
Abstract
Pectobacterium and Dickeya species are pathogens of increasing importance on potatoes worldwide. These Soft Rotting Pectobacteriaceae (SRP) cause soft rot, blackleg, and aerial stem rot of tubers and plants, which can lead to extensive yield losses under conditions favourable for disease development. High priority is placed on continual, regular surveillance of Pectobacterium and Dickeya populations internationally, to ensure that new pathogens are not introduced into potato production areas. A survey was therefore undertaken to identify the primary SRP on potatoes in South Africa. Representative isolates were tested for pathogenicity and symptom expression, singly and in combination, in tuber slice assays and pot trials. Results showed that Pectobacterium brasiliense (Pb) remains the dominant and most pathogenic species in the country. The presence of Pectobacterium carotovorum (Pc) and Pectobacterium parmentieri was confirmed, although these species were far less common than Pb. Pectobacterium versatile (Pv) was reported for the first time on potatoes in South Africa. Pv caused soft rot in tuber slice assays and blackleg symptoms in pathogenicity pot trials, albeit at low incidences. Some strains identified as Pc were recently reclassified as a novel species, Pv. Pectobacterium carotovorum is more often associated with aerial stem rot than blackleg, which might explain low blackleg incidence in the trials, as well as the aerial stem rot symptoms from which the initial isolations were made. The lack of detection of Dickeya solani in any of the samples suggests that it remains absent from the country.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The blackleg-soft rot-aerial stem rot disease complex is a serious threat to potato production across South Africa (SA) and is set to become even worse in the foreseeable future (van der Waals et al. 2013). Tuber soft rot, which occurs in the field or storage, is characterised by macerated tuber tissue of a creamy consistency that darkens with time and develops a putrid odour. Blackleg is characterised by slimy black stem (haulm) lesions that start at the stem base and rapidly spread upwards to consume the entire stem, resulting in haulm desiccation. It is associated with wilting and chlorosis and is often limited to only one of the stems. Blackleg can result in damping-off and stunting when occurring early in the season (Hélias et al. 2000; Pérombelon 2002; Kumvinit and Akarapisan 2019). Aerial stem rot is characterised by slimy brown stem lesions that originate on aerial stem parts and lead to haulm desiccation (De Boer et al. 2012).
The disease complex is caused by species of the Pectobacterium and Dickeya genera, collectively known as the Soft Rot Pectobacteriaceae (SRP) (Kabir et al. 2020). The highly pathogenic Pectobacterium brasiliense (Pb) is regarded as the predominant species in the country since its first report in 2010 (van der Merwe et al. 2010). Other species also found in SA are Pectobacterium carotovorum (Pc), Pectobacterium parmentieri (Pp), and two Dickeya species: Dickeya zeae and Dickeya dianthicola (van der Merwe et al. 2010; Ngadze et al. 2012; Moleleki et al. 2013). An important species that has not yet been found in SA is Dickeya solani. It is highly pathogenic, particularly in warm climates (Toth et al. 2011).
Differences in pathogenicity among the species are widely reported in the literature, often with contrasting findings (Del Pilar Marquez-Villavicencio et al. 2011; De Boer et al. 2012; van der Wolf et al. 2017). Differences in symptom appearance are less apparent (Czajkowski et al. 2013), although atypical symptoms have been reported for Dickeya species, consisting of rotten upper stems, petioles, and leaves, while the stem base appeared healthy but had a rotten pith (Laurila et al. 2010). It is also believed that Pc seldom causes blackleg but aerial stem rot instead (Pérombelon 2002; De Boer et al. 2012; Charkowski 2018). Co-infections with multiple species can also occur and influence the incidence and severity of symptoms (De Boer et al. 2012). Both synergistic and antagonistic effects have been reported (van der Wolf et al. 2017). In such co-infections, factors such as the prevailing temperatures and initial inoculum levels influence which taxa predominate (Czajkowski et al. 2011).
The various Pectobacterium and Dickeya species have different temperature preferences. Pectobacterium brasiliense has an optimum and growth range of 31–32 °C and 20–39 °C, respectively. This is similar to Pc, with a range of 20–40 °C and an optimum of 31–34 °C, while Pp has a range of 20–34 °C and an optimum of 29 °C. Pectobacterium atrosepticum prefers cooler temperatures, with a range of 18–31 °C and an optimum of 26–27 °C (du Raan et al. 2016). Conversely, Dickeya species prefer warm conditions, with Dickeya dadantii having a temperature range and optimum of 20–39 °C and 31–34 °C, respectively, and D. solani, 24–42 °C and 35 °C, respectively (du Raan et al. 2016). The warm South African climate, which is optimal for the highly pathogenic Pb, might explain the predominance of this species in the country (van der Merwe et al. 2010; van der Waals et al. 2013). Considering the highly aggressive nature of D. solani under warm conditions (Tsror et al. 2013), it is likely to have a devastating impact on potato production if introduced into the country.
The SRP are mainly spread by latently infected seed tubers (Czajkowski et al. 2011). Pb is proposed to have entered the country in this manner (van der Merwe et al. 2010). The pathogen remains latent in the tuber, even at high inoculum levels, and certain conditions need to be met for bacterial multiplication and symptom expression to occur. The most important of these is the presence of water on the tuber (Pérombelon 2002; Czajkowski et al. 2011). This causes anaerobic conditions that inhibit host defences and stimulate bacterial multiplication. Once a critical cell density is reached, acyl-homoserine lactone (AHL)-mediated quorum sensing stimulates the production of pectinolytic enzymes, causing tissue maceration and symptom appearance (Pérombelon 2002; van der Merwe et al. 2010). Soft rot can progress rapidly under favourable conditions, consuming entire tubers within 2 to 3 days (Charkowski 2018). This can result in non-emergence (blanking), the incidence of which increases with higher inoculum concentrations (Bain et al. 1990). Blackleg does not develop in the absence of soft rot of the mother tuber (Pérombelon 2002). The higher the initial inoculum, the higher the incidence and the faster the onset of symptoms which, under favourable conditions, can kill a plant within hours of appearing (Charkowski 2018).
Management of this disease complex relies primarily on the production of certified, disease-free seed tubers (Czajkowski et al. 2011), due to a lack of resistant commercially produced cultivars (Ngadze and van der Waals 2012; Kubheka et al. 2013) and few effective chemical treatments (Czajkowski et al. 2013; Charkowski 2018). Certification involves field and postharvest inspections, testing, and roguing of infected plants and tubers (Agrios 2005).
Various approaches exist for detection and identification of SRP. Isolation on the semi-selective medium, crystal violet pectate (CVP) agar, is used to differentiate SRP from other species based on the characteristic pits that they form on the agar (Hélias et al. 2012). For more specific identification, polymerase chain reaction (PCR) protocols have been developed for most species. A multiplex protocol, developed by Potrykus et al. (2014) is successful at identifying Dickeya spp., Pa, Pc, and Pp, although it cannot differentiate between the latter two species. PCR protocols have also been developed for Pp (Kim et al. 2012) and Pb (Duarte et al. 2004). Recently, Portier et al. (2019) used multi-locus sequence analysis (MLSA) of the concatenated sequences of the dnaX-leuS-recA housekeeping genes to construct phylogenies of the SRP taxa. This resulted in the elevation of the sub-species of P. carotovorum to species level and the proposal of new species. The newly proposed species of Pectobacterium polaris, Pectobacterium odoriferum, Pectobacterium actinidiae, and Pectobacterium versatile (Pv) were distinguished from the newly emended description of Pectobacterium carotovorum. A gapA PCR-sequencing assay has also been developed that rapidly and successfully differentiates between most of the known SRP taxa (Cigna et al. 2017).
The introduction of new species into SA and the taxonomic reclassification of existing ones warrant a revision of the current SRP species composition in the potato industry. Certification and diagnostic services require updated information in order to test for the correct species and make accurate diagnoses. Knowledge regarding differences in symptom appearance between the species might allow for the early detection of newly introduced species during field inspections by certification services. Furthermore, as none of the SRP species are on the list of prohibited organisms, as defined in the Agricultural Pests Act (Act No. 36 of 1983) (Department of Agriculture 2013), there is no barrier to their entry into the country. Therefore, the highly pathogenic D. solani might already be present. Accordingly, a survey was conducted to determine the current SRP species composition in the potato industry in South Africa and, subsequently, to look for differences in symptom expression among these species. Koch’s postulates were carried out on species identified for the first time from potatoes in South Africa.
Materials and Methods
SRP Survey
Sample Collection
Throughout 2020 and 2021, samples of diseased tubers and haulms were collected from 10 of the 16 potato production regions in SA, namely, North West, Northern Cape, Sandveld, South Western Free State, Eastern Free State, Mpumalanga, Limpopo, Western Free State, Gauteng, and Marble Hall. Primary isolations were made on CVP single-layer agar (Hélias et al. 2012), containing Dipecta AG 366 pectin (BIOCOM Africa). The presence of characteristic pits after 24 to 48 h of incubation at 28 °C constituted a positive identification of SRP. In the absence of pits, the plates were further incubated at 25 °C for 24 to 48 h, to screen for species with lower cardinal temperatures. Individual pits were then sub-cultured onto nutrient agar (NA) to obtain pure colonies. The NA plates were incubated at 28 °C for 48 h before pure colonies were big enough for DNA extraction and storage. All colonies used for DNA extraction were also stored by suspending part of the colony in a sterile 80:20 (v/v) nutrient broth (NB)–glycerol solution and placing in a − 40 °C freezer.
Isolate Identification
DNA extractions were performed with the Zymo Research Quick-DNA™ Fungal/Bacterial Miniprep Kit (Zymo Research) according to the manufacturer’s protocol. The extracted DNA samples were stored at − 20 °C until used for PCR. The PCRs were performed using the 2720 Thermal Cycler from Applied Biosystems (Thermo Fisher Scientific Corp). Each DNA sample was subjected to two PCR reactions: the first, for the detection of Pb, and the second, a multiplex, for the detection of Pa, Pc/Pp, and Dickeya species. Taxon-specific primers that were used were BR1f (5′ GCG TGC CGG GTT TAT GAC CT 3′) and L1r (5′ CAA GGC ATC CAC CGT 3′) for Pb (Duarte et al. 2004), Y45 (5′ TCA CCG GAC GCC GAA CTG TGG CGT 3′) and Y46 (5′ TCG CCA ACG TTC AGC AGA ACA AGT 3′) for Pa, Df (5′ AGA GTC AAA AGC GTC TTG 3′) and Dr (5′ TTT CAC CCA CCG TCA GTC 3′) for Dickeya species, and ExPcF (5′ GAA CTT CGC ACC GCC GAC CTT CTA 3′) and ExPcR (5′ GCC GTA ATT GCC TAC CTG CTT AAG 3′) for Pc and Pp (Potrykus et al. 2014). The Pb PCR was performed in a reaction volume of 25 µl, containing 2 µl of DNA template (0.5 to 500 ng), 1 × OneTaq®Standard Reaction Buffer, 200 µM mixed dNTP solution, 0.625 U of OneTaq®DNA Polymerase (New England Biolabs), and primer concentrations of 0.2 µM for BR1f and L1r. The PCR was run according to the following thermal regime: denaturation at 94 °C for 2 min followed by 25 cycles of denaturation (94 °C for 45 s), annealing (62 °C for 45 s), extension (72 °C for 90 s), and a final, single extension at 72 °C for 10 min (Duarte et al. 2004). The multiplex PCR was performed in a reaction volume of 25 µl, containing 2 µl of DNA template (0.5 to 500 ng), 1 × OneTaq®Standard Reaction Buffer, 80 µM of each of the four dNTPs, and 1 U of OneTaq®DNA Polymerase. The primer concentrations were 1.2 µM each of ExPcF and ExPcR, 0.1 µM each of Y45 and Y46, and 0.32 µM each of Df and Dr. The PCR was run according to the following thermal regime: an initial denaturation at 95 °C for 4 min, followed by 30 cycles of denaturation (94 °C for 45 s), annealing (62 °C for 90 s), extension (72 °C for 90 s), and a final single extension at 72 °C for 3 min (Potrykus et al. 2014). Following the PCR, the amplified DNA was stored at 4 °C, awaiting gel electrophoresis.
The PCR products were separated on 1% (w/v) agarose gel (LONZA) in 1 × TAE buffer (40 mM Tris–acetate, 1 mM EDTA), containing 0.05 µl ml−1 Roti®GelStain and run at 140 V and 250 mA for 50 min. The bands were visualised with a UV transilluminator (Bio-Rad Laboratories). The presence of band sizes of 420 bp (Pa), 130 bp (Dickeya species), and 550 bp (Pc/Pp) in the multiplex (Potrykus et al. 2014) and 322 bp in the singleplex for Pb (Duarte et al. 2004) indicated successful identification of the respective species. Positive controls for Pb, Pc, and Pa were obtained from the Bacterial Culture Collection (BCC) of the Centre for Microbial Ecology and Genomics (CMEG) at the University of Pretoria (Suppl. Table 1). The identified isolates were stored in a bacterial freezing medium (OPS Diagnostics) at − 40 °C.
The DNA samples that produced amplicons in the Pb and multiplex PCRs were subjected to a PCR targeting the gapA gene, using the primers gapA-7-F (5′ ATC AAA GTA GGT ATC AAC GG 3′) and gapA-938-R (5′ TCR TAC CAR GAA ACC AGT T 3′) (Cigna et al. 2017). The PCR was performed in a reaction volume of 25 µl, containing 2 µl of DNA template (0.5 to 500 ng), 1X OneTaq®Standard Reaction Buffer, 200 µM mixed dNTP solution, 0.625 U of OneTaq®DNA Polymerase (New England Biolabs), and primer concentrations of 0.2 µM for both gapA-7-F and gapA-938-R. The PCR was run according to the following thermal regime: denaturation at 94 °C for 5 min followed by 40 cycles of denaturation (94 °C for 30 s), annealing (54 °C for 30 s), extension (72 °C for 30 s), and finally, single extension at 72 °C for 5 min (Cigna et al. 2017). Gel electrophoresis was performed exactly as for the previous PCRs. The presence of a 932-bp amplicon indicated successful amplification (Cigna et al. 2017). A PCR clean-up was then performed on the amplified DNA, using ExoSAP (Exonuclease I and FastAP) (Thermo Fisher Scientific). The cleaned PCR reaction mixtures were sequenced in both directions, using the same gapA PCR primers and the BigDye Terminator v.3.1. Cycle Sequencing Kit (Applied Biosystems). The sequencing PCR reaction mixture consisted of 4 µl cleaned PCR product, 2.5 µl of 5 × Sequencing Buffer (Applied Biosystems), 0.5 µl of v3.1 ABI PRISM® BigDye Terminator (Applied Biosystems), 0.5 µl of 10 µM forward or reverse primer, and 4.5 µl of nuclease-free water (nfH2O) (Qiagen), adding up to a final volume of 12 µl. The sequencing PCR was run according to the following thermal regime: initial denaturation at 96 °C for 30 s, followed by 24 cycles of denaturation (96 °C for 10 s), annealing (55 °C for 5 s), and elongation (60 °C for 4 min). The sequencing PCR product was precipitated with 3 M sodium acetate (NaOAc) (pH 5.2) and ethanol (Merck). The purified DNA was stored at − 20 °C before being submitted to the DNA Sanger sequencing facility at the University of Pretoria for sequencing by the ABI 3500xl genetic analyser (Applied Biosystems). The BioEdit Sequence Alignment Editor v 7.2.5 (Hall 1999) was used to trim and construct consensus sequences. The resulting sequences were then compared to those in the GenBank nucleotide sequence database, using the Basic Local Alignment Search Tool (BLASTn), and the species identified (Altschul et al. 1990).
The identity of isolates, SRP 1, SRP 2, SRP 3, SRP 4, and SRP 6, were further confirmed by MLSA, as, apart from SRP 1 and SRP 4, their closest BLASTn matches were previously unreported species in the country (Table 1). The inclusion of SRP 1 was due to its role as a positive control in subsequent pathogenicity trials. Although SRP 4 was identified as Pc by BLASTn, its inclusion in the MLSA was due to its previous identification as P. actinidiae in an unrelated gapA phylogeny. Three housekeeping genes, namely, atpD, dnaX, and infB, were amplified using primers shown in Suppl. Table 2. Reactions in a volume of 25 µl were performed comprising the following: 1 × DreamTaq (incl. 20 mM MgCl2), 200 µM mixed dNTP solution, 0.04 U of DreamTaq DNA Polymerase (Thermo Fisher Scientific); 0.2 µM of each primer, 1.0 µl of template DNA (0.5 to 500 ng), and nuclease-free water were added to make up the final volume of 25 µl. The cycle conditions were as follows: initial denaturation at 94 °C for 3 min, 35 cycles of denaturation at 94 °C for 1 min, annealing temperature at 59 °C (dnaX), 55 °C (atpD and infB) for 1 min, elongation at 72 °C for 2 min (dnaX) and 1 min (atpD and infB), and a further 5 min of elongation at 72 °C. The purification and subsequent sequencing of PCR products were performed as described previously.
Sequences of the three protein coding genes of the putative Pectobacterium isolates from this study were edited using BioEdit Sequence Alignment v 7.2.5 (Hall 1999). A multiple sequence alignment including partial sequences of the putative Pectobacterium isolates from this study together with reference type strains, where possible, of Pectobacterium and Dickeya spp., in the Pectobacteriaceae was performed using the default settings of the online version of MAFFT (v7) (Katoh et al 2019). The aligned sequences were exported to BioEdit Sequence Alignment Editor v 7.2.5 (Hall 1999) to trim overhang sequences to a nucleotide (nt) length of 641 for atpD, 443 for dnaX, and 573 for infB. Maximum likelihood trees were constructed for the individual genes and concatenated dataset using MEGA X v 10.2.6 (Kumar et al. 2018) using the GTR model including gamma shape (+ G) and invariable sites (+ I). A bootstrap analysis of 1000 replicates was also performed to provide support for the different branching points. The Cronobacter sakazakii CS-931 strain was included as an outgroup.
Symptom and Pathogenicity Comparative Trials
Isolate Selection
All isolates identified up to 21 July 2020 (Table 1), together with reference strains obtained from the CMEG culture collection (Suppl. Table 1), were evaluated for their maceration ability on tuber slices. At that stage, only Pb isolates were available, along with one strain each of Pc and Pa. Class 1 cv. Sifra tubers were peeled and disinfested by submerging in 5% sodium hypochlorite (NaOCl) for 20 min, followed by two washes of five minutes each, in sterile, distilled water (sdH2O), followed by a final wash in 70% ethanol for 1 min. Tubers were allowed to air-dry on sterile paper towels in a laminar flow. Once dry, the tubers were cut into 7-mm-thick slices and placed on sterile filter paper discs with 0.5 ml sdH2O, in 90 mm Petri dishes. A sterile cork borer was used to make a single 5 mm diameter hole, approximately 3.5 mm deep, in the centre of each tuber slice. For each isolate, the 48-h-old colonies on NA were re-suspended in sdH2O, and the optical density (OD) adjusted to 0.1 (equivalent to 8 × 107 cfu/ml) (Skelsey et al. 2018). A volume of 100 µl of the inoculum suspension was pipetted into the hole made in the tuber slice. A single tuber slice was inoculated per isolate. The Petri dish lids were replaced and sealed with Parafilm. They were incubated at 28 °C for 72 h, allowing time for lesions to develop. Thereafter, the lesion diameter was measured in two perpendicular directions and the average per tuber slice calculated. The Pb isolate SRP 1 (Table 1) had the largest average lesion diameter and was selected, along with the Pc and Pa strains from CMEG, for subsequent pot trials and tuber slice assays.
Greenhouse Pot Trials
The pot trials consisted of eight different treatments, comprising inoculation with the three species (Pb, Pc, and Pa) separately, and in all possible combinations. The treatments were thus (1) Pb, (2) Pc, (3) Pa, (4) Pb + Pc, (5) Pb + Pa, (6) Pc + Pa, (7) Pb + Pc + Pa, and (8) untreated control. The experimental layout was a randomised complete block design (RCBD), consisting of five blocks (replicates), each with two sub-samples per treatment. A sub-sample consisted of a single potted plant. The blocks were arranged perpendicular to a potential gradient created by an air conditioner at the northern end of the greenhouse compartment. The trial was repeated once. Trial 1 ran from 29 July 2020 to 25 August 2020 and trial 2 from 10 September 2020 to 30 September 2020.
The Pc and Pa reference strains and selected Pb isolate were cultured on NA for 48 h, inoculated into nutrient broth (NB) and incubated in a shaker, at 28 °C (Pb and Pc) or 25 °C (Pa), and 250 rpm, overnight. The following day, bacteria were sub-cultured into new NB and incubated under the same conditions as above, until their concentrations reached an OD of 1 (De Boer et al. 2012). The NB cultures were transferred to 50-ml falcon tubes and centrifuged at 5000 rpm and 4 °C for 15 min, to pellet the bacteria. The supernatant was discarded, and the pellet re-suspended in an equal volume of ¼ strength Ringer’s solution and kept at 4 °C to prevent further bacterial growth. On the same day, surface disinfested (submerged in 1% NaOCl for 10 s followed by 2 consecutive washes in sdH2O), sprouted disease-free minitubers of cv. Mondial, were vacuum infiltrated with the prepared inoculum, according to a protocol adapted from van der Wolf et al. (2017). The tubers were immersed in 2 l of a bacterial suspension in sdH2O, and a vacuum of 85 kPa was applied for 25 min. In trial 1, the initial inoculum concentration (OD of 1) was diluted 100 times to a final OD of 0.01, equivalent to approximately 8 × 106 cfu/ml. In trial 2, it was diluted 1000 times to a final OD of 0.001, or 8 × 105 cfu/ml, due to poor emergence in the first trial (van der Wolf et al. 2017). In the co-inoculation treatments, the same bacterial concentrations per species were used as in the single species treatments. For the negative control, ¼ strength Ringer’s solution was used instead of inoculum. After vacuum infiltration, the tubers were allowed to air-dry overnight. The final bacterial concentration in the suspension was confirmed by direct plate count on CVP agar. One tuber was planted per 1 l pot in sandy loam soil at a depth of 10 cm. The temperature in the greenhouse was kept at 28 °C ± 2 °C, and the plants were exposed to the normal day-night light cycle. The pots received 400 ml water every second day, which was supplemented with Dr Fisher’s Multifeed Classic 19:8:16 fertiliser at a rate of 2.5 g/l once a week.
Symptoms were recorded from their first appearance, around two weeks post-plant, until the plants started to die. This amounted to a period of 5 days in trial 1 and 8 days in trial 2. Recordings were made every second day. In trial 1, only three recordings were taken before the death of the first plants. In trial 2, five recordings were taken. Symptoms assessed were stem rot (blackleg), wilting, chlorosis, stunting, damping-off, non-emergence, and atypical blackleg symptoms. The first three symptoms, i.e., stem rot, wilting, and chlorosis, were rated on a scale of 0–4, representing different proportions of severity of the most diseased haulm per plant (Hélias et al. 2000). Specifically, 0 represented no symptoms; 1, less than 25% of the surface area affected; 2, between 25 and 50% affected; 3, between 50 and 75% affected; and 4, between 75 and 100% affected (Azadmanesh et al. 2016). Stunting was rated on a scale of 0–3, representing different proportions of stunting relative to the untreated control plants in the block. Specifically, 0 represented no stunting; 1, less than 50% stunted; 2, between 50 and 99% stunted; and 3, more than 99% stunted. Damping-off, non-emergence, and atypical blackleg symptoms were scored as present or absent (Hélias et al. 2000). These ratings were used to calculate incidence and mean severity.
Tuber Slice Assay
A tuber slice assay was performed to compare the same treatments as in the pot trials for their maceration ability. An RCBD experimental layout was used. A replicate consisted of a single tuber slice per block, of which there were five. The same inoculum used in the vacuum infiltration was used in the tuber slice assay. The original inoculum was diluted 10 times, to an OD of 0.1 (equivalent to 8 × 107 cfu/ml), and 50 µl aliquots were inoculated into holes in the tuber slices, as previously described (Skelsey et al. 2018). The tuber slices were incubated at 28 °C for 72 h in an incubator and thereafter assessed for lesion mass. This was accomplished by weighing the slices, before and after being rinsed under running tap water to remove the decayed tissue. The lesion mass was equivalent to the difference between the mass before and mass after rinsing (Duarte et al. 2004). The mean lesion mass per treatment was then calculated. The assay was repeated once.
Data Analysis
Incidence was calculated as the percentage (%) of sub-samples within a treatment affected by a particular symptom. Before mean severity was calculated, the severity scores were converted to their mid-point % values. Therefore, for stem rot, wilting, and chlorosis, 0 = 0%, 1 = 12.5%, 2 = 37.5%, 3 = 62.5%, and 4 = 87.5%, and for stunting, 0 = 0%, 1 = 25%, 2 = 74.5%, and 3 = 99.5%. The formula used to calculate stem rot, wilting, and chlorosis mean severities was Mean severity (%) = {0(n0) + 12.5(n1) + 37.5(n2) + 62.5(n3) + 87.5(n4)/(Ntotal)}, and for stunting, it was Mean severity (%) = {0(n0) + 25(n1) + 74.5(n2) + 99.5(n3)/(Ntotal)}, where Ntotal was the total number of sub-samples rated and nx was the total number of sub-samples of rating score x (with x representing the mid-point % values). The incidences and severities of the different symptoms were analysed by standard analysis of variance (ANOVA) (α = 0.05). The means for the different treatments were compared by Tukey’s HSD mean comparison test (α = 0.05). Area under disease progress curve (AUDPC) values were calculated from disease progress curves of stem rot severity in trial 2. In trial 1, not enough recordings were taken for this. These values were also analysed by standard ANOVA (α = 0.05) and means separated by Tukey’s HSD mean comparison test (α = 0.05). In the tuber slice assays, ANOVAs (α = 0.05) were performed on the lesion mass data and mean lesion masses compared with Tukey’s HSD test (α = 0.05). All ANOVAs and Tukey’s HSD tests were performed with ARM 2020 Revision 2020.3 software (Gylling Data Management Inc.).
Fulfilling Koch’s Postulates for First Report
Greenhouse Pathogenicity Trials
A greenhouse pathogenicity trial and its repeat were conducted between 28 October 2021 and 28 December 2021 in the same compartment to fulfil Koch’s postulates for two isolates of Pv (SRP2 and SRP3), identified by MLSA, not yet reported on potatoes in SA. An isolate of Pb (SRP1) was also included as a positive control for blackleg symptoms. The experimental layout was an RCBD, consisting of three blocks and one plant per treatment per block. There were eight different treatments, consisting of three separate isolates and two inoculation methods, namely vacuum infiltration and stem injection. An untreated control was included for each inoculation method.
Bacterial colonies grown on NA for 2 days at 28 °C were re-suspended in ¼ strength Ringer’s solution and the OD adjusted to 1. On the same day, surface-disinfested, sprouted, disease-free minitubers of cv. Mondial were vacuum infiltrated with the prepared inoculum. The tubers were immersed in 2 l of a bacterial suspension, at an OD of 0.001 (8 × 105 cfu/ml) (van der Wolf et al. 2017), in sdH2O and a vacuum of 85 kPa applied for 25 min. For the negative control, ¼ strength Ringer’s solution was used instead of inoculum. After vacuum infiltration, the tubers were allowed to air-dry overnight. The final bacterial concentration in the suspension was confirmed by direct plate count on CVP agar. For the following day, the inoculated tubers were planted at a depth of 10 cm in potting media (Culterra) in 1 l pots. Two tubers were planted per pot to ensure at least one emerged. The temperature in the greenhouse was kept at 28 °C ± 2 °C, and the plants were exposed to the normal day-night light cycle. The pots received 400 ml water every second day. Five weeks after emergence, inoculation of the remaining treatments was performed by injecting a single stem per pot with 10 µl of 8 × 108 cfu/ml (OD = 1) inoculum and subsequently covering the wound with Parafilm (Tsror et al. 2020).
Symptoms of blackleg were scored as present or absent from their first appearance until the plants started to die. Isolations and DNA extractions from symptomatic haulms were performed as previously described. Repetitive element palindromic PCR (rep-PCR) of the repetitive BOX element was performed to confirm the identity of the reisolates with the original isolates. The BOX A1R primer (5′ CTA CGG CAA GGC GAC GCT GAC G 3′), designed from the boxA subunit of BOX was used (Versalovic et al. 1994). The PCR was carried out in a 25 µl reaction volume comprising 12.5 µl of OneTaq® Hot Start 2X Master Mix with Standard Buffer (New England Biolabs), 2 µM of the BOX A1R primer, and 20 ng template DNA, added to nfH2O (Qiagen). The thermocycling regime, adapted from Rademaker et al. (2004), consisted of initial denaturation at 95 °C for 7 min, followed by 35 cycles of denaturation (94 °C for 60 s), annealing (53 °C for 60 s), extension (72 °C for 90 s), and a final extension at 72 °C for 10 min. The amplified fragments were resolved on 2% (w/v) Agarose gel (LONZA) in 1 × TAE buffer (40 mM Tris–acetate, 1 mM EDTA), containing 0.05 µl ml−1 Roti®GelStain and run at 60 V and 30 mA for 16 h at 4 °C. The bands were visualised with a UV transilluminator (Bio-Rad Laboratories), and the profiles of the re-isolates and original isolates were compared to fulfil Koch’s postulates.
Tuber Maceration Trials
Tuber maceration trials were performed to compare the maceration ability of the two isolates of Pv (SRP 2 and SRP 3), one isolate of Pc (SRP 4), and one isolate of Pb (SRP 6) to each other and to the maceration ability of Pb (SRP 1), the positive control. A negative control, which consisted of sdH2O, was also included. Therefore, there were six treatments in total. A RCBD experimental layout was used, as in the pot trials. Three replicates, each consisting of a single tuber slice, were used for each of the six treatments.
The protocol described in the isolate selection section was used with a few modifications. Class 1 Sifra tubers were disinfested by submerging in 1% NaOCl for 20 min, followed by two washes of 5 min each in sdH2O. Tubers were allowed to air-dry on sterile paper towels under a laminar flow. Once dry, the tubers were cut into 7-mm-thick slices and placed on filter paper discs with 0.2 ml sdH2O in 90 mm Petri dishes. For each isolate, 50 µl was inoculated onto a separate tuber slice, into 5 mm (diameter) holes made with a sterile cork borer. The same inoculum used in the greenhouse pathogenicity trials was used in the tuber maceration trials. The original inoculum was diluted 10 times to an OD of 0.1 (equivalent to 8 × 107 cfu/ml). Once tuber slices were inoculated, the Petri dish lids were replaced and sealed with Parafilm. Tuber slices were incubated at 28 °C for 72 h, allowing time for lesions to develop. The assay was repeated once.
After incubation, the slices were rinsed under running tap water to remove the decayed tissue. The extent to which the surface area of the tuber slice was removed due to soft rot was assessed on a rating scale of 0 to 3. A rating of 0 was equivalent to no soft rot, 1 was equivalent to soft rot of less than 5% of the surface area, 2 was equivalent to soft rot of 5 to 50% of the surface area, and 3 was equivalent to soft rot of more than 50% of the surface area.
Results
SRP Survey
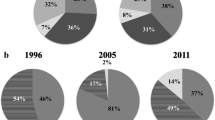
The Pb-specific and multiplex PCRs identified 50 out of 70 isolates as Pb and 14 as Pc or Pp, while six isolates yielded no amplicons (Table 1 and Fig. 1). The gapA PCR-sequencing assay identified 33 out of 70 isolates as Pb, 10 as Pc, three as Pp, two as Dickeya chrysanthemi, and two as Pv. Five isolates belonging to the Enterobacteriaceae family were also identified. These were isolates MCC1 and B1.5 (Enterobacter ludwigii), isolate MCC2.1 (Enterobacter cloacae), isolate MCC5.2 (Kosakonia sacchari), and isolate CWK2B (Enterobacter sp.). Another 15 isolates were not identified by gapA sequencing, either due to not amplifying in the PCR, having sequences with low Phred quality scores, or because they were not sequenced (Table 1 and Fig. 2). According to BLASTn, isolates SRP 5 and SRP 6 shared 98.91% and 99.21% identity with a Dickeya chrysanthemi isolate (Accession no. CP001655.1), while isolates SRP 2 and SRP 3 both shared 99.86% identity with a Pv isolate (Accession no. CP024842.1). The identification of five Pectobacterium isolates was further corroborated by the MLSA of the three housekeeping genes, i.e., atpD, dnaX, and infB, (Fig. 3) with a few exceptions. Based on the concatenated phylogeny, SRP 4 strain shared the same clade as the type strain of Pectobacterium carotovorum CFBP2046T with 99% branch support. Similarly, the SRP 2 and SRP 3 strains shared the same clade with the type strain of Pv CFBP6051T with 99% branch support. In contrast, both the SRP 1 and SRP 6 strains shared a clade with the type strain of Pb LMG21371T with 100% branch support. These findings were supported by the individual gene trees (Suppl. Figures 1, 2, and 3).

Frequency of isolates per species as identified by Pb-specific and multiplex PCR, the latter unable to distinguish between Pc and Pp. ‘Unidentified’ refers to isolates that did not yield amplicons in these PCR assays

Frequency of isolates per species as identified by BLASTn of the gapA sequence. ‘Unidentified’ refers to isolates that either did not yield amplicons in the gapA PCR, had sequences with low Phred quality scores, or were not sequenced. ‘Other’ refers to five isolates belonging to the Enterobacteriaceae family, namely, Enterobacter ludwigii, Enterobacter cloacae, Kosakonia sacchari, and Enterobacter sp.

A maximum-likelihood phylogenetic tree of concatenated atpD, dnaX, and infB housekeeping genes (1 657 bp). GTR + I + G was used as the evolutionary model on MEGA X to construct the tree, and bootstrap values are indicated as percentages. Cronobacter sakazakii CS-931 was included as an outgroup. The scale bar represents substitutions per site
Symptom and Pathogenicity Comparative Trials
Greenhouse Pot Trials
Treatment was significant in the ANOVAs for damping-off incidence, stem rot incidence and mean severity, stunting incidence and mean severity, and wilting incidence and mean severity for both trials, and for non-emergence incidence in trial 1,and AUDPC in trial 2 (p < 0.05). No chlorosis or atypical blackleg occurred, and stem rot colour was always black. Disease progress curves were not constructed for trial 1 due to too few recordings being taken (less than 4) before the plants started to die.
In both trials, stem rot only occurred in the treatments involving Pb and its complexes (Pb + Pc; Pb + Pa; Pb + Pc + Pa) (Table 2). In trial 1, the incidences and mean severities of the mentioned treatments differed significantly from the treatments of only Pc and/or Pa and the untreated control (p < 0.05). They did not, however, differ significantly from one another. In trial 2, only Pb, Pb + Pc, and Pb + Pc + Pa had incidences and mean severities significantly higher than the treatments involving only Pc and/or Pa and the negative control, which did not display this symptom at all (p < 0.05). The incidence but not severity of Pb + Pa was significantly higher than the treatments that did not develop stem rot. The AUDPC values, calculated from the disease progress curves of stem rot in trial 2 (Suppl. Figure 4), were significantly different from the untreated control only for the treatments involving Pb and its complexes (p < 0.05) (Table 2).
In trial 1, Pb had an incidence of non-emergence that was significantly higher than for any of the other treatments, including the untreated control. The only other treatment that had an incidence significantly higher than the untreated control was Pb + Pa (p < 0.05) (Table 3). Trial 2 had very few instances of non-emergence, and differences between treatments were not significant.
In both trials, damping-off occurred only in the treatments of Pb and its complexes (Pb + Pc; Pb + Pa; Pb + Pc + Pa) (Table 3). In trial 1, inoculation with Pb alone had the highest incidence, as the only plant that emerged, succumbed to damping-off. The treatments of Pb + Pa, Pb + Pc, and Pb + Pc + Pa all had lower incidences than Pb, albeit not significantly different. All the mentioned treatments had significantly higher incidences of damping-off than the treatments involving only Pc and/or Pa and the negative control, which did not display this symptom at all (p < 0.05). In trial 2, the same treatments as in trial 1 had significantly higher incidences of damping-off than the untreated control, although only Pb + Pa and Pb + Pc + Pa were significantly higher than the treatments involving only Pc and/or Pa which did not display this symptom at all (p < 0.05).
In both trials, only treatments involving Pb and its complexes (Pb + Pc; Pb + Pa; Pb + Pc + Pa) had significantly higher incidences and mean severities of stunting than the untreated controls (p < 0.05) (Table 3). Although stunting did occur in the treatments of Pc, Pa, and its complex, Pc + Pa, neither the incidences nor severities were significantly different from the untreated control which had no such symptoms.
In both trials, only treatments of Pb and its complexes (Pb + Pc; Pb + Pa; Pb + Pc + Pa) had wilting incidences and mean severities significantly higher than the untreated controls (p < 0.05) (Table 3). The only exception to this was the mean severity of Pb + Pa in trial 2, which was not significantly higher than the untreated control.
Tuber Slice Assay
Treatment was significant in the ANOVA for soft rot lesion mass in both trials (p < 0.05) (Table 4). In trial 1, the treatments of Pb, Pb + Pc, Pb + Pa, Pb + Pc + Pa, and Pc + Pa had mean lesion masses that were significantly higher than the untreated control, although Pc + Pa was not significantly higher than the treatments of Pc and Pa (p < 0.05). In trial 2, the treatments Pb, Pc, Pb + Pa, Pc + Pa, and Pb + Pc + Pa had mean lesion masses that were significantly higher than the untreated control (p < 0.05).
Fulfilling Koch’s Postulates for First Report
Greenhouse Pathogenicity Trials
Stem inoculation treatments resulted in a slightly higher overall incidence of blackleg than vacuum infiltration (Fig. 4). The Pb positive control (SRP 1) had the highest incidence of blackleg of all the isolates tested, with 3/3 plants affected in both trials for stem inoculation and 2/3 in both trials for vacuum infiltration. For the other treatments, the incidences of blackleg were much lower or entirely absent, with 1/3 plants affected in the stem inoculation treatment of SRP 2 (Pv) in trial 2. Comparison of the rep-PCR genomic fingerprints of the reisolates to those of the original isolates confirmed their identities.

Incidence of blackleg in a greenhouse pathogenicity trial and its repeat, conducted to fulfil Koch’s postulates. SRP 1, Pb positive control; SRP 2, P. versatile isolate 1; SRP 3, P. versatile isolate 2; (V), vacuum infiltration; (S), stem inoculation
Tuber Maceration Trials
In both trials, soft rot occurred in all the treatments involving SRP 1 (Pb), SRP 2 (Pv), SRP 3 (Pv), SRP 4 (Pc), and SRP 6 (Pb). Also, the extent of decayed tissue removed from the tuber slices after rinsing was more than 50% of the surface area in all the treatments (Table 5). In trial 1, however, two of the three replicates for the negative control displayed a small amount of decayed tissue. In trial 2, none of the negative control replicates indicated any signs of decay.
Discussion
Findings from the survey suggest that Pb remains the dominant species in the country, as was previously reported (van der Merwe et al. 2010; Ngadze et al. 2012; Moleleki et al. 2013). Until recently, Pv formed part of Pc (Portier et al. 2019), which would explain why it was not detected previously. Pectobacterium carotovorum is known to be present in SA (van der Merwe et al. 2010; Ngadze et al. 2012; Moleleki et al. 2013). The lack of detection of D. solani in any of the samples, suggests that it remains absent from the country. The identification of five isolates belonging to the Enterobacteriaceae is most likely due to contaminated cultures.
Although Pv caused blackleg symptoms in the pathogenicity trials, it was at a much lower incidence than the Pb positive control. The recent description of Pv, formerly Pc (Portier et al. 2019), which is more often associated with aerial stem rot than blackleg (Pérombelon 2002; De Boer et al. 2012; Charkowski 2018), might explain its low blackleg incidence in the trials as well as the aerial stem rot symptoms from which the initial isolations were made. The higher incidence of blackleg found in association with stem inoculation than with vacuum infiltration might be due to the direct, unrestricted access to the xylem created by stem injection. Such a breach is required for aerial stem rot development (De Boer et al. 2012). Although stem inoculation is the standard method used in pathogenicity trials, vacuum infiltration was included as it is the method that most closely resembles natural infection (Taylor et al. 2021).
The confirmation of the identity of the reisolates by rep-PCR fulfilled Koch’s postulates. This is thus the first report of Pv as a causal agent of potato blackleg and soft rot in South Africa, as has been reported abroad (Ma et al. 2021, 2022). Results in this study also indicate that Pv isolates found in South Africa can cause soft rot to the same extent as Pb.
Findings from the symptom and pathogenicity comparative pot trials indicate that, out of the three species that were tested, Pb appeared to be the main driver behind the variation in incidence and severity of the symptoms that were observed. As the treatments of Pc, Pa, and Pc + Pa produced no significant symptoms in the pot trials, and no atypical symptoms were observed among the other treatments, no conclusions could be drawn regarding differences in symptom appearance between the three species. Most strains of Pc are not known to produce blackleg but rather, what is known as aerial stem rot, which does not originate from infected tubers, but instead from aerial stem parts (Pérombelon 2002; De Boer et al. 2012; van der Wolf et al. 2017). This could explain the absence of symptoms in the treatments with this species, as no aerial stem parts were inoculated. A possible explanation for the absence of symptoms associated with the Pa treatments was the high temperature (28 °C) in the greenhouse compartment. It is known that Pa prefers cool conditions (van der Merwe et al. 2010). It was found to have an optimal in vitro temperature of 26–27 °C and a temperature growth range of 18 to 31 °C (du Raan et al. 2016). The only significant differences between the treatments were in the incidence and severity of the symptoms. In most cases, the treatments involving Pb or its complexes had significantly higher incidences and mean severities than the untreated controls. This was nowhere more apparent than in the AUDPC values observed, which is a clear indicator of disease intensity over time. This corresponds to findings by van der Wolf et al. (2017), that Pb is more virulent than the other SRPs, including Pc and Pa, under most environmental conditions. The high incidence of non-emergence in trial 1, in contrast to trial 2, was most likely due to the difference in inoculum concentrations used. This correlates with the findings of Bain et al. (1990) that showed that higher inoculum concentrations in tubers before planting resulted in higher incidences of non-emergence. The lack of significant differences in incidence and mean severity between treatments of individual species and their complexes indicated a lack of possible interactions between species. A potential antagonistic interaction would have resulted in a complex with significantly lower incidence and/or severity than the individual species it was composed of, as has been observed with co-infections of D. solani and Pc (van der Wolf et al. 2017).
Findings from the tuber slice assays of the symptom and pathogenicity comparison trials corresponded to those from the pot trials. The significantly higher mean lesion masses in most of the treatments with Pb, and its complexes further reinforce the conclusion that Pb is the most pathogenic of the species tested. It is noteworthy that the treatments of Pc + Pa in both trials and Pc in trial 2 also had significantly higher mean lesion masses than the untreated controls. This indicates that, although these two species and their combination were unable to cause blackleg, they were still capable of causing tuber soft rot under the in vitro conditions. This is possibly due to the tuber periderm, a physical barrier to infection, being bypassed, with inoculation occurring directly into the pith in the tuber slice assays (Ngadze and van der Waals 2012). Also, many Pc strains, although unable to cause blackleg, are still known to cause soft rot (Pérombelon 2002; De Boer et al. 2012; van der Wolf et al. 2017).
Conclusion
A 2-year survey of the pectinolytic Dickeya and Pectobacterium species affecting the South African potato industry revealed that P. brasiliense remains most prevalent (47% of isolates). Other species that were identified included Pc (14%), Pp (4%), and Pv (3%). The latter is a first report in South Africa, and pathogenicity of the P. versatile isolates was confirmed through Koch’s postulates. Disease expression of single species and all possible combinations of Pb, Pc, and Pa was investigated in pot trials. In both trials, stem rot, non-emergence, damping-off, stunting, and wilting symptoms only occurred in the treatments involving Pb and its complexes (Pb + Pc; Pb + Pa; Pb + Pc + Pa), confirming the aggressive nature and competitive ability of Pb under favourable environmental conditions. Lack of detection of D. solani in any of the samples tested suggests that this destructive pathogen is not yet present in the South African potato industry. Studies such as this one, which provide more information about the distribution, dispersal, survival, and other epidemiological characters of the SRPs present in a country, are needed to ensure the sustainability of potato production on regional, national, and international levels.
References
Agrios GN (2005) Control of plant diseases. Plant Pathology, 5th edn. Elsevier Academic Press, Boston, pp 297–298
Altschul SF, Gish W, Miller W et al (1990) Basic local alignment search tool. J Mol Biol 215:403–410. https://doi.org/10.1016/S0022-2836(05)80360-2
Azadmanesh S, Mozafari J, Hasanzadeh N, Moslemkhani C (2016) In vitro evaluation of potato genotypes for resistance against bacterial soft rot (Pectobacterium carotovorum) - a new tool for studying disease resistance. J Plant Prot Res 57:1–8. https://doi.org/10.1515/jppr-2017-0001
Bain RA, Pérombelon MCM, Tsror L, Nachmias A (1990) Blackleg development and tuber yield in relation to numbers of Erwinia carotovora subsp. atroseptica on seed potatoes. Plant Pathol 39:125–133
Charkowski AO (2018) The changing face of bacterial soft-rot diseases. Annu Rev Phytopathol 12:269–288. https://doi.org/10.1146/annurev-phyto-080417
Cigna J, Dewaegeneire P, Beury A et al (2017) A gapA PCR-sequencing assay for identifying the Dickeya and Pectobacterium potato pathogens. Plant Dis 101:1278–1282. https://doi.org/10.1094/PDIS-12-16-1810-RE
Czajkowski R, de Boer WJ, van der Wolf JM (2013) Chemical disinfectants can reduce potato blackleg caused by ‘Dickeya solani’. Eur J Plant Pathol 136:419–432. https://doi.org/10.1007/s10658-013-0177-8
Czajkowski R, Pérombelon MCM, Van Veen JA, van der Wolf JM (2011) Control of blackleg and tuber soft rot of potato caused by Pectobacterium and Dickeya species: a review. Plant Pathol 60:999–1013. https://doi.org/10.1111/j.1365-3059.2011.02470.x
De Boer SH, Li X, Ward LJ (2012) Pectobacterium spp. associated with bacterial stem rot syndrome of potato in Canada. Phytopathol 102:937–947. https://doi.org/10.1094/PHYTO-04-12-0083-R
Department of Agriculture (2013) South African seed potato scheme. Government Gazette 449:1–9. http://www.potatocertification.co.za/SiteResources/documents/Scheme%202013.pdf. Accessed 16 May 2022
Del Pilar M-V, Groves RL, Charkowski AO (2011) Soft rot disease severity is affected by potato physiology and pectobacterium taxa. Plant Dis 95:232–241. https://doi.org/10.1094/PDIS-07-10-0526
Duarte V, De Boer SH, Ward LJ, De Oliveira AMR (2004) Characterization of atypical Erwinia carotovora strains causing blackleg of potato in Brazil. J Appl Microbiol 96:535–545. https://doi.org/10.1111/j.1365-2672.2004.02173.x
du Raan S, Coutinho TA, van der Waals JE (2016) Cardinal temperature differences, determined in vitro, between closely related species and subspecies of pectinolytic bacteria responsible for blackleg and soft rot on potatoes. Eur J Plant Pathol 144:361–369. https://doi.org/10.1007/s10658-015-0773-x
Hall TA (1999) BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl Acids Symp Ser 41:95–98
Hélias V, Andrivon D, Jouan B (2000) Development of symptoms caused by Erwinia carotovora ssp. atroseptica under field conditions and their effects on the yield of individual potato plants. Plant Pathol 49:23–32
Hélias V, Hamon P, Huchet E et al (2012) Two new effective semiselective crystal violet pectate media for isolation of Pectobacterium and Dickeya. Plant Pathol 61:339–345. https://doi.org/10.1111/j.1365-3059.2011.02508.x
Kabir MN, Taheri A, Dumenyo CK (2020) Development of PCR-based detection system for soft rot Pectobacteriaceae pathogens using molecular signatures. Microorg 8:358. https://doi.org/10.3390/microorganisms8030358
Katoh K, Rozewicki J, Yamada KD (2019) MAFFT online service: multiple sequence alignment, interactive sequence choice and visualization. Brief Bioinform 20(4):1160–1166. https://doi.org/10.1093/bib/bbx108
Kim MH, Cho MS, Kim BK et al (2012) Quantitative real-time polymerase chain reaction assay for detection of Pectobacterium wasabiae using YD repeat protein gene-based primers. Plant Dis 96:253–257. https://doi.org/10.1094/PDIS-06-11-0511
Kubheka GC, Coutinho TA, Moleleki N, Moleleki LN (2013) Colonization patterns of an mcherry-tagged Pectobacterium carotovorum subsp. brasiliense strain in potato plants. Phytopathol 103:1268–1279. https://doi.org/10.1094/PHYTO-02-13-0049-R
Kumar S, Stecher G, Li M, Knyaz C, Tamura K (2018) MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Biol Evol 35:1547–1549. https://doi.org/10.1093/molbev/msy096
Kumvinit A, Akarapisan A (2019) Characterization of blackleg and soft rot from potato in northern Thailand. J Phytopathol 167:655–666. https://doi.org/10.1111/jph.12859
Laurila J, Hannukkala A, Nykyri J et al (2010) Symptoms and yield reduction caused by Dickeya spp. strains isolated from potato and river water in Finland. Eur J Plant Pathol 126:249–262. https://doi.org/10.1007/s10658-009-9537-9
Ma X, Brazil J, Rivedal H et al (2022) First report of Pectobacterium versatile causing potato soft rot in Oregon and Washington. Plant Dis 106:1292. https://doi.org/10.1094/PDIS-08-21-1635-PDN/SUPPL_FILE/PDIS-08-21-1635-PDN_SF2.PDF
Ma X, Stodghill P, Gao M et al (2021) Identification of Pectobacterium versatile causing blackleg of potato in New York State. Plant Dis 105:2585–2594. https://doi.org/10.1094/PDIS-09-20-2089-RE
Moleleki LN, Onkendi EM, Mongae A, Kubheka GC (2013) Characterisation of Pectobacterium wasabiae causing blackleg and soft rot diseases in South Africa. Eur J Plant Pathol 135:279–288. https://doi.org/10.1007/s10658-012-0084-4
Ngadze E, Brady CL, Coutinho TA, van der Waals JE (2012) Pectinolytic bacteria associated with potato soft rot and blackleg in South Africa and Zimbabwe. Eur J Plant Pathol 134:533–549
Ngadze E, van der Waals JE (2012) Role of polyphenol oxidase, peroxidase, phenylalanine ammonia lyase, chlorogenic acid, and total soluble phenols in resistance of potatoes to soft rot. Plant Dis 96:186. https://doi.org/10.1094/PDIS-02-11-0149
Pérombelon MCM (2002) Potato diseases caused by soft rot erwinias: an overview of pathogenesis. Plant Pathol 51:1–12. https://doi.org/10.1046/j.0032-0862.2001.Shorttitle.doc.x
Portier P, Pédron J, Taghouti G et al (2019) Elevation of Pectobacterium carotovorum subsp. odoriferum to species level as Pectobacterium odoriferum sp. nov., proposal of Pectobacterium brasiliense sp. nov. and Pectobacterium actinidiae sp. nov., emended description of Pectobacterium carotovorum and description of Pectobacterium versatile sp. nov., isolated from streams and symptoms on diverse plants. Int J Syst Evol Microbiol 69:3207–3216. https://doi.org/10.1099/ijsem.0.003611
Potrykus M, Sledz W, Golanowska M et al (2014) Simultaneous detection of major blackleg and soft rot bacterial pathogens in potato by multiplex polymerase chain reaction. Ann Appl Biol 165:474–487. https://doi.org/10.1111/aab.12156
Rademaker JLW, Louws FJ, Versalovic J, De BFJ (2004) Characterization of the diversity of ecologically important microbes by rep-PCR genomic fingerprinting. In: Kowalchuk GA, de Bruijn FJ, Head IM, Akkermans AD, van Elsas JD (eds) Molecular Microbial Ecology Manual, 2nd edn. Kluwer Academic Publishers, Dordrecht, pp 1–33
Skelsey P, Humphris SN, Campbell EJ, Toth IK (2018) Threat of establishment of non-indigenous potato blackleg and tuber soft rot pathogens in Great Britain under climate change. PLoS ONE 13:1–18. https://doi.org/10.1371/journal.pone.0205711
Taylor D, Charkowski AO, Zeng Y (2021) Laboratory assays used to rank potato cultivar tolerance to blackleg showed that tuber vacuum infiltration results correlate with field observations. Plant Dis 105:585–591
Toth IK, van der Wolf JM, Saddler G, Lojkowska E, Hélias V, Pirhonen M et al (2011) Dickeya species: an emerging problem for potato production in Europe. Plant Pathol 60:385–399. https://doi.org/10.1111/j.1365-3059.2011.02427.x
Tsror L, Ben-Daniel B, Chalupowicz L et al (2013) Characterization of Dickeya strains isolated from potato grown under hot-climate conditions. Plant Pathol 62:1097–1105. https://doi.org/10.1111/ppa.12030
Tsror L, Erlich O, Mordechai-Lebiush S et al (2020) First report of Pectobacterium parmentieri one of the causal agents of potato blackleg and tuber soft rot diseases in Israel. Plant Dis 104:2288. https://doi.org/10.1094/pdis-02-20-0226-pdn
van der Merwe JJ, Coutinho TA, Korsten L, van der Waals JE (2010) Pectobacterium carotovorum subsp. brasiliensis causing blackleg on potatoes in South Africa. Eur J Plant Pathol 126:175–185. https://doi.org/10.1007/s10658-009-9531-2
van der Waals JE, Krüger K, Franke AC et al (2013) Climate change and potato production in contrasting South African Agro-Ecosystems 3. Effects on relative development rates of selected pathogens and pests. Potato Res 56:67–84. https://doi.org/10.1007/s11540-013-9231-3
van der Wolf JM, de Haan EG, Kastelein P et al (2017) Virulence of Pectobacterium carotovorum subsp. brasiliense on potato compared with that of other Pectobacterium and Dickeya species under climatic conditions prevailing in the Netherlands. Plant Pathol 66:571–583. https://doi.org/10.1111/ppa.12600
Versalovic J, Schneider M, de Bruijn FJ, Lupski JR (1994) Genomic fingerprinting of bacteria using repetitive sequence based PCR (rep-PCR). Meth Cell Mol Biol 5:25–40
Funding
Funding for this work was provided by Potatoes South Africa.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Theron, E., Bophela, K.N., Bisschoff, J. et al. Survey of Soft Rot Pectobacteriaceae Infecting Potatoes in South Africa. Potato Res. 66, 723–749 (2023). https://doi.org/10.1007/s11540-022-09598-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11540-022-09598-1