
Abstract
Purpose
The water-level-fluctuating zone (WLFZ) is the buffer zone of energy and material exchange between terrestrial and aquatic ecosystems. Artificial vegetation restoration of WLFZ can improve the interception capacity of P pollution. The purpose of this study is to explore the effect of artificial vegetation restoration on the bioavailability of soil phosphorus (P) in the WLFZ.
Material and methods
Soil samples from different spatial locations (natural vegetation zone, artificial vegetation restoration zone) and different altitudes of the WLFZ were collected in the Three Gorges Reservoir (TGR) region, Chongqing, China. Soil P fraction, microbial biomass P (MBP), and phosphatase activity were measured.
Results and discussion
Artificial vegetation restoration changed the spatial distribution patterns of soil bioavailable P (Bio-P) in the WLFZ. The soil bioavailable inorganic P (Bio-Pi) in the artificial vegetation restoration zone was significantly higher than those at the natural vegetation zone (p < 0.05) and its content decreased with the decrease of altitude. The content of bioavailable organic P (Bio-Po) in the two transects was not significantly different in general, but was different at different altitudes. Phosphodiesterase (PDE) activity was negatively correlated with Bio-Po in artificial vegetation restoration zone (p < 0.01, R2 = 0.21), but significantly positively correlated with in natural vegetation zone (p < 0.05, R2 = 0.17); this suggests that the relationship between Bio-Po and PDE activity was altered by vegetation restoration. Moreover, the factors controlling the bioavailability of P in the WLFZ are discussed.
Conclusion
Artificial vegetation restoration and altitude are the control factors of soil P fractions and bioavailability in WLFZ. Vegetation restoration can increase soil TP and Bio-Pi in general but has little effect on Bio-Po.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
The water-level-fluctuating zone (WLFZ) is a transition zone between aquatic and terrestrial ecosystems and is also an active zone for elemental transport and transformation (Hill et al. 2000; Li et al. 2019). The WLFZ plays an important role in controlling soil erosion and relieving ecological stress (Hale et al. 2007; Cheng et al. 2015). Water level fluctuation affects not only the physicochemical properties of the soils in the zone, but also the transport and deposition of materials in the water column, thus affecting the biogeochemical cycling of elements at the water-land interface (Tang et al. 2014; Evtimova et al. 2016; Zhu et al. 2022).
Phosphorus (P) regulates the primary productivity of terrestrial and aquatic ecosystems and has an important impact on ecosystem functioning (Lembi et al. 2001; Cleveland et al. 2011). Studies on agricultural, forest, grassland, and wetland ecosystems have shown that drying and rewetting (DRW) conditions altered soil P fractions and increased the amount of labile P in the soil, providing nutrients to vegetation but increasing the risk of its loss with runoff (Bünemann et al. 2013; Brödlin et al. 2019; Khan et al. 2022). Due to the frequent DRW cycle in the WLFZ of the reservoir after dam construction, the contents of labile P in the soil increase (Wang et al. 2021). DRW causes structural disruption of soil aggregates, releasing large amounts of molybdate reactive phosphorus and molybdate unreactive phosphorus bound to the soil matrix (Bünemann et al. 2013). Drastic changes in intracellular osmotic pressure after soil DRW will lead to the death of microorganisms and the release of P from the microorganisms as solutes in organic (Po) and inorganic (Pi) form, leading to an increase in labile P (Birch 1958; Turner et al. 2003).
The development of hydropower in the Yangtze River basin has led to significant impacts on the water ecosystem (Cheng et al. 2015; Wang et al. 2018, 2020b; Shen et al. 2022). To meet the needs of flood control, power generation, and navigation, the unique scheduling mechanism of the Three Gorges Reservoir (TGR) creates a WLFZ of up to 30 m during the rainy season when flood discharge is maintained at low water levels and during the dry season when water storage is maintained at high water levels (Zhang et al. 2012; Tang et al. 2018). As a result of the anti-seasonal cycle of DRW, the original vegetation in the zone dies due to long-term flooding and habitat transformation, posing a serious threat to the ecology of the reservoir area (Wu et al. 2004). In order to prevent soil erosion and maintain ecosystem stability, extensive vegetation restoration has been carried out in the WLFZ of TGR. The natural vegetation zone vegetation is dominated by herbaceous plants, and the artificial vegetation restoration zone vegetation consists of herbs, trees, and shrubs (Ye et al. 2014). Previous studies have shown that soil microbial biomass of C, N, and P in the WLFZ is significantly increased after vegetation restoration, and that vegetation type has a significant impact on microbial load, usually grassland > cropland and woodland > bare ground (Yang et al. 2017).
The WLFZ is sensitive to natural and human activities and becomes the hotspot of global P cycling (Jeppesen et al. 2009; Vidon et al. 2010). The spatial and temporal distribution of P in the WLFZ of TGR has been extensively studied, but its variability pattern remains controversial (Zhang et al. 2012; Wu et al. 2016; Ye et al. 2019). For example, Wu et al. (2016) suggest that the bioavailable P (Bio-P) in soils in the permanent backwater zone (near the dam) is higher than in the fluctuant backwater zone (away from the dam). However, Ye et al. (2019) found that Bio-P had higher concentration levels in soils in the midstream WLFZ of the reservoir area. Due to periodic reservoir scheduling, soils in the higher altitude WLFZ take about 125 days longer to fall dry than those at lower altitudes. The study on Pengxi River, a tributary of TGR, showed that Bio-P increased with the decrease of altitude, and MBP, OM, and Fe oxides were the main factors affecting Bio-P in the WLFZ (Wang et al. 2020b). Previous studies have focused on natural vegetation zone, and fewer studies have been conducted on the quality changes of artificially restored soils, especially the effect of artificial vegetation restoration on the bioavailability of P in WLFZ soils has rarely been reported.
The long-term anti-seasonal operation of the TGR has led to the degradation of the structure and function of the ecosystem in the WLFZ and thus causing a series of ecological problems, such as the degradation of the original vegetation, the change of soil physical and chemical properties, and the loss of water and soil. In recent years, with the development of many kinds of vegetation restoration measures in the WLFZ of the TGR area, the habitat condition of WLFZ has been well improved, but P pollution is still a major water environmental and ecological problem in the basin (Xue et al. 2020). Therefore, it is of great significance and necessity for deepen understanding the dynamic changes of soil P fractions in the WLFZ. Take two typical transects in the TGR area as examples: natural vegetation zone, artificial vegetation restoration zone; additionally, combined different altitudes and soil depths, soil P was fractioned to study its bioavailability.
Here, we made the following hypotheses: (1) artificial vegetation restoration would influence the change of soil P fraction and then affect its bioavailability, and (2) vegetation restoration could affect soil phosphatase activity and thus affect P cycle.
2 Materials and methods
2.1 Study sites and sampling
The study site is located in the WLFZ on the right bank of the Yangtze River in Qingxi Town, Fuling, Chongqing (29° 48′ N, 107° 28′ E).The climate is humid subtropical monsoon, with an average temperature of 18.1 °C and an average precipitation of 1072 mm. The regional soil type is mainly yellow brown soil (Ye et al. 2014). Two representative transects were selected: (1) artificial vegetation restoration zone and (2) natural vegetation zone. There are little effects of human activities on the natural vegetation zone, and the dominant species of plants are the annual plant Echinochloa crus-galli, perennials Acorus gramineus Soland, and Cynodon dactylon. The artificial vegetation restoration zone is about 5 km away from the natural vegetation zone. At the altitude of 165–175 m a.s.l., where mainly planted native plants, such as Morus alba and Salix matsudana Koidz. The regions of below the 165 m a.s.l., the submersion time is relatively long, approximately 7–8 months of each year. Here, annual Polypogon fugax and Cynodon dactylon were planted. The WLFZ is relatively flat above 170 m a.s.l., the slope of the middle and lower part is 20° and 25°, and the lighting conditions and flooding gradient are basically the same.
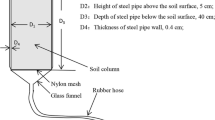
In the summer of 2022, July (during the low water level of the TGR) soil samples were collected along each transect at different depths (0–10, 10–30, 30–50 cm) at four altitudes of 145, 155, 165, and 175 m a.s.l (Fig. 1). Within each plot, three 1 m × 1 m soil sampling subplots were made and then mixed into one composite sample, totaling 24 samples. Soil samples were collected using foil sampler. The specific operation is inserting the foil sampler into the soil and extracting intact cores from sides of the quantitative soil pits. Fresh soil samples were sealed in polythene bags and stored at 4 °C and transported to the laboratory immediately.

The sampling sites (red and purple dots) in the water-level-fluctuation zone of Three Gorges Reservoir, China
2.2 Laboratory analysis
Soil pH was measured from suspension (soil:water, 1:2.5) using a combination pH electrode. The bulk weight (BD) was determined by the cutting ring method. Soil particle size was measured using a laser particle size analyzer (Mastersizer 2000; Malvern, UK); each sample was pretreated with 10% H2O2 and 10% HCl to remove organic matter and carbonate. The particle size of the samples was summarized as coarse sand (200–2000 μm), fine sand (20–200 μm), silt (2–20 μm), and clay (0.0–2 μm). Soil moisture content (MC) was measured by drying at 105 °C to a constant weight. Soil porosity is determined using the pycnometer method, soil porosity (%) = (1 − BD soil specific gravity−1) × 100.
Referring to the kit instruction offered by Shanghai Enzyme-linked Biotechnology Co., Ltd, the activity of phosphatase activity was measured. The activity of alkaline phosphatase (ALP), acid phosphatase (ACP), phosphodiesterase (PDE), and phytase (PAE) in the samples was calculated from the standard curves by measuring the absorbance at 450 nm with the spectrophotometric method of enzyme standardization.
Soil MBP was determined by chloroform fumigation (Brookes et al. 1982). Five grams (dry weight) of fresh soil was weighed into a Petri dish and placed in a vacuum desiccator together with 50 ml of ethanol-free chloroform and evacuated to vacuum. The vacuum desiccator was incubated in the dark for 24 h at room temperature and the chloroform was removed. The P of the supernatant was determined by ICP-OES by extracting MBP with 0.5 M NaHCO3 at a soil to water ratio of 1:10 and shaking for 30 min. Unfumigated soils of the same control were also extracted as described above. The P content in the fumigated soil was subtracted from the corresponding P content in the unfumigated soil and divided by 0.4 to calculate the MBP.
The soil P fractions are determined by the modified Hedley sequential P fraction method (Tiessen and Moir 1993). Briefly, after drying, grinding, and passing through a 100-mesh sieve, 0.5 g of soil was weighed into a centrifugal tube, extracted by successive additions of different reagents (H2O, 0.5 M NaHCO3, 0.1 M NaOH, 1 M HCl) at a soil to water ratio of 1:60, shaken for 16 h, centrifuged, and passed through a 0.45-μm filter membrane. The extracts were digested with potassium persulfate at 121 °C to determine the total P (TP). The phosphate in each extract was determined using the molybdenum blue colorimetric method and the Po in each extraction state was the difference between TP and Pi. The extracted P fractions were designated as H2O-P, NaHCO3-P, NaOH-P, and HCl-P. Bioavailable phosphorus (Bio-P) is considered to be the bioavailable fractions of soil P and is also more readily released into water (Cross and Schlesinger 1995).
In this study, the equations for Bio-Pi and Bio-Po are shown below (Wang et al. 2020a).
2.3 Statistical analyses
Statistical analyses of the data were carried out using SPSS. One-way ANOVA was carried out to test differences P fractions and phosphatase activities among the altitudes and transects. The unary linear regression was conducted to determine the relationship among phosphatase activity and P fractions. Pearson correlation was used to explain the relationship between P fractions and particle size.
3 Results and discussion
3.1 Physicochemical properties of soil samples
As shown in Table 1, soil pH in the WLFZ ranged from 7.37 to 8.75, with little variation overall, all being weakly alkaline. Natural vegetation zone was generally higher than artificial vegetation restoration zone, but the difference did not reach a significant level. Soil BD ranged from 1.01 to 1.52 g cm−3. Porosity ranged from 33.88 to 64.02%. MC ranges from 5.14 to 31.08%, with lower altitude soils (145 m, 155 m) subject to frequent periodic inundation and higher water content than higher altitude soils (165, 175 m). Soil median particle size ranged from 5.05 to 55.61 μm, with the mean median grain particle increasing with elevation in both sections (Fig. S1). Due to periodic water level fluctuations, fine-grained suspended particulate matter is more likely to be deposited in the WLFZ at lower altitude (Tang et al. 2018).
3.2 Distribution of P fractions in WLFZ soil
Artificial vegetation restoration had a significant effect on P fractions in the WLFZ, which varied with elevation (Fig. 2). Overall, inorganic P is the main fractions of soil P in the WLFZ (152.23 to 614.91 mg/kg). The contents of H2O-Pi, NaHCO3-Pi, NaOH-Pi, and HCl-Pi were 0.20 to 7.29 mg kg−1, 0.15 to 55.55 mg kg−1, 0.09 to 100.38 mg kg−1, and 130.32 to 631.49 mg kg−1. The TP content of artificial vegetation restoration zone was significantly higher than that of natural vegetation zone (508.92 ± 62.63 mg kg−1, 490.06 ± 120.72 mg kg−1, p < 0.05, Fig. 2a).

Box plot of P content in different transects and altitudes. Lower and upper box boundaries represent the quartiles (25% and 75% quantiles, respectively), the square is the mean value, and the solid lines are the median. Asterisks indicate significant difference in P content between the two transects (*, p < 0.05; ***, p < 0.001). A, artificial vegetation zone; N, natural vegetation zone
H2O-Pi is weakly adsorbed P, a status of P present in soil porewater and physically adsorbed on the surface of fine soil particles. NaHCO3-Pi is potentially active inorganic P, more stable than H2O-Pi. At each altitude, the contents of H2O-Pi and NaHCO3-Pi in the artificial vegetation restoration zone soil were higher than those in the natural vegetation zone (Fig. S1). The NaOH-Pi is metal oxide-bound inorganic P, which is easily converted to labile P and released into the water column under anaerobic conditions. At 155 m a.s.l., NaOH-Pi contents in the artificial vegetation restoration zone were higher than that in the natural vegetation zone, but the result was opposite at other altitudes. HCl-Pi is Ca-bound inorganic P, which is mainly found in rock-forming minerals and can be converted to soil Bio-P over a longer time scale through weathering and dissolution processes. In this study, HCl-Pi was the main component (44–100%) of the soil P pool in the WLFZ. The effects of artificial vegetation restoration measures and variations of altitude on HCl-Pi were not significant, indicating that its source was mainly soil minerals, which is consistent with previous results in the TGR area (Zhang et al. 2012). HCl-Pi is refractory P that is slow to participate in the P cycle, and on the other hand, its content is relatively high, and it is not sensitive to vegetation restoration and periodic DRW cycle. The Bio-Pi content of artificial vegetation restoration zone soil was significantly higher than that of natural vegetation zone at each altitude, except for 155 m a.s.l. (85.81 ± 30.51 mg kg−1, 51.81 ± 28.28 mg kg−1, p < 0.05, Fig. 2b). Possible reasons for the above phenomenon are as follows: (1) the vegetation cover after the artificial vegetation restoration reduced the amount of soil erosion and P loss, and (2) the vegetation restoration has facilitated the transformation of different fractions of P in the soil of the WLFZ, making the P fractions migrate in a direction more conducive to plant growth and uptake.
In the WLFZ of the TGR, Ca-P was formed and released due to the reduction and dissolution of Fe–P during the flooding period, and the mineralization of soil organic P was intensified during the drying period, which resulted in the massive depletion of soil organic P (Ma et al. 2008). This explains why the content of organic P in the WLFZ soil is significantly lower than that of inorganic P in this study (Fig. S1). The content of H2O-Po, NaHCO3-Po, NaOH-Po, and HCl-Po ranged from 0 to 13.17 mg kg−1, 2.06 to 41.75 mg kg−1, 0.97 to 51.08 mg kg−1, and 0 to 121.90 mg kg−1. H2O-Po and NaHCO3-Po were both labile organic P (Gao et al. 2022; Bauke et al. 2022), easily migrated and mineralized in the soil and entered the water column under flooding conditions, with no significant pattern of variation between altitudes and transects (Fig. S2). NaOH-Po is bioavailable Po and HCl-Po is mainly Ca-bound organic P (Wang et al 2020a; Bauke et al. 2022). The contents of NaOH-Po in artificial vegetation restoration zone decreased with the decreasing of altitude, while the content of HCl-Po in natural vegetation zone increased with the decreasing of altitude, showing an opposite variation pattern in transects along altitude, which may be related to the change of the response relationship between phosphatase activity and organic P in vegetation restoration transects. There was no significant difference in Bio-Po contents between the two transects overall, but the distribution along altitude showed a certain pattern (Fig. 2c, f). At low altitudes (145 m, 155 m), the soil Bio-Po contents in artificial vegetation restoration zone were lower than that in natural vegetation zone, while at high altitudes (155 m, 165 m) it was the opposite (Fig. 2f). The content of soil Bio-Po in artificial vegetation restoration zone decreased with the decreasing of altitudes, while no significant change was found in natural vegetation zone, indicating that the enhancement of DRW cycle in the soil after vegetation restoration led to the enhancement of Bio-Po mineralization, and this distribution pattern is different from that in the study of Pengxi River (Gao et al. 2022). In terms of distribution along depth, while each form Pi did not vary significantly from Po (Fig. S3).
Studies have shown that particle size is an important factor controlling the distribution patterns of P fractions, and that as particle size decreases, Bio-P content of the soil/sediment increases (Zhang et al. 2012). In this study, the soil median particle size increased with altitudes in both transects (Fig. 3), and Bio-P and Bio-Pi showed a significant positive correlation (Table 2, p < 0.05), which is consistent with the findings of Wang et al. (2020b).

Particle size of the samples at different transects a artificial vegetation restoration zone and b natural vegetation zone. The black squares represent the median particle size. a: 0–10 cm, b: 10–30 cm, and c: 30–50 cm
3.2.1 MBP in WLFZ soil
Soil MBP is the labile fractions of soil organic P and is dominated by nucleic acids, phospholipids and is generally highly associated with NaHCO3-P (Khan and Rainer 2012). Compared with other organic P in the soil, MBP is more easily to be mineralized and absorbed by plants, becoming an important source of soil Bio-P (Oehl et al. 2001; George et al. 2010). In this study, MBP ranged from 2.54 to 57.60 mg kg−1, accounting for 0.45 to 13.74% of TP (Fig. 4a). In the two transects, the highest MBP content was found at 165 m a.s.l., and the content of MBP decreased with the decreasing of altitude (Fig. 4b). Previous studies have demonstrated that the soil MBP content within the TGR area exhibits variation based on the presence of different vegetation covers, however, the spatial and temporal distribution of MBP in the region does not follow a consistent pattern (Jia et al. 2016; Ren et al. 2018). As the soil in lower altitudes experienced frequent DRW cycle, a large amount of water-soluble P was released after the lysis of cells, which will also affect the absorption of available P by vegetation, and the MBP content in the two sections shows a similar variation trend along the altitude. This finding is different from previous reports (Jia et al. 2016; Yang et al. 2017).

Distribution of MBP content in a transect and b altitude. Lower and upper box boundaries represent the quartiles (25% and 75% quantiles, respectively), the square is the mean value, and the solid lines are the median. Asterisks indicate significant difference in P content between the two transects (*, p < 0.05; **, p < 0.01). A, artificial vegetation zone; N, natural vegetation zone
The contents of MBP in artificial vegetation restoration zone were significantly lower than that in natural vegetation zone at each altitude (p < 0.05, Fig. 4a), which was contrary to the dynamic variation of Bio-Pi (Fig. 2e). The soil MBP content was significantly positively correlated with NaOH-Po and Bio-Po in artificial vegetation restoration zone (p < 0.01, Fig. 5a; p < 0.01, Fig. 5b), but not in natural vegetation zone (p > 0.05, Fig. 5a, b), indicating that MBP became the component of NaOH-Po and Bio-Po in the WLFZ soil after vegetation restoration. There was no correlation between MBP component and NaHCO3-P and NaOH-Pi, which was different from previous studies (Ren et al. 2018). Numerous studies have shown that both plant uptake and litter accumulation may lead to transformation of soil MBP (Chen et al. 2003; Cleveland et al. 2007). The dynamic balance between vegetation nutrient and soil nutrient was affected by vegetation type, changed the transformation relationship between MBP and Bio-Po, which resulted in the decrease of soil MBP content in the WLFZ after artificial vegetation restoration.

Relationships between MBP and the content of a NaOH-Po and b Bio-Po
3.2.2 Phosphatase activity of WLFZ soil
Soil phosphatase is a kind of hydrolase that catalyzes and accelerates mineralization of soil organic P, and it is an important index to measure the mineralization rate of organic P and the bioavailability of P (Nannipieri et al. 2011; Burns et al. 2013). Soil phosphatase activities in the WLFZ are shown in Fig. S4; the activity of ALP, ACP, PDE, and PAE was 1.71 to 2.60 μmol·(g·h) −1, 0.53 to 0.76 μmol·(g·h) −1, 4.35 to 6.58 μmol·(g·h) −1, and 1.74 to 1.89 μmol·(g·h) −1. Generally, ALP and PAE activities were similar, ACP activity was the lowest, and PDE activity was the highest.
Both ALP and ACP are belonged to phosphomonoesterase, ALP is mainly released by microorganisms, while ACP is mainly produced by plants, and they have different adaptations to soil pH, catalyzing the hydrolysis of phosphate monoester in soil organic P to release phosphate molecules (Dinkelaker et al. 1992; Nannipieri et al. 2011). PDE can catalyze the hydrolysis of phospholipids and nucleic acids in soil organic P to release phosphomonoester, and the hydrolyzed products can be further hydrolyzed by ALP and ACP (Turner and Haygarth 2005; Spohn et al. 2013). PAE catalyzes the degradation of soil phytic acid to myo-inositol phosphates or inorganic P and is a specific phosphomonoesterase. Numerous studies have shown that soil pH is an important factor affecting enzyme activity (Dinkelaker et al. 1992; Turner and Haygarth 2005). In this study, the soil in WLFZ is weakly alkaline, which may explain the lower ACP activity. In general, vegetation restoration can increase soil phosphatase activity (Fu et al. 2020; Zhang et al. 2021), but our results show that there is no significant difference in soil phosphatase activity at different altitudes between the two transects (Fig. S4). This finding may be related to that P is not a limiting nutrient for the productivity of soil in WLFZ of TGR, which is consistent with the study of Ren et al. (2018). Although the artificial vegetation restoration had no significant effect on phosphatase activity, the PDE activity increased with the decreasing of altitude. Gao et al. (2022) found that soil phosphatase activity in the WLFZ of Pengxi River, a tributary of the TGR, was affected by plant and microbial biomass and increased with the increasing of altitude, which was contrary to the results of this study. Frequent DRW cycling cannot only intensify soil Bio-P mineralization, but also lead to the dissolution of reducing metal ions, thus promoting or inhibiting phosphatase activity (Pulford and Tabatabai 1988; Wang et al. 2021). The vegetation restoration in the WLFZ may further enhance these effects, resulting in higher soil phosphatase activity at lower altitudes compared to higher altitudes.
Soil orthophosphate absorbed by plants to maintain growth, and organic P mineralized to provide nutritional support when inorganic P deficiency. Therefore, the level of soil phosphatase activity directly affects the turnover of organic P (Turner and Haygarth 2005; Li et al. 2021; Wang et al. 2021). The activity of PDE was the highest in the two transects; one possible explanation is that the soil organic P in this study were mainly phosphodiesters. The H2O-Pi was significantly negatively correlated with the PDE activity in artificial vegetation restoration zone (p < 0.05, Fig. 7a) and natural vegetation zone (p < 0.05, Fig. 7a). H2O-Pi is the most easily uptaked and utilized P fractions in soil by plants, which indicates that when the soil inorganic P contents in the WLFZ are low, plants and microorganisms will be prompted to secrete phosphatase and raise the PDE activity to promote the process of organic P mineralization.
There is a significant correlation between soil P content and enzyme activity, but due to the wide range of soil enzyme sources and the heterogeneity of soil physicochemical properties, the correlation between P fractions and enzyme activity is currently difficult to establish (Adams et al. 1992; Allison et al. 2007). On the one hand, the production of phosphatase promoted by microorganisms usually leads to depletion of organic P (Nannipieri et al. 2011); on the other hand, lower concentrations of easily degradable substrates may inhibit enzyme activity (Turner and Haygarth 2005; Boitt et al. 2018; Wang et al. 2021). The soil Bio-Po content in artificial vegetation restoration zone and natural vegetation zone was different at each altitude, but not significant (Fig. 2f), indicating that the inhibition of PDE activity had no relation with substrate concentration. Phosphodiester in soil must be sequentially hydrolyzed by PDE and phosphomonoesterase to release inorganic phosphate for plant uptake and utilization (Turner and Haygarth 2005). In natural vegetation zone, PDE activity was positively correlated with NaOH-Po and Bio-Po (p < 0.05, Fig. 7f; p < 0.05, Fig. 7i), while the distribution of soil phosphatase activity and Bio-P did not change regularly along altitude (Fig. S4). After the artificial vegetation restoration, the original vegetation pattern in the WLFZ was changed, and the distribution of soil microbial community along altitude also succession correspondingly. There was a significant negative correlation between PDE activity and NaOH-Po and Bio-Po (p < 0.05, Fig. 7e; p < 0.01, Fig. 7h); the distribution of Bio-Po and PDE activity showed an opposite pattern along altitude (Fig. 2f, Fig. 6a). Soil phosphomonoesterase (ALP, ACP) activity and Bio-Po in the two transects revealed the same correlation as PDE and Bio-Po, through the indirect influence of PDE and phosphomonoesterase (Fig. 7). The evidence presented indicates that artificial vegetation restoration has enhanced the response of PDE activity to the DRW cycle. Moreover, it has influenced the distribution pattern of Bio-Po across different altitudes. Furthermore, a notable positive correlation between MBP and Bio-Po was observed (p < 0.01, Fig. 5a; p < 0.05, Fig. 5b), providing further support for this perspective.

Distribution of PDE activity along altitude in a artificial vegetation restoration zone and b natural vegetation zone. Different lowercase letters represent significant differences between different altitudes (p < 0.05)

Relationships between soil P fractions and phosphatase activity
Previous studies have shown that soil phytate chelates with metal cations such as Ca2+, Mg2+, and Zn2+ and can release inorganic P through phytase catalyzed hydrolysis to provide P sources for plant growth (Perkins and Underwood 2001; Zhu et al. 2017). HCl-Po is a resistant organic P containing phytic (He et al. 2006). In this study, soil PAE activity in natural vegetation zone had a significant positive correlation with HCl-Po (p < 0.05, Fig. 7b). In the artificial vegetation restoration zone, PAE activity and HCl-Po distribution along the altitude are not regular, and there is no significant correlation between them. One possible explanation is that the vegetation restoration years in the WLFZ is short, and phytase mineralization of organic P is not fully established. Unfortunately, our study was insufficient to investigate the relationship between enzyme activity and P fractions on the vegetation restoration years.
4 Conclusion
The study showed that the TP and Bio-Pi contents in the WLFZ of TGR were significantly increased after artificial vegetation restoration, and the distribution of Bio-Pi was mainly affected by particle size. The variation in Bio-Po content was not significantly different between the transects in general, but the distribution was different between different altitudes. Artificial vegetation restoration exerts an impact on the mineralization of soil OP within the WLFZ, thereby altering the biological bioavailability of phosphorus (P). Furthermore, this effect varies with altitude due to the influence of the DRW cycle. This change may protect water from the threat of eutrophication in the WLFZ. Artificial vegetation restoration resulted in a significant decrease in MBP content and an increase in PDE activity, and both showed regular changes along the altitude gradient. The PDE activity mainly controls the soil Bio-Po mineralization, but the mechanism of action in the two transects is different. Through these studies, vegetation restoration can be better selected to improve the ecological function of WLFZ wetlands at different altitudes, providing a new perspective for understanding the soil P cycle in the WLFZ.
References
Adams MA (1992) Phosphatase activity and phosphorus fractions in karri (Eucalyptus diversicolor F. Muell.) forest soils. Biol Fertil Soils 14(3):200–204
Allison VJ, Condron LM, Peltzer DA, Richardson SJ, Turner BL (2007) Changes in enzyme activities and soil microbial community composition along carbon and nutrient gradients at the Franz Josef chronosequence. New Zealand Soil Biol Biochem 39(7):1770–1781
Bauke SL, Wang Y, Saia SM, Popp C, Tamburini F, Paetzold S, Amelung W, Sperber C (2022) Phosphate oxygen isotope ratios in vegetated riparian buffer strip soils. Vadose Zone J 21(3):e20193
Birch HF (1958) The effect of soil drying on humus decomposition and nitrogen availability. Plant Soil 10(1):9–31
Boitt G, Black A, Wakelin SA, McDowell RW, Condron LM (2018) Impacts of long-term plant biomass management on soil phosphorus under temperate grassland. Plant Soil 427(1):163–174
Brödlin D, Kaiser K, Kessler A, Hagedorn F (2019) Drying and rewetting foster phosphorus depletion of forest soils. Soil Biol Biochem 128:22–34
Brookes PC, Powlson DS, Jenkinson DS (1982) Measurement of microbial biomass phosphorus in soil. Soil Biol Biochem 14(4):319–329
Bünemann EK, Keller B, Hoop D, Jud K, Boivin P, Frossard E (2013) Increased availability of phosphorus after drying and rewetting of a grassland soil: processes and plant use. Plant Soil 370(1):511–526
Burns RG, DeForest JL, Marxsen J, Sinsabaugh RL, Stromberger ME, Wallenstein MD, Weintraub MN, Zoppini A (2013) Soil enzymes in a changing environment: current knowledge and future directions. Soil Biol Biochem 58:216–234
Chen CR, Condron LM, Davis MR, Sherlock RR (2003) Seasonal changes in soil phosphorus and associated microbial properties under adjacent grassland and forest in New Zealand. For Ecol Manag 177:539–557
Cheng F, Li W, Castello L, Murphy BR, Xie S (2015) Potential effects of dam cascade on fish: lessons from the Yangtze river. Rev Fish Biol Fish 25:569–585
Cleveland CC, Liptzin CD (2007) C: N: P stoichiometry in soil: is there a “redfield ratio” for the microbial biomass? Biogeochemistry 85:235–252
Cleveland CC, Townsend AR, Taylor P, Alvarez-Clare S, Bustamante MMC, Chuyong G, Dobrowski SZ, Grierson P, Harms KE, Houlton BZ, Marklein A, Parton W, Porder S, Reed SC, Sierra CA, Silver WL, Tanner EVJ, Wieder WR (2011) Relationships among net primary productivity, nutrients and climate in tropical rain forest: a pan-tropical analysis. Ecol Lett 14:1313–1317
Cross AF, Schlesinger WH (1995) A literature review and evaluation of the Hedley fractionation: applications to the biogeochemical cycle of soil phosphorus in natural ecosystems. Geoderma 64(3):197–214
Dinkelaker B, Marschner H (1992) In vivo demonstration of acid-phosphatase-activity in the rhizosphere of soil-grown plants. Plant Soil 144:199–205
Evtimova VV, Donohue I (2016) Water-level fluctuations regulate the structure and functioning of natural lakes. Freshw Biol 61:251–264
Fu D, Wu X, Duan C, Chadwick DR, Jones DL (2020) Response of soil phosphorus fractions and fluxes to different vegetation restoration types in a subtropical mountain ecosystem. CATENA 193:104663
Gao YL, Fang F, Tang ZC, Zhang R, Jiang YX, Guo JS (2022) Distribution characteristics of soil phosphorus forms and phosphatase activity at different altitudes in the soil of water-level-fluctuation zone in Pengxi River, Three Gorges Reservoir. Huan Jing Ke Xue 43(10):4630–4638
George TS, Turner BL, Gregory PJ, Cademenun BJ, Richardson AE (2010) Depletion of organic phosphorus from Oxisols in relation to phosphatase activities in the rhizosphere. Eur J Soil Sci 57:47–57
Hale BW, Adams MS (2007) Ecosystem management and the conservation of river–floodplain systems. Landsc Urban Plan 80(1–2):23–33
He Z, Fortuna AM, Senwo ZN, Tazisong IA, Honeycutt CW, Griffin TS (2006) Hydrochloric fractions in Hedley fractionation may contain inorganic and organic phosphates. Soil Sci Soc Am J 70(3):893–899
Hill AR (2000) Stream chemistry and riparian zones. Streams and ground waters 83–110
Jeppesen E, Kronvang B, Meerhoff M, Søndergaard M, Hansen KM, Andersen HE, Lauridsen TL, Liboriussen L, Beklioglu M, Özen A, Olesen JE (2009) Climate change effects on runoff, catchment phosphorus loading and lake ecological state, and potential adaptations. J Environ Qual 38(5):1930–1941
Jia GM, He L, Cheng H, Wang ST, Xiang HY, Zhang XF, Xi Y (2016) Ecological stoichiometry characteristics of soil microbial biomass carbon, nitrogen and phosphorus under different vegetation covers in three gorges reservoir area. Res Soil Water Conserv 23:23–27
Khan KS, Joergensen RG (2012) Relationships between P fractions and the microbial biomass in soils under different land use management. Geoderma 173:274–281
Khan SU, Hooda PS, Blackwell MSA, Busquets R (2022) Effects of drying and simulated flooding on soil phosphorus dynamics from two contrasting UK grassland soils. Eur J Soil Sci 73(1):e13196
Lembi CA (2001) Limnology, lake and river ecosystems. J Phycol 37(6):1146–1147
Li SZ, Deng Y, Shi FN, Hu MM, Pang BH, Wang YC, Li K, Peng WQ, Liang XD, Bao YF, Meng JJ (2019) Research progress on water-level-fluctuation zones of reservoirs: a review. Wetl Sci 17:689–696
Li JB, Xie T, Zhu H, Zhou J, Li CN, Xiong WJ, Xu L, Wu YH, He ZL, Li XZ (2021) Alkaline phosphatase activity mediates soil organic phosphorus mineralization in a subalpine forest ecosystem. Geoderma 404:115376
Ma LM, Zhang M, Teng YH, Zhao JF (2008) Characteristics of phosphorous release from soil in periodic alternately waterlogged and drained environments at WFZ of the Three Gorges Reservoir. Huan Jing Ke Xue 29(4):1035–1039
Nannipieri P, Giagnoni L, Landi L, Renella G (2011) Role of phosphatase enzymes in soil. Phosphorus in Action, Springer, Berlin, Heidelberg 2011:215–243
Oehl F, Oberson A, Probst M, Fliessbach A, Roth HR, Frossard E (2001) Kinetics of microbial P uptake in cultivated soils. Biol Fert Soils 34:31–41
Perkins RG, Underwood GJC (2001) The potential for phosphorus release across the sediment–water interface in an eutrophic reservoir dosed with ferric sulphate. Water Res 35(6):1399–1406
Pulford ID, Tabatabai MA (1988) Effect of waterlogging on enzyme activities in soils. Soil Biol Biochem 20(2):215–219
Ren QS, Song H, Yuan ZX, Ni XL, Li CX (2018) Changes in soil enzyme activities and microbial biomass after revegetation in the three gorges reservoir. China Forests 9(5):249
Shen YF, Cheng RM, Xiao WF, Zeng LX, Wang LJ, Sun PF, Chen T (2022) Temporal dynamics of soil nutrients in the riparian zone: effects of water fluctuations after construction of the Three Gorges Dam. Ecol Indic 139:108865
Spohn M, Kuzyakov Y (2013) Distribution of microbial- and root-derived phosphatase activities in the rhizosphere depending on P availability and C allocation–coupling soil zymography with 14C imaging. Soil Biol Biochem 67:106–113
Tang Q, Bao Y, He X, Zhou H, Cao Z, Gao P (2014) Sedimentation and associated trace metal enrichment in the riparian zone of the Three Gorges Reservoir, China. Sci Total Environ 479:258–266
Tang Q, Collins AL, Wen AB, He XB, Bao YH, Yan DC, Long Y, Zhang YS (2018) Particle size differentiation explains flow regulation controls on sediment sorting in the water-level fluctuation zone of the Three Gorges Reservoir, China. Sci Total Environ 633:1114–1125
Tiessen HJWB, Moir JO (1993) Characterization of available P by sequential extraction. Soil Sampl Methods Anal 7:5–229
Turner BL, Haygarth PM (2005) Phosphatase activity in temperate pasture soils: potential regulation of labile organic phosphorus turnover by phosphodiesterase activity. Sci Total Environ 344(1–3):27–36
Turner BL, Driessen JP, Haygarth PM, Mckelvie ID (2003) Potential contribution of lysed bacterial cells to phosphorus solubilisation in two rewetted Australian pasture soils. Soil Biol Biochem 35:187–189
Vidon F, Allan C, Burns D, Duval TP, Gurwick N, Inamdar S, Lowrance R, Okay J, Scott D, Sebestyen S (2010) Hot spots and hot moments in riparian zones: potential for improved water quality management. J Am Water Resour Assoc 46(2):278–298
Wang Y, Zhang N, Wang D, Wu J, Zhang X (2018) Investigating the impacts of cascade hydropower development on the natural flow regime in the Yangtze River, China. Sci Total Environ 624:1187–1194
Wang C, Fang F, Yuan Z, Zhang R, Zhang W, Guo J (2020a) Spatial variations of soil phosphorus forms and the risks of phosphorus release in the water-level fluctuation zone in a tributary of the Three Gorges Reservoir. Sci Total Environ 699:134124
Wang Y, Zhang N, Wang D, Wu J (2020b) b) Impacts of cascade reservoirs on Yangtze river water temperature: assessment and ecological implications. J Hydrol 590:125240
Wang CQ, Xue L, Jiao RZ (2021) Soil phosphorus fractions, phosphatase activity, and the abundance of phoC and phoD genes vary with planting density in subtropical Chinese fir plantations. Soil till Res 209:104946
Wu JG, Huang JH, Han XG, Gao XM, He FL, Jiang MX, Jiang ZG, Primack RB, Shen ZH (2004) The Three Gorges Dam: an ecological perspective. Front Ecol Environ 2(5):241–248
Wu Y, Wang X, Zhou J, Bing H, Sun H, Wang J (2016) The fate of phosphorus in sediments after the full operation of the Three Gorges Reservoir, China. Environ Pollut 214:282–289
Xue L, Hou P, Zhang Z, Shen M, Liu F, Yang L (2020) Application of systematic strategy for agricultural non-point source pollution control in Yangtze River basin. China Agr Ecosyst Environ 304:107148
Yang WH, Qing H, Ren Q, He Y, Li X, Li C (2017) Characteristics of soil microbial biomass C and N under revegetation in the hydro-fluctuation belt of the Three Gorges Reservoir Region. Acta Ecol Sin 37(23):7947–7955
Ye C, Cheng X, Zhang Q (2014) Recovery approach affects soil quality in the water level fluctuation zone of the Three Gorges Reservoir, China: implications for revegetation. Environ Sci Pollut Res 21(3):2018–2031
Ye C, Chen C, Butler OM, Rashti MR, Esfandbod M, Zhang DuM, Q, (2019) Spatial and temporal dynamics of nutrients in riparian soils after nine years of operation of the Three Gorges Reservoir, China. Sci Total Environ 664:841–850
Zhang B, Fang F, Guo J, Chen Y, Li Z, Guo S (2012) Phosphorus fractions and phosphate sorption-release characteristics relevant to the soil composition of water-level-fluctuating zone of Three Gorges Reservoir. Ecol Eng 40:153–159
Zhang Y, Li Y, Wang S, Umbreen S, Zhou C (2021) Soil phosphorus fractionation and its association with soil phosphate-solubilizing bacteria in a chronosequence of vegetation restoration. Ecol Eng 164:106208
Zhu J, Qu B, Li M (2017) P mobilization in the Yeyahu wetland: phosphatase enzyme activities and organic P fractions in the rhizosphere soils. Int Biodeterior Biodegradation 124:304–313
Zhu K, Li W, Yang S, Ran Y, Lei X, Ma M, Wu S (2022) Intense wet-dry cycles weakened the carbon sequestration of soil aggregates in the riparian zone. CATENA 212:106117
Funding
This study was sponsored jointly by the National Key Research and Development Project (2021YFC3201002), the Strategic Priority Research Program of CAS (XDB40020400), the National Natural Science Foundation of China (41977296, 42277253, 51809287, U2040211), the Guizhou Science and Technology Department Fund ([2020] 4Y006, [2019]1042), the Central Leading Local Science and Technology Development Fund Project ([2021]4028), the Youth Innovation Promotion Association CAS (No. 2019389), and the Follow-up Work of the Three Gorges Project (2136902).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Responsible editor: Wenzhi Cao
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Qiu, S., Wang, Y., Wang, J. et al. Effects of artificial vegetation restoration on the fractions and availability of soil phosphorus in the water-level-fluctuating zone of Three Gorges Reservoir, China. J Soils Sediments 24, 1826–1837 (2024). https://doi.org/10.1007/s11368-023-03603-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11368-023-03603-x