps), respectively. These cultivars differ in growth (Fernandes et al. 2010; Silva et al. 2013), nutrient requirements (Fernandes et al. 2011; Silva et al. 2020), nutrient utilisation efficiencies (Fernandes and Soratto 2013), and tuber yield in response to N fertilisation (Fontes et al. 2010; Oliveira et al. 2020). In addition, these cultivars exhibit different patterns of dry matter partitioning to tubers according to the N source (Silva et al. 2013).
The most suitable source and rate of N application during sidedressing must be precise and synchronised with a short period of maximum N uptake by the potato crop, which is the initial stage of tuberization (Fernandes et al. 2011). The application of granulated ammonium nitrate during preplanting; of nitrate, urea, or liquid ammonium during the initiation of potato tuber growth (Zotarelli et al. 2014; Rens et al. 2016); of granulated ammonium nitrate dissolved in water for drip irrigation according to nutritional demands (Makani et al. 2020); and four applications of urea during the crop cycle (Liu et al. 2021) are examples of N management with sources, rates, and application times of N in subtropical sandy soils in the Northern Hemisphere. In Brazil, sidedressing N fertilisation is usually applied to potato crops immediately before the hilling operation, approximately 30 days after planting (DAP) (Souza et al. 2019; 2020; Yagi et al. 2020), and urea or ammonium sulfate (AMS) is commonly used (Souza et al. 2019; 2020; Fernandes et al. 2021). Fertiliser incorporation into the soil by adding N fertilisers with or without other nutrients, such as potassium (K) and chlorine (Cl), is a common practice in Brazil and can affect both the dynamics and the availability of the main inorganic forms of N, as well as the soil chemical attributes of the ridge microenvironment.
Partial nitrification of NH4+-N, resulting from sidedressing N application or soil organic matter mineralisation, results in the concentration of NH4+-N and NO3−-N at the top of the ridge immediately after hilling, increasing the NO3−-N levels at the base of the ridge until vine killing (Zebarth and Milburn 2003). The NH4+-N should be the N form that is metabolically preferred by potato plants, as it does not need to be reduced to be assimilated but it is harmful to plant growth when it is available at high concentrations and exclusively available at any growth stage (Davis et al. 1986). Potato plants tolerate and increase the dry matter of stems, leaves, and tubers in a less wide range with NH4+-N than with NO3−-N (Cao and Tibbitts 1998). Furthermore, as the pH decreases, the electrical conductivity of the medium increases when adding NH4+-N compared with NO3−-N (Serio et al. 2004). At moderate concentrations, NH4+-N favours shoot growth before tuber initiation (Gao et al. 2014) and stimulates tuber bulking (Osaki et al. 1995a, b) and earlier tuber initiation (Qiqige et al. 2017). On the other hand, NO3−-N promotes the plant branching and growth of stolons (Osaki et al. 1995a, b), which swell or form tubers earlier (Gao et al. 2014) and in greater quantity than plants fed with NH4+-N (Serio et al. 2004; Gao et al. 2014; Qiqige et al. 2017).
The NH4+-N uptake decreases calcium (Ca) concentration in the shoots of potato plants more than NO3−-N nutrition (Polizotto et al. 1975; Davis et al. 1986; Cao and Tibbitts 1993), which motivates an additional supply of Ca to prevent possible deficiencies (Cao and Tibbitts 1993). In this sense, calcium nitrate (CN) has been evaluated as a source of Ca for potato plants with supplementary input of NO3−-N (Simmons and Kelling 1987; Giletto et al. 2022). This fertiliser has been used in split applications during plant emergence and tuber initiation stages in sandy soil in temperate climates (Bundy et al. 1986). According to the latter authors, there were no different effects of split-applied CN, AMS, urea, and ammonium nitrate on total and graded tuber yields over 5 years. However, on average over the years, they found that AMS increased total tuber yields by 3.5 Mg ha−1 (6.2%) in relation to the average of the other N sources. Given the above, we hypothesise that in medium-textured soils of a subtropical environment, the effects of sidedressed ammoniacal, nitric, and amidic N sources may be distinct and vary according to the potato cultivar, with these effects being observed in terms of tuber yield and size, plant nutritional status, and soil chemical attributes.
The objective of this study was to evaluate the effects of sidedressing N sources and rates on soil fertility, plant nutrition, and the tuber yield and size of Agata and Atlantic potato cultivars in subtropical southern Brazil.
Materials and methods
Site, Experimental Design, and Crop Management
The study was carried out during the 2020–2021 spring–summer season in a commercial potato field in Porto Amazonas, Paraná State, southern Brazil (25° 34′ 06″ S; 49° 55′ 54″ W; altitude of 817 m). Two experiments were conducted concurrently in adjacent areas, one with Agata and another with the Atlantic cultivar. The soil is classified as a Typic Inceptisol (Soil Survey Staff 2022). Representative soil samples from the area of each experiment were collected before planting to determine their chemical (Pavan et al. 1992) and granulometric (EMBRAPA 1997) attributes (Table 1). The climate in the region is humid subtropical Cwb (mild summer, evenly distributed rainfall and no dry season), according to the Köppen classification system. Daily air temperature and rainfall were recorded at a weather station located approximately 1.5 km from the experimental areas (Figure S1).
In both experiments, the treatments comprised the combinations of three N rates (70, 140, and 210 kg N ha–1) with three N fertiliser sources [urea, 45% N; ammonium sulfate (AMS), 20% N + 24% S; and calcium nitrate (CN), 16% N + 19% Ca, sidedressed immediately before hilling], plus a control without sidedressed N fertilisation. The treatments were arranged in a 3 × 3 + 1 factorial scheme, with a randomised block design and four replications. Each plot comprised four 5-m-long rows with 0.80-m row distance. Two of the centre rows were used for the experimental measurements. In each plot, the two border rows and the 0.5-m end rows were not used for measurements.
The areas were cultivated with black oat (Avena strigosa Schreber) before the planting. This grass was killed with 1500 g ha−1 glyphosate, and the soil was tilled with subsoiling, ploughing, and light harrowing operations. As soil base saturation was higher than 60%, a critical value for liming according to regional criteria (NEPAR-SBCS 2017), no limestone was applied.
The furrows were mechanically opened with a furrower, and the starter fertiliser was applied manually and incorporated into the furrows in all plots with a hoe at an average depth of 0.15 m. Starter fertiliser (a mix of single superphosphate and a multinutrient formula in the same granule containing 6% N, 12% P, 5% K, 8% Ca, 8.4% S, 0.03% B, 0.05% Mn, and 0.1% Zn) provided 40 kg N, 183 kg P, 33 kg K, 349 kg Ca, 218 kg S, 0.201 kg B, 0.335 kg Mn, and 0.670 kg Zn ha−1. Starter fertilisation was calculated in accordance to recommendations for potato crop (Soratto et al. 2016; NEPAR-SBCS 2017) and to meet S demand by potato crop (Souza et al. 2019). Most of the K fertilisation [i.e. 91 kg K ha−1 (KCl, 50% K)] was sidedressed in all plots prior to hilling (Yagi et al. 2020; NEPAR-SBCS 2017).
Immediately after planting fertiliser, seed tubers of the potato cultivars Agata and Atlantic, weighing approximately 35 g and with diameters between 30 and 40 cm, were planted with a spacing of 0.30 m between them on 19 October 2020. Pesticides use was guided by pest management recommendations commonly adopted in Brazil (Lopes et al. 2016), using drones with sprayings in an ultralow volume system. At 40 DAP, N treatments were manually applied prior to hilling. At 60 DAP, Mn, Cu, Zn, and B, at the respective rates of 165, 55, 42, and 150 g ha−1, were sprayed to the crop foliage using a drone. At 40 DAP, N treatments were manually applied, plus KCl in all plots, prior to the hilling operation. At 60 DAP, Mn, Cu, Zn, and B foliar applications were performed using a drone at the respective rates of 165, 55, 42, and 150 g ha−1. Supplementary irrigation was not necessary due to the adequate distribution of rainfall during the experimental period (Figure S1). A total of 595 mm of rainfall was recorded throughout the entire crop cycle and 480 mm (80.7%) after the application of treatments and hilling.
Sampling and Analysis
Plant nutritional status was evaluated via diagnostic leaf sampling at 34 and 30 days after emergence (DAE) respectively in experiments with the Agata and Atlantic cultivars. The third expanded leaf (leaflets plus petioles) from the apex of 12 plants selected randomly from each plot was collected. The leaves were rinsed with deionised water, dried in an oven with forced air circulation at 65 °C for 72 h, ground to pass a 1.0-mm sieve and then chemically analysed (Malavolta et al. 1997). After nitric-perchloric digestion (HNO3 + HClO4), the concentrations of P, Ca, Mg, S, Cu, Mn, and Zn were determined via atomic absorption spectrophotometry, while K concentration was determined via flame photometry. Leaf N concentration was determined via extraction with sulfuric acid (H2SO4) digestion and quantified via the semimicro-Kjeldahl method (Malavolta et al. 1997). After ashing the leaf samples at 550 °C and dissolving the ash in dilute HCl, B concentration was colorimetrically determined using the azomethine-H method (Malavolta et al. 1997).
After the vine killing, soil samples were taken (0.0–0.2 m) by collecting 12 simple samples in each plot between the potato plants in the ridge. The soil samples were air-dried, sieved, and chemically analysed using methods described in Pavan et al. (1992). The active acidity (pH) was estimated potentiometrically with a CaCl2 solution of 0.01 mol L−1 (ratio 1:2.5; soil:CaCl2 solution). The potential acidity (H + Al) was estimated potentiometrically by the SMP buffer method, and the exchangeable acidity (Al3+) was estimated via extraction with 1 mol L−1 KCl solution (1:10 ratio; soil:KCl solution) and titration with 0.0125 mol L−1 NaOH. Exchangeable Ca and Mg (Ca2+ and Mg2+) were extracted with 1 mol L−1 KCl and quantified via atomic absorption spectrophotometry. Exchangeable K (K+) was extracted with Mehlich-1 solution and determined by flame photometry. The cation exchange capacity (CEC) was calculated as the sum of H + Al, Ca2+, Mg2+, and K+ values. The base saturation indices (i.e. Ca2+, Mg2+, and K+ saturation) of the ion exchange complex was calculated according to the formula: base saturation (%) = ((Ca2+ + Mg2+ + K+)/CEC) × 100.
To harvest tubers, the two 4 m long of the centre rows of each plot were considered. Thus, rows with 13 to 14 plants were thinned and standardised to 12 plants per row in both experiments, excluding the plants at the ends of the rows. Harvesting was carried out at 150 DAP in a semimechanized manner with a two-line tractorized potato harvester followed by manual collection. According to the transverse diameter, the tubers were classified as marketable (> 45 mm) or nonmarketable (< 45 mm) and were subsequently weighed. The total tuber yield was obtained by adding the yields of marketable and nonmarketable tubers.
Agronomic efficiency (AE) was calculated as a measure of the yield gained (marketable and total tuber yield) per unit of sidedressing N applied [AE (kg tuber increased kg−1 N applied) = (yield with sidedressing N application − yield without sidedressing N application)/sidedressing N application rate].
Statistical Analyses
A combined analysis of variance using the two experiments (cultivars) resulted in significant interactions for almost all the measured variables; therefore, individual analyses of variance were separately performed for each cultivar using Agroestat software (Barbosa and Maldonado Junior 2015). The blocks and all of the block interactions were considered random effects. The sidedressing N rate and source were considered fixed effects. To maintain a balanced experimental design, the data from the control (without sidedressing N fertilisation) were excluded from the tests of the main effect of the N source and N rate × N source interaction in a (3 × 3) + 1 factorial scheme. The effect of the quantitative factor was evaluated by polynomial regression analyses independently of the analysis of variance (Finkelman et al. 1977; Piepho and Edmondson 2018); therefore, the effects of N application were split among the three N sources for all dependent variables. For this purpose, the control (without sidedressing N fertilisation) was considered the zero rate common for all the independent variables. Significant (p ≤ 0.05) regression equations relating the N rates and dependent variables were plotted, and the most significant polynomial equation was selected between linear or quadratic functions.
The N source means were compared using Tukey’s test (p < 0.05), according to the interaction (p < 0.05) between the factors according to the F test of the analysis of variance.
Results
Plant Nutritional Status
There were no differences between the sidedressing N sources on leaf N concentrations in either experiments (Table S1). There was no significant effect of any N source on the leaf N concentration of the Agata cultivar (Fig. 1a). In the Atlantic potato experiment, the mean leaf N concentrations increased linearly and quadratically with urea-N and CN-N rates, respectively (Fig. 2b). The respective maximum increments under an applied N rate of 210 kg N ha−1 were 6.0 (15.2%) and 6.5 g kg−1 (16.5%) in relation to the control treatment without N in sidedressing.

Concentrations of N (a), Ca (b), Mn (c), and Zn (d) in the diagnostic leaves of potato cultivar Agata as affected by sidedressing nitrogen rates and sources. Vertical bars indicate the standard deviation of the mean. Different letters indicate differences at p ≤ 0.05 according to Tukey’s test to separate the means in the factorial 3 N sources × 3 N rates (70, 140, and 210 kg N ha). Ns, * and ** indicate nonsignificant and significant at 5% and 1% probability, respectively, considering the treatment without N in sidedressing, as a common treatment for all N sources.
 : urea;
: urea;
 : ammonium sulfate;
: ammonium sulfate;
 : calcium nitrate
: calcium nitrate

Concentrations of N (a), Ca (b), Mg (c), B (d), and Mn (e) in the diagnostic leaf of the potato cultivar Atlantic as affected by sidedressing nitrogen rates and sources. Vertical bars indicate the standard deviation of the mean. Different letters indicate differences at p ≤ 0.05 according to Tukey’s test to separate the means in the factorial 3 N sources × 3 N rates (70, 140, and 210 kg N ha−1). Ns, * and ** indicate nonsignificant and significant at 5% and 1% probability, respectively, considering the treatment without N in sidedressing, as a common treatment for all N sources.
 : urea;
: urea;
 : ammonium sulfate;
: ammonium sulfate;
 : calcium nitrate
: calcium nitrate
The leaf Ca concentrations of the Agata cultivar were 15.4% greater on average under urea and CN than under AMS (Table S1). The AMS and CN rates linearly decreased and increased, respectively, the leaf Ca concentrations (Fig. 1b). With 210 kg N ha−1 as CN, a 12.4% increase in the leaf Ca concentration was obtained, while with AMS, there was a decrease of 18.9%. In the leaves of Atlantic potato plants, the Ca concentrations differed among the three N sources, with higher concentrations in response to CN, intermediate with urea and lower with AMS (Fig. 2b). Linear decreases with CN-N rates (14.0% with 210 kg N ha−1) and quadratic decreases with AMS-N (37.3% with 175 kg N ha−1) and urea-N rates (12.1% with 100 kg N ha−1) were observed for leaf Ca concentrations. Unlike those of foliar Ca, leaf Mg concentrations were 14.0 to 15.9% (depending on the cultivar) lower with sidedressing urea than sidedressing AMS and CN (Table S1). The quadratic effects of the CN-N and AMS-N rates on the leaf Mg concentrations of Atlantic cultivar were antagonistic, increasing first and decreasing last (Fig. 2c). According to the equations, the maximum and minimum estimated leaf Mg concentrations were 4.8 and 3.6 g kg−1 with 95 and 108 kg N ha−1, respectively.
The rates of AMS-N linearly increased leaf B concentrations of Atlantic cultivar, which increased by 8.4% in relation to the average concentration calculated for the other N sources (Table S1; Fig. 2d). Significant interactions between sidedressing N sources and rates were observed for leaf Mn and Zn concentrations in the Agata experiment and for leaf Mn concentration in the Atlantic experiment (Table S1, Figs. 1c, d and 2e). Leaf Mn concentrations of Agata cultivar did not follow a definite trend among the N sources, and the N rates’ effect did not fit any polynomial regression model (Fig. 1c). At the rate of 70 kg N ha−1, urea promoted a higher Mn concentration compared to CN, whereas at the rate of 140 kg N ha−1 AMS promoted higher Mn concentration compared to urea. Similarly, leaf Zn concentrations showed no definite trend at these intermediate N rates (Fig. 1d). For the Atlantic cultivar, there were quadratic effects of the AMS-N and CN-N rates on the leaf Mn concentrations (Fig. 2e).
Soil Chemical Attributes
In the Agata experiment, the soil chemical attributes in the ridge at vine killing time were not influenced by the treatments (Table S2), except the Al3+ concentration that increased linearly with AMS rates (Fig. 3a). Similar results were verified for Al3+ concentration in the Atlantic experiment (Fig. 3b). In the experiments with Agata and Atlantic cultivars, the Al3+ concentrations increased to 0.38 and 0.24 cmolc dm−3, respectively, at a rate of 210 kg ha−1 AMS-N, calculated according to the respective linear regression equations. Soil Al3+ concentrations were higher with AMS in relation to the averages of other N sources in each experiment (which did not differ) (Table S2). Thus, Al3+ concentrations with AMS application were 107.0% and 72.9% higher than the averages of urea and CN, respectively, in the Agata and Atlantic experiments.

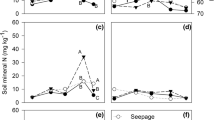
Concentration of exchangeable aluminium (Al3+) in the soil of the experiments with cultivars Agata (a) and Atlantic (b), base saturation index (c), pH (CaCl2) (d), and concentrations of exchangeable K (K+) (e) and Mg (Mg2+) (f) in the soil of the experiment with cultivar Atlantic, as affected by sidedressing nitrogen rates and sources. Vertical bars indicate the standard deviation of the mean. Different letters indicate differences at p ≤ 0.05 according to Tukey’s test to separate the means in the factorial 3 N sources × 3 N rates (70, 140, and 210 kg N ha−1). Ns, * and ** indicate nonsignificant and significant at 5% and 1% probability, respectively, considering the treatment without N in sidedressing, as a common treatment for all N sources.
 : urea;
: urea;
 : ammonium sulfate;
: ammonium sulfate;
 : calcium nitrate
: calcium nitrate
In the Atlantic experiment, there was also an effect of the N sources on the soil pH(CaCl2) values and the interaction of the N sources and rates on the soil Mg2+ concentration (Table S2). The rates of CN-N and AMS-N increased and decreased the soil pH (CaCl2) values up to a rate of 75 kg N ha−1, with maximum and minimum values of 5.2 and 4.9, respectively (Fig. 3d). At rates of 70 and 140 kg N ha−1, the pH (CaCl2) under AMS decreased by 0.3 (4.8%) and 0.4 (7.5%), respectively, in relation to the means obtained in response to urea and CN, which did not differ from each other (Table S2). Similarly, the base saturation values also decreased quadratically with increasing AMS-N rate, reaching a minimum value of 55% at 95 kg N ha−1 and then increasing to 63% at 120 kg ha−1 (Fig. 3c). The Mg concentrations also fit a quadratic model with the AMS-N rates, decreasing by up to 0.6 cmolc dm−3 with 71 kg ha−1 of N but subsequently increasing by up to 1.7 cmolc dm−3 with the maximum rate of N applied as sidedressing. This value was 81.9% higher than the average urea and CN also at 210 kg N ha−1 (Fig. 3f).
Soil K+ concentration increased with the application of urea at a rate of 70 kg N ha−1 followed by a decrease at rates of 140 and 210 kg N ha−1 (Fig. 3e). In relation to the control treatment, there was an increase of 28.5% in K+ (0.77 cmolc dm−3 with 130 kg ha−1 urea-N; Fig. 3e) in addition to decreases of 8.3% (55% base saturation with 95 kg ha−1 AMS-N; Fig. 3c) and 39.0% in Mg2+ (0.59 cmolc dm−3 with 74 kg ha−1 AMS-N; Fig. 3f). With 210 kg N ha−1 applied as a sidedressing, however, there was an increase of 95.2% in Mg2+ (1.9 cmolc dm−3; Fig. 3f), where AMS increased the Mg concentration by 55.6% and 134.7% in relation to CN and urea, respectively (Fig. 3f).
Tuber Yield and Agronomic Efficiency
Nonmarketable, marketable, and total potato tuber yields of any potato cultivars were not influenced by N sources or by the N source × N rate interactions (Table S3). However, the potato tuber yields of the Agata and Atlantic cultivars responded distinctly to the N rates applied with the sources tested (Fig. 4).

Total (a, b), marketable (c, d), and nonmarketable (d, e) potato tuber yields of cultivars Agata (a, c, e) and Atlantic (b, d, f), as affected by sidedressing nitrogen rates and sources. Vertical bars indicate the standard deviation of the mean. Ns, * and ** indicate nonsignificant and significant at 5% and 1% probability, respectively, considering the treatment without N in sidedressing, as a common treatment for all N sources.
 : urea;
: urea;
 : ammonium sulfate;
: ammonium sulfate;
 : calcium nitrate
: calcium nitrate
The total tuber yields of the Agata potato cultivar increased linearly with urea-N and quadratically with AMS-N and CN-N, resulting in maximum yields of 40.7, 35.5 and 34.2 Mg ha−1 at 210, 152, and 152 kg N ha−1, respectively (Fig. 4a). From these results, AE values of 194, 234, and 225 kg tubers kg−1 applied N were calculated. Similarly, the marketable tuber yields varied with respect to the total yields; linearly, for the urea-N rates; and quadratically, for the AMS-N and CN-N rates (Fig. 4c). Maximum marketable tuber yields of 30.1, 25.0, and 24.2 Mg ha−1 at respective rates of 210, 161, and 145 kg N ha−1 (Fig. 4a) were estimated from the equations. From these results, AE values of 143, 155, and 167 kg tubers kg−1 applied N were calculated. The yields of nonmarketable tubers increased linearly up to 14.5 Mg ha−1 with urea-N rates of 210 kg ha−1 N and did not vary (p > 0.05) with rates of N in the form of AMS and CN (Fig. 4e).
In the Atlantic experiment, the total tuber yield increased linearly with increasing AMS rate and quadratically with increasing CN rate, resulting in maximum yields of 35.7 and 38.2 Mg ha−1, respectively, at rates of 210 and 130 kg N ha−1 (Fig. 4b). From these situations, the AE values of 170 and 294 kg tubers kg−1 of applied N were estimated. Only the CN rates affected marketable tuber yields, which in a quadratic manner resulted in a maximum value of 34.8 Mg ha−1 with 129 kg N ha−1 (Fig. 4d) and in a respective AE of 270 kg tubers kg−1 applied N. There was no effect of rate for any N source on nonmarketable tuber yield (Fig. 4f). Atlantic tuber yields did not change linearly or quadratically as a function of the urea-N rate applied (Fig. 4b, d, f).
There were interactions between N sources and rates for AE (in terms of total and marketable tuber yields of Agata cultivar) (Table S3; Fig. 5a, c). Only with 70 kg N ha−1 there were differences between N sources for the AE for marketable (Fig. 5a). With this sidedressing N rate, AMS presented respectively AE’s 45.8% and 70.4% greater than that of urea, with no differences between these N sources and CN (Fig. 5a, c). This same N rate presented an AE that was 105.9% higher in terms of the Agata potato marketable tuber yield compared to the average of the other N rates (Table S3). For the Atlantic cultivar, the CN presented average AE values 118.8% and 101.1% higher in terms of the respective marketable (Fig. 5b) and total tuber yield (Fig. 5a) in relation to the means obtained with AMS and urea, which did not differ from each other.

Agronomic efficiency (AE) for marketable (a, b) and total (c, d) tuber yields of the Agata and Atlantic potato cultivars as affected by sidedressing nitrogen (N) rates and sources. Vertical error bars indicate the standard deviation of the mean. Uppercase letters compare N sources within each N rate (a, c) or according to average N rates (b, d) whereas lowercase letters compare N rates within each N source (a, c) at p < 0.05 according to Tukey’s test
Discussion
The results of this study reveal that the effects of the sources and rates of N applied during sidedressing on potato crops are highly dependent on the soil‒plant interface. The plant nutritional status, soil chemical attributes in preharvest ridges, and tuber yields varied systematically and interrelatedly but distinctly among the potato cultivars. The Agata and Atlantic cultivars, the main cultivars planted in Brazil for fresh market use and for frying in the form of chips, respectively (Fontes et al. 2010), have different nutrient requirements throughout their cycles, especially after the beginning of tuberization, highlighting the fact that the Atlantic cultivar has the highest amounts of N and basic cations compared to Agata (Fernandes et al. 2011; Silva et al. 2020). This characteristic becomes relevant because, in addition to N transformations in the soil, the export of basic cations are identified as the two main causes of acidification in subtropical soils (Dong et al. 2022).
Thus, the greater extractions of Ca, Mg, and K by the Atlantic cultivar and greater the application of AMS in conjunction with sidedressing may have been joint determinants of the decreases in pH (Fig. 3d) and base saturation (Fig. 3c) in the ridges, considering that these effects on soil chemical attributes were less intense or non-significant with rates of AMS applied as sidedressing for Agata potato (Table S2), except for the Al3+ concentration in the soil (Fig. 3a). Greater S extractions by the Atlantic cultivar in relation to Agata, as corroborated by work on absorption marches of these cultivars (Fernandes et al. 2011; Silva et al. 2020), also justify a greater effect of AMS rates on soil Al3+ solubilisation in the experiment with Agata potato, represented by the highest angular coefficient in the respective linear regression (Fig. 3a).
Soil acidification, particularly related to AMS application, can be attributed both to NH4+-N nitrification (Mizuno et al. 2000) and to the release of organic acids and protons in the root zone in response to NH4+-N uptake to maintain the internal ionic balance (Kissel et al. 2020). One should also consider the greater affinity of sulfate anions (SO42−-S) than of NO3−-N for basic cations in the formation of zero-charge ionic pairs susceptible to leaching (Foloni et al. 2010), in addition to the colloidal exchange complex in planting furrows. Furthermore, soil acidification under AMS-N rates in the Atlantic potato experiment may also have been responsible for the increases in leaf B and Mn concentrations, similar to what was observed by Maier et al. (2002).
Although soil acidification with sidedressing AMS was identified only in the experiment with the Atlantic cultivar (Table S2; Fig. 3c, d), the Al3+ concentration increased linearly with AMS-N rates during sidedressing in both experiments. However, it should be noted that these increases in Al3+ concentrations occurred even with the application of 889 kg ha−1 agricultural gypsum (data not shown) in the planting furrows together with phosphate fertilisation with single superphosphate mixed with NPK formula as base fertiliser. Consequently, the lower leaf Ca concentration in both potato cultivars and the lower Mg concentration in Atlantic plants with higher rates of AMS may be related to the greater amount of Al3+ ester provided by these treatments. Increased availability of Al3+ from the soil can also decrease the concentrations and accumulation of Ca and Mg in the shoots and roots of potato plants due to the competition of these cations for the same absorption sites in the roots (Lee 1971). Furthermore, NH4+-N uptake decreases Ca and Mg concentrations in the shoots of potato plants compared to NO3−-N nutrition (Polizotto et al. 1975; Davis et al. 1986; Cao and Tibbitts 1993) since the supply of NH4+-N inhibits the absorption of Ca and Mg by plants to maintain the internal ionic balance (Polizotto et al. 1975). The results obtained in the present study corroborate those obtained in a greenhouse by Maier et al. (2002), who also observed lower leaf concentrations of Ca and Mg and lower soil pH values after harvest in pot experiments with the Atlantic cultivar fertilised with AMS in relation to urea, ammonium nitrate, and CN. Souza et al. (2022) also observed lower concentrations of Mg in tubers of the Agata cultivar fertilised with AMS than in those fertilised with urea, both split at planting and as sidedressing.
Decreases in leaf Ca concentrations in both potato cultivars according to the increase in sidedressing N rates, which were opposite in Agata plants and less intense in the Atlantic cultivar when both were fertilised with CN (Figs. 1b and 2b), may easily be attributed to a dilution effect of N fertilisers. However, even at the highest N rate applied, the leaf Ca concentrations in both cultivars were above the lower limit of 7.5 g Ca kg−1, which is considered adequate (NEPAR-SBCS 2017). This difference between potato cultivars in the assimilation of Ca through the applied CN can be attributed to the lower accumulation of dry matter in the leaves of Agata plants than in those of Atlantic plants after the application of CN (Silva et al. 2013), justifying the absence of a dilution effect and increases in leaf Ca concentrations in the former. Furthermore, potato plants treated with NH4+-N have greater leaf area (Qiqige et al. 2017) and lower concentrations of Ca and Mg in their shoots (Polizotto et al. 1975; Davis et al. 1986; Cao and Tibbitts 1993, 1994) than do those treated with NO3N. This may also explain the more pronounced decreases in leaf concentrations or lower concentrations of Ca in both potato cultivars and Mg in the Atlantic cultivar with rates of AMS-N in relation to the other sources of N (Figs. 1b and 2b). Similar results to those of this study were obtained by Maier et al. (2002) in three experiments in a greenhouse, where CN application systematically provided higher leaf concentrations of Ca and Mg than urea, AMS, and ammonium nitrate in Russet Burbank and Atlantic cultivars.
Unlike leaf Ca concentrations, the dilution effect does not justify the absence of treatment effects on leaf N concentrations in the Agata cultivar, as this cultivar has leaves with lower dry matter production than Atlantic potato leaves (Fernandes et al. 2010; Silva et al. 2013). In the present experiment involving the Agata cultivar (Fig. 1a), only trends of linear increases in leaf N concentrations were detected urea-N (R2 = 0.77; p = 0.094) and AMS (R2 = 0.98; p = 0.138), as was a quadratic effect for CN-N rates (R2 = 0.99; p = 0.139) (data not shown). In this case, the assessment of the nutritional N status of the plants may have been hampered by the difference between the dates of emergence of the plants in the two experiments, by the difference in the volume of rainfall between the application of the treatments and the leaf sampling, and by the differences between the system roots of the studied cultivars. Leaf samplings were carried out 18 and 28 days after the application of N as a sidedressing in the respective experiments with the Agata and Atlantic cultivars. During the period from the application of treatments to the collection of diagnostic leaves in both experiments, 154 mm of rainfall was recorded, 47.5% (73 mm) of which occurred in intense events during the five days following the application of treatments. This rainfall regime at these different time intervals for the nutritional diagnosis of the plants, associated with the decreased accumulation of roots of the Agata cultivar in relation to that of the Atlantic potato (Fernandes et al. 2010; Silva et al. 2013), justifies both the absence of variations in leaf N concentrations in the Agata cultivar and the occurrence of more significant variations in nutrient concentrations in the experiment with the Atlantic cultivar. The absence of differences between N sources in the leaf N concentrations of the Agata cultivar was also attributed to the short time interval between top dressing and leaf sampling by Souza et al. (2019) in the evaluation of AMS, urea, and AMS with nitrification inhibitors. Except for the respective control treatments, the leaf N concentrations in both potato cultivars (Figs. 1a and 2a) are within the range considered adequate, from 40 to 60 g kg−1 (NEPAR-SBCS 2017). However, the maximum leaf N concentrations observed in this study for the Agata (44.0 g kg−1 with 210 kg urea-N ha−1) and Atlantic (45.8 g kg−1 210 kg CN-N ha−1) cultivars were lower than those observed in studies evaluating the application of N at planting in potato crops. For example, considering the maximum rate of urea applied at planting in the present study (210 kg N ha−1), leaf concentrations of 65.1 g N kg−1 were measured in Agata potato (Coelho et al. 2022) and 48.9 and 50.6 g N kg−1 were measured in the diagnostic leaves of Agata and Atlantic potatoes, respectively (Oliveira et al. 2020). Thus, later assessments of the nutritional diagnosis of N for potato plants, in addition to the recommended standard time for the crop, are necessary for more accurate assessments in studies on sidedressing N fertilisation for potato cultivars. The results obtained by Souza et al. (2019) corroborate this argument based on N concentrations in diagnostic leaves of the Agata cultivar in three experiments in three local years with collections 8, 10, and 20 days after the application of N. These authors measured leaf N concentrations of 45.3, 46.9, and 53.9 g kg−1 under AMS and of 43.9, 44.0, and 50.6 g kg−1 under urea. According to Fernandes et al. (2022), later sampling of diagnostic potato leaves after traditional sidedressing N fertilisation is feasible and necessary to maintain N availability throughout tuberization to obtain higher tuber yields.
The responses of tuber yield as a function of N source and rates in sidedressing were consistent with the interrelationships between plant nutritional diagnoses and soil chemical attributes, both of which are influenced by the different nutritional requirements of the studied potato cultivars. Despite being less demanding of macro- and micronutrients (Fernandes et al. 2011; Silva et al. 2020) and having a smaller root system than the Atlantic cultivar (Fernandes et al. 2010; Silva et al. 2013), the Agata cultivar has higher nutrient utilisation efficiency, i.e. it produces more fresh tubers with less absorbed nutrients (Fernandes and Soratto 2013). This is probably the justification for this potato cultivar to have shown a greater AE (Fig. 5a, b), in addition to greater responses and magnitudes to the rates of N sources in sidedressing in relation to the Atlantic potato (Fig. 4). For example, the maximum increases in the total tuber yields of Agata and Atlantic of 67.5% (14.3 Mg ha−1) and 55.9% (12.8 Mg ha−1) with AMS and of 69.3% (14.0 Mg ha−1) and 36.9% (10.3 Mg ha−1), respectively, with CN.
The effect of sidedressing N fertilisation on the Agata potato crop was dependent on the N source and the applied rate (Fig. S3). With higher AEs in terms of marketable tuber yields and total tuber yields, 70 kg N ha−1 applied as AMS was advantageous, and between 140 and 210 kg N ha−1, the total yields or marketable tuber yields and their respective AEs were similar between N sources (Fig. 5a, c). However, with the application of 210 kg N ha−1 urea, there was still an increase in tuber productivity, which did not occur with AMS or CN at this same rate (Fig. 4a). With this rate and source of N, there were increases in total productivity, of 14.7% (5.2 Mg ha−1) and 19.1% (6.5 Mg ha−1), and in marketable tubers, of 20.5% (5.1 Mg ha−1) and 24.2% (5.9 Mg ha−1), compared to the respective maximum tuber yields obtained with AMS and CN.
Possible increases in the availability of NH4+-N at the beginning of tuberization and of NO3−-N at the end of the cycle with the application of a high rate of urea-N in sidedressing may have provided the respective filling and late initiation of tubers of the Agata cultivar. Regarding the application of urea in sidedressing, 137.2% and 91.2% increases were observed in marketable (Fig. 4b) and nonmarketable tuber yields (Fig. 4e), respectively, with 210 kg N ha−1. The hydrolysis of urea to NH4+-N occurs quickly within 5 days, while most nitrification occurs 28 to 56 days after the addition of urea depending on temperature (MacLean and McRae 1987). The release of NH4+-N or ammonia (NH3) occurs with the application of urea-N according to pH conditions and alters the dry matter partitioning of the Agata and Atlantic cultivars in a hydroponic system, stimulating vegetative growth to the detriment of tuberization (Silva et al. 2013). The Agata cultivar matures early to very early (ECPD 2023), and fertilisation with NH4+-N at the beginning of tuberization causes the plants to develop tubers early (Qiqige et al. 2017). In response, N nutrition must be precise and suitable because, while the supply of NO3−-N favours the growth of shoots and the initiation of stolons and stems, the supply of NH4+-N stimulates tuber filling (Osaki et al. 1995a) but can also be toxic to potato plants (Cao and Tibbits 1998) or harmful to plant growth at any stage of development at excessive concentrations (Polizotto et al. 1975; Davis et al. 1986).
In the experiment with the Atlantic cultivar, the application of CN-N was the best option for sidedressing N fertilisation because it resulted in more than twice the AE in marketable and total tuber yields in relation to the other N sources due to the deleterious effects of AMS-N at the soil‒plant interface and the absence of responses in tuber yields with urea-N rates (Fig. 5b, d).
A supply of NO3−-N with CN-N requires a greater energy cost for reducing NO3−-N in plants and decreasing shoot growth, contrary to what occurs with urea-N (Silva et al. 2013). After absorption, NO3−-N is initially distributed to leaves, where it is transformed into soluble amino acids, mainly asparagine, aspartic acid, glutamine, and glutamic acid, which are subsequently stored in petioles and stems and subsequently increase the number of branches and stolons and tuber growth (Osaki et al. 1995b) and the number of tubers per plant (Qiqige et al. 2017; Gao et al. 2014). Bundy et al. (1986) observed the superiority of AMS for total tuber yield in 2 out of 5 years of study and for the average of these 5 years in relation to urea, CN, and ammonium nitrate. According to these same authors, the leaching of NO3—N with CN, or loss of nonhydrolysed urea or NO3−-N by leaching due to faster nitrification with urea than with AMS, or loss of NH3 with urea, which are all associated with the superficial rooting of the plants, justified the obtained results.
Conclusions
This study demonstrated that the effects of sidedressing N sources and rates on potato crops under subtropical conditions were soil‒plant interface dependent and differed between potato cultivars due to their specific nutritional requirements. For Agata potato cultivation, the best source of N was dependent on the application rate. Compared with urea, only the sidedressing AMS treatment showed higher AE for total tuber yields, with yields up to 70 kg N ha−1 and a maximum technical efficiency of 152 kg N ha−1. However, increases in marketable and nonmarketable tuber yields were still obtained with up to 210 kg urea-N ha−1, indicating that this source and rate of N are indicated for the Agata cultivar. For the Atlantic cultivar, there was a lack of response in tuber yield with urea-N, and sidedressed AMS-N was shown to be harmful for increasing the exchangeable acidity of the soil, consequently worsening the plant Ca and Mg status. In contrast, CN did not negatively affect soil chemical attributes; favoured plant nutrition in terms of Ca and Mg; and provided the highest AE in total and marketable tuber yields, which reached a maximum at 130 kg CN-N ha−1, indicating that this N source and rate are suitable for the Atlantic cultivar. Our results provide insights that more nutritionally demanding cultivars interact negatively and synergistically with acidifying N fertilisers in soils with the potential availability of toxic Al, impairing Ca and Mg nutrition, and the tuber bulking.
Data Availability
The datasets analysed during the current study are available from the corresponding author on reasonable request.
References
Barbosa JC, Maldonado Junior W (2015). AgroEstat: System for Statistical Analysis of Agronomic Trials. Jaboticabal, Brazil (in Portuguese)
Bender FD, Sentelhas PC (2020) Assessment of regional climate change impacts on Brazilian potato tuber yield. Int J Plant Prod 14:647–661. https://doi.org/10.1007/s42106-020-00111-7
Bundy LG, Wolkowski RP, Weis GG (1986) Nitrogen source evaluation for potato production on irrigated sandy soils. Am Potato J 63:385–397. https://doi.org/10.1007/BF02854101
Cao W, Tibbitts TW (1993) Study of various NH4+/NO3- mixtures for enhancing growth of potatoes. J Plant Nutr 16:1691–1704. https://doi.org/10.1080/01904169309364643
Cao W, Tibbitts TW (1994) Responses of potatoes to solution pH levels with different forms of nitrogen. J Plant Nutr 17:109–126. https://doi.org/10.1080/01904169409364713
Cao W, Tibbitts TW (1998) Response of potatoes to nitrogen concentrations differ with nitrogen forms. J Plant Nutr 21:615–623. https://doi.org/10.1080/01904169809365429
Coelho ARF, Marques AC, Pessoa CC, Daccak D, Luís IC, Silva MM, Simões M, Reboredo FH, Pessoa MF, Legoinha P, Ramalho JC, Campos OS, Pais IP, Semedo JN, Lidon FC (2022) Natural enrichment of solanum tuberosum l with calcium—monitorization of mineral interactions in plant tissues. Biol Life Sci Forum 11:28. https://doi.org/10.3390/IECPS2021-11972
Davis JM, Loescher WH, Hammond MW, Thorton RE (1986) Response of potatoes to nitrogen form and to change in nitrogen form at tuber initiation. J Amer Soc Hort Sci 111:70–72. https://doi.org/10.21273/JASHS.111.1.70
Dong Y, Yang J, Zhao X, Yang S, Mulder J, Dörsch P, Peng X, Zhang G (2022) Soil acidification and loss of base cations in a subtropical agricultural watershed. Sci Total Environ 2022(827):154338. https://doi.org/10.1016/j.scitotenv.2022.154338
Dongyu Q (2022) Role and potential of potato in global food security. In: 11th World Potato Congress 2022 May 30 - Jun 2 Dublin, Leinster. https://www.fao.org/3/cc0330en/cc0330en.pdf. Accessed in 2023.
ECPD (2023) Agata. The European cultivated potato database, Edinburgh, Scotland. European Coop. Programme for Plant Genetic Resources. https://www.europotato.org/varieties/. Accessed 20 July 2023.
Embrapa (1997) Centro Nacional de Pesquisa de Solos. Soil Analysis Methods Handbook (In Portuguese), 2nd edn. Embrapa, Rio de Janeiro, Brazil. https://ainfo.cnptia.embrapa.br/digital/bitstream/item/169149/1/Manual-de-metodos-de-analise-de-solo-2-ed-1997.pdf. Accessed in 2023.
FAOSTAT (2023) Statistics Division. Food and Agriculture Organization of the United Nations. http://www.fao.org/faostat/en/#home. Accessed in 2023.
Fernandes AM, Soratto RP (2013) Absorption and nutrients use efficiency by potato cultivars. Biosci J 29:91–100
Fernandes AM, Soratto RP, Silva BL, Souza-Schlick GD (2010) Growth and dry matter accumulation and distribution in potato cultivars during the winter crop season. (In Portuguese, with English abstract.). Pesq Agropec Bras 45:826–835. https://doi.org/10.1590/S0100-204X2010000800008
Fernandes AM, Soratto RP, Silva BL (2011) Nutrient extraction and exportation by potato cultivars: I - macronutrients. (In Portuguese, with English abstract). R Bras Ci Solo 35:2039–2056. https://doi.org/10.1590/S0100-06832011000600020
Fernandes FM, Soratto RP, Fernandes AM, Souza EFC (2021) Chlorophyll meter-based leaf nitrogen status to manage nitrogen in tropical potato production. Agron J 113:1733–1746. https://doi.org/10.1002/agj2.20589
Fernandes FM, Soratto RP, Sandaña P, Souza EFC, Yagi R (2022) Optimization of leaf nitrogen concentration and monitoring time for sustainable potato production in tropics. J Soil Sci Plant Nutr 22:2765–2775. https://doi.org/10.1007/s42729-022-00843-7
Finkelman JM, Wolf EH, Friend MA (1977) Polynomial regression analysis as an alternative to ANOVA for data characterized by lower-order trends. Hum Factors 19:279–281. https://doi.org/10.1177/001872087701900310
Foloni JSS, Rosolem CA, Garcia RA (2010) Effect of liming and ammonium sulfate in cotton: II - Concentration of cations and anions in the soil solution and plant nutrient uptake. (In Portuguese, with English abstract). R Bras Ci Solo 2006(30):433–442. https://doi.org/10.1590/S0100-06832006000300005
Fontes PCR, Braun H, Busato C, Cecon PR (2010) Economic optimum nitrogen fertilization rates and nitrogen fertilization rate effects on tuber characteristics of potato cultivars. Potato Res 53:167–179. https://doi.org/10.1007/s11540-010-9160-3
Gao Y, Jia L, Hu B, Alva A, Fan M (2014) Potato stolon and tuber growth influenced by nitrogen form. Plant Prod Sci 17:138–143. https://doi.org/10.1626/pps.17.138
Giletto CM, Kloster Erreguerrena M, Ceroli P, Carciochi W, Silva SE, Rodriguez S, Salvagiotti F, Reussi Calvo NI (2022) Holistic assessment of calcium fertilization in potato: diagnostic, productivity, and tuber quality. J Soil Sci Plant Nutr. https://doi.org/10.1007/s42729-022-01060-y
Haverkort AJ, Struik PC (2015) Yield levels of potato crops: recent achievements and future prospects. Field Crops Res 182:76–85. https://doi.org/10.1016/j.fcr.2015.06.002
Hijmans RJ (2003) The effect of climate change on global potato production. Amer J Potato Res 80:271–279. https://doi.org/10.1007/BF02855363
Kissel DE, Bock BR, Ogles CZ (2020) Thoughts on acidification of soils by nitrogen and sulfur fertilizers. Agrosyst Geosci Environ 3:e20060. https://doi.org/10.1002/agg2.20060
Kleinwechter U, Gastelo M, Ritchie J, Nelson G, Asseng S (2016) Simulating cultivar variations in potato yields for contrasting environments. Agric Syst 145:51–63. https://doi.org/10.1016/j.agsy.2016.02.011
Lee CR (1971) Influence of aluminum on plant growth and mineral nutrition of potatoes1. Agron J 63:604–608. https://doi.org/10.2134/agronj1971.00021962006300040027x
Liu KL, Du JX, Zhong YJ, Shen Z, Yu XC (2021) The response of potato tuber yield, nitrogen uptake, soil nitrate nitrogen to different nitrogen rates in red soil. Sci Rep 11:1–10. https://doi.org/10.1038/s41598-021-02086-5
Lizana CX, Sandaña P, Behn A, Ávila-Valdés A, Ramírez DA, Soratto RP, Campos H (2020) Potato. In: Sadras VO, Calderini DF (eds) Crop physiology: case histories for major crops. Academic Press, San Diego, USA, pp 551–587
Lopes CA, Reis A, Lima MF (2016) Diseases and control methods. In: Silva GO, Lopes CA (eds) Potato production system. (In Portuguese). Embrapa Hortaliças, Brasília, DF, Brazil. https://ainfo.cnptia.embrapa.br/digital/bitstream/item/132923/1/Sistema-de-Producao-da-Batata.pdf. Assessed in 2023.
MacLean AA, McRae KB (1987) Rate of hydrolysis and nitrification of urea and implications of its use in potato production. Can J Soil Sci 67:679–686. https://doi.org/10.1038/srep09697
Maier NA, McLaughlin MJ, Heap M, Butt M, Smart MK (2002) Effect of nitrogen source and calcitic lime on soil pH and potato yield, leaf chemical composition, and tuber cadmium concentrations. J Plant Nutr 25:523–544. https://doi.org/10.1081/PLN-120003380
Makani MN, Zotarelli L, Sargent SA, Huber DJ, Sims CA (2020) Nitrogen fertilizer rate affects yield and tuber quality of drip-irrigated tablestock potatoes (Solanum tuberosum L.) grown under subtropical conditions. Am J Potato Res 97:605–614. https://doi.org/10.1007/s12230-020-09809-w
Malavolta E, Vitti GC, Oliveira AS (1997) Evaluation of the nutritional status of plants: principles and applications (In Portuguese). Piracicaba, SP, Brazil
Mizuno N, Yoshida H, Tadano T (2000) Efficacy of single application ammonium sulfate in suppressing potato common scab. Soil Sci Plant Nutr 46(3):611–616. https://doi.org/10.1080/00380768.2000.10409126
NEPAR-SBCS (2017) Handbook of fertilization and liming for the State of Parana (In Portuguese). Curitiba, PR, Brazil
Oliveira RC, Luz J, Queiroz M, Lana R, Quintao M, Silva JRRD, Castoldi R (2020) Yield of potato cultivars as a function of nitrogen rates. Rev Caatinga 33:954–963. https://doi.org/10.1590/1983-21252020v33n410rc
Osaki L, Shirai J, Shinano T, Tadano T (1995a) 15N-allocation of 15NH4-N and 15NO3-N to nitrogenous compounds at the vegetative growth stage of potato plants. Soil Sci Plant Nutr 41(4):699–708. https://doi.org/10.1080/00380768.1995.10417020
Osaki M, Shirai J, Shinano T, Tadano T (1995b) Effects of ammonium and nitrate assimilation on the growth and tuber swelling of potato plants. Soil Sci Plant Nutr 41:709–719. https://doi.org/10.1080/00380768.1995.10417021
Pavan MA, Bloch MFD, Zempulski HCD, Miyazawa M, Zocoler DC (1992) Manual of soil chemical analysis and quality control. Instituto Agronômico do Paraná, Londrina, PR, Brazil (in Portuguese)
Piepho HP, Edmondson RN (2018) A tutorial on the statistical analysis of factorial experiments with qualitative and quantitative treatment factor levels. J Agron Crop Sci 204:429–455. https://doi.org/10.1111/jac.12267
Polizotto KR, Wilcox GE, Jones CM (1975) Response of growth and mineral composition of potato to nitrate and ammonium nitrogen. J Am Soc Hort Sci 100:165–168. https://doi.org/10.21273/JASHS.100.2.165
Qiqige S, Jia L, Qin Y, Chen Y, Fan M (2017) Effects of different nitrogen forms on potato growth and development. J Plant Nutr 40:1651–1659. https://doi.org/10.1080/01904167.2016.1269345
Rens LR, Zotarelli L, Cantliffe DJ, Stoffella PJ, Gergela D, Burhans D (2016) Marketable evaluation of seasonal distribution of nitrogen fertilizer for potato. Potato Res 59:1–20. https://doi.org/10.1007/s11540-015-9304-6
Serio F, Elia A, Signore A, Santamaria P (2004) Influence of nitrogen form on yield and nitrate content of subirrigated early potato. J Sci Food Agric 84:1428–1432. https://doi.org/10.1002/jsfa.1835
Silva JG, França MGC, Gomide FTF, Magalhaes JR (2013) Different nitrogen sources affect biomass partitioning and quality of potato production in a hydroponic system. Am J Potato Res 90:179–185. https://doi.org/10.1007/s12230-012-9297-5
Silva CD, Soares MEP, Ferreira MH, Cavalcante ACP, Andrade GAV, Aquino LA (2020) Dry matter and macronutrient extraction curves of potato varieties in the Alto Paranaíba region, Brazil. Rev Bras Eng Agrícola Ambient 24:176–186. https://doi.org/10.1590/1807-1929/agriambi.v24n3p176-186
Simmons KE, Kelling KA (1987) Potato Responses to Calcium Application on Several Soil Types Amer Potato J 64:119–136. https://doi.org/10.1007/BF02854208
Soil Survey Staff (2022) Keys to Soil Taxonomy. USDA-Natural Resources Conservation Service, 13th edn. https://www.nrcs.usda.gov/sites/default/files/2022-09/Keys-to-Soil-Taxonomy.pdf. Assessed in 2024.
Soratto RP, Silva GO, Fernandes AM (2016) Fertilization. In: Silva GO, Lopes C (eds) Potato Production System. (In Portuguese). Embrapa Hortaliças, Brasília, DF, Brazil. https://ainfo.cnptia.embrapa.br/digital/bitstream/item/132923/1/Sistema-de-Producao-da-Batata.pdf. Accessed in 2023.
Souza EFC, Soratto RP, Fernandes AM, Rosen CJ (2019) Nitrogen source and rate effects on irrigated potato in tropical sandy soils. Agron J 111:378–389. https://doi.org/10.2134/agronj2018.03.0198
Souza EFC, Soratto RP, Sandaña P, Venterea RT, Rosen CJ (2020) Split application of stabilized ammonium nitrate improved potato yield and nitrogen-use efficiency with reduced application rate in tropical sandy soils. Field Crops Res 254:107847. https://doi.org/10.1016/j.fcr.2020.107847
Souza EF, Soratto RP, Fernandes AM, Gupta SK (2022) Performance of conventional and enhanced-efficiency nitrogen fertilizers on potato tuber mineral composition and marketability. J Sci Food Agric 102(8):3078–3087. https://doi.org/10.1002/jsfa.11648
Yagi R, Nazareno NRX, Soratto RP (2020) Agronomic and economic interactions between sidedressed nitrogen and potassium fertilizations on ‘Atlantic’ potato. Braz Arch Biol Technol 63:1–11. https://doi.org/10.1590/1678-4324-2020190640
Zebarth BJ, Milburn PH (2003) Spatial and temporal distribution of soil inorganic nitrogen concentration in potato hills. Can J Soil Sci 83:183–195. https://doi.org/10.4141/S02-061
Zotarelli L, Rens LR, Cantliffe DJ, Stoffella PJ, Gergela DM, Fourman D (2014) Nitrogen fertilizer rate and application timing for chipping potato Atlantic. Agron J 106:2215–2226. https://doi.org/10.2134/agronj14.0193
Acknowledgements
We are thankful to the potato grower (Alexandre Dzierwa), who provided the area for this investigation.
Funding
We thank the National Council for Scientific and Technological Development (CNPq) for granting awards for excellence in research to the third author. The Yara International ASA partially funded this research.
Author information
Authors and Affiliations
Contributions
Conceptualization, methodology, and data curation: R.Y., B.B., and D.S.A. Formal analysis and investigation: R.Y. and R.P.S. Writing, original draft: R.Y. Writing, review and editing: R.Y., B.B., R.P.S., T.R.S.C., and D.S.A. Supervision: R.Y.
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Yagi, R., Bagio, B., Soratto, R.P. et al. Impacts of Nitrogen Sources and Rates on Soil Chemical Attributes and Potato Nutrition and Yield in a Subtropical Environment. Potato Res. (2024). https://doi.org/10.1007/s11540-024-09714-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11540-024-09714-3