
Abstract
Recent studies have shown that multiple sclerosis (MS) and schizophrenia share similarities in some respects, including white matter damage and neuroinflammation. On the other hand, adenosine was reported to promote oligodendrocyte precursor maturation and remyelinating while influencing microglia activation. The aim of the present study was to examine possible beneficial effects of adenosine on the recovery of cuprizone (CPZ)-exposed mouse which has been used as an animal model of MS and schizophrenia as the CPZ-exposed mouse presents demyelination, oligodendrocyte loss, microglia accumulation, as well as behavioral changes. As reported previously, C57BL/6 mice, after fed CPZ for 5 weeks, showed salient demyelination and oligodendrocyte loss in the cerebral cortex (CTX) and hippocampus, in addition to displaying anxiety-like behavior, spatial working memory deficit, and social interaction impairment. Administration of adenosine for 7 days during the recovery period after CPZ withdrawal promoted the behavioral recovery of CPZ-exposed mice and accelerated the remyelinating process in the brains of mice after CPZ withdrawal in a dose-dependent manner. In addition, the effective dose (10 mg/kg) of adenosine inhibited microglia activation and suppressed abnormal elevation of the pro-inflammatory cytokines IL-1β and TNF-α in CTX and hippocampus, but increased levels of the anti-inflammatory cytokines IL-4 or IL-10 in the same brain regions during the remyelinating process. These results provided an evidence-based rationale for the application of adenosine or its analogues as add-on therapy for schizophrenia.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Cuprizone (CPZ) is a copper chelator and has been shown to selectively induce oligodendrocyte loss and damage to the myelin sheath in the central nervous system (CNS) of rodents (Matsushima and Morell 2001; Yang et al. 2009). The CPZ-exposed mouse has been used as an animal model of multiple sclerosis (MS), a chronic inflammatory demyelinating disease of human CNS, with focal lesions in the gray and white matter and diffuse neurodegeneration in the entire brain (Kipp et al. 2017; Lassmann 2018; Remington et al. 2007; Selvaraju et al. 2004). In most of cases, MS starts as a relapsing remitting disease following an attack against the CNS which leads to focal inflammatory lesions and neurological symptoms, i.e., MS relapses (Nylander and Hafler 2012). Within the following 10–15 years, more than 60% of relapsing MS patients proceed to develop secondary progressive MS leading to relentless disease progression and accumulation of disability (Tutuncu et al. 2013). It was demonstrated that both resident microglia and blood-derived macrophages, via release of pro-inflammatory cytokines and reactive oxygen species (ROS), lead to oxidative injury of mitochondria, oligodendrocyte damage and degeneration of neurons (Fischer et al. 2013; Gandhi et al. 2010; Ransohoff et al. 2015).
Interestingly, the CPZ-exposed mouse has been also established as a new animal model of schizophrenia, a devastating and complex CNS disorder, which involves multiple pathological components such as neuroinflammation (Monji et al. 2009), mitochondrial dysfunction (Ben-Shachar 2002), and oligodendroglia dysfunction (Davis et al. 2003), in addition to dysfunctions of the dopaminergic system (van Rossum 1967) and glutamatergic system (Kim et al. 1980). First, the CPZ-exposed mouse showed behavioral changes in multiple domains of high-order brain functions including the increased CNS activity (Chang et al. 2017; Xu et al. 2009), the reduced anxiety response to a novelty challenge test (Franco-Pons et al. 2007), the impaired prepulse inhibition, the spatial working memory deficit, as well as the social interaction decrease (Tezuka et al. 2013; Xiao et al. 2008; Xu et al. 2009; Zhang et al. 2016). These behavioral changes are reminiscent of some symptoms seen in schizophrenia patients, including those pertaining to mood, affect, and cognitive impairments (Xu and Li 2011), thus demonstrated a face validity of the CPZ-exposed mouse as an animal model of schizophrenia. Secondly, the CPZ-induced behavioral and morphological changes were alleviated by some of the extant antipsychotics such as olanzapine and quetiapine (Chandran et al. 2012; Xiao et al. 2008; Xu et al. 2010, 2011; Zhang et al. 2012), suggesting a predictive validity of this animal model. More interestingly, the CPZ-exposed mice also showed dopaminergic dysfunction (Chang et al. 2017; Xu et al. 2009); oxidative stress (Xuan et al. 2015), neuroinflammation (Shao et al. 2015; Tezuka et al. 2013), and mitochondrial dysfunction (Faizi et al. 2016; Xuan et al. 2014, 2015; Zhang et al. 2018) in the brain, showing the construct validity of the CPZ-exposed mouse as a new animal model in schizophrenia research.
An inference from the above literature review is that MS and schizophrenia share similarities in some respects. In support of this inference, large-scale hospital-based, epidemiological and case studies have suggested a relationship between psychosis and MS through demonstrating their higher than chance co-occurrence, their temporal relationship, and their association with particular structural abnormalities in the brain (Kosmidis et al. 2010), as evidenced by most of magnetic resonance imaging (MRI) studies in schizophrenia suggesting the involvement of white matter pathology in multiple cerebral regions in the neurobiology of this condition (Parnanzone et al. 2017); levels of pro-inflammatory cytokines have been found to be relatively high in schizophrenia patients. And a low-level neuroinflammatory process is accompanied with microglial activation demonstrated in neuroimaging studies (Müller 2018; Müller et al. 2015). Furthermore, some antipsychotics showed anti-inflammatory and immunomodulatory effects in animal and human studies (Shao et al. 2015; Zhang et al. 2018), while anti-inflammatory medications have been used as add-on drugs in the treatment of antipsychotic-resistant patients (Xu 2015).
Adenosine is a nucleoside widely distributed in the organism with neuromodulative and neuroprotective activity in CNS. It acts through four types of membrane receptors named A1, A2a, A2b and A3 (Fredholm et al. 2001). High levels of adenosine inside the CNS lesions are supposed to be the consequence of its increased release and reduced removal and seem to be protective against neural damage (Laghi et al. 2000; Lynch et al. 1998; Robertson et al. 2001). For example, adenosine was shown to act as a potent neuron-glial transmitter, inhibit oligodendrocyte progenitor cells (OPC) proliferation, stimulate differentiation, and promote the formation of myelin (Stevens et al. 2002). Similarly, adenosine A1 receptor agonist, N6-cyclohexyladenosine (CHA), protected myelin and induced remyelinating in an experimental model of rat optic chiasm demyelination (Asghari et al. 2013). Relevant to the protective effects of CHA on the rat demyelination, studies have demonstrated that A1 and A2a receptors modulate neuroinflammation in experimental autoimmune encephalomyelitis (EAE) and MS (Du and Xie 2012; Mills et al. 2012). A2b and A3 receptors, though not well characterized, have also been suggested to interact with inflammatory mediators, such as interleukin-6 (IL-6) (Janes et al. 2015; Merighi et al. 2017).
The aim of the present study was to examine possible beneficial effects of adenosine on the recovery of mice after withdrawing from CPZ exposure which has been shown to cause demyelination, oligodendrocyte loss, microglia and astrocyte accumulation, as well as behavioral changes as reviewed above. Based on the previous studies showing protective effects of adenosine on demyelination and the modulation of adenosine receptors on inflammation as summarized before, we hypothesized that adenosine would promote the recovery of mice from the CPZ-induced white matter damage and behavioral changes via modulating neuroinflammatory components in the CNS lesions.
Materials and Methods
Animals and Drugs
All animal procedures applied in this study were in accordance with the guidelines set up by the Animal Care and Use Committee of Shantou University Medical College and approved by this committee.
Young (5-weeks old) male C57BL/6 mice were purchased from the Experimental Animal Center of Sun Yat-sen University (Guangzhou, China). The male and young C57BL/6 mouse is most sensitive to the neurotoxicity of CPZ as evidenced by oligodendrocyte loss and demyelination in the brain thus has been applied in most of previous animal studies (Matsushima and Morell 2001; Praet et al. 2014). A 7-days acclimatization to the new environment was allowed for the mice before subjected to the experimental procedures. During the acclimatization period, they were housed in groups (5 mice/cage of 29 cm × 17.8 cm × 16 cm) at the standard laboratory conditions of a 12 h light-dark cycle, the constant room temperature of 23.0 ± 1 °C, and the relative humidity of 50–60%, with free accesses to food and water. The bedding of cage was changed every other day.
Both CPZ and adenosine were purchased from Sigma-Aldrich (St. Louis, MO). CPZ was mixed into a standard powdered rodent chow at the final concentration of 0.2% (w/w). Adenosine was freshly prepared (dissolved in sterile saline) and administered intraperitoneally (i.p; 1 ml/100 g body weight) to mice once a day at various doses.
Experimental Design
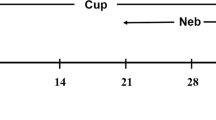
The experiment consisted of the following six animal groups (n = 22–24/group): The normal Control group (Control), in which mice were given the standard rodent diet for 5 weeks; the CPZ intoxication group (CPZ), in which mice were fed the CPZ-containing (0.2% w/w) diet for 5 weeks; the vehicle control group (CPZ-VEH), in which mice were fed the CPZ-containing diet for 5 weeks, then returned to normal diet for 1 week during which period they were given (i.p.) sterile saline once a day; CPZ-AD1, CPZ-AD5, and CPZ-AD10 groups. In these CPZ-AD(x) groups mice consumed the CPZ-containing rodent chow for 5 weeks, then returned to the normal rodent diet without CPZ for 7 days during which period mice were given (i.p.) adenosine at three different doses (1.0, 5.0, and 10.0 mg/kg, dissolved in sterile saline) once a day.
For the CPZ-VEH, CPZ-AD1, CPZ-AD5, and CPZ-AD10 groups, 1 day after the last adenosine or saline injection, mice were subjected to the open-field test and social interaction test with an interval of 4–6 h between, followed by the Y-maze test next day. Mice in the Control and CPZ groups were subjected to the same behavioral tests on the same time points after CPZ withdrawal.
Mice were euthanized with sodium pentobarbital (80 mg/kg, i.p.) after finishing the last behavioral test. The brain tissue of mice was used for immunohistochemical, immunoflurescence, Western-blot, and/or ELISA analysis (described later).
Behavioral Tests
The open-field test was performed to measure the locomotor activity, exploratory behavior and anxiety-related behavior of mice. The open box was 25 cm (L) × 25 cm (W) × 30 cm (H), and evenly illuminated at 15 lx. The travel paths of the animal were recorded by a video camera placed above the arena. The distances travelled on the whole arena and the central zone (12.5 cm × 12.5 cm), as well as the time spent in the central zone were analyzed by the video tracking system (DigBehav System, Yishu Co Ltd., Shanghai, China). The open field was cleaned with 70% alcohol after each trial.
The social interaction test procedure was described previously (Berton et al. 2006). The test was carried out in the same open-field arena used for open-field test. A wire-mesh cage (6 cm × 8 cm × 8 cm) was positioned at one of the corners. Each mouse was introduced into the open field and its trajectory was video-tracked for two 2.5-min sessions which were separated by a 1-min interval. In the first session, the open field contained an empty wire-mesh cage. In the second session, the conditions were identical except that a social target (unfamiliar C57Bl/6 mouse) had been placed into the wire-mesh cage. Between the two sessions, the experimental mouse was removed from the arena, and was placed back into its home cage for approximately one min. The video-tracking data resulted from the first and second sessions were used to determine the time spent by the experimental mouse in the “interaction zone” (a 5-cm-wide corridor surrounding the cage) and referred to as “E” and “A”, respectively.
The spatial working memory of mice was assessed by Y-maze test. A Y-maze with three arms (30 cm long 8 cm wide) surrounded by white Plexiglas walls (15 cm high) was used. Individual mouse was placed at the center to which three arms joint and allowed to explore all the arms freely for an 8-min testing period. The series of entries and the total number of entries were recorded. An actual entry was defined as the animal placing all four paws in an arm. A correct alternation was defined as visits into all three arms on consecutive occasions. The theoretical maximal number of correct alternations was the total number of arm entries minus 2. The spontaneous alternation was calculated as the number of correct alternations divided by the theoretical maximal number of correct alternations and expressed as percentage (%).
Immunohistochemical Staining
As described in one of our recent studies (Shao et al. 2015), the immunohistochemical staining was performed. Serial coronal sections (25 μm) of the brain were cut using a cryostat microtome (LeicaCM1850, Wetzler, Germany) and collected in the PBS-filled wells of 6-well plates. The floating brain sections were washed with 0.01 M PBS for 5 min × 3 and steeped in 0.6% hydrogen peroxide in methanol at room temperature to wipe off endogenous peroxidase activity. After rinsing in PBS, the sections were incubated with a goat serum blocking solution for 30 min at room temperature. The primary antibody to myelin basic protein (MBP) (Abcam, Cambridge, UK) or pi form of glutathione-S-transferase (GST-π) (Boster Biotechnology, Wuhan, China) in the 0.3% Triton X-100 with PBS was applied at 4 °C overnight, followed by incubation at room temperature for 30 min. GST-π is a biomarker for mature oligodendrocytes and MBP is a primary structural protein of the myelin sheath. After rinsing in PBS, sections were incubated with the secondary antibody associated with horseradish peroxidase (HRP; Zhongshan Gold Bridge Biology Company, Beijing, China) at 37 °C for 30 min. Then, the sections were visualized by adding diaminobenzidine solution as recommended by the manufacturer (Zhongshan Gold Bridge Biology Company, Beijing, China). After rinses in PBS, the sections were pasted onto the glass slides, dehydrated by gradient ethyl alcohol and cleared by xylene. After mounted by neutral balsam, immunohistochemical staining was observed and recorded with a Zeiss microscope (Zeiss instruments Inc., Germany) and analyzed with the Image-Pro-Plus 6.0 software (Media Cybernetics, Rockville, Maryland).
Immunofluorescence Staining
As described in one of our recent studies (Shao et al. 2015), the immunofluorescence staining was performed. The free-floating sections were washed with PBST (PBS/0.5% Tween 20) for 5 min × 3. Then the sections were incubated with goat serum working solution (Beyotime, Biotechnology, Shanghai, China) for 30 min at room temperature. Subsequently, the primary rabbit polyclonal anti-Iba-1(Iba-1 is a microglia activation marker; Wako, Osaka, Japan) was applied overnight at a 1:1000 dilution at 4 °C. After rinsing three times in PBST, sections were incubated with Cy3-conjugated goat anti-rabbit antibody (Beyotime Biotechnology, Shanghai, China) at room temperature for 60 min. After rinsing in PBST, the sections were pasted and mounted by the Fluor mount (Abcam, Cambridge, UK). The immunofluorescence was observed under a fluorescence microscope (Zeiss instruments Inc., Germany) and images were recorded with the system. The slides were preserved under 4 °C and shielded from light. The number of Iba-1 positive cells was counted with the Image-Pro Plus 6.0 software (Media Cybernetics, Rockville, Maryland).
Image Analysis
As described in one of our recent studies (Shao et al. 2015), the quantitative analysis of immune-histochemical and immunofluorescence staining was performed. The examined brain regions included the cerebral cortex (CTX, including frontal cortex and parietal cortex) and the CA3 of hippocampus. For each brain region, three tissue sections were chosen. For each section, three images were recorded under the same conditions. The Image-Pro Plus 6.0 software (Media Cybernetics, Rockville, Maryland) was used to automatically read out the measurement values. The data of MBP-like immunoreactivity were expressed as integrated optical density (IOD), which equals to area x average optical density. The data of GST-π and MBP were normalized to the percentage of Control or N-folds of CPZ-VEH, whereas that of Iba-1 positive cells were normalized with the Control as the comparator.
Western-Blot Analysis
Frontal cortex samples were homogenized in RIPA lyses buffer (50 mM Tris, 150 mM NaCl, 1% NP40, 0.5% sodium deoxycholate, and 0.1% SDS) with freshly added protease inhibitor cocktail (Sigma, St. Louis, MO, USA). The suspension was collected after centrifugation at 12,000 rpm for 10 min at 4 °C. Total protein concentration was quantified using a BCA kit (Pierce, Nepean, Ontario, Canada), and protein samples were loaded onto 10% SDS-polyacrylamide gel and subjected to electrophoresis at 70 V and 110 V until the bromophenol blue reached the bottom of the gel. The proteins were then transferred to PVDF membranes. Membranes were incubated in a blocking solution of 5% no-fat milk powder in PBS for 1 h at 22 °C, followed by incubation in a blocking solution containing the primary antibody to MBP (1: 10,000; Santa Cruz Biotechnology, CA, USA) overnight at 4 °C. The membranes were then washed with PBS three times, followed by incubation in the blocking solution containing the rabbit anti-goat secondary antibodies (1: 10,000). After three rinses in PBS, immunoreactivity bands were developed using an ECL detection kit (Amersham Biosciences, Baie d′Urfe Quebec). To confirm equal amounts of loading samples, β-actin was also labeled (1:5000; Sigma, St. Louis, MO, USA) through the same procedures as described above.
Enzyme-Linked Immunosorbent Assay
The concentrations of the cytokines IL-4, IL-10, IL-1β and TNF-α were measured using the ELISA kits (Boster, Wuhan, China) and following the operation steps offered by the manufacturer. Briefly, the brain tissue samples were homogenized in PBS (0.01 M, pH = 7.4) at 10% (w/v). Then, centrifuged at 2800 g for 20 min at 4 °C, and the supernatant was collected. The standards and samples in duplicate (100 μl/well) were added into the ELISA plates and incubated for 90 min at 37 °C. Then, the liquid was thrown away, 100 μl biotin-labeled anti-mouse IL-4, IL-10, IL-1β or TNF-α working solution were added into the wells and incubated for 60 min at 37 °C. Then washing with PBS three times, followed by the addition of 100 μl ABC (avidin-biotin-peroxidase complex) working solution and incubation for 30 min at 37 °C. Washing with PBS three times again, followed by the addition of 90 μl TMB (3,3',5,5'-tetramethylbenzidine) working solution and reaction for 20–25 min at 37 °C while shielded from light, and stopped with 100 μl TMB buffer. The plates were read by the microplate reader (Bio-Rad laboratories, Inc., California, USA) at 450 nm. The concentrations of IL-4, IL-10, IL-1β and TNF-α in the samples were calculated by referring to the standard curve and expressed as pg/mg tissue.
Statistical Analysis
The GraphPad Prism software (version 7; Cabit Information Technology Co., LTD, Shanghai, China) was employed for data analyses. Student t test was performed for the comparisons between Control and CPZ groups, or between CPZ-VEH and CPZ-AD10. For the social recognition data, repeated-measure ANOVA (analysis of variance) was used to compare values under E and A conditions. For the other data, one-way ANOVA was performed, followed by Tukey HSD test for multiple comparisons. A normality test was performed before t-test and one-way ANOVA. p < 0.05 was considered statistically significant.
Results
Adenosine Promotes the Recovery of C57BL/6 Mice from the CPZ-Induced Behavioral Changes
The open-field test resulted in two indices for each group of animals, including TD (distance travelled on the whole open field) and time spent on the central zone. Student t test indicated a significant difference between Control and CPZ groups in terms of the time spent on the central zone of the open-field (42.57 ± 15.06 vs 22.75 ± 11.74, t = 2.74, p = 0.018; Fig. 1a), but not on TD. In terms of the Y-maze test results, Student t test indicated a significant difference between Control and CPZ groups in spontaneous alternation (67% ± 15 vs 54% ± 7%, t = 2.30, p = 0.040; Fig. 1b), but not in the number of arm visiting. In the social interaction test, only the mice in the Control can differentiate the two conditions of with (A) and without (E) an unfamiliar conspecific inside the wire-mesh cage (Fig. 1c). This was further confirmed by significant difference between the two groups in terms of the ratio of time spent in the interaction zone when an unfamiliar conspecific was inside the wire-mesh cage there over that when the cage was empty (SIR; 1.38 ± 0.48 vs 0.86 ± 0.19, t = 2.67, p = 0.020; Fig. 1d).

Cuprizone-induced behavioral changes in mice. a CPZ-exposed mice spent less time on the central zone of the open-field, indicating a relatively high level of anxiety compared to Control group. b CPZ-exposed mice presented a significantly lower rate of spontaneous alternation in the Y-maze test compared to Control group, indicating the presence of sptail working memory impairment. c Only the mice in the Control group can differentiate the tow conditions of without (E) and with (A) an unfamiliar conspecifc inside the metal mesh cage. d CPZ-exposed mice showed a lower SIR in the social interation test compared to Control group, i.e. they spent less time when an unfamiliar conspecific was in the cage at the interaction zone and/or much time when the cage was empty. N = 9–10/group. Data are presented as mean ± SE. *p < 0.05. SIR, ratio of social interaction time under the two different conditions
One-way ANOVA indicated no significant effect of adenosine administration on TD, but showed significant effect on time spent on the central zone [F (3, 38) = 3.960, p = 0.016]. In regard of the time spent on the central zone, Tukey’s multiple comparison test showed significant differences between CPZ-AD10 (54.77 ± 4.77) and the other groups (34.73 ± 5.91/CPZ-VEH; 37.28 ± 3.66/CPZ-AD1; 41.17 ± 3.52/CPZ-AD5) (Fig. 2a). And the groups CPZ-AD10 and CPZ-AD5 reached the same levels as that of the Control group, indicating a complete recovery.

Adenosine promoted the recovery of CPZ-induced behavioral changes in mice. a Mice in the CPZ-AD10 group spent much more time on the central zone of the open-field compared to the other groups, indicating a lowest level of anxiety. b Mice in the CPZ-AD10 group showed a highest rate of spontaneous alternation in the Y-maze test, indicating a best performance in the spatial working memory. c Only the mice in the CPZ-AD10 group could tell an empty cage from that with an unfamiliar conspecific in the social interation test, indicating the intactness of their social interaction function. d All adenosine treated mice showed significantly higher SIR compared to the CPZ-VEH group. N = 9–10/group. Data are presented as mean ± SE. *p < 0.05, **p < 0.01, comparisons were made between CPZ-AD10 and CPZ-VEH, or between E and A. E, an empty cage; A, a cage with an unfamiliar conspecific inside it
As for the Y-maze test, one-way ANOVA indicated no significant effect of adenosine administration on the number of arm visiting, but showed significant effect on spontaneous alternation [F (3, 31) = 3.98, p = 0.02]. Tukey’s multiple comparison test showed significant differences between CPZ-AD10 (72.31 ± 3.22) and CPZ-VEH (59.60 ± 4.25), or CPZ-AD1 (56.84 ± 2.55) (Fig. 2b). Both the groups CPZ-AD10 and CPZ-AD5 reached the same levels as that of the Control group, indicating a complete recovery.
For the social recognition test, the repeated-measure one-way ANOVA was done to compare the time spent by each group (mice) in the interaction zone under the two conditions of an empty cage (E) and a cage with an unfamiliar conspecific (A). Only did the mice in CPZ-AD10 group tell the E from A condition (73.32 ± 7.99 vs 101.56 ± 6.52, p < 0.05] (Fig. 2c), as shown in the Control group. As for SIR, all adenosine-treated groups showed significantly higher SIR compared to the CPZ-VEH group (Fig. 2d).
Adenosine Promotes the Remyelinating Process of C57BL/6 Mice after CPZ Withdrawal
In accordance with previous studies by our team (Xiao et al. 2008; Xu et al. 2009; Yang et al. 2009) and the other investigators (Chandran et al. 2012; Matsushima and Morell 2001), CPZ-exposed mice showed obvious oligodendrocyte loss and myelin breakdown in various brain regions. Shown here are representative images showing the immunohistochemical staining of oligodendrocytes and myelin sheath in cerebral cortex and CA3 region of the hippocampus labeled by the antibodies against GST-π and MBP, respectively (Fig. 3, upper panel). The statistical analysis of their quantile data indicated significant differences between the Control and CPZ groups in terms of the number of GST-π positive cells and IOD of MBP-immunostaining (Fig. 3, lower panel).

Cuprizone-induced oligodendrocyte loss and demyelination. The microphotographs in the upper panel are representative ones from CTX and CA3 of the hippocampus of CPZ-exposed mice and normal Control, respectively. The bar charts in the lower panel show the statistical analysis results of the quantitative data of immunostained oligodendrocytes and MBP in the above-mentioned brain regions. N = 6/group. Data are presented as mean ± SE. **p < 0.01
For the quantitative data of the groups CPZ-VEH, CPZ-AD1, CPZ-AD5, CPZ-AD10, one-way ANOVA indicated significant effects of adenosine administration on the number of GST-π positive cells in both CTX [F (3, 31) = 14.203, p < 0.001] and CA3 [F (3, 31) = 11.060, p < 0.001]. Tukey’s multiple comparison test showed significant increases in GST-π positive cells in CTX of all adenosine-treated C57BL/6 mice relative to the CPZ-VEH group (Fig. 4a). In the case of CA3, adenosine at 5 and 10 mg/kg, but not at 1.0 mg/kg, significantly increased GST-π positive cells compared with the CPZ-VEH group (Fig. 4b). However, only the group CPZ-AD10 reached the same levels of that in Control group, indicating a complete recovery in the CPZ-induced oligodendrocyte loss. In addition, one-way ANOVA indicated significant effects of adenosine administration on the IOD of MBP-immuno-reactivity in both CTX [F (3, 31) = 11.234, p < 0.001] and CA3 [F (3, 31) = 3.652, p = 0.024]. Tukey’s multiple comparison test showed significant increases in IOD of MBP in CTX of all adenosine-treated C57BL/6 mice relative to the CPZ-VEH group (Fig. 4c). In the case of CA3, only adenosine at 10 mg/kg significantly increased the IOD of MBP compared to the CPZ-VEH group (Fig. 4d). However, only the group CPZ-AD10 reached the same levels of that in Control group, indicating a complete recovery in the CPZ-induced myelin breakdown.

Adenosine promoted the recovery of CPZ-induced oligodendrocyte loss and demyelination. a Compared to CPZ-VEH group, all adenosine treated mice (CPZ-AD1, CPZ-AD5, and CPZ-AD10) presented many more oligodendrocyte cells in CTX. b Compared to CPZ-VEH group, the mice in CPZ-AD5 and CPZ-AD10 presented many more oligodendrocyte cells in CA3. c Compared to CPZ-VEH group, all adenosine treated mice (CPZ-AD1, CPZ-AD5, and CPZ-AD10) presented higher levels of MBP-like immunoreactivity in CTX. d Compared to CPZ-VEH group, the mice in CPZ-AD10 group presented higher levels of MBP-like immunoreactivity in CA3. N = 7–8/group. Data are presented as mean ± SE. *p < 0.05, **p < 0.01, ***p < 0.001, compared to CPZ-VEH
To confirm the beneficial effect of adenosine administration on remyelinating in the CPZ-exposed mice as shown in MBP-immunohistochemical staining, Western-blot analysis was performed with the tissue samples of CTX and hippocampus. The representative Western-blot images made from CTX and hippocampus tissue were shown in Fig. 5a and b. The quantitative data of MBP protein levels were analyzed by one-way ANOVA, which indicated significant effects of adenosine administration on this index in both CTX [F (3, 21) = 8.01, p = 0.0013] and hippocampus [F (3, 21) = 8.86, p = 0.0006]. Tukey’s multiple comparison test showed significant increases in MBP protein levels in CTX of all adenosine-treated C57BL/6 mice relative to the CPZ-VEH group (Fig. 5c). In the case of hippocampus, adenosine at 5 and 10 mg/kg significantly increased MBP levels as compared to the CPZ-VEH group; whereas adenosine at 1.0 mg/kg exerted no influence on this index (Fig. 5d).

Adenosine increased MBP expression in CTX and hippocampus of CPZ-exposed mice. a and b are representative western blot images showing MBP immunoreactive bands of CTX and hippocampus tissue samples, respectively. c and d show the corresponding quantitative data of all four groups. N = 7–8/group. Data are presented as mean ± SE. *p < 0.05, **p < 0.01, compared to CPZ-VEH
Adenosine Modulates the Expression of Inflammatory Cytokines while Inhibiting Microgliosis in CTX and Hippocampus
To test our hypothesis that the beneficial effects of adenosine on the recovery of mice from the CPZ-induced behavioral and morphological changes may result from its anti-inflammatory actions on the CNS lesions, we examined the microglia, the resident immune cells in the CNS, by means of immuno-fluorescence staining. In addition, we measured levels of cytokines IL-1β, TNF-α, IL-4, and IL-10 in CTX and hippocampus tissue. Of them, the former two are pro-inflammatory mediators, whereas the latter two are anti-inflammatory ones.
The representative immunofluorescent images of Iba-1 positive cells in CTX and CA3 of the hippocampus from the mice of Control, CPZ, CPZ-VEH, and CPZ-AD10 groups were shown in Fig. 6a and b. The student t-test was performed to compare quantitative data of Iba-1 positive cells between Control and CPZ, as well as between CPZ-VEH and CPZ-AD10. The results showed significant increases in number of Iba-1 positive cells in both CTX and CA3 of CPZ treated mice compared to Control mice. In contrast, the numbers significantly decreased in the CPZ-AD10 group relative to the CPZ-VEH group (Fig. 6c and d).

CPZ exposure for 5 weeks induced microglia activation, but adenosine (10 mg/kg) administration for 7 days suppressed the activation. a and b are representative microphotographs of Iba-1 immunofluorescent staining in CTX and hippocampal CA3, respectively. c and d show the corresponding quantitative data of all four groups. N = 7–8/group. Data are presented as mean ± SE. *p < 0.05, **p < 0.01, ***p < 0.001, between groups indicated
As for levels of cytokines in CTX, student t-test showed no significant difference between the normal Control and CPZ groups, and between CPZ-VEH and CPZ-AD10.0 groups, in terms of IL-1β (Fig. 7a). However, CPZ-exposure for 5 weeks significantly increased levels of the pro-inflammatory cytokine TNF-α in CTX relative to the normal Control group (2.796 ± 0.198 vs 0.212 ± 0.026, p < 0.001). In contrast, adenosine (10.0 mg/kg) administration for 1 week after CPZ-withdrawal significantly decreased TNF-α levels as compared to the CPZ-VEH group (0.738 ± 0.146 vs 2.214 ± 0.262, p < 0.001) (Fig. 7b). For levels of the anti-inflammatory cytokine IL-4 in CTX, student t-test showed no significant difference between the normal Control and CPZ groups, and between CPZ-VEH and CPZ-AD10.0 groups (Fig. 7c). However, CPZ-exposure for 5 weeks significantly decreased levels of the anti-inflammatory cytokine IL-10 in CTX relative to the normal Control group (0.532 ± 0.114 vs 1.113 ± 0.103, p < 0.01). In contrast, adenosine (10.0 mg/kg) administration for 1 week after CPZ-withdrawal significantly increased IL-10 levels as compared to the CPZ-VEH group (1.182 ± 0.069 vs 0.635 ± 0.077, p < 0.001) (Fig. 7d).

Effects of CPZ exposure and adenosine (10 mg/kg) administration after CPZ withdrawal on levels of inflammatory cytokines in CTX of the mouse. There were no differences between Control and CPZ, as well as between CPZ-VEH and CPZ-AD10 in levels of IL-1β in CTX (a). CPZ exposure for 5 weeks increased, while adenosine administration for 7 days decreased, TNF-α levels in CTX (b). There were no differences between Control and CPZ, as well as between CPZ-VEH and CPZ-AD10 in levels of IL-4 in CTX (c). CPZ exposure for 5 weeks decreased, while adenosine administration for 7 days increased, IL-10 levels in CTX (d). N = 7–8/group. Data are presented as mean ± SE. **p < 0.01, ***p < 0.001, between groups indicated
For levels of IL-1β in the hippocampus, student t-test showed a significant increase in CPZ group relative to the normal Control group (3.665 ± 0.354 vs 1.652 ± 0.206, p < 0.001). In contrast, adenosine (10 mg/kg) administration for 1 week after CPZ-withdrawal significantly decreased IL-1β levels compared to the CPZ-VEH group (2.251 ± 0.331 vs 4.553 ± 0.384, p < 0.001) (Fig. 8a). Similarly, CPZ-exposure for 5 weeks significantly increased levels of TNF-α in the hippocampus relative to the normal Control group (3.318 ± 0.253 vs 2.390 ± 0.270, p < 0.05). However, there was no significant difference between CPZ-VEH and CPZ-AD10.0 groups (2.888 ± 0.308 vs 2.387 ± 0.217, p = ns) (Fig. 8b). As for levels of IL-4 in the hippocampus, student t-test showed no significant difference between the normal Control and CPZ groups, but a significant increase in the CPZ-AD10.0 group relative to the CPZ-VEH group (3.298 ± 0.383 vs 1.792 ± 0.390, p < 0.05). (Fig. 8c). CPZ-exposure for 5 weeks significantly increased levels of IL-10 in the hippocampus relative to the normal Control group (6.680 ± 0.562 vs 4.790 ± 0.262, p < 0.05). There was no significant difference between the CPZ-VEH and CPZ-AD10.0 groups (6.37 ± 0.643 vs 6.43 ± 0.921, p = ns). The both groups kept the high levels of IL-10 in the hippocampus (Fig. 8d).

Effects of CPZ exposure and adenosine (10 mg/kg) administration after CPZ withdrawal on levels of inflammatory cytokines in hippocampus of the mouse. CPZ exposure for 5 weeks increased, while adenosine administration for 7 days decreased, IL-1β levels in hippocampus (a). CPZ exposure for 5 weeks increased TNF-α levels in hippocampus, but adenosine administration for 7 days had no effect on this index (b). There were no differences between Control and CPZ in levels of IL-4, while adenosine administration increased this index in the hippocampus (c). CPZ exposure for 5 weeks increased IL-10 levels in hippocampus, but adenosine administration for 7 days had no effect on this index (d). N = 7–8/group. Data are presented as mean ± SE. *p < 0.05, ***p < 0.001, between groups indicated
Discussion
As reviewed in the introduction section, both MS and schizophrenia involve white matter abnormalities and neuroinflammation in the brain. Because of these two pathological features and the concurrent behavioral changes, the CPZ-exposed mouse has provided a novel animal model of demyelination and remyelinating, as well as to test the neuroinflammation and oligodendrocyte dysfunction hypotheses of schizophrenia. In the present study, CPZ-exposure for 5 weeks induced dramatic decrease in oligodendrocytes and salient demyelination in the cerebral cortex and hippocampus, accompanied by behavioral changes indicative of high levels of anxiety, impaired spatial working memory, and social interaction deficit. All these changes replicated the findings in previous studies by our team (Xiao et al. 2008; Xu et al. 2009; Zhang et al. 2008) and the others (Matsushima and Morell 2001; Tezuka et al. 2013). More significantly, administration of adenosine for 7 days promoted the behavioral recovery of CPZ-exposed mice and accelerated the remyelinating process in the brains of mice after CPZ withdrawal in a dose-dependent manner. In addition, the effective dose (10 mg/kg) of adenosine inhibited microglia activation and suppressed abnormal elevation of the pro-inflammatory cytokines IL-1β and TNF-α in CTX and hippocampus, but increased levels of the anti-inflammatory cytokines IL-4 or IL-10 in the same brain regions during the remyelinating process.
In a previous animal study looking at the recovery process of mice after CPZ exposure, the behavioral changes induced by CPZ exposure for 5 weeks showed various recovery processes. Spatial working memory impairment gradually recovered during a three-week post-CPZ recovery period; the antipsychotics clozapine, olanzapine, and quetiapine, but not haloperidol, significantly promoted the recovery process. Social interaction deficit always kept at lower levels during the recovery period and showed no response to the antipsychotics mentioned above. In contrast, the performance alterations of mice in the elevated plus maze spontaneously recovered to normal control level in 2 weeks after CPZ withdrawal (Xu et al. 2011). In another study, the CPZ-induced spatial working memory impairment showed a similar recovery process while the locomotor activity kept at a constant high level during a four-week post-CPZ recovery period (Zhang et al. 2012). In the present study, only the CPZ-AD10 group showed significantly improved performances in open-field, Y-maze, and social interactions tests, and these mice performed comparable to those in Control group, indicating a complete recovery in the behaviors tested. The promoting effects of adenosine at the high dose on the recovery of the CPZ-induced behavioral changes accomplished within 1 week after CPZ withdrawal, faster than the recovery-promoting effects of some antipsychotics. Furthermore, the complete recovery of social interaction deficit in the CPZ-AD10 group suggests a great potential of adenosine to treat this abnormal behavior which was shown to be persistent and resistant to antipsychotics in previous studies (Xu et al. 2011; Zhang et al. 2012).
The CPZ intoxication model has a consistent pattern of demyelination and remyelinating. After 5 or 6 weeks of continuous CPZ-exposure, demyelination is maximal and remyelinating begins. Withdrawal of CPZ at this stage results in significant remyelinating over the course of about 4 weeks (Matsushima and Morell 2001; Stidworthy et al. 2003). As such, recent studies have examined effects of some extant drugs on remyelinating process in the search for a potent approach to promoting remyelinating in the CPZ-exposed mouse. Two examples relevant to the present study are quetiapine and bucladesine. The former is an atypical antipsychotic and the latter is a cell-permeable analogue of cAMP (cyclic adenosine monophosphate). Quetiapine was shown to significantly accelerate remyelinating after CPZ withdrawal (Wang et al. 2015; Xu et al. 2011; Zhang et al. 2012), while bucladesine treatment exhibited a protective effect on myelination and OPC (oligodendrocyte precursor cell) maturation (Vakilzadeh et al. 2015). Like bucladesine, treatment with rolipram, a cAMP-dependent phosphodiesterase inhibitor, promoted OPC maturation (Sun et al. 2012) and increased the number of remyelinated axons around the lesion borders (Syed et al. 2013). In the present study, adenosine administration for 7 days accelerated the remyelinating process or OPC maturation after CPZ withdrawal as evidenced by many more mature oligodendrocytes (GST-π positive cells) and significantly higher MBP-immunoreactivity and MBP protein expression levels in CPZ-AD groups compared to CPZ-VEH group. All three doses of adenosine applied in this study resulted in increased levels of the above indices in CTX as compared to CPZ-VEH group, while only middle (5 mg/kg) or high (10 mg/kg) dose of adenosine showed significant effects in the hippocampus, suggesting CTX is more sensitive to adenosine treatment than the hippocampus. In line with these beneficial effects of adenosine on remyelinating, all adenosine receptor subtypes are expressed by OPCs (Fredholm et al. 2011; Stevens et al. 2002) and a key role of adenosine in oligodendrocyte maturation has been recognized (Burnstock and Ulrich 2011).
In the brain tissue of CPZ-exposed mice, loss of myelin and oligodendrocytes is followed or accompanied with recruitment of microglia, macrophages and astrocytes (Shao et al. 2015; Stidworthy et al. 2003; Tezuka et al. 2013; Zhang et al. 2008). In accordance with the previous studies, CPZ exposure for 5 weeks significantly increased number of Iba-1 positive cells while the same treatment induced demyelination and oligodendrocyte loss in both CTX and hippocampus of the mouse. Together, the results of the present study and previous studies suggest the involvement of microglia, macrophages and astrocytes in the demyelination and remyelinating processes. In support of this suggestion, CPZ-AD10 group showed significant fewer numbers of Iba-1 positive cells while presented many more GST-π positive cells in both CTX and hippocampus as compared to CPZ-VEH group. In line with these results, previous studies showed that quetiapine inhibited demyelination and oligodendrocyte loss while suppressed microglia recruitment and astrogliosis in CPZ-exposed mice (Shao et al. 2015; Zhang et al. 2008). Also, this antipsychotic inhibited microglial activation while promoted myelin repair in the CPZ-exposed mice (Wang et al. 2015).
Microglia are resident immune cells of the CNS. These small cells have several functions, including pathogen recognition, phagocytosis, antigen presentation, and synapse remodeling (Boche et al. 2013). In non-pathological conditions microglia are of quiescence or rest although they constantly survey the CNS environment (Marshall et al. 2013; Nimmerjahn et al. 2005). In response to neuronal injury or even subtle perturbations of the CNS environment, resident microglia react with a set of stereotypic changes in their physiological state (Kreutzberg 1996; Marshall et al. 2013). Activated microglia may become hyper-ramified or ameboid/phagocytic (Boche et al. 2013), and synthesize and release pro-inflammatory molecules, including IL-1β, TNF-α, IL-6, superoxide radicals, glutamate, and nitric oxide (Barger et al. 2007; Takaki et al. 2012). These molecules, via enhancing oxidative stress and activating cell death pathways through stimulation of kinases and caspase cascades, exert deleterious effects on susceptible cell populations, especially oligodendrocytes and neurons (Monji et al. 2013). Alternatively, activated microglia may release anti-inflammatory cytokines such as IL-4, IL-10, insulin-growth factor-1, transforming growth factor-b, and neurotrophic factors (Boche et al. 2013; Ekdahl 2012; Hu et al. 2015), which facilitate healing and limit neuronal injury (Devinsky et al. 2013). The former phenotype is termed as the classical M1 activation and the latter M2 activation.
Based on the knowledge about microglia as reviewed above, the CPZ-induced increases in Iba-1 positive cells in CTX and hippocampus, and the suppression of these increases by adenosine following CPZ withdrawal in the present study, it is plausible to speculate that adenosine applied in this study might inhibit M1 activation and facilitate M2 activation of microglia during the recovery period after CPZ withdrawal. In support of this speculation, CPZ exposure for 5 weeks increased levels of TNF-α in CTX, and levels of IL-1β and TNF-α in hippocampus, indicating the M1 activation of microglia; adenosine (10 mg/kg) administration for 1 week decreased levels of TNF-α in CTX and of IL-1β in hippocampus, suggesting the inhibition of M1 activation by adenosine; and adenosine increased levels of IL-10 in CTX and of IL-4 in the hippocampus, implying the M2 activation of microglia by adenosine.
In line with these in vivo data, adenosine inhibited the lipopolysaccharide-induced expression of iNOS, but increased the expression of Arg-1 in RAW264.7 macrophage cells in a dose-dependent manner, demonstrating the effects of adenosine on the polarizations of the RAW264.7 cells (unpublished data).
Although we did not explore the underlying mechanisms for the effects of adenosine on remyelinating and microglia activation in the CPZ-exposed mouse, the beneficial effects shown in this study can be explained in the context of the findings from recent studies addressing roles of A1 and A2a receptors in OPC maturation and microglia activation. In a study by Asghari et al. (2013), CHA (an A1 receptor agonist) reduced the extent of demyelination during demyelination phase, but increased remyelinating during remyelinating phase in an experimental model of rat optic chiasm demyelination. In addition, the activation of A1 receptor has been reported to induce OPC migration (Othman et al. 2003). In a more recent study, two A2a receptor agonists (CGS21680 and ATL313) significantly attenuated progression of motor symptoms following a single intrathecal administration at the onset of motor symptoms induced by EAE. In the meanwhile, the treatment decreased markers of activation in microglia and monocyte-lineage cells (Loram et al. 2015). Relevant to this finding, mice lacking A2a receptor showed increased chemokine production with subsequent greater immune cell infiltration into the spinal cord during EAE (Mills et al. 2012; Yao et al. 2012). Therefore, we may speculate that adenosine promoted remyelinating via acting on A1 receptor while modulated microglia activation via its action on A2a receptor in brain tissue of the CPZ-exposed mice. Both the effects of promoting remyelinating and modulating microglial activation may contribute to the beneficial effects of adenosine on behavioral recovery process in the CPZ-exposed mice.
A1 and A2a receptors also play key roles in regulating the entry of adenosine into the brain. In a recent study, the activation of A2a receptor was shown to increase accumulation of drugs into the brain in a time- and dose-dependent manner. Also, a broad-spectrum agonist NECA increased BBB permeability and supported macromolecule delivery to the CNS in experimental setting (Carman et al. 2011), suggesting a possible role of A1 receptors. In a more recent study, A2a activation by Lexiscan (one of A2a receptor agonists) rapidly increased accumulation of the chemotherapeutic drug and P-gp substrate epirubicin in the mouse brain by downmodulating the expression level of P-gp (Kim and Bynoe 2016), one of multidrug-resistant transporters highly expressed on brain endothelial cells and hinder the effective delivery of drugs into the CNS. We were aware of the short half-life of adenosine, up to 90% of which is metabolized in the brain within 10 min (Pardridge et al. 1994), and the fact that BBB permeation induced by NECA treatment (half-life, 4 h) lasted significantly longer than that induced by Lexiscan treatment (half-life, 2.5 min) (Carman et al. 2011). Interestingly, despite its short half-life, extracellular adenosine permeabilized the BBB to entry of 10 kDa dextran (Bynoe et al. 2015).
In conclusion, short-term administration of adenosine promoted behavioral recovery and remyelinating while it modulated microglia activation and inflammatory cytokines in CPZ-exposed mice. These results provide experimental in vivo evidence for an important role of activated microglia in remyelinating process. The beneficial effects of adenosine on the CPZ-exposed mice may be of relevance to the treatment of MS and schizophrenia, both of which involve white matter abnormalities and neuroinflammation in the brain.
References
Asghari AA, Azarnia M, Mirnajafi-Zadeh J, Javan M (2013) Adenosine A1 receptor agonist, N6-cyclo- hexyladenosine, protects myelin and induces remyelination in an experimental model of rat optic chiasm demyelination; electrophysiological and histopathological studies. J Neurol Sci 325:22–28
Barger SW, Goodwin ME, Porter MM, Beggs ML (2007) Glutamate release from activated microglia requires the oxidative burst and lipid peroxidation. J Neurochem 101:1205–1213
Ben-Shachar D (2002) Mitochondrial dysfunction in schizophrenia: a possible linkage to dopamine. J Neurochem 83:1241–1251
Berton O, McClung CA, Dileone RJ, Krishnan V, Renthal W, Russo SJ et al (2006) Essential role of BDNF in the mesolimbic dopamine pathway in social defeat stress. Science 311:864–868
Boche D, Perry VH, Nicoll JA (2013) Review: activation patterns of microglia and their identification in the human brain. Neuropathol Appl Neurobiol 39:3–18
Burnstock G, Ulrich H (2011) Purinergic signaling in embryonic and stem cell development. Cell Mol Life Sci 68:1369–1394
Bynoe MS, Viret C, Yan A, Kim DG (2015) Adenosine receptor signaling: a key to opening the blood-brain door. Fluids Barriers CNS 12:20. https://doi.org/10.1186/s12987-015-0017-7
Carman AJ, Mills JH, Krenz A, Kim DG, Bynoe MS (2011) Adenosine receptor signaling modulates permeability of the blood-brain barrier. J Neurosci 31:13272–13280
Chandran P, Upadhyay J, Markosyan S, Lisowski A, Buck W, Chin CL et al (2012) Magnetic resonance imaging and histological evidence for the blockade of cuprizone-induced demyelination in C57BL/6 mice. Neuroscience 202:446–453
Chang H, Liu J, Zhang Y, Wang F, Wu Y, Zhang L et al (2017) Increased central dopaminergic activity might be involved in the behavioral abnormality of cuprizone exposure mice. Behav Brain Res 331:143–150
Davis KL, Stewart DG, Friedman JI, Buchsbaum M, Harvey PD, Hof PR et al (2003) White matter changes in schizophrenia: evidence for myelin-related dysfunction. Arch Gen Psychiatry 60:443–456
Devinsky O, Vezzani A, Najjar S, De Lanerolle NC, Rogawski MA (2013) Glia and epilepsy: excitability and inflammation. Trends Neurosci 36:174–184
Du C, Xie X (2012) G protein-coupled receptors as therapeutic targets for multiple sclerosis. Cell Res 22:1108–1128
Ekdahl CT (2012) Microglial activation - tuning and pruning adult neurogenesis. Front Pharmacol 3:41
Faizi M, Salimi A, Seydi E, Naserzadeh P, Kouhnavard M, Rahimi A, Pourahmad J (2016) Toxicity of cuprizone a Cu(2+) chelating agent on isolated mouse brain mitochondria: a justification for demyelination and subsequent behavioral dysfunction. Toxicol Mech Methods 26:276–283
Fischer HJ, Schweingruber N, Lühder F, Reichardt HM (2013) The potential role of T cell migration and chemotaxis as targets of glucocorticoids in multiple sclerosis and experimental autoimmune encephalomyelitis. Mol Cell Endocrinol 380:99–107
Franco-Pons N, Torrente M, Colomina MT, Vilella E (2007) Behavioral deficits in the cuprizone-induced murine model of demyelination/remyelination. Toxicol Lett 169:205–213
Fredholm BB, IJzerman AP, Jacobson KA, Klotz KN, Linden J (2001) International Union of Pharmacology. XXV. Nomenclature and classification of adenosine receptors. Pharmacol Rev 53:527–552
Fredholm BB, Johansson S, Wang YQ (2011) Adenosine and the regulation of metabolism and body temperature. Adv Pharmacol 61:77–94
Gandhi R, Laroni A, Weiner HL (2010) Role of the innate immune system in the pathogenesis of multiple sclerosis. J Neuroimmunol 221:7–14
Hu P, Thinschmidt JS, Caballero S, Adamson S, Cole L, Chan-Ling T et al (2015) Loss of survival factors and activation of inflammatory cascades in brain symp thetic centers in type 1 diabetic mice. Am J Physiol Endocrinol Metab 308:688–698
Janes K, Wahlman C, Little JW, Doyle T, Tosh DK, Jacobson KA et al (2015) Spinal neuroimmune activation is independent of T-cell infiltration and attenuated by A3 adenosine receptor agonists in a model of oxaliplatin-induced peripheral neuropathy. Brain Behav Immun 44:91–99
Kim DG, Bynoe MS (2016) A2A adenosine receptor modulates drug efflux transporter P-glycoprotein at the blood-brain barrier. J Clin Invest 126:1717–1733
Kim JS, Kornhuber HH, Schmid-Burgk W, Holzmüller B (1980) Low cerebrospinal fluid glutamate in schizophrenic patients and a new hypothesis onschizophrenia. Neurosci Lett 20:379–382
Kipp M, Nyamoya S, Hochstrasser T, Amor S (2017) Multiple sclerosis animal models: a clinical and histopathological perspective. Brain Pathol 27:123–137
Kosmidis MH, Giannakou M, Messinis L, Papathanasopoulos P (2010) Psychotic features associated with multiple sclerosis. Int Rev Psychiatry 22:55–66
Kreutzberg GW (1996) Microglia: a sensor for pathological events in the CNS. Trends Neurosci 19:312–318
Laghi A, Catalano C, Panebianco V, Iannaccone R, Iori S, Passariello R (2000) Optimization of the technique of virtual colonoscopy using a multislice spiral computerized tomography. Radiol Med 100:459–464
Lassmann H (2018) Multiple sclerosis pathology. Cold Spring Harb Perspect Med 8. https://doi.org/10.1101/cshperspect.a028936
Loram LC, Strand KA, Taylor FR, Sloane E, Van Dam AM, Rieger J et al (2015) Adenosine 2A receptor agonism: a single intrathecal administration attenuates motor paralysis in experimental autoimmune encephalopathy in rats. Brain Behav Immun 46:50–54
Lynch JJ 3rd, Alexander KM, Jarvis MF, Kowaluk EA (1998) Inhibition of adenosine kinase during oxygen-glucose deprivation in rat cortical neuronal cultures. Neurosci Lett 252:207–210
Marshall SA, McClain JA, Kelso ML, Hopkins DM, Pauly JR, Nixon K (2013) Microglial activation is not equivalent to neuroinflammation in alcohol-induced neurodegeneration: the importance of microglia phenotype. Neurobiol Dis 54:239–251
Matsushima GK, Morell P (2001) The neurotoxicant, cuprizone, as a model to study demyelination and remyelination in the central nervous system. Brain Pathol 11:107–116
Merighi S, Bencivenni S, Vincenzi F, Varani K, Borea PA, Gessi S (2017) A2B adenosine receptors stimulate IL-6 production in primary murine microglia through p38 MAPK kinase pathway. Pharmacol Res 117:9–19
Mills JH, Alabanza LM, Mahamed DA, Bynoe MS (2012) Extracellular adenosine signaling induces CX3CL1 expression in the brain to promote experimental autoimmune encephalomyelitis. J Neuroinflammation 9:193
Monji A, Kato T, Kanba S (2009) Cytokines and schizophrenia: microglia hypothesis of schizophrenia. Psychiatry Clin Neurosci 63:257–265
Monji A, Kato TA, Mizoguchi Y, Horikawa H, Seki Y, Kasai M et al (2013) Neuroinflammation in schizophrenia especially focused on the role of microglia. Prog Neuro-Psychopharmacol Biol Psychiatry 42:115–121
Müller N (2018) Inflammation in schizophrenia: pathogenetic aspects and therapeutic considerations. Schizophr Bull. https://doi.org/10.1093/schbul/sby024
Müller N, Weidinger E, Leitner B, Schwarz MJ (2015) The role of inflammation in schizophrenia. Front Neurosci 9:372. https://doi.org/10.3389/fnins.2015.00372
Nimmerjahn A, Kirchhoff F, Helmchen F (2005) Resting microglial cells are highly dynamic surveillants of brain parenchyma in vivo. Science 308:1314–1318
Nylander A, Hafler DA (2012) Multiple sclerosis. J Clin Invest 122:1180–1188
Othman T, Yan H, Rivkees SA (2003) Oligodendrocytes express functional A1 adenosine receptors that stimulate cellular migration. Glia 44:166–172
Pardridge WM, Yoshikawa T, Kang YS, Miller LP (1994) Blood-brain barrier transport and brain metabolism of adenosine and adenosine analogs. J Pharmacol Exp Ther 268:14–18
Parnanzone S, Serrone D, Rossetti MC, D'Onofrio S, Splendiani A, Micelli V, Rossi A, Pacitti F (2017) Alterations of cerebral white matter structure in psychosis and their clinical correlations: a systematic review of diffusion tensor imaging studies. Riv Psichiatr 52:49–66
Praet J, Guglielmetti C, Berneman Z, Van der Linden A, Ponsaerts P (2014) Cellular and molecular neuropathology of the cuprizone mouse model: clinical relevance for multiple sclerosis. Neurosci Biobehav Rev 47:485–505
Ransohoff RM, Hafler DA, Lucchinetti CF (2015) Multiple sclerosis-a quiet revolution. Nat Rev Neurol 11:134–142
Remington LT, Babcock AA, Zehntner SP, Owens T (2007) Microglial recruitment, activation, and proliferation in response to primary demyelination. Am J Pathol 170:1713–1724
Robertson CL, Bell MJ, Kochanek PM, Adelson PD, Ruppel RA, Carcillo JA et al (2001) Increased adenosine in cerebrospinal fluid after’ severe traumatic brain injury in infants and children: association with severity of injury and excitotoxicity. Crit Care Med 29:2287–2293
Selvaraju R, Bernasconi L, Losberger C, Graber P, Kadi L, Avellana-Adalid V et al (2004) Osteopontin is upregulated during in vivo demyelination and remyelination and enhances myelin formation in vitro. Mol Cell Neurosci 25:707–721
Shao Y, Peng H, Huang Q, Kong J, Xu H (2015) Quetiapine mitigates the neuroinflammation and oligodendrocyte loss in the brain of C57BL/6 mouse following cuprizone exposure for one week. Eur J Pharmacol 765:249–257
Stevens B, Porta S, Haak LL, Gallo V, Fields RD (2002) Adenosine: a neuron-glial transmitter promoting myelination in the CNS in response to action potentials. Neuron 36:855–868
Stidworthy MF, Genoud S, Suter U, Mantei N, Franklin RJ (2003) Quantifying the early stages of remyelination following cuprizone-induced demyelination. Brain Pathol 13:329–339
Sun X, Liu Y, Liu B, Xiao Z, Zhang L (2012) Rolipram promotes remyelination possibly via MEK-ERK signal pathway in cuprizone-induced demyelination mouse. Exp Neurol 237:304–311
Syed YA, Baer A, Hofer MP, González GA, Rundle J, Myrta S et al (2013) Inhibition of phosphor- diesterase-4 promotes oligodendrocyte precursor cell differentiation and enhances CNS remyelination. EMBO Mol Med 5:1918–1934
Takaki J, Fujimori K, Miura M, Suzuki T, Sekino Y, Sato K (2012) L-glutamate released from activated microglia downregulates astrocytic L-glutamate transporter expression in neuroinflammation: the ‘collusion’ hypothesis for increased extracellular L-glutamate concentration in neuroinflammation. J Neuroinflammation 9:275
Tezuka T, Tamura M, Kondo MA, Sakaue M, Okada K, Takemoto K et al (2013) Cuprizone short-term exposure: astrocytic IL-6 activation and behavioral changes relevant to psychosis. Neurobiol Dis 59:63–68
Tutuncu M, Tang J, Zeid NA, Kale N, Crusan DJ, Atkinson EJ, Siva A, Pittock SJ, Pirko I, Keegan BM, Lucchinetti CF, Noseworthy JH, Rodriguez M, Weinshenker BG, Kantarci OH (2013) Onset of progressive phase is an age-dependent clinical milestone in multiple sclerosis. Mult Scler 19:188–198
Vakilzadeh G, Khodagholi F, Ghadiri T, Darvishi M, Ghaemi A, Noorbakhsh F et al (2015) Protective effect of a cAMP analogue on behavioral deficits and neuropathological changes in cuprizone model of demyelination. Mol Neurobiol 52:130–141
van Rossum JM (1967) The effect of psychostimulants on the central and autonomic nervous system. Schweiz Z Sportmed 15:26–40
Wang H, Liu S, Tian Y, Wu X, He Y, Li C et al (2015) Quetiapine inhibits microglial activation by neutralizing abnormal STIM1-mediated intercellular calcium homeostasis and promotes myelin repair in a cuprizone-induced mouse model of demyelination. Front Cell Neurosci 9:492
Xiao L, Xu H, Zhang Y, Wei Z, He J, Jiang W et al (2008) Quetiapine facilitates oligodendrocyte development and prevents mice from myelin breakdown and behavioral changes. Mol Psychiatry 13:697–708
Xu H (2015) Neuroinflammation in schizophrenia focused on the pharmacological and therapeutic evidence. Pharmacologia 6:438–453
Xu H, Li XM (2011) White matter abnormalities and animal models examining a putative role of altered white matter in schizophrenia. Schizophr Res Treatment 2011:826976. https://doi.org/10.1155/2011/826976
Xu H, Yang HJ, Zhang Y, Clough R, Browning R, Li XM (2009) Behavioral and neurobiological changes in C57BL/6 mice exposed to cuprizone. Behav Neurosci 123:418–429
Xu H, Yang HJ, McConomy B, Browning R, Li XM (2010) Behavioral and neurobiological changes in C57BL/6 mouse exposed to cuprizone: effects of antipsychotics. Front Behav Neurosci 4:8
Xu H, Yang HJ, Rose GM, Li XM (2011) Recovery of behavioral changes and compromised white matter in C57BL/6 mice exposed tocuprizone: effects of antipsychotic drugs. Front Behav Neurosci 5:31
Xuan Y, Yan G, Peng H, Wu R, Xu H (2014) Concurrent changes in 1H MRS metabolites and antioxidant enzymes in the brain of C57BL/6 mouse short-termly exposed to cuprizone: possible implications for schizophrenia. Neurochem Int 69:20–27
Xuan Y, Yan G, Wu R, Huang Q, Li XM, Xu H (2015) The cuprizone-induced changes in (1) H-MRS metabolites and oxidative parameters in C57BL/6 mouse brain: Effects of quetiapine. Neurochem Int 90:185–192
Yang HJ, Wang H, Zhang Y, Xiao L, Clough RW, Browning R et al (2009) Region-specific susceptibilities to cuprizone-induced lesions in the mouse forebrain: Implications for the pathophysiology of schizophrenia. Brain Res 1270:121–130
Yao SQ, Li ZZ, Huang QY, Li F, Wang ZW, Augusto E et al (2012) Genetic inactivation of the adenosine A(2A) receptor exacerbates brain damage in mice with experimental autoimmune encephalomyelitis. J Neurochem 123:100–112
Zhang Y, Xu H, Jiang W, Xiao L, Yan B, He J et al (2008) Quetiapine alleviates the cuprizone-induced white matter pathology in the brain of C57BL/6 mouse. Schizophr Res 106:182–191
Zhang Y, Zhang H, Wang L, Jiang W, Xu H, Xiao L et al (2012) Quetiapine enhances oligodendrocyte regeneration and myelin repair after cuprizone-induced demyelination. Schizophr Res 138:8–17
Zhang Q, Li Z, Wu S, Li X, Sang Y, Li J, Niu Y, Ding H (2016) Myricetin alleviates cuprizone-induced behavioral dysfunction and demyelination in mice by Nrf2 pathway. Food Funct 7:4332–4342
Zhang L, Xu S, Huang Q, Xu H (2018) N-acetylcysteine attenuates the cuprizone-induced behavioral changes and oligodendrocyte loss in male C57BL/7 mice via its anti-inflammation actions. J Neurosci Res 96:803–816
Acknowledgments
This work was supported by grants from the National Natural Science Fund (30770771) and the Medical Scientific Research Program of Guangdong, China (A2015242).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Zhang, J., Yang, L., Fang, Z. et al. Adenosine Promotes the Recovery of Mice from the Cuprizone-Induced Behavioral and Morphological Changes while Effecting on Microglia and Inflammatory Cytokines in the Brain. J Neuroimmune Pharmacol 13, 412–425 (2018). https://doi.org/10.1007/s11481-018-9799-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11481-018-9799-0