
Abstract
Recently, a hybrid breeding system was developed for diploid potato. We compared performance of diploid hybrids with commercially available tetraploid cultivars. Therefore, seedling tubers were produced from true hybrid seeds in field conditions. In the subsequent year, diploid hybrids grown from seedling tubers showed a yield potential comparable with commercial tetraploid cultivars: the highest yielding diploid hybrids showed a yield comparable with the lower yielding tetraploid cultivars. Yields of hybrids and commercial tetraploid cultivars were broken down into different yield components and the interactions with growing conditions were quantified. The stability of yield and other relevant traits in different growing conditions was similar between hybrids and commercial cultivars. The contribution of the different yield components to total yield over different environments was compared between diploid hybrids and tetraploid cultivars. In diploid hybrids as well as tetraploid cultivars, more tubers per stem resulted in the highest yield gain, while an increase in tuber size resulted in a relatively smaller increase of total yield.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Potato (Solanum tuberosum L. and its tuber bearing crossable species) is the third food crop after rice and wheat (Zaheer and Akhtar 2016). A large proportion of the production is used for other purposes such as seed, industrial processing and feed (Haverkort and Struik 2015). Despite its importance, yield increase by genetic gain has been limited for the last century, especially in cultivars for consumption (Douches et al. 1996; Piepho et al. 2014). The most important factor limiting genetic gain is the lack of an efficient breeding system. An advanced system is hybrid breeding based on crosses between two homozygous inbred lines (Lindhout et al. 2011). By choosing contrasting parents, yield contributing alleles can be combined (Lindhout et al. 2011). The segregation of potato traits after several generations of inbreeding was described by Meijer et al. (2018). Until now, the absence of a self-compatibility system hampered the creation of homozygous inbred lines in potato. Since the discovery of the Sli-gene (Hosaka and Hanneman 1998), it has become possible to develop a diploid hybrid breeding system (Lindhout et al. 2018). The discovery not only creates opportunities for commercial breeding programmes, it also greatly facilitates genetic research by using advanced segregating populations such as introgression libraries and backcross populations (Jeuken and Lindhout 2004; Endelman and Jansky 2016; Prinzenberg et al. 2018). Diploid germplasm allows high throughput phenotyping and efficient QTL detection, accelerating the breeding process (Meijer et al. 2018; Prinzenberg et al. 2018; Su et al. 2019).
With the development of this hybrid breeding system, which is new for potato, using diploid hybrids instead of tetraploid clones, a system of testing new cultivars needs to be developed. Commonly in breeding systems, new cultivars are grown at several locations from tubers and their performance is benchmarked against a set of already existing commercial tetraploid cultivars (Tiemens-Hulscher et al. 2013). To determine relevant genetic gain, performance of diploid hybrids needs to be benchmarked against tetraploid cultivars commonly used by farmers. Assessing the genotype by environment interactions in farmers’ fields is an important aspect of acceptance of new (types of) cultivars.
Besides the use of new genetic material in diploid breeding, the starting material is also different from the traditional breeding germplasm. In tetraploid breeding, a cross is made only once and the true seeds derived from this cross are grown to seedling tubers (Tiemens-Hulscher et al. 2013). After that, genotypes are clonally propagated, which means that starting material of tetraploid cultivars always consists of seed tubers. Hybrid breeding in potato results in true (botanical) seeds. Plant development is highly affected by the type of starting material (true seed or seed tuber) and by its physiological, physical, genetic and phytosanitary quality. Size and physiological age of seed tubers have large effects on ware crop yield (Arsenault et al. 2001; Struik and Wiersema 2001). Therefore, seedling tubers need to be produced from diploid hybrids to evaluate the performance of hybrids in comparison to existing benchmark cultivars.
Although for most field crops, dry matter yield is the most important economic output, for potato fresh tuber yield, tuber size distribution, dry matter content and tuber shape are main output determinants. These traits are important criteria whether a cultivar is suitable for fry or chips production (Tiemens-Hulscher et al. 2013). Dry matter percentage should range between 22 and 25% for chips (crisps) and 21 and 23% for fries. Tubers that are processed into chips should be round, while tuber shape of frying cultivars should be oblong for optimal processing (Tiemens-Hulscher et al. 2013). Potato yield can be dissected into different components and be expressed as the product of tuber weight, tubers per stem and stems per unit area (Haverkort et al. 1990; Lynch et al. 1995; Struik and Wiersema 2001; Asghari-Zakaria et al. 2007). Potato production is highly affected by the environment (Haverkort 1990; Kooman et al. 1996). To investigate the specific development aspects of diploid hybrids in different environments, the contribution of these components that make up potato yield was investigated.
In this paper, we show the first results of such a hybrid breeding programme focusing on yield and yield components. As testing environments, we have chosen farmers’ fields in The Netherlands, Belgium and France, representative for the north-western European potato cultivation environment. This region is known for its high technological level of potato cultivation and its conducive climate, resulting in high yields.
The objectives of this study were to (1) evaluate the performance of diploid hybrids in comparison to existing tetraploid cultivars, (2) to compare the environmental effect on yield and yield components between diploid hybrids and tetraploid cultivars and (3) to evaluate the contribution of different yield components to total yield.
Materials and Methods
Genetic Material
Diploid heterozygous Solanum tuberosum accessions were crossed with the self-compatibility donor S. chacoense (Hosaka and Hanneman 1998). Inbred lines of diploid potato were developed in a dedicated diploid breeding programme as described by Lindhout et al. (2011, 2018). The inbred parents were crossed after 5 to 7 generations of self-pollinations to produce the test hybrids. These inbreds should theoretically be more than 95% homozygous, but this was not tested (Lindhout et al. 2011). In winter 2015–2016, crosses between parent lines were made manually to produce diploid hybrid seeds and 572 hybrids were grown in one location in a first year trial to select the most uniform hybrids. This resulted in a panel of 65 diploid hybrids that was used in the present study. These hybrids were selected based on a wide variation in yield and yield components to represent the genetic variation among these hybrids and to allow a detailed analysis.
Nineteen commercial tetraploid cultivars were used as controls for the different potato product types, representing the frying, chip and fresh consumption markets: Annabelle, Lady Cristil, Lady Claire, Pirol, Innovator, Lady Rosetta, Agata, Arsenal, Hermes, Brooke, Nicola, Bintje, Markies, Russet Burbank, Fontane, Spunta, Mozart, Agria and Milva. The main differences between these cultivars are dry matter content, shape and tuber size. The diploid hybrids were not yet differentiated for the different market segments. These control cultivars are commonly grown in the regions where the trials took place. Hence, these controls are a realistic representation of farmers’ practice.
Seed tubers of commercial cultivars were classified in class A in the NAK (Nederlandse Algemene Keuringsdienst voor zaaizaad en pootgoed van landbouwgewassen, The Dutch General Inspection Service) classification system (NAK 2018). Seed tubers were produced and stored in the conditions optimal for the specific cultivar for a realistic representation of the cultivar’s potential, so storage conditions were not identical for all cultivars. Diploid hybrids were all stored at 4 °C as optimal conditions for the specific hybrids were yet unknown. Seed tubers of equal size and sprout distribution were selected for the trial. To assess the suitability of the seedling tubers for a ware crop trial, sprouting was scored after 6 months of storage (data not shown) (Tiemens-Hulscher et al. 2013). Generally, seed tubers should be planted when dormancy has been broken (Struik and Wiersema 2001). Seedling tubers with a sprouting score between 3 and 7 were used in the trial; sprouting of seed tubers of tetraploid cultivars was comparable with the sprouting score of 5 of the diploid hybrids.
Seedling Tuber Production
Field Trial Description
Seedling tuber production from true diploid hybrid seeds was performed on a heavy marine clay in Wolphaartsdijk [NL], in 2016. The botanical seeds of the hybrids were extracted from the berries and sown into 104 plug trays (2.5 × 2.5 × 3.8 cm) filled with soil (Horticoop) in a greenhouse compartment in the first week of April 2016. Germination after 2 weeks was between 54 and 98%. Six weeks after sowing the seedlings were mechanically transplanted into the open field, into ridges of 75 cm width and a plant distance of 30 cm within the row. The crop was treated as a standard seed-tuber crop, according to farmers’ practices. Irrigation was applied at 1 and 3 days after planting to prevent the seedlings from drying out. Pests and diseases were chemically controlled. Both mechanical weeding and hand-weeding were applied twice. After 90 growing days in the field, the haulm was chemically killed. Three weeks after haulm killing, the seed tubers of all diploid hybrids were lifted by a harvesting machine. The seed tubers were stored in a climate-controlled storage room at 4 °C from October 1st onwards until the end of February.
Design and Measurement
Seedling tuber production was done in a non-replicated design with a plot size between 90 and 488 plants, depending on hybrid seed availability. Seedlings were transplanted semi-mechanically with an adjusted 4-row cabbage planter with a planting distance of 30 cm between plants. After harvest, seedling tubers were sorted by size and yield was determined for tubers > 28 mm. Phytosanitary control on quarantine diseases was routinely carried out by the national inspection service (NAK at Emmeloord, NL). To assess the quality of the seedling tubers, sprouting was scored in February, at the preparation of the yield trials according to a 1–9 scale, with 9 being completely dormant and 1 abundantly sprouting (Tiemens-Hulscher et al. 2013).
Yield Trials
Field Trial Description
In 2017, yield trials were conducted using the seedling tubers as starting material that were produced in 2016. Certified seed tubers (class A) of the tetraploid cultivars were obtained to serve as control cultivars in the trials. The field trials were conducted at five different sites (Warmeriville [FR], Berthem [BE], Hilvarenbeek [NL], Est [NL], Giethoorn [NL]). These sites differed in soil type, climate and crop management (Table 1). Two of the sites were typical sandy soils, one a loamy soil, one a light clay and one a heavy clay soil, covering the full range of physical soil types on which potatoes are grown in NW Europe. At all sites, the trial was laid out within a farmers’ field, hence crop management was according to farmers’ practice. Typically, the tubers were planted in ridges, spaced at 75 cm, except for the experiment in Warmeriville, where the distance between ridges was 90 cm (Table 1). Spacing between seed(ling) tubers in all locations was 25 cm. In Hilvarenbeek, the farmer had adopted a bed system, which was better adapted to his conditions. Due to the fine sand at this location, ridges would be prone to erosion.
Design and Measurements
At preparation of the field trials, the weight of the seed(ling) tubers for each plot was recorded. Trials were planted mechanically in a randomized complete block design with three repetitions and 20 plants per plot, divided over two rows. Between each plot, two border plants of the cultivar Miss Blush were planted to obtain an equal border effect over all plots. The bi-colored cultivar Miss Blush was chosen to avoid mixture at harvest, as test cultivars and diploid hybrids showed uniform skin color.
The whole experiment was bordered by two ridges of Miss Blush at the sides and one full plot of Miss Blush at the beginning and end of each ridge. During the growing season, stem number was counted per plot. After harvest, tuber number and size were measured automatically using a 3D camera (RMA Techniek, ‘s Heer Arendskerke, Netherlands). Total tuber weight was determined per plot. A subsample of approximately 5 kg was used to determine dry matter content (see below for formulas).
Data Analysis
Data of the yield trials were statistically analyzed using R (R Core Team 2017). A component analysis was done on total yield (Formula 1) to determine how these components contribute to total yield in each cultivar. Yield and yield components were adjusted for spatial field trends with Spatial Analysis of Field Trials with Splines (SpATS) (Rodríguez-Álvarez et al. 2018). In this model, differences in plant development due to environmental variation within the field are distinguished from differences in the genetic compositions between the cultivars. In SpATS, variation in the field is analyzed in two directions (rows and columns of the trial) using P-splines; effects of spatial variation are described by a standard mixed model (Rodríguez-Álvarez et al. 2018). By adjusting the data using SpATS, effects of environmental variation within trial fields were reduced, as the focus of this analysis was the genetic differences and the differences between trial fields. To meet the assumptions of normality of the SpATS model, data were log transformed, means and standard errors were back-transformed to linear data for graphical presentation. To examine to what extent the different yield components contribute to total yield, the variation in each yield component was compared with variation in total yield by linear regression. The contribution was defined as the change in yield by changing one unit of the yield component. This was determined by the slope of a linear model, which was fitted through the data of all sites. Genotype by environment interaction was calculated with the AMMI analysis (Additive Main effects and Multiplicative Interaction models), using the Agricolae package in R (Crossa 1990). Results of the AMMI analysis are presented in Fig. 2 and in Table 2. For the AMMI analysis, the raw data were used; the rest of the presented data of yield, stems/m2, tubers/stem and tuber size were adjusted for spatial effects within the field using SpATS. Under water weight was used to determine dry matter content (Formula 3, 4) (Tiemens-Hulscher et al. 2013). Tuber shape was determined using Formula 5.
Results
Seedling Tuber Production from Seedlings
In contrast to regular potato seed tuber productions, seedling tubers were produced from diploid hybrid seedlings. After transplanting to the field, seedlings grew rapidly and hardly any plant died. For most hybrids, the canopy was more or less closed at the end of June. After harvest of the seedling-tuber production, the number of seedling tubers was assessed (Fig. 1). Although diploid hybrids were not selected for yielding capacity, a good seedling tuber production was achieved.

Frequencies of number of seedling tubers (35–55 mm) produced per plant
Yield Trials
Seed tubers produced from true hybrid seeds in 2016 were used to conduct multi-location yield trials in 2017. In these trials, the performance of diploid hybrids grown from tubers was compared to the performance of tetraploid cultivars in different environments. Diploid hybrids that were used in this experiment were the first test hybrids derived from a hybrid breeding system based on homozygous inbred lines (Lindhout et al. 2011). It is known that the correlation between yield of single greenhouse-grown plants and larger blocks in the field of the same cultivar is low. Therefore, traits for high yield were not considered as selection criteria during the selection of parent lines for these hybrids.
Yield
Among the different sites, significant differences (p < 0.001) in yield were found (Table 2). Highest yielding site was Hilvarenbeek with on average 54 Mg/ha, while lowest average yield was measured in Berthem with 27 Mg/ha. Average yield for the Netherlands and France was 43 Mg/ha, in Belgium 50 Mg/ha (Eurostat), so differences in production between sites were due to local environmental and field management differences.
Fresh tuber yield differed significantly among cultivars (p < 0.001). Diploid hybrids yielded on average between 16 and 52 Mg/ha. For the tetraploid cultivars, average yield was between 52 and 101 Mg/ha. Performance of cultivars among the different sites varied; indeed, a significant interaction (GxE, p < 0.001) in yield between cultivar and site was found (Table 2).
The genotype × environment interaction of the hybrids and cultivars was analyzed using the AMMI analysis. The AMMI parameters can be used to make a biplot in which the interaction effects of genotype and environment are shown (Gauch and Zobel 1997). In the biplot (Fig. 2), 62.9% of the GxE interaction is explained in the principal component axis PC1. Also, the principal component axis PC2 explained a significant part of the G × E interaction (23.6%); the rest of the PC axes were not significant. On average, the yield of diploid hybrids was lower than the yield of tetraploid cultivars; however, the highest yielding hybrids were similar to the lowest yielding tetraploid cultivars. For these higher yielding hybrids, the interaction with the environment was also similar to several tetraploid cultivars (Fig. 2). The large distance between the trial locations in Fig. 2 shows that they differed in environmental effect, except for Warmeriville and Berthem that were very close to each other (Fig. 2).

Biplot of yield and PC1 of the AMMI analysis (Additive Main effects and Multiplicative Interaction models)
Dry Matter
Dry matter percentage (DM%) was measured in each genotype on all fields separately. A wide genetic variation for DM% was observed, as well as variation in DM% among the different sites. Diploid hybrids showed a broad range of DM%, between 15.8 and 21% (Fig. 3). This was in range with most of the tetraploid cultivars, except for three chip cultivars with a DM% of more than 22.5% (Brooke, Pirol and Lady Rosetta). The rest of the chip and frying cultivars had a DM% between 19.2 and 21.3%, 12 of the diploid hybrids also had a DM% within this range. The rest of the hybrids had a DM% comparable with the tetraploid table cultivars that produced a DM% between 19.2 and 15.6. So in general, in this test panel, the diploid hybrids had similar DM% to the tetraploid cultivars.

Average dry matter (DM) percentage and coefficient of variation (CV) of DM% of diploid hybrids and tetraploid cultivars
To compare the extent of the environmental effect on genotypes, the coefficient of variation (CV) of DM% over the sites was calculated for each diploid hybrid and tetraploid cultivar (Fig. 3). Variation of DM% between fields in most diploid hybrids was in the same range as the variation of DM% in tetraploid cultivars. The most stable diploid hybrid (Hybrid 1) had a CV of 2.7%, which was similar to the CV of the tetraploid cultivar Innovator, while the DM% in hybrid 1 was even higher than in Innovator (21% and 19.4%, respectively). So, dry matter percentage and stability in different environments were as good as, and sometimes even better in hybrids than in tetraploid cultivars.
Tuber Shape
Tuber shape is an important trait for cultivars in the chip and the frying industry, because tubers need to be suitable for the processing machines (Tiemens-Hulscher et al. 2013). Tuber shape is here expressed by Formula 4, in which the length, width and height of individual tubers are taken into account. The result is a dimensionless number between 1 and 0, with 1 representing a perfect sphere. Without selection for specific tuber shape in the diploid hybrids during the breeding process, the same range from round to long was found in hybrids as in tetraploid cultivars (Fig. 4). The variation for long tubers for fries represented by frying cultivars like Innovator and Russet Burbank was similar to the variation of several hybrids, as well as a round shape represented by several chip cultivars like Hermes and Lady Rosetta.

Average tuber shape and coefficient of variation (CV) of tuber shape of diploid hybrids and tetraploid cultivars. Tuber shape is a dimensionless number between 1 and 0, with 1 representing a perfect sphere
The uniformity of the shape is important for mechanical processing. Therefore, the coefficient of variation of the shape data of the individual tubers was calculated for each hybrid and cultivar (Fig. 4). Most diploid hybrids had a higher CV than the tetraploid cultivars. On average, the CV for the hybrids was 20%, and for the tetraploid cultivars, it was 17%; however, the most stable hybrids showed overlap with the least stable tetraploid cultivars. To gain insight into the effects of the different environments on the distribution of shape, we show boxplots of two cultivars (Fig. 5). Both Hybrid 9 and Bintje had a stable shape across the different environments. The differences in mean shape and variation between the five locations were smaller for Hybrid 9 than for cultivar Bintje.

Boxplots of shape of individual tubers of tetraploid cultivar Bintje (n = 3484) and diploid hybrid 9 (n = 2782) grown at five different locations in 2017
Yield Components
Potato yield was analyzed as the product of different plant components that can contribute to yield. Individual tuber weight, number of tubers per stem and number of stems per surface unit were chosen as relevant yield components to compare the growth of diploid hybrids and tetraploid cultivars. These yield components were compared between tuber grown diploid hybrids and commercial tetraploid cultivars that grew in yield trials on five different locations.
Generally, the yield components differed between the tetraploid cultivars and the diploid hybrids. The mass of the individual tubers of diploid hybrids was lower than that of the tetraploid controls (71 vs. 177 g on average). The hybrids had on average a higher number of stems (25.6 stems/m2) than the tetraploids (14.9 stems/m2). The number of tubers per stem was higher in the tetraploids, compared to the diploids (2.9 and 1.8, respectively). This resulted in approximately equal number of tubers per unit area (45.2 and 43.5 tubers/m2 for diploids and tetraploids, respectively) and all hybrids performed with the number of tubers/m2 within the range of the tetraploid cultivars.
When comparing yield components of individual tetraploid cultivars and diploid hybrids, the differences were small. A positive correlation was found between yield and tubers per stem (p < 0.001). Indeed, the higher yielding hybrids had a relatively high number of tubers per stem. Hybrid 9 for example produced 2.3 tubers per stem on average, which was nearly as much as Annabelle (2.6) and more than Innovator (1.9). Tuber size also was positively correlated with yield (p < 0.001); also for the diploid hybrids, the larger tubers resulted in higher yield, even though all hybrid tubers were relatively small.
The cultivars with higher yield also had a relatively high number of tubers per m2. For example, Bintje showed a high number of tubers/m2 (60.3) and the highest yield (7.2 kg/m2) across all cultivars, while Hybrid 1 had the lowest yield (1.6 kg/m2) and a low number of tubers m-2 (31.2). In higher yielding hybrids, the number of tubers/m2 was also high, with both Hybrid 9 and tetraploid cultivar Bintje having the largest number of tubers/m2.
Contribution of Yield Components to Yield
Variation in yield was found across sites as well as between cultivars. The variation in yield was due to the difference in responses of individual genotypes to the environments. Although there are trade-offs and interdependencies between yield components, they explain differences in yield between cultivars and environments.
The number of tubers per stem showed the largest variation across sites (CV = 24.8%); between Berthem and Hilvarenbeek, the difference was almost a factor two. For stems/m2, the variation was much lower (CV = 15.3%) and the weight of the tubers was rather stable (CV = 3.7%). In conclusion, the yield component that was affected most by the environment was the number of tubers per stem.
To compare the extent to which different yield components were affected by the environment between cultivars, the contribution of individual yield components to total yield over the different sites was determined by calculating the slope between total yield and yield component across sites (Table 3 and Table S1). It estimated the yield gain for a single yield component for each cultivar, with a positive value indicating a contribution to higher yield when the specific yield component increases, and a negative value indicating a loss in total yield when increasing a specific yield component.
To compare the relative effects of yield components on yield between the tetraploid cultivars and the diploid hybrids, correlations between yield components and yield were calculated. Every pair of average yield and yield component value of each location was used in the regression analysis, data are shown in the last three columns of Table 3. It compared the difference in contribution to yield of yield components. Number of tubers per stem explained 80% of the variation in yield in tetraploids and 90% in the diploid hybrids. The relative effect of number of stems per m2 on yield was much smaller: 18% in tetraploid cultivars and 5% in hybrids. Tuber size affected yield least in tetraploids and hybrids (2% and 5%, respectively). So, on average, hybrids and tetraploid commercial cultivars shared the most important contributing factor to yield.
A higher number of tubers per stem contributed positively to total yield in all cultivars (Table 3). For diploid hybrids in the selection of Table 3, the contribution was between 426 and 3161 g yield increase per m2 for each extra tuber per stem. In the tetraploids, this was between 1164 and 3387 g/m2 yield increase/(tuber/stem). More stems per m2 led in some hybrids to a decrease in total yield while in others, total yield increased with more stems per m2 (contribution between − 174 and 867 g/m2 yield increase/(stems/m2) in Table 3). In the tetraploid cultivars, higher stem numbers were positively correlated with yield with contributions between 165 and 956 yield g/m2 yield increase/(stems/m2). There was a large variation in the contribution of larger tubers to total yield between diploid hybrids as well as in tetraploid cultivars, with values between − 8 and 175 g/m2 yield increase/(g/tuber) for hybrids and − 101 and 5 g/m2 yield increase/(g/tuber) for tetraploid cultivars. Overall, the range of yield contribution by the different yield components was overlapped between diploid hybrids and tetraploid cultivars.
Variation was found between the different cultivars in total yield as well as in the contribution of the separate yield components to yield. To examine whether there is an optimal value for each yield component for high yield, the gain in yield with an increase of the yield component was compared with the average value of that yield component for each cultivar. Although yield gain and increase in yield component are not completely independent, it provides insight into whether there are optimal yield component values.
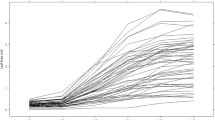
Genotypic variation was found for the contribution of yield components to total yield. The effect of differences in yield components between genotypes on the correlation between yield component and yield was examined (Figs 6, 7 and 8). Most genotypes showed a positive correlation between yield and tubers/stem when the average number of tubers/stem was between 1.0 and 3.8, so for this dataset, more tubers per stem contributed positively to total yield in the whole range.

Contribution of more tubers per stem to total yield, expressed in yield increase per extra tuber/m2, compared to average number of tubers per stem over five sites (Berthem, Giethoorn, Hilvarenbeek, Est, Warmeriville)

Contribution of more stems per m2 to total yield, expressed in total yield increase per extra stem/m2, compared to average number of stems/m2 over five sites (Berthem, Giethoorn, Hilvarenbeek, Est, Warmeriville)

Contribution of larger tubers to total yield, expressed in total yield increase per extra g tuber (tuber size), compared to average tuber size over five sites (Berthem, Giethoorn, Hilvarenbeek, Est, Warmeriville)
In contrast to the number of tubers/stem, larger tubers also affected yield negatively in some cultivars (Fig. 8). A negative relation was found between contribution of larger tubers to total yield and average tuber size of a genotype (Fig. 8). Until a tuber size of about 90 g, the production of larger tubers resulted in a higher total yield. When a genotype already produced large tubers on average, larger tubers decreased total yield. So, with increasing average tuber size of a genotype contribution of even larger tubers to total yield decreased.
Discussion
In this research, we have presented results of replicated field trials across multiple locations of diploid hybrids in comparison with tetraploid cultivars. We have analyzed the yield components to explain differences in yield level and stability.
Hybrid Seedling-Tuber Production
To perform yield trials in which diploid hybrids could be benchmarked against tetraploid commercial cultivars, we first produced seedling tubers from diploid hybrid seeds. This was done to decrease the difference in starting material, as plant performance is largely affected by the starting material (Arsenault et al. 2001; Struik and Wiersema 2001). The breeding process of the test hybrids that were used in this trial was done exclusively in the greenhouse, so no selection for yield was applied.
Seedling tubers that were produced were used in the subsequent year for yield trials. The physiological age of seed tubers is one of the main determinants for seed tuber quality (Struik and Wiersema 2001) and it is affected by many genetic and environmental factors such as management of the seed tuber crop, storage conditions, seed tuber size and the state of dormancy (Struik 2007). Before the seed tubers were selected for the ware crop trial, sprouting was scored. The results showed that dormancy was broken in all tubers; however, there was some variation probably due to different genetic background of the diploid hybrids.
To minimise the experimental error, caused by differences in the physiological conditions of the tubers, seed tubers were stored under optimal conditions and equally sprouting tubers were selected for the yield trial. If present, the largest unexplained error would occur in the diploid hybrids, due to differences in physiological conditions as the tubers of the different hybrids were not stored under optimal conditions for each hybrid. In addition, some genetic variation within the hybrids may still occur as the parents were not 100% homozygous. However, the within-plot variation of the hybrids was not higher than this variation within commercial cultivars (data not shown). So, this supports the conclusion that the differences between the hybrids and the commercial cultivars were due to genetic differences.
Yield Trials
Yield trials were conducted to examine yield potential of diploid hybrids, the GxE interaction and to compare yield components between diploid hybrids and tetraploid cultivars. The trials were conducted at five sites in France, Belgium and Netherlands (3 sites) from which insight into the variation due to genetics, the different environments and their interactions was obtained.
Performance of Diploid Hybrids and Tetraploid Cultivars
For a long time, the assumption that tetraploid potatoes are inherently higher yielding than diploid potatoes was based in a large part on the untested hypothesis that having four allelic variants per locus contributes to yield superiority (Bani-Aameur et al. 1991; Jong and Tai 1991; Werner and Peloquin 1991; Buso et al. 1999). When grown in a farmer-managed ware crop, we have shown that two of the diploid hybrids and cultivar Annabelle show similar average yields with 48, 52 and 52 Mg/ha, respectively, thereby proving the yield potential of diploid hybrids. This was suprising; as diploid hybrids were never selected for high yielding under field conditions, they were not expected to keep up with yield of tetraploid commercial cultivars in which selection for high yield has been performed for hundreds of years.
These results are supported by trials of Hutten (1994) and Jansky et al. (2016) in which they demonstrated yield potential of diploid potato.
Next to fresh tuber yield, other important criteria that determine the economic value of a cultivar were assessed. The diploid hybrids that were used in this trial had a range for DM% as well as for tuber shape that overlapped with the tetraploid cultivars for chips and fries (Hermes and Innovator). As heritability of tuber shape is high (Bisognin et al. 2012; Lindqvist-Kreuze et al. 2015; Meijer et al. 2018), it is a trait that can be selected for efficiently.
Environmental Effect on Plant Performance
Insight into the stability of desired traits across different environments is important to be able to select for desired trait properties efficiently in the breeding process. These diploid hybrids were a set of first test hybrids, developed while the breeding programme was still immature. Therefore, more variation was expected in the hybrids than in the tetraploid cultivars, which were clones. This expected variation was investigated for yield as well as a number of relevant traits.
Yield of diploid hybrids and tetraploid cultivars was affected by the environment. In line with the results found in this study, potato tuber yield is known to be highly influenced by different environments (Haverkort 1990). Several approaches of measuring yield stability are possible (Lin et al. 1986). In this study, the AMMI analysis was used as a useful method to distinguish the effects of genotype, environment and their interaction (Gauch et al. 2008). The first principal component axis PC1 explained 62.9% of the GxE interaction and is therefore used to discuss the interaction. Although we cannot attribute differences in performance between hybrids or tetraploid commercial cultivars to specific environmental conditions, we were able to determine that yields of both diploid hybrids and tetraploid cultivars were affected by GxE. The magnitude on the PC1 axis was comparable between hybrids and cultivars; however, the scores of the tetraploid cultivars generally were higher. A higher score of a cultivar in a specific environment indicates a better performance in that environment, while genotypes that score close to zero perform more constant across environments (Gauch and Zobel 1997). Even though the hybrids generally had lower yields and performed less stable across the environments than cultivars, there were also hybrids that perform similar to tetraploid cultivars in both yield and stability in this set of unselected test-hybrids.
Dry matter content of both diploid hybrids and tetraploid cultivars was affected similarly by the environment. When stability of a cultivar for DM% is high, efficient selection for specific dry matter percentages is possible (Wang et al. 2017). Although there was still genetic variability within the diploid hybrids and the hybrids were never tested in the field before, overall stability of the performance was similar to clonally propagated tetraploid cultivars. This genetic variability may have led to the slightly higher CV of shape in diploid hybrids compared to tetraploid cultivars; however, it was small in effect, which is in line with earlier reports (Yildirim and Celal 1985; Love et al. 1997).
The variation of yield components in the different environments was compared between the diploid hybrids and the tetraploid cultivars. The relation of the three different yield components was similar for tetraploids and diploids: for both, the number of tubers that was produced per stem was affected most by the environment, followed by number of stems per m2 and finally tuber size.
No difference was found in the average effect of environment on yield components between hybrids and tetraploid cultivars. This comparison shows that diploid hybrid cultivars express similar variation to various environments as commercial tetraploid cultivars despite the genetic variation that is still present in these first test hybrids, and the difference in starting material (seedling tubers compared to normal seed tubers). Moreover, it shows that yield and dry matter are highly affected by the environment, so multiple environments are needed for selection, while tuber shape is more stable across different environments.
Contribution of Yield Components to Total Yield
The variation of yield components in different environments was compared to the difference in total yield between those environments to evaluate the contribution of each yield component to the total yield. An increase in number of tubers produced per stem showed the strongest positive effect on yield in diploid hybrids as well as in tetraploid cultivars. For tetraploid cultivars, a similar effect of number of tubers per plant was found by Asghari-Zakaria et al. (2007). In contrast to the effect of the number of stems, the effect of tuber size on yield differed between tetraploid cultivars and hybrids. The commercial tetraploid cultivars already produced relatively large tubers and larger tubers did not result in more yield or affected yield even negatively. This finding is in contrast to the results of Asghari-Zakaria et al. (2007) who showed a positive correlation between tuber size and yield. However, as both studies contained only 1 year of field trials, different findings could be explained by year effects. The results suggest that there is an optimum in tuber size above which larger tuber size has a neutral or negative effect on total yield.
The distribution of yield components was different between diploid hybrid cultivars and tetraploid cultivars. Diploids had smaller tubers and fewer tubers per stem, but more stems/m2 than tetraploids. The large number of stems/m2 in diploid hybrids even affected total yield negatively, while in all tetraploid cultivars, an increase of stem density had a positive effect on yield. Firman and Allen (2007) showed that increasing stem density up to 10 stems/m2 resulted in a higher total yield, but higher densities did not affect total yield, and in our study, we found an optimum of 18 stems/m2. With larger stem densities, the tuber size distribution can be steered: higher density results in more smaller tubers (Firman and Allen 2007). Indeed, in this experiment, a negative correlation was found between stem density and tuber size.
As most commercial tetraploids cultivars produced larger tubers than diploid hybrids, the effect of increased tuber weight was larger in hybrids than in tetraploid cultivars. The estimated optimal tuber size was 100 g/tuber and larger tubers did not contribute to higher total yield or even resulted in lower yield. A larger number of tubers per stem resulted in a higher total yield in most diploid hybrids and tetraploid commercial cultivars. This comparison shows the similarity of distribution of yield components contributing to yield in diploid hybrids and tetraploid cultivars. In addition, it gives insight in which yield components need most focus in the diploid hybrid cultivars to improve yield potential.
Implications for Breeding
The analysis of yield components was performed for different environments and the variation of specific plant properties in different environments was evaluated. With this information, we determined traits that need to be tested in several environments to make reliable selections, for example yield, dry matter and tuber number were affected by the environment. Specific yield components that can be used as selection criteria in a breeding program were identified. In this trial, optimal configurations of yield components for the tested environments were found: a tuber size of approximately 100 g/tuber and a stem density of about 18 stems/m2, with as many tubers as possible as more tubers always increased total yield. These configurations can be used as breeding targets. Moreover, we were able to identify more stable hybrids over more variable ones, and hence we can start selecting for trait stability.
Conclusion
In this study, we compared the performance of diploid test-hybrid cultivars to tetraploid commercial ones in field trials at representative locations in the NW-European potato-growing area. In these trials, some diploid hybrids have shown a yield potential comparable to the tetraploid cultivars. For major economically relevant traits such as dry matter percentage, tuber number/m2 and tuber size, the diploid hybrids only slightly underperformed compared to the tetraploid cultivars. For yield, the best hybrids performed as good as the lower yielding tetraploid cultivars, the diploid hybrid cultivars having never been selected under field conditions. Stability of these traits across different locations was remarkably similar to the that of tetraploid cultivars. Moreover, the stability was different for the measured traits. For making selections in breeding, this information helps to determine the number of environments that is needed to test a certain trait. There was a large overlap in performance of the diploid hybrids and tetraploid cultivars for all the yield components analyzed (stems per m2, tubers per stem and tuber size). In both tetraploid cultivars and diploid hybrids, the number of tubers per stem affected the total yield most, followed by stems/m2. Tuber size had the smallest effect on total yield. With the variation for these traits present in diploids, desired properties can be combined in a cultivar through hybrid breeding which is competitive with the current commercial tetraploid cultivars.
References
Arsenault WJ, LeBlanc DA, Tai GCC, Boswall P (2001) Effects of nitrogen application and seedpiece spacing on yield and tuber size distribution in eight potato cultivars. Am J Potato Res 78(4):301–309
Asghari-Zakaria D, Fathi M, Hasan-Panah D (2007) Sequential Path Analysis of Yield Components in Potato. Potato Res. 49(4):273–279
Bani-Aameur F, Lauer FI, Veilleux RE, Hilali A (1991) Genomic Composition of 4x–2x Potato Hybrids: Influence of Solanum Chacoense. Genome 34(3):413–420
Bisognin DA, Rigão MA, Lopes SJ, Storck L (2012) Heritability and Correlation among Potato Tuber Traits. Crop Breed. Appl. Biotechnol. 12(3):215–219
Buso JA, Boiteux LS, Peloquin SJ (1999) Multitrait Selection System Using Populations with a Small Number of Interploid (4x-2x) Hybrid Seedlings in Potato: Degree of High-Parent Heterosis for Yield and Frequency of Clones Combining Quantitative Agronomic Traits. Theor Appl Genet 99(1–2):81–91
Crossa J (1990) Statistical analysis of multilocation trials. Adv. Agron. 44:55–85
Douches DS, Maas D, Jastrzebski K, Chase RW (1996) Assessment of potato breeding progress in the USA over the last century. Crop Sci 26(6):1544–1552
Endelman JB, Jansky SH (2016) Genetic Mapping with an Inbred Line-Derived F2 Population in Potato. Theor Appl Genet 129(5):935–943
Firman DM, Allen EJ (2007) Potato Biology and Biotechnology: Advances and Perspectives. Chapter 33: Agronomic practices. Elsevier, Amsterdam. 719–38
Gauch H, Piepho H, Annicchiarico P (2008) Statistical Analysis of Yield Trials by AMMI and GGE: Further Considerations. Crop Sci 48. https://doi.org/10.2135/cropsci2007.09.0513
Gauch HG, Zobel R (1997) Identifying mega-environment and targeting genotypes. Crop Sci 37:381–385
Jong H, Tai GCC (1991) Evaluation of Potato Hybrids Obtained from Tetraploid-Diploid Crosses I. Parent-Offspring Relationships. Plant Breeding 107(3):177–182
Haverkort AJ (1990) Ecology of Potato Cropping Systems in Relation to Latitude and Altitude. Agr. Syst. 32(3):251–272
Haverkort AJ, Struik PC (2015) Yield Levels of Potato Crops: Recent Achievements and Future Prospects. Field Crop. Res. 182:76–85
Haverkort AJ, van de Waart M, Bodlaender KBA (1990) Interrelationships of the Number of Initial Sprouts, Stems, Stolons and Tubers per Potato Plant. Potato Res 33(2):269–274
Hosaka K, Hanneman RE (1998) Genetics of self-compatibility in a self-compatible wild diploid potato species Solanum chacoense. 2. Localization of an S- locus inhibitor (Sli) gene on the potato genome using DNA markers. Euphytica 103:265–271
Hutten RCB (1994) Basic Aspects of Potato Breeding via the Diploid Level. Wageningen
Jansky SH, Charkowski AO, Douches DS, Gusmini G, Richael C, Bethke PC, Spooner DM, Novy RG, De Jong H, De Jong WS, Bamberg JB, Tomphson AL, Bizimungu B, Holm DG, Brown CR, Haynes KG, Sathuvalli VR, Veilleux RE, Miller C, Bradeen JM, Jiang J (2016) Reinventing Potato as a Diploid Inbred Line-Based Crop. Crop Sci 56(4):1412–1422
Jeuken MJW, Lindhout P (2004) The Development of Lettuce Backcross Inbred Lines (BILs) for Exploitation of the Lactuca Saligna (Wild Lettuce) Germplasm. Theor Appl Genet 109(2):394–401
Kooman PL, Fahem M, Tegera P, Haverkort AJ (1996) Effects of Climate on Different Potato Genotypes 2. Dry Matter Allocation and Duration of the Growth Cycle. Eur J Agron 5(3–4):207–217
Lin CS, Binns MR, Lefkovitch LP (1986) Stability Analysis: Where Do We Stand? Crop Sci 26:894–900
Lindhout P, de Vries M, ter Maat M, Su Y, Viquez-Zamora M, van Heusden S (2018) Hybrid Potato Breeding for Improved Varieties. Achieving Sustainable Cultivation of Potatoes 1(1):1–24
Lindhout P, Meijer D, Schotte T, Hutten RCB, Visser RGF, van Eck HJ (2011) Towards F 1 Hybrid Seed Potato Breeding. Potato Res 54(4):301–312
Lindqvist-Kreuze H, Khan A, Salas E, Meiyalaghan S, Thomson S, Gomez R, Bonierbale M (2015) Tuber Shape and Eye Depth Variation in a Diploid Family of Andean Potatoes. BMC Geneti 16(1):1–10
Love SL, Werner BK, Pavek JJ (1997) Selection for Individual Traits in the Early Generations of a Potato Breeding Program Dedicated to Producing Cultivars with Tubers Having Long Shape and Russet Skin. Am Potato J 74(3):199–213
Lynch DR, Foroud N, Kozub GC, Fames BC (1995) The Effect of Moisture Stress at Three Growth Stages on the Yield, Components of Yield and Processing Quality of Eight Potato Varieties. Am Potato J 72:375–385
Meijer D, Viquez-Zamora M, van Eck HJ, Hutten RCB, Su Y, Rothengatter R, Visser RGF, Lindhout WH, van Heusden AW (2018) QTL Mapping in Diploid Potato by Using Selfed Progenies of the Cross S. Tuberosum × S. Chacoense. Euphytica 214(7):121
NAK (2018) Inspection of seed potatoes; https://www.nak.nl/wp-content/uploads/2018/10/Inspection-of-seed-potatoes-DIS-2018-definitief.pdf
Piepho HP, Laidig F, Drobek T, Meyer U (2014) Dissecting Genetic and Non-genetic Sources of Long-term Yield Trend in German Official Variety Trials. Theor Appl Genet 127(5):1009–1018
Prinzenberg AE, Víquez-Zamora M, Harbinson J, Lindhout P, van Heusden S (2018) Chlorophyll Fluorescence Imaging Reveals Genetic Variation and Loci for a Photosynthetic Trait in Diploid Potato. Physiol Plant 164(2):163–175
R Core Team (2017) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org/
Rodríguez-Álvarez MX, Boer MP, van Eeuwijk FA, Eilers PHC (2018) Correcting for Spatial Heterogeneity in Plant Breeding Experiments with P-Splines. Spat Stat 23:52–71
Struik PC (2007) The Canon of Potato Science: 40. Physiological Age of Seed Tubers. Potato Res 50:375–377
Struik PC, Wiersema SG (2001) Seed Potato Technology. Wageningen Academic Publishers, The Netherlands
Su Y, Viquez-Zamora M; den Uil D; Sinnige J; Kruyt H; Vossen J; Lindhout P; van Heusden S (2019) Introgression of Genes for Resistance against Phytophthora infestans in Diploid Potato. American Journal of Potato Research
Tiemens-Hulscher M, Delleman J, Eising J, Lammerts van Bueren ET (2013) Potato Breeding. Drukkerij De Swart, Den Haag
Wang Y, Snodgrass LB, Bethke PC, Bussan AJ, Holm DG, Novy RG, Pavek MJ, Porter GA, Rosen CJ, Sathuvalli V, Thompson AL, Thornton MT, Endelman JB (2017) Reliability of Measurement and Genotype × Environment Interaction for Potato Specific Gravity. Crop Sci 57:1–7
Werner JE, Peloquin SJ (1991) Yield and Tuber Characteristics of 4x Progeny from 2x x 2x Crosses. Potato Res 34(3):261–267
Yildirim MB, Celal F (1985) Genotype x environment interactions in potato. Am Potato J62(7):371–375
Zaheer K, Akhtar MH (2016) Potato Production, Usage, and Nutrition—A Review. Crit Rev Food Sci Nutr 56(5):711–721. https://doi.org/10.1080/10408398.2012.724479
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
ESM 1
(DOCX 38.1 kb)
Rights and permissions
About this article

Cite this article
Stockem, J., de Vries, M., van Nieuwenhuizen, E. et al. Contribution and Stability of Yield Components of Diploid Hybrid Potato. Potato Res. 63, 345–366 (2020). https://doi.org/10.1007/s11540-019-09444-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11540-019-09444-x