
Abstract
The success of any potato breeding programme depends upon the selection of superior parents and cross combinations, and superior clones from the segregating progenies. However, due to the tetrasomic inheritance and heterozygous nature of cultivated potato, potato breeding has remained empirical, cumbersome and time-consuming. Potato breeders have used a number of approaches to simplify and improve the efficiency of potato breeding. These include use of mid-parent/mid-self values, genetic diversity, combining ability, estimated breeding values and progeny tests for the selection of parents and cross combinations. The selection of superior clones from the segregating progeny has been hampered by low efficiency of selection in early generations particularly for tuber yield and its components. The possibility of in vitro selection and production of microtubers for early large scale trials has been explored. The procedures used and constraints observed are reviewed and discussed in this paper.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Potato (Solanum tuberosum L.), the most important non-cereal food crop of the world, is grown in over 140 countries on an area of 19.32 million ha with world production of 368.37 million tons (FAO 2013). It has high nutritional value, therefore, has been declared food for future by the FAO, especially in the developing countries. The average world productivity of potato is only 19 t/ha, which is much lower than the potential yield of 120 t/ha (Pandey et al. 2005). Genetic improvement can be one of the way-forwards to increase the productivity and quality of potatoes. However, the development of new potato cultivars through breeding is a laborious and time-consuming process. It is mainly because the methods for potato breeding are not very efficient and have not changed much in the twentieth century (Neele et al. 1988; Caligari 1992; Tarn et al. 1992; Douches and Jastrzebski 1993; Gopal 2006).
Potato breeding is based on a phenotypic recurrent selection scheme that cycle every 5 to 9 years. Another limitation in potato improvement that the breeder has been faced with is the challenge of tetrasomic inheritance, imposed by this auto- or segmental allotetraploid crop (2n = 4x = 48) which results in complicated genetic segregation (Howard 1970; Ross 1986; Matsubayasthi 1991). For a character in tetraploid potato, there is a possibility of involvement of four alleles at each locus. Consequently, each locus has the potential for several first-, second- and third-order allelic interactions (Bonierbale et al. 1993). Moreover, in the most extreme case, eight different alleles may independently segregate in populations, resulting from crosses between two tetraploid parents. This implies that even a single locus can display a continuous variation (Meyer et al. 1998). This decreases the probability of finding transgressive segregants in an autotetraploid population below that of a diploid crop (Wricke and Weber 1986). Nearly 40 traits including yield and quality attributes as well as tolerance to various biotic and abiotic stresses are considered in potato breeding thereby making varietal development in potato very demanding (Gebhardt 2013). However, potatoes also have the advantage of vegetative propagation due to which a clone though initially may be a single plant can be multiplied with all its inter- and intra-allelic gene actions intact, and if found promising it can be adopted as a variety for commercial cultivation.
Two basic steps in any potato breeding programme are to identify (1) superior parents and cross combinations to produce segregating progenies and (2) select superior clones from the progenies generated. However, the choice of parents and cross combination and superior clones from the segregating progeny has always puzzled the potato breeders. Though, recent advances in molecular breeding provide opportunities for rapid genetic gain (Slater et al. 2014a), yet the use of molecular approaches requires quantitative genetic analysis of the highly heterozygous breeding populations for improvement of the complex yield and quality traits with low heritability. Thus, phenotypic selection still remains the common practice in potato breeding programmes. This paper reviews the status and the constraints encountered in conventional potato breeding and the way-forward to improve the efficiency of selection. To our knowledge, this is the first review of its kind on this important topic in potato.
Selection of Superior Parents and Cross Combinations
To conduct a successful potato breeding programmme for a character such as yielding ability, it is necessary to select parental lines capable of transmitting the character to their offspring. The methods used to determine this and their effectiveness in potato are reviewed below.
Combining Ability
Information on combining ability of parents can help breeders to decide their hybridization strategy. Parental lines which transmit superior performance for a character to their offspring when crossed with a wide variety of other clones are said to have a good general combining ability (GCA) for that character. The deviation of a specific cross from what is expected on the basis of the GCA of the parents is called specific combining ability (SCA). When GCA proves significant, parents with good GCA can be used again in future crosses, and continued progress can be sought over a number of generations of multitrait genotypic recurrent selection. When SCA is also present, the progeny tests can be used to identify the best crosses for clonal evaluation and cultivar production.
For many economically important traits, it has been possible to partition genetical variation into components due to GCA and SCA. Combining ability analysis of the tetraploid parents could be performed by crossing them to a common set of testers and determining which produces desirable progeny at the highest frequency. Half-diallel crossing designs were used by Killick (1977), while North-Carolina II design was employed by Plaisted et al. (1962) and Killick and Malcolmson (1973). Line × tester analysis on a pattern similar to North Carolina II designs has been used by Gopal (1998a, b). In these test cross methods, a large set of crosses is required to be made in which selected females are crossed with a random sample of testers in a specific pattern based on the design used. To simplify GCA estimation in potato, Gopal et al. (2008) studied the effectiveness of top cross vs. poly cross as alternative to test cross method. They compared GCA of 12 females estimated based on matings with specific single testers (top cross) and three types of bulk pollen (poly cross) with those estimated by 72 (12 × 6) test crosses, using common testers in three types of matings. The study was conducted for 2 years at each of the two locations representing short-days sub-tropical conditions in plains and long-days temperate conditions in hills of India. Matings based on selected individual single testers (top cross) resulted in GCA estimates similar to those based on a number of testers as were evident from the significant and high correlation coefficients (Table 1). Among the six testers used, the tester CP 2110 could estimate the GCA of the females with reasonably high accuracy both for tuber yield and general impression, and CP 2351 for reaction to late blight. However, bulk pollen matings (poly cross) were not an effective alternative for estimating GCA of females based on their matings with the bulked testers individually (test cross). This shows that top cross involving a selected tester can be an effective alternative to test cross method, and this could substantially reduce the number of crosses required for estimating the GCA. More studies are required to identify the effective testers for other characters and environmental conditions for top cross method of GCA estimation.
In making comparisons between the estimates of GCA and SCA variances, it is tempting to evaluate their relative size. A combining ability model states that the observed phenotype of an individual is equal to a mean from which all other effects are deviations, an average or additive effect of each of the alleles, a dominance effect, an epistatic effect of the interaction of alleles at different loci, an effect due to environment in which the individual is growing and an effect due to the interaction between the genotype of the individual and its environment. The additive effect of a gene may be looked upon as the average effect of a gene substitution of one allele for another taken over all possible combinations of remaining genotypes weighted by their relative frequency. The dominance effect is the interaction between alleles at the same locus. The epistatic effects are the special effects produced by combinations of genes at different loci that are different from the total average effects of the allelic combinations present at each of the loci. Epistatic effects can be subdivided according to the number of loci which are considered at a time. If only the epistatic effects of two loci are considered, the total epistatic effect is the sum of interaction between the additive effects of two different loci (additive × additive), the additive effect of one locus and the dominance effect of the other (additive × dominance) and the dominance effects of two loci (dominance × dominance). Thus, when a clone is selected as having good GCA, this means that the algebraic sum of the additive, dominance and epistatic effects passed on to its offspring produced a favourable result in excess of the average of all the clones tested.
Several studies of combining abilities are available for a number of characters in potato (Table 2). There is some evidence that combining ability is an important feature of the expression of certain disease resistances also (Tung et al. 1990; Gopal 1998b; Gopal et al. 2008). However, contradictory results have been obtained with regard to the relative importance of GCA and SCA for various characters (Table 2).
Gopal (1998a) studied the GCA and its repeatability over three early generations, i.e. seedling generation (S), first clonal generation (C1) and second clonal generation (C2) in potato on 72 progenies generated from 18 × 4 (line × tester) factorial matings. They reported that repeatability for GCA as judged from correlation coefficients between generations ranged between 0.18 and 0.82 for tuber yield and its components. Correlation coefficients between generations for GCA for breeders’ preference score, plant vigour, tuber colour, tuber shape and uniformity in tuber colour were higher and ranged between 0.27 and 0.95. They concluded that GCA had moderate repeatability over generations for various characters. Kumar and Gopal (2006) studied combing ability in Andigena potatoes by evaluating the progenies of 11 × 3 factorial matings from seedling to third clonal generations. They found that both GCA and SCA could be reliably estimated for uniformity in tuber shape, uniformity in tuber size, uniformity in tuber colour, predominant tuber shape and predominant tuber size in early generations starting from the S. Whereas these parameters for tuber yield, average tuber weight and tuber number could be reliably estimated from C2 onward. Ruiz de Galarreta et al. (2006) made a partial diallele between 14 potato cultivars and found that both GCA and SCA were significant in all early generations. The correlations between generations and between characters showed that for tuber yield no relation was obtained between generations; however, average tuber weight and yield were positively associated in all generations. They concluded that appropriate selection criteria depend strongly on the particular cross.
In potato, diploid clones which form both n and 2n gametes are available. This suggests the feasibility of studying their combining abilities simultaneously for diploid and tetraploid levels. A given diploid clone would be mated to other diploids and to tetraploids. The former series of matings give rise to diploid progenies whose performance evaluates the breeding value of n gametes produced by that diploid parent. The latter series of matings yield tetraploid progenies which, upon evaluation of their performance, are expected to provide information on the breeding value of the 2n gametes produced by the same diploid parent. Mendiburu and Peloquin (1977) reported that in 2x-2x and 4x-2x matings non-additive genetic effects were predominant in determination of tuber yields at both the diploid and the tetraploid levels. Keijzer van der Stoel et al. (1991) reported that in 2x-2x crosses the SCA variance for tuber yield appears to be more important than the GCA variation, while the opposite was true for tuber dry matter. In the 4x-2x crosses, however, the GCA variance was greater than the SCA variances. Keijzer van der Stoel et al. (1991) also found that the GCA of the parents for tuber yield in the 2x-2x and 4x-2x crosses in S did not show a similar pattern as in the C1. The GCA for tuber dry matter, however, showed a greater correspondence between the generations. Digenic interaction exists in the 2n gametes. Such interactions are expected to be strong in those formed by the first division restitution (FDR) mechanism as they preserve a large proportion of heterozygous loci between the homologous chromosomes of 2n gamete producing diploid parents and 4x parents.
This literature survey shows that the answer to the question whether GCA or SCA is the more important in the same quantitatively inherited characters was not always same. According to Plaisted et al. (1962), larger estimates of SCA variance than the corresponding GCA variance are the characteristics of the tetraploid potatoes. They further suggested that informal previous selection which narrows the genetic base of the tested genotypes may also lead to greater estimates of SCA variance for various characters. Killick and Malcolmson (1973), using a concept developed in evolutionary population genetics, suggested that traits subjected to directional selection would be expected to show little additive genetic variance, but a large degree of dominance and epistasis, whereas the reverse would be true for traits subjected to stabilizing selection. The study conducted by Gopal (1998a) endorsed both of these concepts. However, based on various reports on combining ability, it is concluded that the relative importance of GCA or SCA in a population may depend on kind of material, experimental design and/or environmental conditions.
Nevertheless, the combining ability analysis has proven to be helpful in identifying the promising parents for use in potato breeding programmes. However, keeping in view poor repeatability of GCAs, the information on combining ability needs to be sought afresh for identifying promising parents. Further, combining ability analysis requires making the crosses in specific pattern depending upon the design used, which is generally difficult in practical potato breeding programmes due to problems of large scale sterility and incompatibility in potato resulting in many years of work to complete a combining ability study (Plaisted et al. 1962; Plaisted and Patterson 1963; Killick and Malcolmson 1973; Killick 1977; Gopal 1994, 2006). In such situations, progeny test is an alternative to planned combining ability studies.
Progeny Test
Since genetic analysis has proven difficult in potato, progeny testing is an effective tool to evaluate the breeding value of parents and specific cross combinations both at tetraploid, diploid and tetraploid × diploid levels. The concept of progeny evaluation is not new and has been suggested in other crops (Simmonds 1979, 1996). The progeny tests which have been developed and validated in potato (Bradshaw and Mackay 1994; Bradshaw et al. 1995; Gopal 1997) are of great value for genetical studies because they can be done within a year of making crosses as part of a breeding programme. These can be used to identify the best crosses particularly for traits that display continuous variation and for which, in general, non-additive inheritance is more important. De Jong and Tai (1991) reported that in 4x-2x crosses, a relatively low degree of determination (R 2) of both 4x and 2x parents on the performance of their 4x-2x hybrid progenies was found for various quantitative traits including tuber yield implying thereby that test crossing and subsequent progeny analysis is necessary before such parents can be used in a breeding programme.
Tai and De Jong (1980) conducted multivariate analyses of ten hybrid families of group Tuberosum × (group Phureja × haploid group Tuberosum) parents. The homogeneity of within family VAR-COV matrices and the significance of the mean vector between families confirmed the need of identifying superior progenies before raising a large population of seedlings for selection work. The progeny testing was also found to be useful for assessing the value of parents as in the canonical diagram no clear relation existed between the hybrid families and the parents.
The correlation coefficients between progeny means in different generations of potato breeding have been reported to be weak (Caligari et al. 1986), but also moderate to strong (Brown and Caligari 1986, 1989; Maris 1969, 1988). Gopal (1997) reported that moderately effective selection based on progeny means could be conducted for plant vigour, tuber yield, tuber number, average tuber weight, breeders’ preference score, tuber colour, tuber shape, and uniformity in tuber colour, tuber shape and tuber size. The correlation coefficients between progeny means in different generations for tuber dry matter were reported to be strong (Maris 1962, 1969; Neele and Louwes 1989).
The glasshouse progeny test for breeders’ preference score has been shown to correlate well with progeny mean preference scores made in the C1 and C2 at both seed and ware sites (Brown et al. 1988; Brown and Caligari 1989). This univariate cross prediction method had been tested by Caligari and Brown (1986) also. They reported that actual numbers of clones falling in the desired category were also a good cross predictor as a reasonably good agreement between predicted and observed numbers was seen over sites and years for breeders’ preference score. Based on similar studies, Brown et al. (1987a, b) and Bradshaw et al. (1998) reported that seedling progeny evaluation by breeders’ preference score could be used to reject entire crosses on the ground that they were less likely than others to contain clones of commercial worth. Brown and Caligari (1989) found this method to be effective for total tuber weight (yield) also, but not for mean tuber weight and number of tubers. Total tuber weight has been reported to be an important component of breeders’ preference score in early generations (Brown and Caligari 1986; Neele et al. 1991; Bradshaw et al. 1998). Tai (1975) reported that out of five characters recorded, tuber appearance and average weight of marketable tubers made important contribution to the mean visual scores (breeder’s preference score).
Brown and Caligari (1988) examined three methods of cross prediction namely multivariate probabilities, the sum of ranks and the frequency of genotypes in a sample that transgress set target values. The characters tuber weight, mean tuber weight, number of tubers and regularity of tuber shape were examined. It was found that a sample as small as 25 clones provided good predictions (as judged by the observed frequencies in larger progeny samples examined in various environments). The best estimates were obtained using multivariate probabilities based on the means, within progeny variation and the phenotypic correlation between variates. The rankings of the crosses according to these multivariate probabilities provided good indications of the number of clones which survived selection in an actual breeding scheme. Brown et al. (1988) found that the progeny mean was a slightly better predictor of superior crosses over generations than the mean and within progeny phenotypic variance. Caligari and Brown (1986) and Gopal (2001b) had also found that within progeny variation was not a major component in prediction. Gopal (2001b) reported that family selection based on progeny mean is almost as effective as the one based on both progeny mean and within family standard deviation.
Brown et al. (1988) had reported that predictions based on the expected proportions of clones that would transgress a given target value was better than the prediction based on the observed frequency of desirable clones in a progeny sample. However, Gopal (1997) did not find much difference in the effectiveness of these two types of proportions with regard to effectiveness of prediction. From a practical point, breeders would be mainly interested in knowing whether the proportion of clones expected to transgress a particular target value in later generations can be predicted from the observed or predicted proportions transgressing the target value in an earlier generation. Further, all these studies (Caligari and Brown 1986; Brown et al. 1988; Gopal 1997) had reported that progeny means were better than the proportions for cross prediction.
Neele and Louwes (1989) reported that progeny selection is effective for chip colour when glucose tests strips were used for indirect selection of chip colour. Hayes and Thill (2002) reported that family selection for chip colour is effective even in greenhouse grown progenies as they found significant correlations between family means of greenhouse grown and field grown tubers. Cross prediction based on progeny mean has also been practised for tuber blight and foliage blight (Caligari et al. 1983) as well as potato cyst nematodes Globodera pallida (Phillips 1981).
Haynes et al. (2012) studied if parents that produce progeny with broad adaptation could be identified early in a potato breeding programme. For this seed of all second field generation clones were distributed to five locations for selection. Based on common selections made at all the locations, they concluded that early generation selection can be used to identify parents that produce broadly adapted progeny. Because potato displays such strong G × E interactions, this selection approach may be particularly beneficial for selecting potato cultivars with broad adaptation.
Estimated Breeding Values
Best linear unbiased prediction (BLUP) is commonly practised in animal breeding. Recently, Slater et al. (2014b) used BLUP in potato breeding after modifying it to suit autotetraploid potato as suggested by Kerr et al. (2012). It is an advanced biometric technique that permits the analysis of a large data set derived from pedigree for the calculations of expected breeding values (EVP). Since BLUP considers information from all relatives in the analysis, it improves the accuracy of analysis, particularly for low heritability traits. Slater et al. (2014b) showed that BLUP analysis based on a large pedigree, provided EBVs for the families, and all cultivars in the pedigree and the individual genotypes themselves, potentially enabling breeding from the best progeny. EBVs also improved cross-generational predictions and superior genotypes could be identified for retention that would otherwise have been lost by the rejection of the entire low scoring families thereby demonstrating the advantage of using EBVs over progeny means.
Mid-parent and Mid-self Values
In the absence of progeny means, mid-parent values can be used at the start of the breeding programme to predict the mean performance of crosses (Bradshaw et al. 2003). As early as 1929, Krantz and Hutchin reported that average maturity of F1 potato progenies was closely associated with the mid-parent maturity value. Mid-parent values, however, are effective only if the character(s) under improvement are controlled by mainly additive gene actions, e.g. foliage maturity (Killick 1977). As most of the quantitative characters in potato are non-additive in nature, mid-parent values act as only preliminary screens to compliment the progeny means which are required to be estimated later. For tuber yield correlation coefficients between progeny means and mid-parent values, or parental values were found to be moderate or low (Brown and Caligari 1989; McHale and Lauer 1981; Maris 1989; Schroeder and Peloquin 1983; Veilleux and Lauer 1981). For tuber dry matter, however, mid-parent values were moderately to strongly associated with progeny means (Maris 1989; Neele and Louwes 1989). Gopal (1998c) found that the mid-parent values were ineffective as correlation coefficient between mid-parent values and progeny means was in general not significant for most of the characters. Bradshaw et al. (2003) reported that when an outlier was excluded from the data for fry colour, a moderate correlation of 0.71 was found for the mid-parent values and offspring means, which was assessed in different years. An even higher correlation of 0.86 was found by Bradshaw et al. (2000) in a diallel set of crosses in which parents and progenies were assessed in the same trial. They suggested that the use of mid-parent values for fry colour is justified. Neele (1990) reported that time of tuber initiation of the progenies was associated with the time of tuber initiation of the earliest tuberizing parent. The mean increase rate of the harvest index of the progenies was equal to the mean values of parents. He thus suggested that differences in harvest index between progenies can be predicted with a high degree of precision by using the harvest index parameters of the parents. This estimate of harvest index was further found to have a fairly good correlation to tuber yield.
In analogy to mid-parent, mid-self values have also been tested for cross prediction. Compared with test crosses, this method has the advantage that it requires only one offspring population per evaluated parent, but it can only be applied to male-fertile parental clones. As compared with mid-parent value, evaluation for mid-self values, however, is more time- and labour-consuming than simply evaluating the parents. Brown and Caligari (1989) reported that mid-self values provided better prediction than mid-parent values for mean tuber weight and number of tubers. Gopal (1998c) too reported that in contrast to mid-parent values, mid-self values were, in general, better for cross prediction. This, they suggested, may be because mid-self values reflect the potential of the parents based on the performance of new genotypic combinations produced after selfing, whereas parents per se performance/mid-parent values are based on the phenotypic performance of the parents, which may or may not be reflected in the progeny. Neele et al. (1991), however, reported that mid-self value did not improve the prediction of progeny means compared with the mid-parent value. They further reported that predictions based on the test crosses were the best and surpassed those of the mid-parent value.
Genetic Diversity of Parents
The positive relation between genetic diversity of parents and tuber yield of their progeny has been confirmed by many authors in potato, including Rowe (1967), Mendoza and Haynes (1974b), Mok and Peloquin (1975), Cubillos and Plaisted (1976), De Jong et al. (1981), Veilleux and Lauer (1981), Landeo and Hanneman (1982), Schmiediche et al. (1982), Estrada-Ramos (1984), Gopal and Minocha (1997a) and Luthra et al. (2001, 2005). Negative heterotic effects (inbreeding depression) for yield and other characters are common for within Gp. Tuberosum crosses (Maris 1969, 1989; Tarn and Tai 1983; Tai 1994); as a result new varieties have not made a major impact on yield improvement of potatoes (Van der Plank 1946; Seiffert 1957; Howard 1963a, b; Gopal 2006) in most of the countries. This is attributed to the relatively narrow genetic base of this subspecies (Mendoza and Haynes 1974a); most tuberosum varieties, then, are more or less closely related to each other (Gopal and Oyama 2005) being descended from a small number of introductions of South American potatoes (Simmonds 1962). Gp. Andigena also has narrow genetic base (Maris 1989). Thus, it has been suggested that high inbreeding coefficients in the progenies should be avoided. However, Loiselle et al. (1989) reported that inbreeding coefficient was of little use in predicting a priori the selection pattern of a cross in a multitrait type of breeding programme. They attributed this to several factors. One is the large influence of non-heritable variation of potato clones in visual selection of early breeding generations (Tai and Young 1984; Gopal et al. 1992). Another is the changing emphasis on different traits as the selection process progresses, which results in no significant correlation between selections at succeeding stages (Tai and Young 1984; Gopal et al. 1992). Selection primarily retains genotypes of a ‘commercial’ type. The emphasis is placed on good tuber size and a high marketable yield and not on late maturing, vigorous plants. Inbreeding coefficient is negatively associated with vigour and tuber yield. Thus, heterosis breeding is essential for individual traits like tuber yield although inbreeding coefficient may not be of direct use in selection of parents for a multitrait like breeding programme.
Paz and Veilleux (1997) used RAPD markers to measure genetic distance among doubled monoploids of S. phureja and found that largest parental genetic distance was always associated with the highest total tuber yield among progenies of doubled monoploids. They, thus, suggested that RAPD markers may facilitate the identification of diverse parents to maximize the expression of heterosis. Chimote et al. (2008) used 20 RAPD primers, two chloroplast DNA deletions based and one plastid-based SSR markers and three mitochondrial SCAR markers to differentiate 40 parental lines used in an Indian potato breeding programme aimed at developing early bulking varieties. They reported that three parents with Andigena type cytoplasm and four with wild-type cytoplasm were distinct from the rest. They suggested that divergent parents may be useful in planning hybridization aimed at effective utilization of heterosis.
Gopal and Minocha (1997a) estimated genetic distances among 22 tetraploid clones of potato based on morphological characters recorded on plants grown under six in vitro and four in vivo conditions. They found that genetic distances under in vitro conditions had no relationship with the heterosis for tuber yield observed in the 72 progenies (18 × 4) of these parents in three clonal generations, whereas those under in vivo conditions had good relationship with the observed heterosis. Further, they reported that relationship between genetic distances and heterosis was stronger when parents were evaluated under the conditions similar to those under which crosses were evaluated. This reflects the importance of G × E interaction on the use of apparent genetic diversity determined on phenotypic basis of parents for predicting the progeny’s performance (Gopal 2006). It is concluded that genetic divergence though of use in identifying parents for exploiting heterosis in progeny has only moderate effectiveness and that too when G × E interaction is taken care of.
Selection of Superior Clones in the Segregating Progeny
In most potato breeding programmes, a large number of seedlings are grown from which a few promising clones are retained following selection in early generations. Wenzel et al. (1983) calculated that 500,000 to 2,000,000 seedlings must be evaluated in order to select a single new cultivar. This estimate is still consistent with the practices used for most modern breeding programmes. These populations must be quickly reduced to a manageable number before statistically accurate and consistent tests can be performed. Tai and Young (1984) suggested that it was appropriate to eliminate 99% of the available genotypes in the first 3 years of selection. Unfortunately, many studies (Maris 1962, 1966, 1988; Swiezynski 1968; von Kameke 1975, 1978; Tai 1975; Anderson 1977; Brown et al. 1984, 1987b; Brown and Caligari 1986; Caligari et al. 1986; Neele et al. 1988, 1989; Gopal et al. 1992; Bae et al. 2008) have shown early generation selection to be ineffective in identifying superior genotypes. Davies and Johnston (1965, 1968) reported that the selection in early generations is likely to be greatly affected by bias of the selectors; different selectors selecting from the same material will often select different plants. Anderson and Howard (1981) pointed that although selection for non-subjective traits like resistance to various diseases and pests may be effective in these generations, it would not be so for subjective characters like tuber yield, its components, stolon length, etc. They reported that considerable differences occurred between selectors in intensity of selection when selecting the same potato clones in S, C1 and C2. Comparing clones selected or discarded in 1 year with clones selected or discarded in previous years indicated poor effectiveness of selection. A high proportion of the clones selected in C2 would always have been discarded in S or C1. Davies and Johnston (1974) reported that in C1 stage single-hill system selection was ineffective even when clones were replicated twice, as very small percentage of clones selected from two replications were same. Neele et al. (1988) reported that the use of two-plant plots instead of one-plant ones in C1 helped reduce the environmental variance, but it had only slightly positive effect on the selection efficiency. In two separate studies, looking at the agreement between C1 materials selected in two locations, both Blomquist and Lauer (1962) and Davies and Johnston (1965) found that a small percentage of clones were selected at both locations. Only 2.3% of the clones were selected at two locations in Minnesota (Blomquist and Lauer 1962). In Davies and Johnston’s (1965) study, both researchers selected at the New Brunswick and Ontario, Canada sites: Davies selected 5.2% of the clones at both locations, Johnston selected 7.5% of the clones at both locations. These and many other studies (Maris 1962, 1966, 1988; Swiezynski 1968; von Kameke 1975; 1978; Tai 1975; Anderson 1977; Brown et al. 1984, 1987a; Caligari et al. 1986; Brown and Caligari 1986; Neele et al. 1988, 1989; Gopal et al. 1992; Bae et al. 2008) thus did not suggest stringent selection in early generations. Rather, in some cases, selection in S as well as C1 was at the best random, sometimes even having a negative effect. Hence, Pfeffer (1963), Krug et al. (1974), Tai and Young (1984) and Brown et al. (1984) suggested that seedling stage should only provide seed tubers for growing in the field the following year. In contrast, however, Howard (1963a) had suggested that selection in seedling stage can be made against long stolons, low yield and long tubers. He viewed that clones which produce very low yields as seedlings could be discarded as they were unlikely to produce high yield in C1. But other studies (Brown et al. 1984; Gopal et al. 1992) showed that the seedlings which produce very small tubers should not be discarded at this stage.
The size of tubers produced in S is genetically controlled, however, it effects on the performance of a clone in C1 and later clonal generations is non-genetic thereby adversely influencing efficiency of selection for yield, yield components and breeders’ preference in the early generations of a potato breeding programme (Tai 1975; Brown et al. 1984, 1987a; Brown 1988; Gopal et al. 1992). It is a common practice in a number of potato breeding programmes (Tai and Young 1984; Fitschen 1984; Scholz 1986) to retain and plant a single tuber, usually the largest, from each seedling. Swiezynski (1978) demonstrated that the selection of individual tubers was very effective in the elimination of clones with irregular tuber shape and deep eyes. Previously, Swiezynski (1959) had shown that big tubers are less variable in specific gravity. The size of the tuber planted can influence the number of clones that are visually selected by breeders (Blomquist and Lauer 1962; Brown et al. 1984; Maris 1986). Krantz and Bailey (1929) and Krantz and Hutchins (1929) observed a delay in maturity of C1 plants, associated with reduced seed tuber size and that this effect could be carried over to C2 due to smaller tubers produced in C1. The conditions in which seedlings are grown can also greatly influence the efficiency of selection in C1 (Krug et al. 1974), and it has been suggested that the efficiency of selection particularly for tuber characters, can be increased if seedlings are grown under field conditions or in large pots as this results in more and bigger tubers (Zadina 1971; Swiezynski 1978; Gopal et al. 1992). Hayes and Thill (2002) reported that large amounts of non heritable variation seem to be associated with the clonal chip scores of green house grown tubers and selection for good chipping genotypes by evaluating greenhouse grown tubers would seem to result in at least a high degree of false positive selection. For similar reason, Louwes and Neele (1987) had suggested the use of bigger pots or field if chip quality and specific gravity of the seedling populations are to be evaluated.
Keeping in view the poor efficiency of selection in early generations, studies have been taken to improve the selection efficiency in these generations. For example, Love et al. (1997) reported that the selection for breeders’ preference score in early clonal generations is unreliable because it is based on unreplicated small plots and there is significant G × E interaction and high error variance. In order to improve the efficacy of selection, they suggested separation of breeder’s preference, which has large G × E interaction, into its components traits, some of which may have a small G × E variance. This strategy was also tried by Gopal et al. (1992). However, Neele et al. (1991) found that the expected and observed selection response of the component traits was equivalent to that of breeders’ preference and hence concluded that selection for breeders’ preference could not be improved by separately selecting for component characters like tuber yield and tuber appearance. On the other hand, Gopal et al. (1992) reported that positive selection for tuber colour, tuber shape, skin smoothness, non-cracking etc. can be done in seedling generation whereas no rejection should be done in this generation for tuber yield and its components. They further reported that seedlings of poor vigour may be rejected prior to transplanting in the field as they are likely to have low yields in later generations (Fig. 1). Ortiz and Golmirzaie (2003) too had found significant and positive correlations between plant vigour after transplanting the seedlings and their tuber yield. In order to define the selectability of visible traits, Love et al. (1997) evaluated 27 traits through three clonal generations. They reported that positive selection in early generations was possible for degree of skin russeting and the incidence and severity of skin defects. Negative selection was found to be appropriate for incidence of secondary growth, growth cracks, tuber malformations, tuber curvature, pointed ends, the incidence and severity of russet patchiness, eye depth, tuber shape, yield, tuber number, average tuber weight and flatness. Traits too inconsistent for use in selection in early generations included uniformity of skin russeting, uniformity in tuber shape and uniformity in tuber size. Neele and Louwes (1989) reported that no rejection should be done based on chip colour in S, and for tuber dry matter too, the rejection should not be stringent. They further reported that tuber glucose contents as measured with Reflocheck Glucose test strips were strongly correlated with the colour of the chips, but Gluko-test strips were less reliable. Dalianis et al. (1966)) reported that in early generation selection for after cooking blackening at least four tubers should be cooked. Hayes and Thill (2003) reported that selection for cold chipping was successful in S.

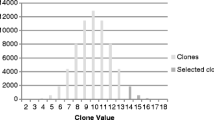
Mean tuber yield in different generations in the clones grouped on the basis of plant vigour in the seedling generation. Source: Gopal (2006)
Tai and Young (1984) suggested that moderate selection intensity over several generations would give a better balance between genetic advance and loss of valuable genotypes. Neele et al. (1989) concluded from selection studies that retaining 20 to 50% of the first year field seedlings followed by a 20% selection in the second clonal year would be the most economical selection procedure. This mild selection would also result in a greater proportion of clones in the late-maturing classes and higher specific gravity. For visual characteristics, Neele et al. (1991) suggested that negative selection be applied according to heritability to avoid rejecting too many valuable genotypes at an early stage.
Swiezynski (1984) reported that selection in S for potato viruses PVX, PVY, PVA and PVS was effective, but not for PLRV before the C1. Possibilities of early selection for PVM resistance are not yet clear. Levy et al. (1991) reported that the selection for heat tolerance was possible in the S. The seedlings that did not form tubers in the hot glasshouse, but were subsequently transferred to the cool glasshouse, tuberization was much enhanced and clones selected through this dual procedure exhibited tolerance to heat in C1. Reynolds and Ewing (1989) used the procedure in which plants were assessed for vigour of shoot growth and later the ability to tuberize under high temperature for selecting heat tolerant genotypes. The heat tolerance was, however, associated with earliness (Levy 1986).
Keeping in view the low efficiency of selection in early generations for agronomic characters, it has been suggested (Gopal 1997) that progeny selection should be practised prior to clonal selection in early generations. As described above, progeny selection can be exploited for making initial choices in identifying the crosses which have a high probability of producing clones with desirable phenotypes. Those progenies which have low probability can be discarded in S. This can be adopted in large breeding programmes: a large number (may be hundreds of crosses) may be evaluated in S by raising as few as 50–60 genotypes per cross. Negative selection (rejection of poor phenotype) may be practised for the characters under evaluation and a larger population of the selected crosses can be raised to practise within-progeny individual clonal selection (Gopal 1997). Gopal et al. (1992, 1994) suggested the following selection procedure for individual clonal selection in early generations.
-
1.
Seedlings of poor vigour may be rejected prior to transplanting in the field as they represent inferior genotypes.
-
2.
Those clones with undesirable tuber colour, tuber shape, eye depth and tuber cracking may be rejected from the S onwards as these characters have high repeatability.
-
3.
No rejection should be done on the basis of tuber yield, average tuber weight or number of tubers in the S.
-
4.
Negative selection for tuber yield and tuber weight can be initiated from the C1 onwards, whereas the number of tubers can be considered for the rejection of undesirable types from the C2 onwards.
-
5.
Since both the number of tubers and tuber size contribute to tuber yield, and number of tubers and tuber size are negatively associated with each other, it is suggested that a standard be fixed for the minimum number of tubers required in the selected types, before employing average tuber weight as a selection parameter for tuber yield, otherwise selection may result in retention of genotypes with oversized and fewer tubers.
In order to further improve the efficiency of selection in early generations, Gopal and Minocha (1997b; c) explored the possibility of using in vitro micropropagation techniques (Gopal et al. 1998). A considerable similarity was observed between in vitro and in vivo system with regard to genetic parameters and character associations in potato (Gopal 2001a). In vitro selection was found to be highly effective for tuber colour, number of eyes, and moderately effective for average tuber weight, plant vigour and foliage senescence under specific conditions (Gopal and Minocha 1997b). They postulated that in vitro selection may be effective even for tuber yield, but need is to simulate and identify the in vitro conditions under which genotypic differences are reflected in phenotypes more realistically as they are expressed in vivo.
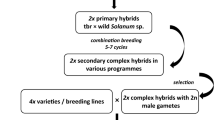
The problem of few tubers in early generations can be overcome by using in vitro propagation to produce microtubers. The production of microtubers, however, will involve cost of tissue culture and labour. In order to reduce this, microtuber production can be started after the harvest of S with the clones selected following the procedure described here. The number of clones required to be multiplied this way can be further reduced if breeding for tuber yield and its components is combined with resistance to diseases such as late blight. Susceptible genotypes can be discarded in the S itself (Caligari et al. 1983; Stewart et al. 1983). Further, commercial scale tissue culture devises likes fermentors and bioreactors can be used to reduce the time and cost of microtuber production (Donnelly et al. 2003). The benefit:cost analysis will depend on the corresponding direct and indirect costs involved in microtuber production and conducting of field trials for extra years due to limited tubers in early generations when microtubers are not produced. In the crop raised from microtubers, selection can be done for general impression (Gopal and Minocha 1997c; Gopal et al. 2002), giving less weightage to tuber yield. This procedure of individual clonal selection would help in an early and safe elimination of unproductive genotypes. The production of microtubers in sufficient numbers, and the subsequent production of normal seed tubers from them, will result in earlier large scale trials thereby substantially reducing the period taken to develop a variety. The suggested selection procedures based on only in vivo propagation and also combined with in vitro propagation is illustrated in Figure 2.

Stepwise procedure for selection of superior clones for agronomic characters on potato breeding indicating the time advantage of combining in vitro and in vivo methods. Source: Gopal (2006)
The selected types from the early generations can be evaluated in bigger and multilocational trials for identifying the types widely adapted over environments. The number of environments and replications can be decided based on the number of genotypes under evaluation and the available number of tubers per genotype. Comstock and Moll (1963) stated that ‘genotype by environment interaction is in some way involved in most problems of quantitative genetics and many problems of plant breeding’. Genotype by environment interactions have been recognized by plant breeders for many years as reducing the efficiency of selection (Hill 1975). In practical potato breeding programmes, limited seed supply and logistical problems involved in the handling of large number of progeny usually restrict the selection of early generation clones to one location. Further, the selection in later generations is also usually made at one location on each year’s results, and only those genotypes that are selected in 1 year are grown in the subsequent year. It is only very advanced few selections that are evaluated in multilocation trials over years. As in other crops, the best performing genotypes are selected for release as cultivars based on performance over years and locations.
Conclusion
The various strategies for the selection of superior parents and cross-combinations in potato breeding programmes need to be used in combination, as none of these at its own are efficient enough to lead to reliable results, although the progeny test and EBV are good methods to select for superior parents and cross combinations. In the absence of combining ability, EBV or progeny test information, mid-parent/mid-self values and genetic divergence among parents can serve as a preliminary guide particularly in the initial years of a potato breeding programme. In early generations of progeny evaluation for the selection of superior clones, the selection pressure needs to be nil to low, and only very poor types should be rejected (negative selection) in the first three generations for traits like tuber yield and its components. Positive selection can be practised for qualitative characters and disease/pest resistance in early generations itself depending upon the stability of expression of the character(s) concerned. The production of microtubers in sufficient numbers can help in early large scale trials and reduce the time required in development of potato varieties.
Abbreviations
- GCA:
-
General combining ability
- SCA:
-
Specific combining ability
- S:
-
Seedling generation
- C1:
-
First clonal generation
- C2:
-
Second clonal generation
- EBV:
-
Estimated breeding value
- RAPD:
-
Randomly amplified polymorphic DNA
- SSR:
-
Simple sequence repeat
- SCAR:
-
Sequence characterized amplified region
- G × E:
-
Genotype × environment
References
Anderson JAD (1977) Potato breeding. Selection for agronomic characters and resistance to cyst nematodes. MSc Thesis, Univ Camb pp 217
Anderson JAD, Howard HW (1981) Effectiveness of selection in the early stages of potato breeding programmes. Potato Res 24:289–299
Bae J, Jansky SH, Rouse DI (2008) The potential for early generation selection to identify potato clones with resistance to Verticillium wilt. Euphytica 164:385–393
Blomquist AW, Lauer FI (1962) First clonal generation potato progeny performance at two Minnesota locations. Am Potato J 39:460–463
Bonierbale MW, Plaisted RL, Tanksley SD (1993) A test of the maximum heterozygosity hypothesis using molecular markers in tetraploid potatoes. Theor Appl Genet 86:481–491
Bradshaw JE, Mackay GR (1994) Breeding strategies for propagated potatoes. In: Bradshaw JE, Mackay GR (eds) Potato Genetics. CABI Publishing, Wallingford, pp 467–497
Bradshaw JE, Stewart HE, Wastie RL, Dale MFB, Phillips MS (1995) Use of seedling progeny tests for genetical studies as part of a potato (Solanum tuberosum subsp. tuberosum) breeding programme. Theor Appl Genet 90:899–905
Bradshaw JE, Dale MFB, Swan GEL, Todd D, Wilson RN (1998) Early-generation selection between and within pair crosses in a potato (Solanum tuberosum subsp. tuberosum) breeding programme. Theor Appl Genet 97:1331–1339
Bradshaw JE, Todd D, Wilson RN (2000) Use of tuber progeny tests for genetical studies as part of a potato (Solanum tuberosum subsp. tuberosum) breeding programme. Theor Appl Genet 100:772–781
Bradshaw JE, Dale MFB, Mackay GR (2003) Use of mid-parent values and progeny tests to increase the efficiency of potato breeding for combined processing quality and disease and pest resistance. Theor Appl Genet 107:36–42
Brown J (1988) The effect of the weight of the seedling-derived tuber on subsequent clonal generations in a potato breeding programme. Ann Appl Biol 113:69–78
Brown J, Caligari PDS (1986) The efficiency of seedling selection for yield and yield components in a potato breeding programme. Z Pflanzenzuchtung 96:53–62
Brown J, Caligari PDS (1988) The use of multivariate cross prediction methods in the breeding of a clonally reproduced crop (Solanum tuberosum). Heredity 60:147–153
Brown J, Caligari PDS (1989) Cross prediction in a potato breeding programme by evaluation of parental material. Theor Appl Genet 77:246–252
Brown J, Caligari PDS, Mackay GR, Swan GEL (1984) The efficiency of seedling selection by visual preference in a potato breeding programme. J Agric Sci (Camb) 103:239–346
Brown J, Caligari PDS, Mackay GR, Swan GEL (1987a) The efficiency of visual selection in early generation of a potato breeding programme. Ann Appl Biol 110:357–363
Brown J, Caligari PDS, Mackay GR (1987b) The repeatability of progeny means in the early generations of a potato breeding programme. Ann Appl Biol 110:365–370
Brown J, Caligari PDS, Dale MFB, Swan GEL, Mackay GR (1988) The use of cross prediction methods in a practical potato breeding programme. Theor Appl Genet 76:33–38
Caligari PDS (1992) Breeding new varieties. In: Harris PM (ed) The potato crop, 2nd edn. Chapman and Hall, London, pp 334–372
Caligari PDS, Brown J (1986) The use of univariate cross prediction methods in the breeding of a clonal reproduced crop (Solanum tuberosum). Heredity 57:396–401
Caligari PDS, Mackay GR, Stewart HE, Waistie RL (1983) A seedling progeny test for resistance to potato foliage blight Phytophthora infestans (Mont.) de Bary). Potato Res 27:43–50
Caligari PDS, Brown J, Abbott RJ (1986) Selection for yield and yield components in the early generations of a potato breeding programme. Theor Appl Genet 73:218–222
Chimote VP, Kumar R, Sharma N, Kamal S (2008) Nuclear-cytoplasmic diversity in parental genotypes used for Indian early bulking potato (Solanum tuberosum ssp. tuberosum) breeding programme. Ind J Genet Plant Breed 68:171–176
Chitsay M (1984) Inheritance of factors affecting quality of processed potato (Solanum tuberosum group tuberosum L.). Diss Abstr Int B 45:24B
Comstock RE, Moll RH (1963) Genotype-environment interactions. In: Hanson WD, Robinson HF (eds) Statistical genetics and plant breeding. Nat Acad Sci Nat Res Pub 982:164-196
Cubillos AG, Plaisted RL (1976) Heterosis for yield in hybrids between S. tuberosum ssp. tuberosum and S. tuberosum ssp. andigena. Am Potato J 53:143–150
Dalianis CD, Plaisted RL, Patterson LC (1966) Selection for freedom from after cooking darkening in potato breeding programme. Am Potato J 43:207–215
Davies HT, Johnston GR (1965) First clonal generation potato seedling selection at two locations. Am Potato J 42:186–189
Davies HT, Johnston GR (1968) Second clonal generation potato seedling selection at two locations. Am Potato J 45:150–153
Davies HT, Johnston GR (1974) Reliability of potato selection in the first clonal generation. Am Potato J 51:8–11
Dayal TR, Upadhya MD, Chaturvedi SN (1985) Combining ability for tuber yield and other traits in the potato (Solanum tuberosum L.). J Ind Potato Assoc 12:122–137
De Jong H, Rowe PR (1971) Inbreeding in cultivated diploid potatoes. Potato Res 14:74–83
De Jong H, Tai GCC (1991) Evaluation of potato hybrids obtained from tetrapoid-diploid crosses. I. Parent-offspring relationships. Plant Breed 107:177–182
De Jong H, Tai GCC, Russel WA, Jonston GR, Proudfoot KG (1981) Yield potential and genotype-environment interactions of tetraploid-diploid (4x-2x) potato hybrid. Am Potato J 58:191–199
Donnelly DJ, Coleman WK, Coleman SE (2003) Potato microtubers: production and performance: A review. Am J Potato Res 80:103–115
Douches DS, Jastrzebski K (1993) Potato. In: Kaloo G, Bergh BO (eds) Genetic Improvement of Vegetable Crops. Pergamon Press, Oxford, pp 605–644
Estrada-Ramos N (1984) Acaphu: Tetraploid fertile breeding line selected from a Solanum acaule x S. phureja cross. Am Potato J 61:1–8
FAO (2013) FAOSTAT, Food and Agriculture Organization, Rome (www.faostat.fao.org)
Fitschen HJ (1984) Züchtung neuer Kartoffelsorten. Kartoffelbau 35:52–55
Gaur PC, Gopal J, Rana MS (1983) Combining ability for yield, its components and tuber dry matter in potato. Ind J Agric Sci 53:876–879
Gaur PC, Gopal J, Rana MS (1985) Combining ability performance of some recently developed potato hybrids. J Ind Potato Assoc 12:195–198
Gaur PC, Pandey SK, Singh SV (1993) Combining ability study in the development of potato hybrids suitable for processing. J Ind Potato Assoc 20:144–149
Gebhardt C (2013) Bridging the gap between genome analysis and precision breeding in potato. Trends Genet 29:248–256
Gopal J (1994) Flowering behaviour, male sterility and berry setting in tetraploid Solanum tuberosum germplasm. Euphytica 72:133–142
Gopal J (1997) Progeny selection for agronomic characters in early generations of a potato breeding programme. Theor Appl Genet 95:307–311
Gopal J (1998a) General combining ability and its repeatability in early generations of potato breeding programmes. Potato Res 41:21–28
Gopal J (1998b) Heterosis and combining ability analysis for resistance to early blight (Alternaria solani) in potato. Potato Res 41:311–317
Gopal J (1998c) Identification of superior parents and crosses in potato breeding programmes. Theor Appl Genet 96:287–293
Gopal J (2001a) In vitro and in vivo genetic parameters and character associations in potato. Euphytica 118:145–151
Gopal J (2001b) Between and within families variation and family selection in potato breeding programmes. J Genet Breed 55:313–318
Gopal J (2006) Considerations for successful breeding. In: Gopal J, Paul Khurana SM (eds) Handbook of potato production, improvement and postharvest management. Food Product Press, New York, USA, pp 77–108
Gopal J, Minocha JL (1997a) Genetic divergence for cross prediction in potato. Euphytica 97:269–275
Gopal J, Minocha JL (1997b) Effectiveness of in vitro selection for agronomic characters in potato. Euphytica 103:67–74
Gopal J, Minocha JL (1997c) Effectiveness of selection at microtuber crop level in potato. Plant Breed 116:293–295
Gopal J, Oyama K (2005) Genetic base of Indian potato selections as revealed by pedigree analysis. Euphytica 142:23–31
Gopal J, Gaur PC, Rana MS (1992) Early generation selection for agronomic characters in a potato breeding programme. Theor Appl Genet 84:709–713
Gopal J, Gaur PC, Rana MS (1994) Heritability, and inter-generation associations between tuber yield and its components in potato (Solanum tuberosum L.). Plant Breed 112:80–83
Gopal J, Minocha JL, Dhaliwal HS (1998) Microtuberization in potato (Solanum tuberosum L.). Plant Cell Rep 17:794–798
Gopal J, Kumar R, Kang GS (2002) Minituber crop and its effectiveness in selection for agronomic characters in potato breeding programmes. Potato Res 45:145–151
Gopal J, Kumar V, Luthra SK (2008) Top-cross vs. poly-cross as alternative to test-cross for estimating the general combining ability in potato. Plant Breed 127:441–445
Hayes RJ, Thill CA (2002) Co-current introgression of economically important traits in a potato breeding program. Am Potato J 79:173–181
Hayes RJ, Thill CA (2003) Genetic gain from early generation selection for cold chipping genotypes in potato. Plant Breed 122:158–163
Haynes KG, Gergela DM, Hutchinson CM, Yencho GC, Clough ME, Henninger MR, Halseth DE, Sandsted E, Porter GA, Ocaya PC (2012) Early generation selection at multiple locations may identify potato parents that produce more widely adapted progeny. Euphytica 186:573–583
Hill J (1975) Genotype-environment interactions—a challenge for plant breeding. J Agric Sci 85:477–493
Howard HW (1963a) Some potato breeding problems. Rep Pl Breed Inst 5-21
Howard HW (1963b) The significance of breeding in improving quality and yield. In: Ivins ID, Milthorpe FL (eds) The growth of the potato. Butterworths, London, pp 292–302
Howard HW (1970) Genetics of the potato. Logos Press Ltd., London, p 22
Keijzer van der Stoel MC, Pegels van Deelen MW, Neele AEF (1991) An analysis of the breeding value of diploid potato clones comparing 2x-2x and 4x-2x crosses. Euphytica 52:131–136
Kerr R, Li L, Tier B, Dutkowski G, McRae T (2012) Use of the numerator relationship matrix in genetic analysis of autopolyploid species. Theor Appl Genet 124:1271–1282
Killick RJ (1977) Genetic analysis of several traits in potatoes by means of a diallel cross. Ann Appl Biol 86:279–289
Killick RJ, Malcolmson JK (1973) Inheritance in potatoes of field resistance to late blight (Phytophthora infestans). Physiol Plant Pathol 3:121–131
Krantz FA, Bailey RM (1929) Relative productivity of certain types of potato seedling populations. Proceed Potato Assoc Am 56–63
Krantz FA, Hutchins AE (1929) Potato breeding methods II. Selection in inbred lines. Miinnesota Agr Expt Sta Tech Bull 58:1–23
Krug H, Wriedt G, Weber WE (1974) Untersuchungen zur Frühselektion in der Kartoffelzüchtung. II. Merkmalsbeziehungen zwischen den Generationen und innerhalb der Klongenerationen. Z Pflanzenzüchtg 73:163–193
Kumar R, Gopal R (2006) Repeatability of progeny mean, combining ability, heterosis and heterobeltiosis in early generations of a potato breeding programme. Potato Res 49:131–141
Landeo JA, Hanneman RE Jr (1982) Heterosis and combining ability of Solanum tuberosum group Andigena haploids. Potato Res 25:227–237
Levy D (1986) Genotypic variation in the response of potatoes (Solanum tuberosum L.) to high ambient temperatures and water deficit. Field Crop Res 15:85–96
Levy D, Kastenbaum E, Itzhak Y (1991) Evaluation of parents and selection for heat tolerance in the early generations of a potato (Solanum tuberosum L.) breeding program. Theor Appl Genet 82:130–136
Loiselle F, Tai GCC, Christie BR, Tarn TR (1989) Relationship between inbreeding coefficient and clonal selection in potato cultivar development program. Am Potato J 66:747–753
Louwes KM, Neele AEF (1987) Selection for chip quality and specific gravity of potato clones: Possibilities for early generation selection. Potato Res 30:241–251
Love SL, Werner BK, Pavek JJ (1997) Selection for individual traits in the early generations of a potato breeding program dedicated to producing cultivars with tubers having long shape and russet skin. Am Potato J 74:199–213
Luthra SK, Sharma PC, Gopal J, Khurana SMP, Pandey SK (2001) Inter-relationships between genetic parameters for tuber yield in potato. J Ind Potato Assoc 28:15–17
Luthra SK, Gopal J, Sharma PC (2005) Genetic divergence and its relationship with heterosis in potato. Potato J 32:37–42
Maris B (1962) Analysis van aardappelpopulaties ten dienste van de veredeling. Versl. Landbouwk. Onderz. 68.8. Wageningen, pp 208
Maris B (1966) The modifiability of characters important in potato breeding. Euphytica 15:18–31
Maris B (1969) Studies on maturity, yield, under water weight and some other characters of potato progenies. Euphytica 18:297–319
Maris B (1986) The effect of seed tuber weight on characters in the first and the second clonal generations of potato populations. Euphytica 35:465–482
Maris B (1988) Correlations within and between characters between and within generations as a measure for the early generation selection in potato breeding. Euphytica 37:205–224
Maris B (1989) Analysis of an incomplete diallel cross among three ssp. tuberosum varieties and seven long-day adapted ssp. andigena clones of the potato (Solanum tuberosum L.). Euphytica 41:163–182
Matsubayasthi M (1991) Phylogenetic relationships in the potato and its related species. In: Tsuchiya T, Gupta PK (eds) Chromosome engineering in plants: Genetics, breeding, evolution. Part B. Elsevier, Amsterdam, pp 93–118
McHale NA, Lauer FI (1981) Breeding value of 2n pollen from diploid hybrids and phureja in 4x-2x crosses in potatoes. Am Potato J 58:365–374
Mendiburu AO, Peloquin SJ (1977) Bilateral sexual polyploidization in potatoes. Euphytica 26:573–583
Mendoza HA, Haynes FL (1974a) Genetic relationship among potato cultivars grown in the United States. Hortic Sci 9:328–330
Mendoza HA, Haynes FL (1974b) Genetic basis of heterosis for yield in autotetraploid potato. Theor Appl Genet 45:21–25
Meyer RC, Milbourne D, Hackett CA, Bradshaw JE, Mc Nicol JW, Waugh R (1998) Linkage analysis in tetraploid potato and association of markers with quantitative resistance to late blight (Phytophthora infestans). Mol Gen Genet 259:150–160
Mok DWS, Peloquin SJ (1975) Breeding value of 2n pollen (dihaploids) in tetraploid x diploid crosses in potatoes. Theor Appl Genet 46:307–314
Mullin R, Lauer FI (1966) Breeding behaviour of F1 inbred potato clones. Proc Am Soc Hortic Sci 89:449–455
Neele AEF (1990) Study on the inheritance of potato tuber yield by means of harvest index, components and its consequences for choice of parental material. Euphytica 48:159–166
Neele AEF, Louwes KM (1989) Early selection for chip quality and dry matter content in potato seedling populations in greenhouse or screenhouse. Potato Res 32:293–300
Neele AEF, Barten JHM, Louwes KM (1988) Effects of plot size and selection intensity on efficiency of selection in the first Clonal generation of potato. Euphytica S:27–35
Neele AEF, Nab HJ, de Jongh MJ, de Loeuw LKM (1989) Optimising visual selection in early clonal generations of potato based on genetic and economic considerations. Theor Appl Genet 78:665–671
Neele AFE, Nab HJ, Louwes KM (1991) Identification of superior parents in a potato breeding programme. Theor Appl Genet 82:264–272
Ortiz R, Golmirzaie AM (2003) Genetic parameters for agronomic characteristics. I. Early and intermediate breeding populations of true potato seed. Hereditas-Lund 139:212–216
Pandey SK, Singh SV, Sarkar D (2005) Potato (Solanum tuberosum) for sustaining foodand nutritional security in developing world. Ind J Agric Sci 75:3–18
Paz MM, Veilleux RE (1997) Genetic diversity based on RAPD polymorphism and its relationship with the performance of diploid potato hybrids. J Am Soc Hortic Sci 122:740–747
Pfeffer C (1963) Vergleichende Untersuchungen über Auslesemöglichkeiten von in Freiland und in Töpfen kultivierten Kartoffelsämlingen. Züchter 33:6–11
Pfeffer C, Effmert M (1985) Breeding homozygous parents for resistance to potato scab caused by Streptomyces scabies (Thaxt.) Waksman et Henrici. Archiv für Züchtungsforschung15:325–333
Phillips MS (1981) A method of assessing potato seedling progenies for resistance to the white potato cyst nematodes. Potato Res 24:101–103
Phillips MS, Dale MFB (1982) Assessing potato seedling progenies for resistance to the white potato cyst-nematode. J Agric Sci 99:67–70
Phillips MS, Wilson LA, Forrest JMS (1979) General and specific combining ability of potato parents for resistance to the white cyst nematodes (Globodera pallida). J Agric Sci UK 92:255–256
Pika NA, Tarasenko VA (1985) Evaluation of potato varieties for combining ability for starch content by means of a two-tester top cross. Genetika 21:1856–1863
Pika NA, Tarasenko A, Mitsko VN (1984) Combining ability of potato varieties and hybrids for tuber flesh blackening after cooking. Sel Semenovod 7:16–17
Plaisted RL, Patterson LC (1963) Two cycles at phenotypic recurrent selection for high specific gravity. Am Potato J 40:396–402
Plaisted RL, Sanford L, Federer WT, Kerr AE, Peterson LC (1962) Specific and general combining ability for yield in potatoes. Am Potato J 39:185–197
Reynolds MP, Ewing EE (1989) Heat tolerance in tuber bearing Solanum species: a protocol for screening. Am Potato J 66:63–74
Ross H (1986) Potato breeding -problems and perspectives. Adv Plant Breed 13:132
Rowe PR (1967) Performance and variability of diploid and tetraploid potato families. Am Potato J 44:263–271
Ruiz de Galarreta JI, Pascualena J, Legorburu FJ, Barandalla L, Ritter E (2006) The history of potato (Solanum tuberosum L) research in Spain. Potato Res 49:19–25
Schmiediche P, Hawkes JG, Ochoa CM (1982) The breeding of the cultivated potato species Solanum x juzepczukii and S. x curtilobum. II. The resynthesis of S x juzepczukii and S. x curtilobum. Euphytica 31:695–708
Scholz M (1986) General strategy of potato breeding in the German Democratic Republic. In: Jellis GJ, Richardson DE (eds) The production of new potato varieties, technological advances. Cambridge University Press, Cambridge, pp 32–38
Schroeder SH, Peloquin SJ (1983) Parental effects for yield and tuber appearance on 4x families from 4x x 2x crosses. Am Potato J 60:819
Seiffert M (1957) Die Bedeutung der Züchtung für die Ertragssteigerung im Kartoffelbau in den letzten Jahrzehnten. Ein Beitrag zur Methodik der Ermittlung des züchterischen Fortschrittes. Züchter 27:1–22
Simmonds NW (1962) Variability in crop plants, its use and conservation. Biol Rev 37:422–465
Simmonds NW (1979) Principles of crop improvement. Longman Group, London
Simmonds NW (1996) Family selection in plant breeding. Euphytica 90:201–208
Slater AT, Cogan NOL, Hayes BJ, Schultz L, Dale MFB, Bryan GJ, Forster JW (2014a) Improving breeding efficiency in potato using molecular and quantitative genetics. Theor Appl Genet 127:2279–2292
Slater AT, Wilson GM, Cogan NOL, Forster JW, Hayes BJ (2014b) Improving the analysis of low heritability complex traits for enhanced genetic gain. Theor Appl Genet 127:809–820
Stewart HE, Taylor K, Wastie RL (1983) Resistance to late blight in foliage (Phytophthora infestans) of potatoes assessed as true seedlings and as adult plants in the glasshouse. Potato Res 26:363–366
Stewart HE, Wastie RL, Bradshaw JE, Brown J (1992) Inheritance of resistance to late blight in foliage and tubers of progenies from parents differing in resistance. Potato Res 35:313–319
Swiezynski K (1959) Selekeja ziemniakow na ccchy uzytknowe Cz. II. Zmiennose krzakowa. Hod Rosl Aklim Nas 3:685–715
Swiezynski KM (1968) Field production of first year potato seedlings in the breeding of early varieties. Eur Potato J 11:141–149
Swiezynski KM (1978) Selection of individual tubers in potato breeding. Theor Appl Genet 53:71–80
Swiezynski KM (1984) Early generation selection methods used in Polish potato breeding. Am Potato J 61:385–394
Tai GCC (1975) Effectiveness of visual selection for early clonal generation seedlings in potato. Crop Sci 15:15–18
Tai GCC (1976) Estimation of general and specific combining abilities in potato. Can J Genet Crop Sci 18:463–470
Tai GCC (1994) Use of 2n gametes. In: Bradshaw JE, Mackay GR (eds) Potato genetics. CAB International, Wallingford, pp 109–112
Tai GCC, De Jong H (1980) Multivariate analyses of potato hybrids. 1. Discrimination between tetraploid-diploid hybrid families and their relationship to cultivars. Can J Genet Cytol 22:227–235
Tai GCC, Young DA (1984) Early generation selection for important agronomic characters in a potato breeding population. Am Potato J 61:419–434
Tarn TR, Tai GCC (1983) Tuberosum x Tuberosum and Tuberosum x Andigena potato hybrids: comparisons of families and parents, and breeding strategies for Andigena potatoes in long-day temperate environments. Theor Appl Genet 66:87–91
Tarn TR, Tai GCC, De Jong H, Murphy AM, Seabrook JEA (1992) Breeding potatoes for long-day temperate climate. In: Janick J (ed.) Plant Breed. Rev. John Wiley & Sons, New York, 9:217-332
Thompson PG, Mendoza HA (1984) Genetic variance estimates in a heterogenous potato population propagated from true seed (TPS). Am Potato J 61:697–702
Thompson PG, Mendoza HA, Plaisted RL (1983) Estimation of genetic parameters for characters related to potato propagation by true seed (TPS) in an andigena population. Am Potato J 60:393–401
Tung PX, Rasco ET, Vander Zaag P, Schmiediche P Jr (1990) Resistance to Pseudomonas solanacearum in the potato: 1. Effects of sources of resistance and adaption. Euphytica 45:203–210
Van der Plank JE (1946) Origin of the first European potatoes and their reaction to length of day. Nature 157:503–515
Veilleux RE, Lauer FI (1981) Breeding behaviour of yield components and hollow heart in tetraploid-diploid vs conventionally derived potato hybrids. Euphytica 30:547–561
von Kameke K (1975) Untersuchungen zur quantitativen Variabilität in Kreuzungsnachkom menschaften der Kartoffel. Hefte Kartoffelbau 19:58
von Kameke K (1978) Untersuchungen zur Erblichkeit einiger Merkmale bei der Kartoffel. Der Kartoffelbau 5:172–173
Wastie RL, Bradshaw JE, Stewart HE (1993) Assessing general combining ability for late blight resistance and tuber characteristics by means of glasshouse seedling tests. Potato Res 36:353–357
Wenzel G, Bapat VA, Uhrig H (1983) New strategy to tackle breeding problems of potato. In: Sen SK, Giles KL (eds) Plant Cell Culture in Crop Improvement. Plenum Press, New York, pp 337–349
Wricke G, Weber WE (1986) Quantitative genetics and selection in plant breeding. Walter de Gruyter, Berlin
Zadina J (1971) Selecting highly productive potato crosses according to performance of the seedlings. Gen A Slechteni 7:269–273
Compliance with Ethical Standard
Conflict of Interest
The author declares that he has no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Gopal, J. Challenges and Way-forward in Selection of Superior Parents, Crosses and Clones in Potato Breeding. Potato Res. 58, 165–188 (2015). https://doi.org/10.1007/s11540-015-9292-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11540-015-9292-6