
Abstract
Utilization of potato (Solanum tuberosum L.) cultivars with greater phosphorus (P) efficiency and response to application of this nutrient is essential to maximize yield, mainly in tropical soils that usually have low P availability. An experiment was conducted under greenhouse conditions in order to evaluate the P uptake rate and the P efficiency for dry matter (DM) production and tuber yield of five potato cultivars (Agata, Asterix, Atlantic, Markies, and Mondial) under low (15 mg dm−3 P) and high (150 mg dm−3 P) P levels. Plants were grown in pots containing 35 dm3 of a Typic Acrortox soil (31% clay, 4% silt, and 65% sand). Mondial and Agata produced high tuber yield under both P levels due to a high P efficiency. Mondial and Asterix were responsive to P applied regarding the tuber DM production and tuber yield. At high P levels, Asterix, which was inefficient and responsive to P application, had similar tuber yield than Agata, which was P efficient and non-responsive. Potato cultivars, such as Markies and Atlantic, which were inefficient and responsive to P application regarding shoot or whole plant DM production, might not produce high tuber yield if the DM partitioning to the tubers is low.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Phosphorus (P) is a macronutrient essential for plant growth and development, and large amounts of P fertilizers are normally needed for crop production. Mineral phosphate rock, as a raw material for P fertilizer production, is non-renewable and will inevitably be depleted within a few decades (EcoSanRes 2008). Due to the continuously increasing consumption of P fertilizers worldwide, a reduction in the sources of phosphate rocks for P fertilizer production is oriented toward scarcity (Abelson 1999). The global production of P fertilizers from high-grade phosphate rock is estimated to be significantly reduced by 2030 (Cordell et al. 2009). Thus, it is critical and imperative to improve the use efficiency of P applied and use less P fertilizers for sustaining crop production.
Potato (Solanum tuberosum L.) is one of the main sources of food in several regions of the world, and its cultivation has been increasing particularly in developing countries and tropical countries (FAOSTAT 2014). However, due to the potato being a crop with low ability to take P up from soils with low available-P levels (Dechassa et al. 2003), high amounts of P fertilizers are normally applied for an adequate potato plant development and high tuber yield (Alvarez-Sánchez et al. 1999; Dechassa et al. 2003; Balemi and Schenk 2009a). In Brazil, as in most tropical countries, one of the main limiting factors to high potato yield is the low P availability in the soil (Manrique 1993; Rocha et al. 1997). Therefore, research on P efficiency is extremely important for sustainability of potato production, especially in soils with low P availability. Use of genotypes with high P efficiency is an option to obtain high yield when plants are grown in soils with low P availability or high fixation capacity of this nutrient, as normally occurs in tropical soils (Balemi 2011; Sanchez and Uheara 1980).
Phosphorus efficiency is defined as the ability of the plants in attaining high yield under limiting P conditions. This can be divided into P uptake efficiency and P use efficiency (Graham 1984; Wang et al. 2010). The P uptake efficiency is the capacity of the plants to take P up from the soil, while the P use efficiency is the ability of the plants to produce biomass or product of economic interest (e.g., grain and tuber) using the P taken up (Wang et al. 2010).
Studies have demonstrated that genotypes with high P uptake efficiency might not present high yield if they are inefficient in utilizing the P taken up. Balemi and Schenk (2009b) and Balemi (2011) verified that inefficient genotypes showed high P accumulation in the shoots even when grown under low P availability in the soil suggesting that the inefficiency of some potato genotypes was more related to the low P use efficiency than the P uptake efficiency. Therefore, higher uptake and accumulation of P in the shoots do not necessarily result in greater P efficiency, especially not in genotypes with low P use efficiency (Wang et al. 2005; Balemi 2011). Some genotypes with high P efficiency are not responsive to P (Balemi and Schenk 2009a), presenting high yield under P-limiting conditions but low response to application of increasing P rates.
Although several studies have been conducted under P-limiting conditions with a purpose of selecting potato genotypes with high P use efficiency, only the shoot dry matter (DM) production, shoot DM rate, and shoot DM production per unit of P taken up were evaluated (Balemi 2009; Balemi and Schenk 2009a; Balemi and Schenk 2009b; Balemi 2011). However, due to the difference in DM partitioning to tubers (Fernandes et al. 2010a) and water content in the tubers (Quadros et al. 2009; Fernandes et al. 2010b) of potato cultivars, a cultivar with high P use efficiency for shoot DM production might not be high yielding.
The cultivars Agata, Asterix, Atlantic, Markies, and Mondial are the cultivars most planted in Brazil, representing approximately 80% of the potato production in the country (ABBA 2010). However, there is no information on differences in P uptake efficiency and P use efficiency among those cultivars. Generally, the same P rate has been recommended for contrasting potato cultivars (Lorenzi et al. 1997; Fontes 1999; Pereira et al. 2005); however, the response to P fertilization might differ among those cultivars, as they differ in the amount of P taken up (Fernandes et al. 2011). Thus, knowledge on P efficiency and response of potato cultivars grown in tropical regions is essential in order to assist the management of P fertilization. This would enable the use of higher P rates in cultivars with higher response to P application. Therefore, the objective of this study was to evaluate the P uptake rate and the P efficiency for DM production and tuber yield of five potato cultivars grown under low and high P supply conditions.
Materials and Methods
A greenhouse experiment was conducted from May to August, 2011 at São Paulo State University in Botucatu, São Paulo, Brazil using 38-L pots, with depth of 26 cm and five holes at the bottom to drain excess water. Pots contained 35 dm3 of a Typic Acrortox soil (31% clay, 4% silt, and 65% sand). A soil sample was air-dried and analyzed to determine the chemical characteristics (van Raij et al. 2001) (Table 1).
The experimental design was a randomized complete block design in a 2 × 5 factorial scheme, with four replications. The treatments consisted of two P levels in the soil, 15 mg dm−3 (low P) and 150 mg dm−3 (high P), and five potato cultivars, Agata, Asterix, Atlantic, Markies, and Mondial. According to Lorenzi et al. (1997), P concentration (resin extractable) in the soil <25 mg dm−3 is considered low and >60 mg dm−3 is considered high for the potato crop. Each pot with a single plant represented one experimental unit (i.e., one replication of one treatment).
Soil was supplied with dolomitic lime in order to reach 60% base saturation (Lorenzi et al. 1997). Then, the soil was wetted to 80% of maximum water holding capacity, covered with polyethylene film, and incubated for 30 days at 25 °C. Thereafter, P levels (15 and 150 mg dm−3 P) were applied using triple superphosphate (41% P2O5) as source. Along with P fertilization, 150 mg dm−3 K (potassium chloride, 60% K2O), 30 mg dm−3 N (ammonium sulphate, 20% N), 5 mg dm−3 Zn, 1 mg dm−3 B, 0.4 mg dm−3 Cu, 1.7 mg dm−3 Fe, 1.1 mg dm−3 Mn, and 0.06 mg dm−3 Mo (fritted trace elements) were added. Soil samples were collected 10 days after application of fertilizers, air-dried, and analyzed for chemical characteristics (Table 1).
Before planting, carbofuran (2,3-dihydro-2,2-dimethyl-7-benzofuranol methylcarbamate) insecticide was applied in the soil at 2.5 mg a.i. dm−3. Seed tubers (diameter size between 30 and 50 mm) were soaked for 5 min separately in solutions with fungicides and bactericide. Each solution had 3.75 g L−1 mancozeb ([[1,2-ethanediylbis[carbamodithioato]](2-)]manganese with [[1,2-ethanediylbis[carbamodithioato]](2-)]zinc), 2.5 g L−1 pencycuron (N-[(4-chlorophenyl)methyl]-N-cyclopentyl-N'-phenylurea), and 0.02 g L−1 kasugamycin (3-O-[2-amino-4-[(carboxyiminomethyl)amino]-2,3,4,6-tetradeoxy-a-D-arabino hexopyranosyl]-D-chiro-inositol). Potato cultivars were planted on May 27, 2011, at a 15-cm depth, using one seed tuber with vigorous sprouts per pot. Emergence occurred 12 days after planting (DAP). Hilling was performed in the beginning of the stolonization stage at 20 days after emergence (DAE). Nitrogen side-dressing fertilization was applied in two split times, 15 and 39 DAE, using 40 mg dm−3 N (ammonium sulphate, 20% N) in each application.
Soil matric potential was monitored with conventional mercury tensiometers (13-mm diameter, with a ceramic porous cup connected with tubing to a mercury manometer), which were constructed according to Richards (1941) and installed at the planting date at 12-cm depth in all pots. Irrigation was performed manually when the mean matric potential in the soil of each treatment reached −0.02 MPa, according to recommendations by Oliveira and Valadão (1997), and calculated so as to increase tension values up to field capacity.
The greenhouse was maintained at 27/14 °C (day/night temperature) and 12-h photoperiod. The photosynthetically active radiation followed a typical diurnal pattern with the maximum of 1500 μmol m−2 s−1 at approximately 12:00 h. Diseases and insects were controlled with preventive pesticide sprays.
Harvest of the plants was performed at 70 DAE, when approximately 75% of the plants showed haulm yellowing. The plants were washed in deionized water and separated in roots, shoots, and tubers. The tubers were weighted for tuber yield (fresh tubers) determination. All plant parts were dried in an oven with forced air circulation at 65 °C for 96 h. Then, the plant material was weighted for DM production determination. The whole plant DM was calculated by adding the values obtained for roots, shoots, and tubers. Dry matter partitioning among the plant parts was calculated as a percentage of DM accumulated in the roots, shoots, and tubers in relation to the whole plant DM.
After drying, the roots, shoots, and tubers were ground in a Wiley grinder with a 1-mm sieve, and the samples were analyzed for P concentration, according to the methodology described by Malavolta et al. (1997). The amount of P accumulated in the roots, shoots, and tubers was estimated by multiplying the nutrient concentration by the amount of DM accumulated. Phosphorus uptake rate was calculated according to the equation described by Swiader et al. (1994):
Data were subjected to analysis of variance and means were separated using Tukey’s test at the 0.05 probability level. Correlation coefficients were calculated across all cultivars and two levels of P using STATISTICA, version 6 (Statsoft, 1995), to determine the relationships between the parameters.
Methodology by Gerloff (1977) and Fageria and Kluthcouski (1980) that suggests the classification of cultivars according to the use efficiency and response to application of P (efficiency and response, ER) was utilized for differentiation of the cultivars. The cultivars that presented higher DM production or tuber yield than the average of all cultivars under low P level (15 mg dm−3) were considered efficient. The response to P application was obtained according to the equation:
Response index to P applied (RIPA) = [(DM or tuber yield (g) at high P level − DM or tuber yield (g) at low P level) / difference between P levels (mg dm−3)].
A graphical representation in the Cartesian plane was used to classify the cultivars. The DM production or tuber yield from different cultivars at the low P levels was presented in the x-axis and the response to P applied was presented in the y-axis of the Cartesian coordinates system. The origin point of the x-axis was presented by average of DM production or tuber yield at the low P level of all cultivars, while the origin point of the y-axis was presented by average of the response to P applied of all cultivars. The diagram was divided into four quadrants separating four cultivar groups. At the first quadrant, cultivars efficient and responsive; at the second, inefficient and responsive; at the third, inefficient and non-responsive; and at the fourth, efficient and non-responsive are represented.
Results and Discussion
Dry matter of roots and tubers were influenced by cultivar and P level (Table 2). Under high P level, DM of root and tuber was 16 and 60%, respectively, greater than the low P level, indicating that P, apart from promoting further development of the root system, is essential for growth of potato tubers, as was also observed by Alvarez-Sánchez et al. (2001). The greatest root and tuber DM production was obtained in the cultivar Mondial.
Dry matter production of shoot and whole plant was affected by cultivar × P level interaction (Table 2). Shoot DM in Agata, Asterix, and Atlantic was similar under both P levels. However, shoot DM in Mondial under low P level was 28% higher than that under high P level, while shoot DM in Markies under low P was lower than that under high P level. Under low P level, Mondial showed the highest shoot DM compared with the other cultivars (Table 2). Thus, the cultivars Agata, Asterix, Atlantic, and specially Mondial were able to maintain shoot growth, even under P-limiting conditions, due to an increase in allocation of DM to the shoot, which did not occur in Markies (Fig. 1). However, under high P level, Mondial showed shoot DM approximately 29% lower than the cultivars Agata, Asterix, and Markies.

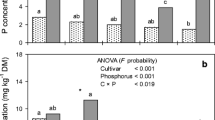
Effect of P supply on dry matter partitioning of potato cultivars. Different lowercase letters indicate significant difference between P levels at the same cultivar, whereas different uppercase letters indicate significant difference between cultivars at the same P level, at P ≤ 0.05, according to Tukey’s test
Whole plant DM of all cultivars was 29% lower under low P level than high P level, demonstrating that the growth of all cultivars was limited under low P supply (Table 2). Balemi (2009) observed reduction from 35 to 75% in DM production of potato genotypes grown under low P availability. Mondial and Agata showed higher whole plant DM production than the other cultivars under low P level. However, under high P level, Mondial, Agata, and Asterix had higher whole plant DM. Therefore, Mondial and Agata seem to have greater capacity of growing in P-deficient soils, possibly due to the ability to maintain cell growth and division, even with lower P concentrations in the plant tissues (Table 3), as observed in other crops (Lynch et al. 1991; Chiera et al. 2002). Nevertheless, Asterix requires high P availability to maintain optimal growth with greater DM production (Table 2).
Tuber yield was affected by cultivar × P level interaction (Table 2). Tuber yield of all cultivars under low P level was on average 32.4% lower than under high P level, indicating that significant reductions in tuber yield occur when potato plants are grown under P-limiting conditions (Alvarez-Sánchez et al. 1999; Dechassa et al. 2003; Fleisher et al. 2012). In the two P levels, the cultivars Mondial and Agata showed the greatest tuber yield compared with the other cultivars (Table 2). The higher productive ability of these cultivars, even under P-limiting conditions, indicates higher P efficiency (Graham, 1984).
Dry matter partitioning to different plant parts was significantly influenced by cultivar × P level interaction (P < 0.001). Dry matter allocation to the roots of all cultivars was higher under low P level than under high P level (Fig. 1), which was also observed by Balemi and Schenk (2009b) and Balemi (2009). When P availability is low, the roots become the preferential sink of photoassimilates in order to improve P uptake efficiency (Horst et al. 1993).
The DM allocation to the shoots of Agata, Asterix, Atlantic, and Mondial was higher under low P, which maintained and/or increased shoot DM production under low P level (Fig. 1 and Table 2). The DM allocation to the shoot in Markies remained unchanged by the P levels; however, the shoot DM production was lower under low P level compared with the high P level (Fig. 1 and Table 2).
The cultivars Asterix, Atlantic, and Mondial showed lower DM allocation to the tubers under low P, while the DM allocation to the tubers in Agata and Markies was not altered by P supply (Fig. 1). Under high P level, Mondial showed greater DM allocation to the tubers and lower DM allocation to the shoots compared with the other cultivars. In all cultivars and P levels studied, the tubers were the main sink of carbohydrates of the plants, representing from 54.8 to 77.2% of the total plant DM (Fig. 1), which is in accordance with Fernandes et al. (2010a). Under both P levels, Atlantic showed higher DM allocation to the roots than the other cultivars, although it did not differ significantly from Asterix under low P level (Fig. 1).
Phosphorus concentration in the roots and shoots was influenced by cultivar × P level interaction (Table 3). All cultivars had higher P concentrations in roots under high P level compared with those under low P level. These results were also observed by Fernandes and Soratto (2012) on potato plants grown in nutrient solution. Under low P level, Atlantic had the greatest P concentration in the roots, while under high P level, the P concentration in the roots was similar in all cultivars.
Phosphorus concentration in the shoots of all cultivars was on average 44% higher under high P level than under low P level (Table 3). Other authors also observed increase in P concentration in the leaves (Balemi 2009; Balemi and Schenk 2009b) and the shoots (Fernandes and Soratto 2012; Balemi 2011) of potato plants grown under high P availability. Under low P level, P concentration in the shoot of Atlantic was higher than that of the other cultivars (Table 3). The high P level promoted the highest P concentration in the shoots of Markies, even though it did not differ significantly from the concentrations in Asterix and Atlantic.
Phosphorus concentration in the tubers was affected only by the P level, with an increase of 46% on average in plants grown under high P level compared with that under low P level (Table 3).
Phosphorus accumulation in the roots, shoots, tubers, and whole plant was affected by cultivar × P level interaction (Table 4). Amounts of P accumulated in the roots, shoots, tubers, and whole plant of all cultivars under high P level were 1.8-, 1.6-, 2.4-, and 2.2-fold, respectively, greater than under low P level, due to the greater plant DM production and P concentration in their tissues (Tables 2, 3, and 4).
Under low P level, roots of Atlantic accumulated more P than roots of the cultivars Asterix, Markies, and Mondial. However, the P accumulated in the roots did not promote root growth of Atlantic (Tables 2, 3, and 4), although under low P availability, retention of P in the roots usually increases in order to maintain the growth of the root system (Marschner 1995). Under low P level, P accumulation in the shoots, tubers, and whole plant was similar in all cultivars, regardless of differences in DM production and P concentration in the shoots (Tables 2, 3, and 4). Under high P level, Markies had lower P accumulation in the roots than the other cultivars; however, the P accumulation in the shoots was greater in Asterix and Markies than in the other cultivars (Table 4).
Under high P level, Mondial accumulated more P in the tubers and whole plant than Agata, Atlantic, and Markies, especially due to the greater DM accumulation in the tubers and the whole plant of Mondial (Tables 2 and 4). This indicates that under high P level, cultivars with greater P uptake could be more productive if the differences in P efficiency are not substantial, because in this condition, the P accumulation in the whole plant has a positive and significant correlation with the whole plant DM production and tuber yield (Table 5). Other authors also observed a positive correlation between total P uptake and DM accumulation in crops such as wheat (Ozturk et al. 2005), rice (Fageria et al. 1988), and potato (Balemi 2011).
Phosphorus uptake rate was influenced by cultivar × P level interaction (Table 4). In all cultivars, P uptake rate was greater under high P level, which increased concentration and accumulation of P in all plant organs of the potato cultivars (Tables 3 and 4). Fernandes and Soratto (2012) also observed an increase in P uptake rate per unit root mass of potato when the P concentration in the nutrient solution was increased.
There was no significant difference in P uptake rate among cultivars under low P level (Table 4), as under low P level, the transport of P to the roots is the main limiting factor to P uptake (Dechassa et al. 2003). The cultivars Asterix, Atlantic, and Markies showed P uptake rate and P accumulation in the whole plant similar to the other cultivars under low P level (Table 4); however, they produced less DM in the whole plant and fresh tubers (Table 2), demonstrating that higher plant growth and tuber yield also depend on the cultivar’s ability to use the taken up P for production of DM or tubers (Wang et al. 2005; Balemi 2011).
Under high P level, the highest P uptake was found in Mondial, and this cultivar also accumulated higher amount of P in the whole plant than Atlantic and Markies, even though Mondial showed root DM similar to the other cultivars (Tables 2 and 4). These results indicate that Mondial seems to be more efficient in P uptake from the soil under high P level, due to its different morphological and physiological characteristics of the roots from other cultivars. Under conditions of low P level in the soil, genotypes with greater yield or biomass production are considered as P efficient in comparison with other genotypes without these characteristics (Gerloff 1977; Fageria and Kluthcouski 1980). Under low P level, only Agata and Mondial were efficient, since they had DM production of shoot, tuber, and whole plant, and tuber yield greater than the average of all cultivars (38.9, 80.5, 129.0, and 475 g plant−1 for DM of shoot, tuber, whole plant, and tuber yield, respectively) (Fig. 2 and Table 2). Greater P efficiency of these cultivars might be related to the capacity to maintain cellular division and growth under low P concentration in the tissues (P use efficient) (Lynch et al. 1991; Chiera et al. 2002), as demonstrated by the lower P concentration in plant tissues (Table 3). Considering the shoot DM production, Asterix was also efficient due to its high DM partitioning to the shoot under low P level (Figs. 1 and 2a). However, tuber DM and tuber yield in Asterix were low (Table 2), which classifies this cultivar as inefficient, considering the tuber DM and tuber yield (Fig. 2b, d). As the tubers represented more than 70% of the whole plant DM at the end of the cycle (Fernandes et al. 2010a), the results of this study indicate that only tubers (tuber DM or tuber yield) should be taken into account for proper evaluation of P efficiency.

Classification of potato cultivars for P efficiency and responsiveness based on shoot dry matter (a), tuber dry matter (b), whole plant dry matter (c), and tuber yield (d). DM, dry matter, RIPA, response index to P applied
Classification of potato genotypes regarding P efficiency by evaluating shoot DM production and rate of shoot DM production under limiting P supply, as well as shoot DM production per unit of P taken up has been performed by several authors (Balemi 2009; Balemi and Schenk 2009a; Balemi and Schenk 2009b; Balemi 2011). However, the results of this study indicate that potato cultivars that are P efficient for shoot DM production (Fig. 2a) do not necessarily show high tuber yield (Table 2 and Fig. 2d), when the cultivars have differences in DM partitioning to tubers (Fernandes et al. 2010a) or in water content in the tubers (Quadros et al. 2009; Fernandes et al. 2010b).
Under low P level, Asterix, Atlantic, and Markies were less productive than other cultivars (Table 2). Since the P uptake rate of these cultivars did not differ from that of the other cultivars (Table 4), the lower productivities were due to the lower use efficiency of P taken up for tuber production (Fig. 2b, d). This demonstrates that the greater plant growth and higher tuber yield also depend on the cultivar’s ability to use the taken up P for production of DM or tubers (Wang et al. 2005; Balemi and Schenk 2009a; Balemi 2011).
The response index to P applied, evaluated by the tuber DM production and tuber yield, indicated that only the cultivars Asterix and Mondial were responsive, as these cultivars had the tuber DM production and tuber yield from 9 to 49% higher than the average of all cultivars (Table 2 and Fig. 2b, d). However, Asterix was inefficient in the use of P for tuber DM production and tuber yield. On the other hand, Agata was efficient and non-responsive because this cultivar showed the highest tuber yield under low P level and low increases in tuber yield under high P level in the soil.
The cultivars Markies and Atlantic were not responsive to P application for tuber DM production and tuber yield, but both cultivars, mainly Markies, showed high response to P for DM production of the shoots and the whole plant (Fig. 2). This is due to the increase in shoot growth and DM allocation to the shoots of Markies grown under high P level (Table 2 and Fig. 1). The results suggest that the classification of potato cultivars regarding the P efficiency and the response to application of this nutrient, with DM production of the shoot or the whole plant as variables (Balemi 2009; Balemi and Schenk 2009a; Balemi and Schenk 2009b; Balemi 2011), might not be an adequate parameter as they differ in DM partitioning to the tubers (Fernandes et al. 2010a), which are the plant organ of commercial interest.
Conclusion
Mondial and Agata showed high tuber yield under both P levels due to a high P efficiency. Mondial and Asterix were responsive to P applied regarding the tuber DM production and tuber yield. At high P levels, Asterix, which was inefficient and responsive to P application, had a tuber yield similar to the one of Agata, which was P efficient and non-responsive. Potato cultivars, such as Markies and Atlantic, which were inefficient and responsive to P application regarding shoot or whole plant DM production, might not produce high tuber yield if the DM partitioning to the tubers is low.
References
ABBA - Associação Brasileira da Batata (Brazilian Association of Potato) (2010) Variedades (Varieties). Rev Batata Show 10:1–4
Abelson PH (1999) A potential phosphate crisis. Science 283:2015
Alvarez-Sánchez E, Etchevers JD, Oritz J, Núñez R, Volke V, Tijerina L, Martínez A (1999) Biomass production and phosphorus accumulation of potato as affected by phosphorus nutrition. J Plant Nutr 22:205–217
Alvarez-Sánchez E, Etchevers JD, Ortiz J, Núñez R, Martínez AG, Castellanos JZ (2001) Phosphorus nutrition of potato and maize seedlings. Terra 19:55–65
Balemi T (2009) Effect of phosphorus nutrition on growth of potato genotypes with contrasting phosphorus efficiency. Afr Crop Sci J 17:199–212
Balemi T (2011) Screening for genotypic variation in potato for phosphorus efficiency. Int Res J Plant Sci 2:233–243
Balemi T, Schenk MK (2009a) Genotypic variation of potato for phosphorus efficiency and quantification of phosphorus uptake with respect to root characteristics. J Plant Nutr Soil Sci 172:669–677
Balemi T, Schenk MK (2009b) Genotypic difference of potato in carbon budgeting as a mechanism of phosphorus utilization efficiency. Plant Soil 322:91–99
Chiera J, Thomas J, Rufty T (2002) Leaf initiation and development in soybean under phosphorus stress. J Exp Bot 53:473–481
Cordell D, Drangert JO, White S (2009) The story of phosphorus: global food security and food for thought. Glob Environ Chang 19:292–305
Dechassa N, Schenk MK, Claassen N, Steingrobe B (2003) Phosphorus efficiency of cabbage (Brassica oleraceae L. var. capitata), carrot (Daucus carota L.), and potato (Solanum tuberosum L.). Plant Soil 250:215–224
EcoSanRes (2008) Closing the loop on phosphorus. Stockholm Environment Institute (SEI) funded by SIDA Stockholm. 2p. http://www.ecosanres.org/pdf_files/ESR-factsheet-04.pdf
Fageria ND, Kluthcouski J (1980) Metodologia para avaliação de cultivares de arroz e feijão para condições adversas de solo (Methodology for evaluation of rice and bean cultivars to adverse soil conditions). Embrapa/CNPAF, Brasília
Fageria NK, Morais OP, Baligar VC, Wright RJ (1988) Response of rice cultivars to phosphorus supply on an oxisol. Fert Res 16:195–206
FAOSTAT (2014) Production: Potatoes. Available at http://faostat.fao.org/site/567/default.aspx#ancor (accessed 20 Jan. 2014). Food and Agriculture Organization of the United Nations, Rome, Italy
Fernandes AM, Soratto RP (2012) Nutrition, dry matter accumulation and partitioning, and phosphorus use efficiency of potato plants grown under phosphorus rates in nutrient solution. Rev Bras Ciênc Solo 36:1528–1537
Fernandes AM, Soratto RP, Evangelista RM, Nardin I (2010a) Qualidade físico-química e de fritura de tubérculos de cultivares de batata na safra de inverno (Physicochemical and frying quality of potato cultivars in winter season). Hort Bras 28:299–304
Fernandes AM, Soratto RP, Silva BL, Souza-Schlick GD (2010b) Crescimento, acúmulo e distribuição de matéria seca em cultivares de batata na safra de inverno (Growth and dry matter accumulation and distribution in potato cultivars during the winter crop season). Pesq Agropec Bras 45:826–835
Fernandes AM, Soratto RP, Silva BL (2011) Extração e exportação de nutrientes em cultivares de batata: I - macronutrientes (Nutrient extraction and exportation by potato cultivars: I – macronutrients). Rev Bras Ciênc Solo 35:2039–2056
Fleisher DH, Wang Q, Timlin DJ, Chun J-A, Reddy VR (2012) Response of potato gas exchange and productivity to phosphorus deficiency and carbon dioxide enrichment. Crop Sci 52:1803–1815
Fontes PCR (1999) Batata (Potato). In: Ribeiro AC, Guimarães PTG, Alvarez V VH (ed) Recomendações para o uso de corretivos e fertilizantes em minas gerais - 5ª Aproximação (Recommendations for the use of soil amendments and fertilizers in the State of Minas Gerais - 5th Approach). CFSEMG, Viçosa, pp 179
Gerloff S (1977) Plant efficiencies in the use of N, P, and K. In: Wrigh M (ed) Plant adaptation to mineral stress in problem soils. Cornell Univ Press, New York, pp 161–174
Graham RD (1984) Breeding for nutritional characteristics in cereals. Adv Plant Nutr 1:57–102
Horst WJ, Abdou M, Wiesler F (1993) Genotypic differences in phosphorus efficiency of wheat. Plant Soil 155(156):293–296
Lorenzi JO, Miranda Filho HS, van Raij B (1997) Raízes e tubérculos (Roots and tubers). In: van Raij B, Cantarella H, Quaggio JA, Furlani AMC (ed) Recomendações de adubação e calagem para o Estado de São Paulo (Lime and fertilizer recommendations for the State of São Paulo). 2nd ed. Tech. Bull. 100. Inst Agronômico Campinas, Campinas pp 221–229
Lynch J, Läuchli A, Epstein E (1991) Vegetative growth of common bean in response to phosphorus nutrition. Crop Sci 31:380–387
Malavolta E, Vitti GC, Oliveira SA (1997) Avaliação do estado nutricional das plantas: princípios e aplicações (Evaluation of nutritional status of plants: principles and applications), 2nd edn. Potafos, Piracicaba
Manrique LA (1993) Constrains for potato productions in tropics. J Plant Nutr 16:2075–2120
Marschner H (1995) Mineral nutrition of greater plants, 2nd edn. Academic Press, London
Oliveira CAS, Valadão LT (1997) Manejo da água do solo no cultivo da batata (Soil water management in the potato cultivation). Tech Commun 3, Embrapa Hortaliças, Brasília
Ozturk L, Eker S, Torun B, Cakmak I (2005) Variation in phosphorus efficiency among 73 bread and durum wheat genotypes grown in a phosphorus-deficient calcareous soil. Plant Soil 269:69–80
Pereira AS, Daniels J, Freire CJS, Bertoncini O, Nazareno NRX, Brisolla AD, Salles LAB, Madail JCM (2005) Produção de Batata no Rio Grande do Sul (Potato production in the State of Rio Grande do Sul). Tech Cir. 48. Embrapa Clima Temperado, Pelotas
Quadros DA, Iung MC, Ferreira SMR, Freitas RJS (2009) Composição química de tubérculos de batata para processamento, cultivados sob diferentes doses e fontes de potássio (Chemical composition of potato tubers for processing, grown in different levels and sources of potassium). Ciênc Tecn Alim 29:316–323
Richards LA (1941) Soil moisture tensiometer materials and construction. USDA-ARS, Washington, DC. http://afrsweb.usda.gov/sp2userfiles/place/53102000/pdf_pubs/p0015.pdf (accessed 5 Mar. 2009)
Rocha FAT, Fontes PCR, Fontes RLF, Reis FP (1997) Critical phosphorus concentrations in potato plant parts at two growth stages. J Plant Nutr 20:573–579
Sanchez PA, Uheara G (1980) Management considerations for acid soils with high phoshphorus fixation capacity. In: Khasawneh, FE, Sample EC, Kamprath, EJ (ed) The role of phosphorus in agriculture. ASA, CSSA, and SSSA, Madison, pp 471–514
Statsoft (1995) STATISTICA Release 5. StatSoft, Tulsa
Swiader JM, Chyan Y, Freiji FG (1994) Genotypic differences in nitrate uptake and utilization efficiency in Pumpkin hybrids. J Plant Nutr 17:1687–1699
van Raij B, Andrade JC, Cantarella H, Quaggio JA (2001) Análise química para avaliação da fertilidade de solos tropicais (Chemical analysis for fertility evaluation of tropical soils). Campinas, Inst Agronômico
Wang QR, Li JY, Li ZS, Chritie P (2005) Screening chinese wheat germplasm for phosphorus efficiency in calcareous soils. J Plant Nutr 28:489–505
Wang X, Shen J, Liao H (2010) Acquisition or utilization, which is more critical for enhancing phosphorus efficiency in modern crops? Plant Sci 179:302–306
Acknowledgments
Acknowledgments are to São Paulo Research Foundation (FAPESP, proc. 2010/04987-6) for supporting this research, and to the National Council for Scientific and Technological Development (CNPq) for providing research grant to the first author.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Soratto, R.P., Pilon, C., Fernandes, A.M. et al. Phosphorus Uptake, Use Efficiency, and Response of Potato Cultivars to Phosphorus Levels. Potato Res. 58, 121–134 (2015). https://doi.org/10.1007/s11540-015-9290-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11540-015-9290-8