
Abstract
Skeletal muscle is a major tissue of glucose consumption and plays an important role in glucose homeostasis. Prenylflavonoids, a component of Macaranga tanarius fruits, have been reported to have antioxidant, antibacterial, and anticancer effects. However, the effects of these compounds on skeletal muscle glucose metabolism are unclear. Here, we isolated five prenylflavonoids from M. tanarius fruits, and investigated the mechanism of action of these compounds on skeletal muscle cells using L6 myotubes. We found that isonymphaeol B and 3′-geranyl naringenin increased glucose uptake in a dose-dependent manner. Furthermore, both isonymphaeol B and 3′-geranyl naringenin increased AMPK phosphorylation but did not affect PI3K-Akt phosphorylation. Isonymphaeol B and 3′-geranyl naringenin also increased Glut1 mRNA expression and plasma membrane GLUT1 protein levels. These results suggest that isonymphaeol B and 3′-geranyl naringenin have beneficial effects on glucose metabolism through AMPK and GLUT1 pathway. Isonymphaeol B and 3′-geranyl naringenin may be potential lead candidates for antidiabetic drug development.
Graphical abstract

Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Diabetes is among the top 10 causes of death worldwide. In 2019, the prevalence of diabetes was estimated to be 473 million worldwide and is expected to increase to 578 million by 2030 [1]. Type 2 diabetes is the most common type of diabetes, accounting for around 90% of all diabetes worldwide. Type 2 diabetes mellitus (T2DM) is a metabolic disease characterized by the impairment of insulin-secretion from pancreatic β cells and insulin resistance in peripheral tissues such as liver, adipose tissue, and skeletal muscle [2] causing hyperglycemia which in turn leads to multiple diabetic complications, including nephropathy, neuropathy, retinopathy, ketoacidosis, cardiovascular disease, and hypertension [3], significantly reducing the quality of life.
Skeletal muscle is a major peripheral tissue, accounting for about 40% of the total body mass and more than 30% of total energy expenditure. In addition, over 80% of insulin-stimulated glucose transport is incorporated to skeletal muscle [4]. Any impairment in glucose uptake in skeletal muscle causes hyperglycemia [5]. Therefore, skeletal muscle and insulin plays an important role in maintaining blood-glucose homeostasis.
The uptake of glucose into tissues is mediated to by a carrier protein called glucose transporter (GLUT) 1–12 [6]. In skeletal muscle, GLUT4 is predominantly expressed and regulate glucose uptake in insulin-dependent and -independent pathways [7]. Translocation of intracellular GLUT4 to the cell membrane is important in glucose uptake [8]. On the other hand, GLUT1 is ubiquitously expressed and thought to be responsible for basal glucose uptake [9].
Translocation of GLUT4 into the cell membrane is regulated by two distinct signaling pathways. The first is the insulin-dependent pathway in which phosphatidylinositol-3 kinase (PI3K) is activated by insulin and then downstream protein kinase B (Akt) is activated and increases GLUT4 translocation. The other is the insulin-independent adenosine 5′-monophosphate activated protein kinase (AMPK) pathway, which is a major cellular and whole-body energy sensor and a master regulator of metabolic homeostasis, and this kinase is activated by increases in the AMP/ATP ratio that occurs during energy shortage [10]. AMPK has also been known as an attractive therapeutic target for metabolic disease including obesity and diabetes [11, 12]. Metformin, one of the therapeutic agents for hyperglycemia, increases glucose uptake by activating AMPK in skeletal muscle [13]. It is also reported that AMPK is activated by natural compounds such as resveratrol contained in red wine [14] and curcumin contained in Curcuma longa [15]. Therefore, natural products and food ingredients are excellent sources of lead compounds in the search for new drugs for the prevention and treatment of various metabolic diseases including type 2 diabetes.
We have investigated the pharmacological activity of various plant resources in Okinawa, Japan. Macaranga tanarius is native in Okinawa and known as source of Okinawan propolis [16]. M. tanarius characteristically contains prenylflavonoids such as nymphaeol A (6-geranyl eriodictyol), nymphaeol B (2′-geranyl eriodictyol), nymphaeol C (6-dimethylallyl-2′-geranyl eriodictyol), isonymphaeol B (5′-geranyl eriodictyol) and 3′-geranyl naringenin [17]. These prenylflavonoids have been reported to have various pharmacological activities such as antioxidative [18], antibacterial [19], anti-inflammatory and α-glucosidase inhibitive activity [20]. However, the effect of these prenylflavonoids in glucose metabolism is unknown. In this study, we examined the effects of prenylflavonoids from M. tanarius fruits on glucose uptake in skeletal muscle cells and the mechanism of action of these compounds.
Materials and methods
Materials
Insulin and Na+/K+ ATPase α-1 antibody were purchased from Sigma-Aldrich (St. Louis, MO, USA). Polyclonal antibodies against GLUT1, GLUT4, phospho-Akt (Ser473), Akt, phospho-AMPKα (Thr172), AMPKα, β-actin, horseradish peroxidase (HRP)-conjugated anti-rabbit IgG and HRP-conjugated anti-mouse IgG were purchased from Cell Signaling Technology Inc. (Beverly, MA, USA). Other reagents were obtained from FUJIFILM Wako Pure Chemical Corporation (Osaka, Japan).
Plant material
Macaranga tanarius fruits used in this study was collected at the University of the Ryukyus in Okinawa, Japan., in May 2016. The plant was identified by one of the authors (Y. Saito). A voucher specimen (Saito and Amano 3520, 10 Apr. 2020) has been deposited in the Herbarium of the Faculty of Education (URO), University of the Ryukyus, Okinawa.
Isolation and identification of prenylflavonoids
The dried fruits of M. tanarius (2.2 g) were ground with a mixer and then extracted with ethanol (100.0 mL) for 5 days. The ethanol extract solution was filtered and concentrated to obtain M. tanarius fruits extract (MFE). MFE (215.4 mg) was separated using reverse-phase HPLC with COSMOSIL 5C18-AR-II (20 × 250 mm) at a flow rate of 5.0 ml/min, UV of 215 nm, gradient condition of 75% methanol containing 0.1% trifluoroacetic acid (TFA) at 0–50 min and 100% methanol at 50–70 min. The fraction eluted with 100% methanol was further separated using reverse-phase HPLC with COSMOSIL 5C18-AR-II (20 × 250 mm) at a flow rate of 5.0 ml/min, UV of 215 nm, gradient condition of 90% methanol containing 0.1% TFA at 0–10 min and 100% methanol at 10–20 min. The fraction eluted with 75% methanol containing 0.1% TFA resulted in the isolation of Compound 1 (33.7 mg), 2 (13.6 mg), 3 (43.8 mg) and 4 (19.7 mg), and the fraction eluted with 90% methanol containing 0.1% TFA resulted in the isolation of compound 5 (24.8 mg). The chemical structures of Compound 1 (nymphaeol B), 2 (isonymphaeol B), 3 (nymphaeol A), 4 (3′-geranyl naringenin) and 5 (nymphaeol C) were determined from their 1H NMR and 13C NMR spectra measured in CDCl3, CD3OD and acetone-d6 by comparison with reported data [21,22,23,24] (Fig. 1a). The 1H and 13C NMR spectra were obtained on AVANCE III 500 NMR spectrometer (Bruker BioSpin Corporation, Billerica, MA, USA).

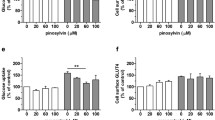
Effects of prenylflavonoids on glucose uptake in L6 myotubes. a Chemical structures of the prenylflavonoids isolated from M. tanarius fruits. b Effects of prenylflavonoids on the viability of L6 myotubes. Cell viability was measured by the MTT assay. c Effects of prenylflavonoids on glucose uptake in L6 myotubes. L6 myotubes were preincubated in glucose-free KHH buffer for 2 h. The cells were then incubated in glucose-containing KHH buffer without or with prenylflavonoids for 24 h, and the glucose uptake was measured as described in Materials and methods. Each value represents the mean ± SD of three independent experiments. *p < 0.05, **p < 0.01 vs Control cells (Con)
Cell culture
L6 rat myoblasts (JCRB Cell Bank, Osaka, Japan) were cultured in Dulbecco’s modified Eagle’s medium (DMEM) (Sigma-Aldrich) supplemented with 10% fetal bovine serum (FBS) (Gibco, BRL, NY) and antibiotics (100 units/mL penicillin and 10 µg/mL streptomycin (Sigma-Aldrich) at 37 ℃ under a humidified 5% CO2 atmosphere. To differentiate myotubes, L6 myoblasts were seeded in 96-well plates (5 × 103 cells/well) or 6-well plates (1 × 105 cells/well) and cultured to confluence for 2 days. Then, the cells were cultured with DMEM supplemented with 0.5% FBS for 9 days. The medium was renewed every 3 days.
Glucose uptake assay
Differentiated L6 myotubes in 96-well plates were washed with PBS and incubated in filter-sterilized glucose-free Krebs–Henseleit-HEPES (KHH) buffer (119 mM NaCl, 4.7 mM KCl, 2.5 mM CaCl2・2H2O, 25 mM NaHCO3, 1.2 mM KH2PO4, 1.2 mM MgSO4, 0.1% bovine serum albumin, 10 mM HEPES, and 2 mM sodium pyruvate, pH 7.4) for 2 h. The cells were then cultured for 24 h in KHH buffer containing 11 mM glucose (glucose-containing KHH buffer) with or without prenylflavonoids. To measure the amount of glucose uptake into the cells, the glucose concentrations in the KHH buffer before and after culture were measured with a microplate reader (Bio Tek, absorbance at 490 nm) using the glucose CII test kit (FUJIFILM Wako). The amounts of glucose uptake were calculated from the differences in glucose concentrations between before and after incubation.
Cell viability
Cell viability was determined using a MTT reagent (Sigma-Aldrich) after the Glucose uptake assay. The absorbance at 570 nm was measured using a microplate reader.
Subcellular fractionation and GLUTs translocation analysis
To assesses the translocation of GLUT4, plasma membrane and cytoplasmic fractions were prepared as described previously [25] with slight modification. L6 myotubes were harvested with buffer A (50 mM Tris–HCl, pH 8.0, 0.5 mM DTT) containing 0.1% (v/v) Nonidet P-40 (NP-40), protease inhibitors (1 mM PMSF, 5 µg/mL leupeptin and 5 µg/mL aprotinin), and a phosphatase inhibitor (10 mM NaF and 1 mM Na3VO4) and homogenized by 10 passages through a 27-gage needle. Each homogenate was centrifuged at 1000 g for 10 min at 4 ℃, and the pellet was resuspended in NP-40-free buffer A. After incubation on ice for 10 min with occasional mixing, the samples were centrifuged at 1000×g for 10 min at 4 ℃. The supernatants from the first and second centrifugations at 1000×g were pooled and centrifuged at 16,000×g for 20 min at 4 ℃. This supernatant was collected as the cytoplasm fraction. The pellet was resuspended in buffer A containing 1% (v/v) NP-40, incubated for 24 h at 4 ℃, and centrifuged at 16,000×g for 20 min at 4 ℃. The supernatant was collected as the plasma membrane fraction.
Western blotting analysis
L6 myotubes cultured in a 6-well plate were washed with PBS and then incubated with glucose-free KHH buffer for 2 h. Next, glucose-free KHH buffer was removed and incubated with glucose-containing KHH buffer for 2 h. The cells were then cultured for prescribed time in glucose-containing KHH buffer without or with prenylflavonoids. The cells were washed with ice-cold PBS and solubilized in 1 × SDS Loading Buffer (Cell signaling) to obtain total cell lysate. The lysates were sonicated for 10 s, boiled at 100 ℃ for 10 min and centrifuged at 15,000 rpm for 10 min. The protein concentrations of the supernatants were measured using RC DC ™ Protein Assay kit (Bio-Rad laboratories Inc., Hercules, CA, USA). Equal amounts of protein (20 µg per lane) were loaded onto 10% Mini-PROTEAN TGX precast gels (Bio-Rad), separated by electrophoresis, and transferred to nitrocellulose membranes. The membranes were washed with Tris buffered saline (TBS) and blocked with 5% BSA in TBS containing 1% Tween 20 (TBS-T) for 1 h. After the blocking, the membranes were washed in TBS-T and incubated with anti-phospho-AMPK, anti-AMPK, anti-phospho-Akt, anti-Akt, anti-β-actin, anti-GLUT1, anti-GLUT4 or anti-Na+/K+ ATPase antibodies overnight at 4℃. The membranes were then washed with TBS-T and incubated with HRP-conjugated anti-rabbit IgG or HRP-conjugated anti-mouse IgG at room temperature for 1 h. Immunoreactive bands were detected using Clarity Western ECL Substrate (Bio-Rad). The intensity of each band was analyzed with a ChemiDoc XRS Plus system coupled with Image Lab software (Bio-Rad).
Real-time reverse transcription-polymerase chain reaction (RT-PCR)
L6 myotubes cultured in a 6-well plate were subjected to total RNA isolation using RNeasy plus mini kit (QIAGEN). The total RNA was reverse transcribed into cDNA using the ReverTra Ace qPCR RT kit (TOYOBO, Osaka, Japan) according to the manufacturer's instructions. Real-time PCR was performed with FastStart SYBR green Master Mix (Roche Diagnostics, Mannheim, Germany) in a StepOnePlus System (Applied Biosystems, USA). The relative expression levels of the target genes against the endogenous reference gene β-actin was calculated using the delta cycle threshold method. The primer sequences are listed in Table 1.
Statistical analysis
All the data are presented as mean ± standard deviation. One-way analysis of variance (ANOVA) followed by Dunnett’s test was used to determine the statistical significance of the differences between the experimental groups. Statistical significance was assumed at p < 0.05, p < 0.01.
Results
Effect of prenylflavonoids from M. tanarius fruits on glucose uptake in L6 myotubes
First, cytotoxicity of prenylflavonoids was assessed by MTT assay. Nymphaeol C decreased cell viability only at highest concentration (30 µM). In contrast, nymphaeol A, nymphaeol B, isonymphaeol B and 3′-geranyl naringenin did not show cytotoxicity up to 30 µM (Fig. 1b). To investigate the effect of prenylflavonoids on glucose metabolism, glucose uptake was measured in L6 myotubes. Nymphaeol A, nymphaeol B and nymphaeol C increased glucose uptake at a concentration of 30 µM. Isonymphaeol B and 3′-geranyl naringenin increased glucose uptake of L6 myotubes in a concentration-dependent manner (Fig. 1c). These results suggest that prenylflavonoids from M. tanarius have a stimulatory effect on glucose uptake with the highest activity shown by isonymphaeol B.
Effects of isonymphaeol B and 3′-geranyl naringenin on PI3K/Akt and AMPK signaling molecules
To study the mechanism of action of prenylflavonoids in the upregulation of glucose uptake, we investigated whether isonymphaeol B and 3′-geranyl naringenin affect insulin PI3K/Akt pathway or noninsulin AMPK pathways by Western blot analysis. Insulin increased the amount of phosphorylated Akt in L6 myotubes, whereas isonymphaeol B and 3′-geranyl naringenin had no effect on phosphorylated Akt (Fig. 2a). In contrast, 5-aminoimidazole-4-carboxamide ribonucleotide (AICAR), an AMPK activator used as a positive control, significantly increased the amount of phosphorylated AMPK. Isonymphaeol B and 3′-geranyl naringenin also increased phosphorylated AMPK in a time-dependent manner (Fig. 2b). These results suggest that isonymphaeol B and 3′-geranyl naringenin activate AMPK pathway but not PI3K/Akt pathway.

Effects of isonymphaeol B and 3′-geranyl naringenin on the phosphorylation of Akt and AMPK in L6 myotubes. a, b Cells were incubated for the indicated times with isonymphaeol B (Iso-B) and 3′-geranyl naringenin (3′-Ger) and the cell lysates were subjected to Western blotting. Each value represents the mean ± SD of three independent experiments. *p < 0.05, **p < 0.01 vs non-treatment cell (−)
Furthermore, to reveal whether the activation of the AMPK pathway is involved in isonymphaeol B and 3′-geranyl naringenin-increased glucose uptake, we examined the effects on isonymphaeol B and 3′-geranyl naringenin-increased glucose uptake using AMPK inhibitor (dorsomorphin). Dorsomorphin partly decreased isonymphaeol B and 3′-geranyl naringenin-increased glucose uptake (Fig. 3). These results suggest that the activation of AMPK contributes to isonymphaeol B and 3′-geranyl naringenin-increased glucose uptake.

Effects of AMPK inhibitor on isonymphaeol B and 3′-geranyl naringenin increased glucose uptake. L6 myotubes were preincubated in glucose-free KHH buffer without or with dorsomorphin (AMPK inhibitor) for 2 h. The cells were then incubated in glucose-containing KHH buffer without or with dorsomorphin (AMPK inhibitor) in presence of isonymphaeol B (Iso-B) and 3′-geranyl naringenin (3′-Ger) for 24 h, and the glucose uptake was measured as described in Materials and methods. Each value represents the mean ± SD of three independent experiments. *p < 0.05, **p < 0.01 vs non-treatment cells (−)
Effects of isonymphaeol B and 3′-geranyl naringenin on GLUT1 and GLUT4 in L6 myotubes
GLUT1 and GLUT4 is the major glucose transporters in muscle. The amount of glucose transporters in plasma membrane is important in the activity of glucose uptake. We prepared protein sample of the plasma membrane and cytoplasmic fraction from L6 myotubes and the expression levels of GLUT1 and GLUT4 proteins were determined by Western blot analysis. Na+/K+ ATPase and β-actin were used as loading control proteins for the plasma membrane fraction and the cytoplasmic fraction, respectively. In the plasma membrane fraction, isonymphaeol B and 3′-geranyl naringenin significantly increased the expression level of GLUT1 protein as compared with control. On the other hand, the expression level of GLUT4 protein did not significantly increased as compared with the control. In addition, isonymphaeol B and 3′-geranyl naringenin significantly increased the expression level of Na+/K+ ATPase protein in the cell membrane fraction as compared to control (Fig. 4a and b). In the cytoplasmic fraction, isonymphaeol B and 3′-geranyl naringenin did not significantly change the protein expression levels of GLUT1 and GLUT4 as compared with the control (Fig. 5a and b). Next, we investigated the effects of isonymphaeol B and 3′-geranyl naringenin on Glut1 and Glut4 mRNA expression in L6 myotubes. Isonymphaeol B and 3′-geranyl naringenin increased Glut1 mRNA expression (Fig. 6a). On the other hand, Glut4 mRNA expression was increased by isonympaheol B but not 3′-geranyl naringenin (Fig. 6b). From these results, it is considered that isonymphaeol B and 3′-geranyl naringenin increased GLUT1 protein by increasing Glut1 mRNA expression, and as a result, the amount of GLUT1 protein on the cell membrane also increased.

Effects of isonymphaeol B and 3′-geranyl naringenin on the protein expression of GLUT1 and GLUT4 in the plasma membrane of L6 myotubes. a, b Cells were cultured for the indicated times with isonymphaeol B (Iso-B) and 3′-geranyl naringenin (3′-Ger). The plasma membrane fraction was subjected to Western blotting and GLUT1, GLUT4 and Na+/K+ ATPase were detected. Each value represents the mean ± SD of three independent experiments. *p < 0.05, **p < 0.01 vs non-treatment cells (−)

Effects of isonymphaeol B and 3′-geranyl naringenin on the protein expression of GLUT1 and GLUT4 in the cytoplasm of L6 myotubes. a, b Cells were cultured for the indicated times with isonymphaeol B (Iso-B) and 3′-geranylnaringenin (3′-Ger). The cytoplasm fraction was subjected to Western blotting and GLUT1, GLUT4 and β-actin were detected. Each value represents the mean ± SD of three independent experiments. *p < 0.05, **p < 0.01 vs non-treatment cells (−)

Effects of isonymphaeol B and 3′-geranyl naringenin on the mRNA expression of Glut1 and Glut4 in L6 myotubes. (a, b) Cells were cultured for the indicated times with isonymphaeol B (30 µM) and 3′-geranyl naringenin (30 µM). Total RNA was isolated and reverse transcribed to cDNA for RT-PCR analysis. The expression levels of each mRNA were normalized to the level of β-actin. Each value represents the mean ± SD of three independent experiments. *p < 0.05, **p < 0.01 vs Control cells (Control)
Discussion
Many flavonoids have been reported to have antidiabetic effects [26]. In this study, we focused on prenylflavonoids from M. tanarius, and investigated the antidiabetic effects of these prenylflavonoids. We first found that prenylflavonoids isolated from M. tanarius fruits increased glucose uptake in L6 myotubes. In particular, isonymphaeol B and 3′-geranyl naringenin promoted glucose uptake at lower concentrations in a concentration-dependent manner compared to the others. These results indicated that isonymphaeol B and 3′-geranyl naringenin might exert some metabolic roles as glucose uptake enhancers within skeletal muscles.
Prenyl groups can affect the cellular uptake and biological function of flavonoids by increasing the hydrophobicity of flavonoids and facilitating interaction with the phospholipid bilayers of biological membranes [27, 28]. In fact, 8-prenyl quercetin and 8-prenyl naringenin have been reported to accumulate more in cells and tissues than quercetin and naringenin [29, 30]. In addition, prenylation has been reported to enhance the estrogenic activity in naringenin and genistein [31]. The five prenylflavonoids isolated from M. tanarius are prenylated derivatives of eriodictyol and naringenin. The theoretical value of the Octanol/Water partition coefficient (log Pow), which are an indicator of the hydrophobicity of a compound, is higher in nymphaeol A (logP = 5.8), nymphaeol B (logP = 5.8), nymphaeol C (logP = 7.7), isonymphaeol B (logP = 5.8) and 3′-geranyl naringenin (logP = 6.4) than eriodictyol (logP = 2.0) and naringenin (logP = 2.4). Eriodictyol and naringenin have been reported to increase glucose uptake in adipocytes [32] or skeletal muscle [33]. We have confirmed that isonymphaeol B and 3′-geranyl naringenin increase glucose uptake in L6 myotubes at lower concentrations compared to eriodictyol and naringenin (data not shown). In addition, isonymphaeol B showed a higher glucose uptake effect on L6 myotubes than nymphaeol A, nymphaeol B and nymphaeol C. These results provide that the presence and location of prenyl groups in flavonoids may affect glucose metabolism in skeletal muscle.
Moreover, since only nymphaeol C showed cytotoxicity at 30 μM, it is possible that the dimethylallyl group at the 6-position is involved in the expression of cytotoxic activity. Nymphaeol C has been reported to activate caspase 3 involved in the induction of cell apoptosis more strongly than nymphaeol A, nymphaeol B and isonymphaeol B [34]. Therefore, nymphaeol C may have shown cytotoxicity by inducing apoptosis in L6 myotubes.
Glucose transport in skeletal muscle is mediated by GLUT1 and GLUT4. GLUT1 is localized primarily at the plasma membrane and, is thought to participate mainly in basal glucose transport in skeletal muscle [9]. The activation of GLUT1 is mainly occurred by the cell stressors such as azide [35, 36], osmotic stress [37, 38], methylene blue [39] and glucose deprivation [40, 41]. Among them, the activation of GLUT1 by hypoxia or azide has been attributed to activation of AMPK [38, 42, 43]. Studies of cultured human skeletal muscle cells have reported a decrease in basal glucose uptake and GLUT1 protein levels in cells from patients with type II diabetes [44]. In contradistinction to GLUT1, under basal conditions, GLUT4 is localized mainly in intracellular vesicles and is translocated to the plasma membrane in response to insulin. In particular, GLUT4 is responsible for most of the glucose transport from blood following a meal. GLUT4 translocation to the plasma membrane is mainly regulated by insulin-dependent and -independent signal pathways. The insulin-dependent signaling pathway induces GLUT4 translocation by activating PI3K and downstream Akt phosphorylation when insulin binds to the insulin receptor [45]. On the other hand, the insulin-independent signaling pathway induces translocation of GLUT4 to the plasma membrane through activation of AMPK [46]. AMPK is activated by an increase in the AMP/ATP ratio due to exercise and muscle contraction, adiponectin secreted by adipocytes [47], and liver kinase B1 (LKB1) and Ca2+/calmodulin-dependent kinase β (CaMKKβ) upstream of AMPK [48]. It was suggested that isonymphaeol B and 3′-geranyl naringenin did not affect PI3K phosphorylation, but increased phosphorylation of AMPK, leading to insulin-independent activation of AMPK. It has been reported that naringenin also induced AMPK activation in L6 myotubes without affecting PI3K, and it is possible that isonymphaeol B and 3′-geranyl naringenin promotes glucose uptake by a similar mechanism [33]. To further investigate the mechanism of AMPK activation, it is necessary to investigate the effects of isonymphaeol B and 3′-geranyl naringenin on LKB1, CaMKKβ, and AMP/ATP ratio.
We measured Na+/K+ ATPase as a loading control in plasma membrane fraction analysis. However, Na+/K+ ATPase protein has been reported to increase with activation of AMPK in L6 myotube cells [49]. Therefore, it is suggested that isonymphaeol B and 3′-geranyl naringenin also increased Na+/K+ ATPase protein through activation of AMPK. Isonymphaeol B and 3′-geranyl naringenin enhanced GLUT1 protein in the plasma membrane of L6 myotubes. Thioredoxin-interacting protein (TXNIP) reduces glucose uptake by promoting GLUT1 endocytosis and suppressing Glut1 mRNA expression [50]. However, it has been reported that phosphorylation of TXNIP by activated AMPK promotes the degradation of TXNIP, resulting in an increase GLUT1 protein in plasma membrane, Glut1 mRNA expression and glucose uptake [50]. Therefore, isonymphaeol B and 3′-geranyl naringenin may have increased GLUT1 protein in plasma membrane and Glut1 mRNA expression by promoting TXNIP degradation via AMPK activation. On the other hand, isonymphaeol B and 3′-geranyl naringenin did not increase GLUT4 protein in plasma membrane. Interestingly, although isonymphaeol B increased Glut4 mRNA expression, it did not increase GLUT4 protein in plasma membrane. Therefore, it is necessary to further investigate the mechanism by which isonymphaeol B and 3′-geranyl naringenin selectively increase GLUT1 protein in plasma membrane. In addition, Na+/K+ ATPase is known to regulate glucose transport via sodium glucose cotransporters (SGLTs) according to the established sodium gradient [51]. Therefore, it is necessary to verify whether SGLT1 expressed in skeletal muscle [52] is involved in the glucose uptake increasing effect of isonymphaeol B and 3′-geranyl naringenin. These results suggest that isonymphaeol B and 3′-geranyl naringenin take up glucose into the cell via GLUT1.
In conclusion, we demonstrated that isonymphaeol B and 3′-geranyl naringenin isolated from M. tanarius fruits increase glucose uptake, expression of Glut1 mRNA and plasma membrane GLUT1 protein in muscle cells by activating the AMPK pathway. We propose that isonymphaeol B and 3′-geranyl naringenin and material (M. tanarius fruits and Okinawan propolis) containing these components are potential lead candidates for the development of antidiabetic drugs.
References
International Diabetes Federation (2019) IDF Diabetes Atlas, 9th edn. International Diabetes Federation, Brussels, Belgium
Kahn CR (1994) Banting Lecture. Insulin action, diabetogenes, and the cause of type II diabetes. Diabetes 43:1066–1084
UK Prospective Diabetes Study Group (1998) Intensive blood-glucose control with sulphonylureas or insulin compared with conventional treatment and risk of complications in patients with type 2 diabetes (UKPDS 33). Lancet 352:837–853
Smith AG, Muscat GE (2005) Skeletal muscle and nuclear hormone receptors: implications for cardiovascular and metabolic disease. Int J Biochem Cell Biol 37:2047–2063
DeFronzo RA, Gunnarsson R, Björkman O, Olsson M, Wahren J (1985) Effects of insulin on peripheral and splanchnic glucose metabolism in noninsulin-dependent (type II) diabetes mellitus. J Clin Invest 76:149–155
Joost HG, Thorens B (2001) The extended GLUT-family of sugar/polyol transport facilitators: nomenclature, sequence characteristics, and potential function of its novel members. Mol Membr Biol 18:247–256
Zaid H, Antonescu CN, Randhawa VK, Klip A (2008) Insulin action on glucose transporters through molecular switches, tracks and tethers. Biochem J 413:201–215
Minokoshi Y, Kahn CR, Kahn BB (2003) Tissue-specific ablation of the GLUT4 glucose transporter or the insulin receptor challenges assumptions about insulin action and glucose homeostasis. J Biol Chem 278:33609–33612
Olson AL, Pessin JE (1996) Structure, function, and regulation of the mammalian facilitative glucose transporter gene family. Annu Rev Nutr 16:235–256
Hardie DG (2008) AMPK: a key regulator of energy balance in the single cell and the whole organism. Int J Obes 32:S7–S12
Hwang JT, Kwon DY, Yoon SH (2009) AMP-activated protein kinase: a potential target for the diseases prevention by natural occurring polyphenols. N Biotechnol 26:17–22
Fogarty S, Hardie DG (2010) Development of protein kinase activators: AMPK as a target in metabolic disorders and cancer. Biochim Biophys Acta 1804:581–591
Song Y, Shi J, Wu Y, Han C, Zou J, Shi Y, Liu Z (2014) Metformin ameliorates insulin resistance in L6 rat skeletal muscle cells through upregulation of SIRT3. Chin Med J 127:1523–1529
Breen DM, Sanli T, Giacca A, Tsiani E (2008) Stimulation of muscle cell glucose uptake by resveratrol through sirtuins and AMPK. Biochem Biophys Res Commun 374:117–122
Na LX, Zhang YL, Li Y, Liu LY, Li R, Kong T, Sun CH (2011) Curcumin improves insulin resistance in skeletal muscle of rats. Nutr Metab Cardiovasc Dis 21:526–533
Kumazawa S, Nakamura J, Murase M, Miyagawa M, Ahn MR, Fukumoto S (2008) Plant origin of Okinawan propolis: honeybee behavior observation and phytochemical analysis. Naturwissenschaften 95:781–786
Kumazawa S, Murase M, Momose N, Fukumoto S (2014) Analysis of antioxidant prenylflavonoids in different parts of Macaranga tanarius, the plant origin of Okinawan propolis. Asian Pac J Trop Med 7:16–20
Kumazawa S, Ueda R, Hamasaka T, Fukumoto S, Fujimoto T, Nakayama T (2007) Antioxidant prenylated flavonoids from propolis collected in Okinawa, Japan. J Agric Food Chem 55:7722–7725
Lee JH, Kim YG, Khadke SK, Yamano A, Woo JT, Lee J (2019) Antimicrobial and antibiofilm activities of prenylated flavanones from Macaranga tanarius. Phytomedicine 63:153033
Shahinozzaman M, Taira N, Ishii T, Halim MA, Hossain MA, Tawata S (2018) Anti-inflammatory, anti-diabetic, and anti-alzheimer’s effects of prenylated flavonoids from okinawa propolis: an investigation by experimental and computational studies. Molecules 23:2479
Chen CN, Wu CL, Lin JK (2004) Propolin C from propolis induces apoptosis through activating caspases, Bid and cytochrome c release in human melanoma cells. Biochem Pharmacol 67:53–66
Chen CN, Weng MS, Wu CL, Lin JK (2004) Comparison of radical scavenging activity, cytotoxic effects and apoptosis induction in human melanoma cells by taiwanese propolis from different sources. Evid Based Complement Alternat Med 1:175–185
Yakushijin K, Shibayama K, Murata H, Furukawa H (1980) New prenylflavanones from Hernandia nymphaefolia (Presl) Kubitzki. Heterocycles 14:397–402
Jayasinghe L, Rupasinghe GK, Hara N, Fujimoto Y (2006) Geranylated phenolic constituents from the fruits of Artocarpus nobilis. Phytochemistry 67:1353–1358
Nishiumi S, Ashida H (2007) Rapid preparation of a plasma membrane fraction from adipocytes and muscle cells: application to detection of translocated glucose transporter 4 on the plasma membrane. Biosci Biotechnol Biochem 71:2343–2346
Al-Ishaq RK, Abotaleb M, Kubatka P, Kajo K, Büsselberg D (2019) Flavonoids and their anti-diabetic effects: cellular mechanisms and effects to improve blood sugar levels. Biomolecules 9:E430
Hendrich AB, Malon R, Pola A, Shirataki Y, Motohashi N, Michalak K (2002) Differential interaction of Sophora isoflavonoids with lipid bilayers. Eur J Pharm Sci 16:201–208
Tammela P, Laitinen L, Galkin A, Wennberg T, Heczko R, Vuorela H, Slotte JP, Vuorela P (2004) Permeability characteristics and membrane affinity of flavonoids and alkyl gallates in Caco-2 cells and in phospholipid vesicles. Arch Biochem Biophys 425:193–199
Mukai R, Fujikura Y, Murota K, Uehara M, Minekawa S, Matsui N, Kawamura T, Nemoto H, Terao J (2013) Prenylation enhances quercetin uptake and reduces efflux in Caco-2 cells and enhances tissue accumulation in mice fed long-term. J Nutr 143:1558–1564
Mukai R, Horikawa H, Fujikura Y, Kawamura T, Nemoto H, Nikawa T, Terao J (2012) Prevention of disuse muscle atrophy by dietary ingestion of 8-prenylnaringenin in denervated mice. PLoS ONE 7:e45048
Kretzschmar G, Zierau O, Wober J, Tischer S, Metz P, Vollmer G (2010) Prenylation has a compound specific effect on the estrogenicity of naringenin and genistein. J Steroid Biochem Mol Biol 118:1–6
Zhang WY, Lee JJ, Kim Y, Kim IS, Han JH, Lee SG, Ahn MJ, Jung SH, Myung CS (2012) Effect of eriodictyol on glucose uptake and insulin resistance in vitro. J Agric Food Chem 60:7652–7658
Zygmunt K, Faubert B, MacNeil J, Tsiani E (2010) Naringenin, a citrus flavonoid, increases muscle cell glucose uptake via AMPK. Biochem Biophys Res Commun 398:178–183
Huang WJ, Huang CH, Wu CL, Lin JK, Chen YW, Lin CL, Chuang SE, Huang CY, Chen CN (2007) Propolin G, a prenylflavanone, isolated from Taiwanese propolis, induces caspase-dependent apoptosis in brain cancer cells. J Agric Food Chem 55:7366–7376
Shetty M, Loeb JN, Vikstrom K, Ismail-Beigi F (1993) Rapid activation of GLUT-1 glucose transporter following inhibition of oxidative phosphorylation in clone 9 cells. J Biol Chem 268:17225–17232
Rubin D, Ismail-Beigi F (2003) Distribution of Glut1 in detergent-resistant membranes (DRMs) and non-DRM domains: effect of treatment with azide. Am J Physiol Cell Physiol 285:C377–C383
Barros LF, Barnes K, Ingram JC, Castro J, Porras OH, Baldwin SA (2001) Hyperosmotic shock induces both activation and translocation of glucose transporters in mammalian cells. Pflugers Arch 442:614–621
Barnes K, Ingram JC, Porras OH, Barros LF, Hudson ER, Fryer LG, Foufelle F, Carling D, Hardie DG, Baldwin SA (2002) Activation of GLUT1 by metabolic and osmotic stress: potential involvement of AMP-activated protein kinase (AMPK). J Cell Sci 115:2433–2442
Louters LL, Dyste SG, Frieswyk D, Tenharmsel A, Vander Kooy TO, Walters L, Whalen T (2006) Methylene blue stimulates 2-deoxyglucose uptake in L929 fibroblast cells. Life Sci 78:586–591
Kumar A, Xiao YP, Laipis PJ, Fletcher BS, Frost SC (2004) Glucose deprivation enhances targeting of GLUT1 to lipid rafts in 3T3-L1 adipocytes. Am J Physiol Endocrinol Metab 286:E568–E576
Roelofs B, Tidball A, Lindborg AE, TenHarmsel A, Vander Kooy TO, Louters LL (2006) Acute activation of glucose uptake by glucose deprivation in L929 fibroblast cells. Biochimie 88:1941–1946
Jing M, Ismail-Beigi F (2007) Critical role of 5’-AMP-activated protein kinase in the stimulation of glucose transport in response to inhibition of oxidative phosphorylation. Am J Physiol Cell Physiol 292:C477–C487
Jing M, Cheruvu VK, Ismail-Beigi F (2008) Stimulation of glucose transport in response to activation of distinct AMPK signaling pathways. Am J Physiol Cell Physiol 295:C1071–C1082
Ciaraldi TP, Mudaliar S, Barzin A, Macievic JA, Edelman SV, Park KS, Henry RR (2005) Skeletal muscle GLUT1 transporter protein expression and basal leg glucose uptake are reduced in type 2 diabetes. J Clin Endocrinol Metab 90:352–358
Farese RV, Sajan MP, Standaert ML (2005) Insulin-sensitive protein kinases (atypical protein kinase C and protein kinase B/Akt): actions and defects in obesity and type II diabetes. Exp Biol Med 230:593–605
Zhou G, Myers R, Li Y, Chen Y, Shen X, Fenyk-Melody J, Wu M, Ventre J, Doebber T, Fujii N, Musi N, Hirshman MF, Goodyear LJ, Moller DE (2001) Role of AMP-activated protein kinase in mechanism of metformin action. J Clin Invest 108:1167–1174
Fang H, Judd RL (2018) Adiponectin Regulation and Function. Compr Physiol 8:1031–1063
Srivastava RA, Pinkosky SL, Filippov S, Hanselman JC, Cramer CT, Newton RS (2012) AMP-activated protein kinase: an emerging drug target to regulate imbalances in lipid and carbohydrate metabolism to treat cardio-metabolic diseases. J Lipid Res 53:2490–2514
Benziane B, Björnholm M, Pirkmajer S, Austin RL, Kotova O, Viollet B, Zierath JR, Chibalin AV (2012) Activation of AMP-activated protein kinase stimulates Na+, K+-ATPase activity in skeletal muscle cells. J Biol Chem 287:23451–23463
Wu N, Zheng B, Shaywitz A, Dagon Y, Tower C, Bellinger G, Shen CH, Wen J, Asara J, McGraw TE, Kahn BB, Cantley LC (2013) AMPK-dependent degradation of TXNIP upon energy stress leads to enhanced glucose uptake via GLUT1. Mol Cell 49:1167–1175
Wright EM (2001) Renal Na(+)-glucose cotransporters. Am J Physiol Renal Physiol 280:F10–F18
Chen J, Williams S, Ho S, Loraine H, Hagan D, Whaley JM, Feder JN (2010) Quantitative PCR tissue expression profiling of the human SGLT2 gene and related family members. Diabetes Ther 1:57–92
Author information
Authors and Affiliations
Corresponding authors
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Natsume, N., Yonezawa, T., Saito, Y. et al. Prenylflavonoids from fruit of Macaranga tanarius promote glucose uptake via AMPK activation in L6 myotubes. J Nat Med 75, 813–823 (2021). https://doi.org/10.1007/s11418-021-01517-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11418-021-01517-x