
Abstract
Use of waste wood biomass for bioenergy produces wood ash as a by-product; this ash is typically landfilled, but can potentially play an important role in soil improvement and forest restoration. In particular, high-carbon wood ash biochar (HCWAB) could supply nutrients, improve substrate water-holding capacity and pH, and emulate the ecosystem benefits of wildfire residues. Thickened tailings sites at metal mines across Canada are subject to stringent restoration regulations that entail planting of native trees to promote rapid reforestation. While HCWAB may prove beneficial in this context, field trials have been very limited to date. We conducted a large-scale, replicated field trial on sand-capped tailings at an operational gold mine in the Canadian boreal forest to assess the impact of HCWAB (at dosages of 0, 6.4, 12.8, and 19.1 t/ha) on survival and growth of four native tree species, as well as substrate chemical properties and element uptake in tree tissues. After 2 years, the survival of planted, native trees was highest at low to moderate application rates; HCWAB dosages above 13 t/ha presented reduced tree survival to levels comparable to unamended substrates. Tree growth was higher across all HCWAB doses relative to growth in samples planted on untreated substrates; tree species and initial size also had large impacts on final tree survival and aboveground growth. The survival of Betula papyrifera was significantly higher than other species, while smaller transplanted trees in general survived in greater numbers compared to larger size classes. Volunteer herbaceous vegetation significantly increased at the higher HCWAB application dosages and tree performance was negatively correlated with vegetation cover, consistent with a resource competition effect. HCWAB additions to sand-capped mine tailings did not significantly alter tree tissue concentrations or substrate availability of potentially toxic metals (Cd, Cu, Al). We conclude that low to moderate dosages of HCWAB on sand-capped tailings, particularly between 6.4 and 12.8 t/ha, may offer benefits to early tree survival, growth, and substrate nutrient status without causing significant risks of phytotoxicity and recommend future field trials focus on strategies to reduce tree competition with competing vegetation.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Modern advances in geological exploration, mining technologies, and ore processing have led to increased mineable resources and extended mine life for operations worldwide (Reid et al. 2009), while also creating additional overburden waste and tailings material. The design of safe and high-capacity tailings impoundments has thus been an emerging research focus; priorities include minimizing surface footprint, optimizing storage and closure costs, and reducing environmental risks (Wei et al. 2016). Recently, thickened or paste tailings have been preferred to conventional slurry to increase stacking height, and reduce spread, as well as lower risks of seepage and structural failure (Simms 2017). However, the low hydraulic conductivity and poor aeration associated with stacked, thickened tailings can dramatically inhibit volunteer plant establishment (Ye et al. 2002; Mendez and Maier 2007) and tree root development (Larchevêque et al. 2014; Tordoff et al. 2000; Borgegard and Rydin 1989), thus ultimately impairing revegetation and restoration. Once mining operations cease, thickened tailings are generally capped with layers of inert materials that lack the organic matter, nutrients, and physical structure necessary for plant life. Establishing native vegetation cover and reforestation are generally mandatory components of mine closure plans despite being exceptionally difficult to achieve on most thickened tailings sites or capped impoundments (Beesley et al. 2011). Broadly, forest restoration aims to shift a damaged ecosystem toward its previous, more natural state (Kuuluvainen and Nummi 2023), and hence relies on the success of volunteer and planted native species. Local plant diversity generally enhances beneficial ecosystem functions, such as productivity and carbon storage, in forests and other environments (Chisholm et al. 2013; Xu et al. 2020) and thus a focus on native species is the benchmark for modern mine tailings reforestation programs. Covering tailings sites with new or salvaged topsoil can allow for long-term planted tree success; however, this material may be unavailable in large quantities, particularly at remote sites; hence, there is an increasing focus on accessible amendments that facilitate the growth and survival of plant life.
The term biochar refers to high-carbon material produced from heating biomass feedstocks at high temperatures under oxygen-limited conditions (Lehmann and Joseph 2015). The impacts of a specific biochar on plants and broader ecosystem functions vary based on feedstock and production conditions (Joseph et al. 2021); however, typical benefits include increased soil pH, water holding capacity, and nutrient retention as well as reduced availability of heavy metal in soils (Beesley and Marmiroli 2011). To date, most research attention has been devoted to biochar’s effects within agricultural systems (Biederman and Harpole 2013), with less focus on forest ecosystems, particularly under field conditions (Thomas and Gale 2015). Biochar on contaminated, degraded mine tailings may enhance plant and tree revegetation as well as substrate remediation objectives, but most data available are from lab and greenhouse trials (e.g., Fellet et al. 2011; Lebrun et al. 2019; Trippe et al. 2021), with very few field trials on mine tailing sites (Liu et al. 2022; Román-Dañobeytia et al. 2021; Williams and Thomas 2023a, 2023b). Improvements in plant survival (e.g., Kuttner and Thomas 2017; Marsh et al. 2023) and growth (Thomas and Gale 2015) have been observed from the addition of biochar in restoration trials at various rates and produced under a range of pyrolysis conditions. Biochar derived from wood feedstocks could be particularly beneficial on mine tailings sites in fire-adapted ecosystems, such as those within Canada’s boreal forest zone (Brais et al. 2015). However, the acquisition and transport of designed, wood biochar for large-scale, remote applications remains financially and logistically impractical.
Wood ash is produced as a by-product of wood-fueled heat and power generation; the ash is comparable to biochar in that it is porous, may be high in important plant nutrients, and has a high liming potential (Pitman 2006). Research conducted to date suggests that the impacts of wood ash on forest soils vary with ash properties, dosage applied, and initial substrate conditions (Augusto et al. 2008). Some wood ashes contain high levels of total carbon (TC), which qualifies them as a biochar based on regulatory guidelines for soil amendments (IBI 2015; Hannam et al. 2018). Inconsistent feedstock and gasification processes contribute to more heterogeneous products relative to conventional pyrolyzed biochars (James et al. 2014) and variation in wood ash properties has been a barrier for more widespread research and application (Huotari et al. 2015), particularly on contaminated mine tailings. Impurities in wood ash products, notably toxic metal/loids, may introduce risk for their use if these become bioavailable in soils. Still, there remain strong motivations for applying high-carbon wood ash biochar (HCWAB) to boreal forest ecosystems: its accessibility in remote mining regions, affordability relative to other amendment options, as well as the potential HCWAB holds to improve to soil nutrient and heavy metal status, and ultimately tree performance (Augusto et al. 2008; Hope et al 2017). On acidic soils in Northern Alberta, Gill et al. (2015) found that low dosages of wood ash, as well as wood ash combined with N, both reduced fertilizer requirements and resulted in increased crop yields, pH, and soil nutrient availability. Indeed, experiments applying wood ash to soils have reported improved available plant nutrients such as calcium and magnesium (Saarsalmi et al. 2004), decreased bioavailability of toxins (DeVolder et al. 2003), and increased herbaceous vegetation (Ferreiro et al. 2011) and tree growth (Omil et al. 2013; Salam et al. 2019).
In the fire-adapted boreal forest, application of wood ash to soils has resulted in increased growth of scots pine seedlings (Hytönen 2003; Moilanen et al. 2002); however, growth responses have been negative or neutral on some N-limited soils (Bieser and Thomas 2019; Jacobson et al. 2014; Saarsalmi et al. 2004). Growth effects from wood ash are also variable based on initial soil pH and tree species. Indeed, a meta-analysis of wood ash trials from managed forests across Europe and North America found that initial soil pH and tree species were the strongest predictors of tree growth, and wood ash amendments only resulted in greater growth for softwoods in very acidic soils (Reid and Watmough 2014). Emilson et al. (2019)’s review of Canadian forest trials found significantly higher wood ash growth response in jack pine relative to spruce species. Growth declines from wood ash have also been reported; on a managed boreal forest plot, Brais et al. (2015) found that spruce growth at high wood ash amendment dosages was significantly reduced, while no treatment growth effects were observed in jack pine over the same 5-year time.
Wood ash from electricity-generating facilities has been applied to mine tailings in greenhouse pot trials (DeVolder et al. 2003; Salam et al. 2019); however, very few field trials have tested application of high-carbon wood ash biochar for reforestation of tailings sites (Williams and Thomas 2023b). The use of HCWAB as part of a mine site restoration plan will depend on its in situ effects, particularly those related to the performance of planted native trees, in addition to tree nutrient and contaminant uptake response. A large-scale, 2-year field experiment was installed on sand-capped tailings at an operational gold mine in Northern Ontario, Canada, in order to evaluate the effect of HCWAB on (1) survival and aboveground growth of planted native locally collected saplings (“wildings”), (2) substrate concentration of available nutrients and potentially toxic elements (PTEs), and (3) levels of nutrients and metals in aboveground tree tissues. We hypothesized that HCWAB amendments would increase the substrate supply of available plant nutrients and that this would result in greater concentrations measured in branch and foliage tissues of planted wildings. We also predicted that concentrations of toxic elements such as heavy metals in both tailings and tree tissues would decrease in response to increased dosages of HCWAB, but that this trend may not extend to the highest dosages applied as trace toxins in the char itself may become mobilized at a threshold HCWAB rate above 15–20 t/ha (Williams and Thomas 2023b). Species-specific survival and growth responses were also expected independent of HCWAB treatment, with high survival and growth predicted in Salicaceae species (willows and poplars) since they are well-adapted to open sites and variable environmental conditions (Mosseler et al. 2014). Lastly, we anticipated that fire-adapted Pinus banksiana wildings would exhibit the strongest survival and growth treatment effects from HCWAB.
Materials and methods
Study site
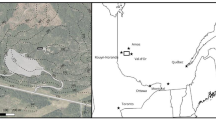
The field experiment was installed on an outer section of the sand-capped tailings zone at Musselwhite Mine, an operational underground gold mine located in Northwestern Ontario’s boreal forest (52.61°N, 90.36°W). The mine’s underground operations commenced in 1997 and have a current life of mine estimation extending to 2028. Expanded mineral reserves motivated a shift to thickened tailings in 2010; however, the experiment was installed on a zone with conventional, 50% solids slurry produced during early operations (Kam et al. 2011). The tailings slurry was evenly capped with a ~ 3-m deep layer of inert sand material from adjacent burrow pits. Average temperature at nearby Pickle Lake weather station ranges from − 19.3 °C in January to 17.7 °C in July, with the minimum daily temperature being above freezing from May to September inclusive (Environment Canada 2023). Topsoil layers around the mine site are mostly impervious and tailings have maintained low oxidation and near neutral pH levels throughout the operations. A natural forest fire impacted large areas around the mine and experiment site in 2011; stands at the time of experimental setup were dominated by regenerating Pinus banksiana, as well as other species typical of boreal forest regions, notably Picea glauca, Picea mariana, Abies balsamea, and Larix laricina and native deciduous species including Betula papyrifera, Populus balsamifera, and Populus tremuloides. Heavy rainfall and aeolian transport in the years prior to experiment installation had led to deposition of some thickened tailings material from the more modern storage pond, which has naturally integrated into the sand layer (D. Achircano Condori, Newmont Musselwhite, pers. comm.).
Experimental design
Site installation
A 4 \(\times\) 4 factorial block design was installed in August 2017 along an accessible, slightly elevated area of the sand-capped tailings storage site. To do so, existing low-lying vegetation and shrubs were removed from the site by hand, then substrate compaction was loosened by tilling with a bulldozer and manually raking the upper 10-cm layer. Twelve 10 m \(\times\) 10 m plots were delineated, separated from east to west by alleys 1-m wide, and north to south by a 3-m wide central corridor. To each plot, one of four HCWAB dosages—0.0, 6.4, 12.8, and 19.1 t/ha—was applied in a randomized block design with three total replications of each treatment. Wood ash biochar was raked into the top 6-cm layer of each plot. A combined total of 432 wildings were planted, all native to the area and readily found in regenerating forested zones of the mine property: Pinus banksiana Lamb (jack pine, hereinafter pine) (67%), Salix bebbiana Sarg. (Bebb’s willow, hereinafter willow) (11%), Populus tremuloides Michx. (trembling aspen, hereinafter aspen) (11%), and Betula papyrifera Marshall (paper birch, hereinafter birch) (11%). A plan-view diagram of the field experiment is included in Fig. 1. The tree cuttings were harvested with roots intact to the degree possible, then placed upright with roots in shallow water, and preserved in a cool, sheltered environment for 12 h prior to planting on the tailings. Once planted, each tree was watered and tagged for monitoring. The selection of specific tree species for planting was motivated by availability in the surrounding forest ecosystem and the planting design was confirmed to be reflective of the company’s ultimate reforestation plans.

Schematic plan-view of experimental installation on sand-capped tailings
HCWAB and tailings characterization
The HCWAB applied on site was bottom ash acquired from a wood gasification co-energy facility located in Kirkland Lake, Ontario. The HCWAB material was packaged in sealed, industrial 0.75-m3 sacks and transported to site then divided into smaller sacks for weighing and distribution onto each experimental plot. HCWAB material was also collected from each package for immediate laboratory analysis. Prior to characterization, HCWAB was washed and oven dried at 105 °C, and then homogenized. Triplicate samples were analyzed at the University of Toronto (Scarborough, Canada) for total carbon (TC) and nitrogen (TN) through Dumas combustion analysis (C:N 628, LECO Instruments, Canada) and results were verified using a Thermo Flash 2000 (Thermo Fisher Scientific, USA), while exchangeable K+, Mg2+, and Ca2+ were extracted with ammonium acetate and filtered through Fisher P8 filter paper for analysis with atomic absorption spectrometry (AAnalyst200, PerkinElmer, USA). After 2 years, substrate samples were collected from the top 6 cm of substrate at three randomly selected points in every plot using a 5-cm diameter soil corer. The samples were sealed in airtight bags, frozen for transport, then dried, pooled, and homogenized for analysis. Both HCWAB and untreated tailings were measured for pH and electrical conductivity (EC) from a 1:20 (pH) and 1:5 (EC) substrate to water solution (Denver Instruments UB-10 Ultra Basic analyzer fitted with a Bluelab pH electrode). Organic carbon content of HCWAB was evaluated at Activation Laboratories (Ancaster, Ontario): carbonate carbon was removed from subsamples by reaction with HCl in a filtering combustion crucible, and the residual analyzed for non-carbonate carbon by infrared gas analysis (using ELTRA Helios and CS-800 instruments). Triplicate samples of both tailings and HCWAB were analyzed for elemental composition at Activation Laboratories (Al, Ag, As, Au, Ba, Be, Ca, Cd, Cr, Cu, Fe, K, Mg, P, Pb, S, Zn, among others—complete list in Tables S1 and S2) using inductively coupled plasma mass spectrometry (ICP-MS) following digestion with a mixture of hydrochloric, nitric, perchloric, and hydrofluoric acids. Moisture, volatile matter, and ash contents of wood ash biochar were measured at the University of Toronto, Department of Forestry, following standardized methodology (ASTM D1762-84): oven drying at 105 °C, determining loss of weight during 950 °C muffle furnace heating, and measuring residue upon combustion at 750 °C, respectively.
Tree growth and physiological measurements
In September 2017, 1 month following installation and planting, the height and root collar diameter (RCD) of each cutting were measured and recorded. In July 2018 and 2019, survival was inventoried, and tree height, leader length, and RCD were measured on living samples in order to quantify growth trends. Aboveground biomass (AGBM) for each sample was estimated based on species-specific allometric equations by Bond-Lamberty et al. (2002):
where AGBM is the tree aboveground biomass in g, RCD is the tree stem diameter at soil surface in cm, and AGE is the approximate stand age in years. Species-specific values for parameters a, b, c, and d are provided by Bond-Lamberty et al. (2002). Average AGBM at planting across all species was 16.2 ± 0.80 g. Twelve months following HCWAB application, ion exchange resins (Plant Root Simulators: WesternAg Innovations, Saskatoon, Canada) were installed in the twelve plots at a frequency of four per plot (two cation probes, two anion probes). The probes were removed in September 2018, 2 months following installation, and packaged in air-tight sealed plastic bags for shipment and analysis. Ground-level volunteer vegetation cover was inventoried at the same time interval as tree survival (12 and 24 months following HCWAB application). The vegetation cover and community composition were quantified by sub-quadrants of each experimental plot; full method details for vegetation sampling are described in Williams and Thomas (2023a).
Twenty-four months following installation, the final survival inventory was collected and select live tree samples of branches, leaves, and needles were harvested, oven-dried for 36 h at 60 °C, and analyzed for elemental composition through laser-ablation coupled to an ICP-MS instrument (LA ICP-MS) at the Department of Earth Sciences, University of Toronto (St. George). LA ICP-MS analyses were performed with a ESL193 excimer-based laser ablation system (Elemental Scientific Lasers, USA) paired with an Agilent 7900 quadrupole mass spectrometer (Agilent Technologies Inc., USA). The laser was manually positioned at a randomly selected point on each intact sample and then set to execute a two-way, 600-µm laser sweep. External calibration was conducted using a NIST610 with glass matrix; the NIST sample was ablated and analyzed across four distinct linear trajectories with two sweeps mapped prior to and following the branch, needle, and leaf sample sets. All laser ablation ICP-MS analyses were conducted using 43Ca as a standard element measured through electron microprobe analysis (EPMA) (JEOL JXA8230 5-WDS, JEOL, Tokyo, Japan) at the Department of Earth Sciences, University of Toronto. Microprobe measurements were taken at three randomly selected points on every needle/leaf and branch, and sample measurements were averaged prior to statistical analysis.
Statistical analysis
Data from field measurements and laboratory testing were analyzed using the R statistical programming environment (R Core Team 2020). Annual survival of planted wildings was analyzed with a generalized linear mixed effects model with binomial distribution using the glmer function in the lme4 R statistical package; other variables were assessed using linear mixed effects models after confirming data normality and homoscedasticity. Analyses treated HCWAB dosage as the fixed, independent variable and a random block effect was included. Analysis of variance (ANOVA) tests on the mixed effects model were first conducted exclusively with the treatment variable and block effect and then repeated with a model that included initial tree biomass as a covariate. The mixed model ANCOVA analysis was also repeated for survival data applying tree species as a covariate term and a treatment \(\times\) species interaction term added to the generalized linear model. Separately, plot-level cumulative tree survival was applied to a generalized linear model with Gaussian distribution and individual models constructed for each tree species. Yearly change in total and plot-level AGBM (growth) was analyzed as a response variable using the same models as tree survival with a Gaussian distribution. In order to study growth and survival dose-dependence, we fitted the variables to individual 2nd-order polynomial models with initial tree AGBM and species included as covariates. We described polynomial functions by employing the glm function with a quadratic model (including linear covariate terms).
Data acquired from PRS probes and LA ICP-MS analysis were both analyzed by fitting linear mixed models to results for each element measurement individually as a function of HCWAB dosage with a random effect. For all analyses, a random block effect was initially included and tested through model comparison using Akaike’s Information Criterion (AIC); the random effect term was maintained as a factor when it reduced AIC by ≥ 1 unit; otherwise, the term was dropped and data were analyzed as a fixed effects model with one-way ANOVA or ANCOVA.
In addition to dose–response analysis, element bioaccumulation factors (BAFs) were considered; the mean leaf/needle uptake for important plant nutrients and metals were expressed individually with respect to the element’s associated substrate concentration. Substrate concentrations were determined by considering direct dose-specific HCWAB inputs to tailings. The bioaccumulation of select metals and plant nutrients was fitted to linear regression models. Post hoc investigations were conducted with p < 0.05 and Dunnett’s test in the “DescTools” package in R to identify means of each response variable significantly different from untreated controls, with multiple comparison t-tests applied using the “multcomp” package in R (Hothorn et al., 2023) with corrected p-values using the false discovery rate procedure for models with a significant random effects term (Benjamini and Hochberg 1995). Analysis of variance and linear regression models are outlined in supplemental tables.
Volunteer vegetation and tree performance data were collected for sub-divided quadrants in each of the twelve experiment plots. First- and second-year cover data were applied with HCWAB dosage to form distinct linear models. Each model tested the relationship of total cover and treatment dose on (1) final tree survival and (2) overall tree growth by ANCOVA. Significant effects of analysis of covariance results were plotted and linear regression analysis was performed for all data. Separate fitted regression lines of each HCWAB dose category were also plotted for the total surviving trees and respective growth data that had significant (p \(\le\) 0.05) within-dose variation across changes in vegetation cover.
Results
Properties of wood ash and sand-capped tailings
Chemical composition and physical properties of the HCWAB and tailings substrate are detailed in Tables S1 and S2, respectively. The wood ash biochar applied was marginally alkaline (pH = 8.6) and had high ash (78.3 ± 0.46%) and TC content (30.4 ± 1.0%; 18.3 ± 1.1% Corg), qualifying it as a “class 3” biochar based on conditions stipulated by the International Biochar Initiative (IBI) (IBI 2015). HCWAB samples measured above the lower limits of IBI’s allowable toxicant concentration thresholds for use on soil for As (29.0 ± 1.5 ppm), Cd (3.36 ± 0.20 ppm), Cu (145.3 ± 6.1 ppm), Mo (10.2 ± 4.6 ppm), and Zn (511.3 ± 23.3 ppm), all of which are also elements with phytotoxicity potential. The sand-capped tailings substrate initially had near neutral pH (7.48 ± 0.03) and was much lower in TC (0.6 ± 0.07%), TN (trace), and other important plant nutrients such as K (1.38 ± 0.01%), P (0.08 ± 0.001%), Zn (39.67 ± 0.67 ppm), and Ca (2.53 ± 0.03%). The substrate measured higher than the HCWAB with respect to numerous elements posing environmental toxicity risk such as Al (6.44 ± 0.06%), Ce (75.0 ± 4.16 ppm), Co (9.70 ± 0.15 ppm), and Ni (33.90 ± 0.17 ppm). Final plot-level TC analyses suggest that the HCWAB amendments remained stable in the upper 6-cm layer over the 2-year experiment, apart from the mid-range dosage plots; substrate surface layer samples from 12.8 t/ha plots were significantly higher in TC than all other areas. Other responses of substrate physical properties are reported in Table S2, with additional characterization described in Williams and Thomas (2023a).
Survival and growth of planted wildings
At the time of initial planting, the average wilding AGBM was 16.2 g; however, there was considerable variation (SD = 16.4 g, CV = 101%) due in part to among-species size differences (F(3, 416) = 12.2, p < 0.001). Among the four species, pine had the largest AGBM at planting, with samples averaging 18.9 ± 17.1 g. Initial average AGBM of other trees were as follows: 16.7 ± 18.6 g (aspen), 9.7 ± 7.9 g (birch), and 5.0 ± 7.16 g (willow) (Fig. S1). An effort was made to allocate all size classes collected within each experimental plot and, so ultimately no significant variation in initial AGBM was recorded based on plot location (p = 0.73).
The total survival of wildings across all HCWAB doses after 2 years was 31.7 ± 1.28%. After 1 year, HCWAB dosage had a significant effect on the rate of tree survival (F(3,431) = 10.16, p = 0.0174; Fig. 2a); however, there was no significant effect on tree survival after 2 years. Consistently highest survival rates were observed in the 6.4 t/ha and 12.8 t/ha plots across both the first and second years of data collection, with final survival rates at these doses reaching 39.8 ± 2.5% and 33.3 ± 2.3%, respectively. Wildings planted in the highest dosage substrate had lowest survival rates after 1 year and rates equivalent to the control plots after 2 years; average final survival in the highest amendment plots was 26.8 ± 1.3%.

Average tree survival during the 2 years of data collection, (a) separated by HCWAB dosage, and (b) grouped by species. Y-axis in (a) is on ln scale and means ± 1SE are plotted exclusively on the combined total data. Means (± 1SE) (b) denoted by a different lowercase (year 1) or uppercase (year 2) letter indicate significant differences between species (p \(\le\) 0.05) based on post hoc comparisons. Species labels on x-axis refer to the following: BEPA, B. papyrifera; PIBA, P. banksiana; POTR, P. tremuloides; SABE, S. bebbiana
We observed significant species-specific differences in survival across the four tree species after 1 year (F(3,431) = 34.4, p < 0.001) and 2 years (F(3,431) = 19.07, p < 0.001); the survival among birch samples was significantly higher than in pine and aspen after 1 year, but exclusively higher than the survival rates of aspen after 2 years (Fig. 2b). Two years after planting, 16.7 ± 4.7% of aspen samples remained alive compared to 50 ± 8.7% of birch wildings. When each species was considered separately, HCWAB dosage did not display significant effects on survival, though survival of each species was highest in the mid-range HCWAB dosages between 6.4 and 12.8 t/ha (Fig. 3). We considered a binomial second-order polynomial model of the form: y = x − x2 + z, where y is the final tree survival, x is the HCWAB dosage, and z is the tree species. Analysis of variance run on the model found that final year tree survival varied significantly based on both the second-order HCWAB treatment term and species covariate term (χ2(3,126) = 3.89, ptmt = 0.049 and χ2(3,126) = 19.18, pspecies < 0.001).

Proportion of wildings surviving 2 years after planting across all HCWAB dosages applied, distinguished by tree species. Means relate to data from each distinct species and are denoted ± 1SE
Tree survival during each year of data collection also varied significantly based on the initial size at the time of planting (year 1: χ2(3,431) = 20.5, p < 0.001; year 2: F(3,431) = 28.46, p < 0.001). Indeed, the smallest wildings at planting—with individual AGBM measures below 15 g—survived at significantly greater rates compared to trees from the largest initial size class (AGBM > 30 g) (Fig. 4). When considering each species independently, this effect was consistently observed, with survival of pine trees most significantly influenced by initial size (χ2(7,431) = 15.34, p = 0.030; Fig. S2). We described tree species and initial size together by fitting a second-order polynomial of the form: y = x − x2 + z, where y is the proportion of trees alive, x is the pooled tree initial AGBM at time of planting, and z is the tree species. The second-order initial AGBM term (χ2(6,431) = 22.7, p = 0.0009) and tree species (χ2(3,431) = 13.3, p = 0.0040) were both found to have significant effects on final survival.

Survival vs. aboveground biomass (allometric estimates at time of planting) of transplanted saplings measured 2 years after planting. Means are plotted ± 1SE, and values denoted by distinct letters indicate significant differences between size classes (p \(\le\) 0.05) based on post hoc comparisons
The average aboveground biomass per tree declined after 1 year followed by increases throughout the second year in every HCWAB dosage category (Fig. 5a). Overall tree growth across all species measured at the time of harvest averaged 10.33 ± 0.87 g and increased with HCWAB dosage, though treatment effects were not statistically significant (Fig. 6). Rather, HCWAB effects on tree growth varied significantly between species (χ2(1,3) = 5.19, p = 0.0021) and were also dependent on initial size of trees at planting (Z2(6,126) = 3.81, p = 0.05). AGBM averages on a plot-level were slightly higher across all treatment dosages relative to the control plots in both years, and averages across the 6.4 t/ha dosages were greatest compared to all other samples (Fig. 5b). Average total tree growth was modeled with HCWAB treatment and initial AGBM as a second-order polynomial function; initial tree size displayed a significant growth effect (χ2(6,431) = 22.7, p = 0.0009).

Average (a) tree-level and (b) plot-level aboveground biomass production measured 2 years after planting; means are plotted ± 1SE. Data is distinguished by (a, b) HCWAB treatment dosage and (c, d) totaled across all dosages

Total growth of live wildings 2 years after planting grouped by HCWAB dosages applied (a) for all live trees and (b) distinguished by tree species. All means are denoted ± 1SE
Ion supply and total carbon
The supply rate of plant nutrients measured by PRS probes did not vary significantly among HCWAB dosages (Table 1); however, nutrients detected by the probes were consistently highest in the mid-range treatment plots (6.4 and 12.8 t/ha). Tailings amended at 6.4 t/ha showed maximum average levels of P, Fe, and Zn ions, while the greatest Ca, K, and Mg supply was measured in tailings with the 12.8 t/ha HCWAB treatment (Fig. S3). We did not observe significant treatment effects on either Total N or NO3− ion supply; however, average nitrate was less than in unamended plots across all HCWAB dosages and declined consistently with increasing HCWAB fitting a 3-parameter log-logistic function (Fig S3b). In the high 19.1 t/ha HCWAB treatment, a 76.8 ± 7.0% decrease in nitrate was measured relative to the control plots. We did not observe statistically significant ion supply dose response from the range of metals and elements of emerging toxicity concern. Average rates of Cu and Cd were highest in both 6.4 t/ha and 12.8 t/ha dosage plots, whereas the supply of Pb was highest in the 12.8 and 19.1 t/ha treatments. The measures for other potentially phytotoxic ions—namely Al, B, and Mn—were inconsistent across treatments.
Total substrate carbon measured 2 years following experimental installation varied significantly with HCWAB dosage (F(1,34) = 6.51, p = 0.015), with greater levels of TC measured in each HCWAB-amended substrate relative to control plots. Measurements of TC were significantly higher in plots with 12.8 t/ha HCWAB doses. Neither plot-level tree survival or initial size (AGBM) could explain variation in TC (F(1,11) = 1.55, psurv = 0.244; F(1,11) = 1.17, psize = 0.304).
Tree tissue chemistry
Concentrations of stored nutrients and PTEs in tree tissues did not exhibit significant effects from HCWAB treatments; however, the average needle and leaf concentrations of several elements were higher across all HCWAB treatments compared to the tissue concentrations from control plots, specifically: Mo, Ba, Cu, As, Pb, and La. Average needle concentrations of important plant nutrients (K, P, Zn, and Mg) were inconsistent across the samples (Table 2). The treatment effect on Fe tissue concentrations was not statistically significant, still Fe levels measured in needle and leaf samples increased with higher HCWAB dosage. The Fe bioaccumulation factor was plotted and fit to a linear regression, which displayed a significant negative relationship between concentrations in tree needles vs, substrates (R2adj = 0.946, F(1,4) = 53.76, p = 0.0181, Fig. 7). All elements measured in tree branch samples were consistently lower than in corresponding tree needle and leaf samples but did not display any significant HCWAB treatment effect. Otherwise, substrate elemental content was not a strong predictor of associated needle or leaf tissue storage based on bioaccumulation factors (Table S3).

Fe needle and leaf concentrations related to (a) Fe concentration in the experimental substrates and (b) HCWAB dosage applied, and (c) Fe branch concentrations related to HCWAB dosage applied. Means representing ± 1SE are plotted
Volunteer vegetation and tree responses
As reported in a prior publication (Williams and Thomas 2023a), volunteer vegetation on the site increased significantly in response to HCWAB additions, with both 12.8 t/ha and 19.1 t/ha plots displaying significantly more cover relative to the control plots. Early (first year) volunteer vegetation was a significant predictor of final tree survival (F(1,47) = 5.51, p = 0.0232, Fig. 8a), with lower survival rates associated with areas of higher vegetation cover. This interactive effect was significant across all data, but most pronounced in the control plots and 12.8 t/ha treatments. Tree growth 2 years following the planting was also significantly lower in quadrants with higher first-year volunteer cover (F(1,47) = 7.85, p = 0.0074, Fig. 8b). In addition, there was an interactive effect of volunteer vegetation and HCWAB dosage on final tree growth, with a more pronounced negative growth effect of volunteer vegetation measured in the two highest HCWAB treatments (12.8 t/ha: F(1,47) = 7.25, p = 0.0226; 19.1: F(1,47) = 5.53, p = 0.0406).

Mean volunteer cover after 1 year related to with (a) mean final tree survival and (b) mean final growth. Regression lines plotted represent total data and exclusively HCWAB dosages with significant (p ≤ 0.05) interaction effects on survival. Points are color-coded by HCWAB dose: 0 t/ha (red), 6.375 t/ha (blue), 12.75 t/ha (green), 19.1 t/ha (magenta). Significant regression relationships are as follows: (a) all doses: y = − 0.9x + 38.44, adjR2 = 0.088, p = 0.0232; control (0 t/ha): y = − 1.75x + 38.9, adjR2 = 0.329, p = 0.030; 12.75 t/ha: y = − 1.43x + 42.2, adjR2 = 0.341, p = 0.0272 and (b) all doses: y = − 1.57x + 37.77, adjR2 = 0.127, p = 0.00742; 12.75 t/ha: y = − 2.19x + 42.87, adjR2 = 0.362, p = 0.0226; 19.1 t/ha: y = − 2.14x + 43.96, adjR.2 = 0.292, p = 0.041
Discussion
Results from our 2-year field experiment on sand-capped mine tailings revealed that applying HCWAB led to higher survival of planted, native wildings at mid-range application rates. However, tree survival declined at dosages over 12.8 t/ha, above which survival closely matched that on unamended substrate for most species. The specific species and initial size of wildings, more so than HCWAB amendment dosage, were most important in determining final tree survivorship and growth. On average, peak pine, birch, and willow survival was observed in the mid-range HCWAB dosages (6.4–12.8 t/ha). In contrast, more aspen wildings planted on control plots survived after 2 years compared to HCWAB-amended plots. Average aboveground biomass production of planted trees was higher in all HCWAB doses compared to untreated plots. Smaller planted trees (initially < 15 g AGBM) survived at higher rates compared to larger size classes, a pattern most apparent in pine. Overall, survivorship was highest in birch and lowest in aspen.
HCWAB additions did not significantly alter substrate availability of potentially toxic metals (Cd, Cu, Al), while the availability of select important plant nutrients (P, Zn, Fe) was higher in amended tailings at all dosages relative to levels in the tailings alone. Our analyses did not reveal significant treatment effects on tree tissue chemistry, with concentrations of potential toxic elements remaining well below toxicity limits for plants and food chain transfer. Taken together, the improved status of available plant nutrients in amended plots combined with limited effects on tree tissue contaminant uptake indicates direct HCWAB benefits for planted trees. However, our results also suggest that changes in tree survival and growth across the site depended on interactive effects of species, initial tree size, and exposure to induced competition on HCWAB-amended plots.
Tree survival and growth
Of the four species planted, pine displayed the highest average growth after 2 years across all treatment doses. Average growth in birch was greatest in high (19.1 t/ha) HCWAB plots, while growth of pine peaked at 12.8 t/ha. In contrast, both aspen and willow exhibited highest growth in the lowest treatment dosage and their growth decreased with additional HCWAB application. Our results correspond to the findings of Bélanger et al. (2021), who applied fly ash on two distinct northern Canadian boreal sites and reported a positive linear growth response in planted jack pines with increasing ash dosages up to 14 t/ha. Emilson et al. (2019)’s review of eight field experiments across Canada found a species-specific tree growth effect of wood ash and highest increases in jack pines compared to other native conifers (white, hybrid, and black spruce). Although Emilson et al.’s results were independent of the stand development stage, Brais et al. (2015) found no growth increases from 5 t/ha application of wood ash to a mature jack pine stand, which suggests jack pine growth responses could be accentuated in younger trees. Jack pines are a fire-adapted, pioneer species and rapidly acquire resources from the proximate environment (Rudolph and Laidly 1990). The jack pine wildings planted on our site may have profited from the increases in available nutrients at higher HCWAB dosages. A study in natural stands near the present field trial found large growth responses of jack pine saplings to variation in pyrogenic carbon from natural fire residues, with a peak response at ~ 30–60 t/ha (Gale and Thomas 2021). In the present study, the AGBM results after 2 years suggest that surviving jack pines rapidly acquired resources from their adjacent substrate, resulting in a higher relative growth. The variation in total carbon measured across plots after 2 years mirrored that of jack pine growth, increasing with HCWAB dosage until a peak at 12.8 t/ha.
Despite increased growth responses, the proportion of surviving pines after 2 years was relatively low and did not display a significant response to HCWAB dosage. Our results are consistent with the findings of Barette et al. (2022), who planted four tree species—jack pine, paper birch, tamarack, and hybrid willow—on boreal gold mine tailings and reported lowest survival rates among the jack pine samples. At a historic asbestos mine in Québec, Grimond et al. (2023) also observed lowest survival of young jack pines compared to five other tree species planted on both waste rock and tailings-derived technosols. Low survival in jack pines is commonly attributed to substrate pH, as the species benefits from slightly acidic soil conditions (Rudolph and Laidly 1990). Substrate pH across our experimental plots ranged from 7.3 to 7.5, likely above optimal for jack pine (Zhang et al. 2015). Although volunteer jack pines are commonly found on newly exposed, moisture- and nutrient-poor sites, they may be succeeded by more selective species, particularly if substrate conditions improve (Rudolph and Laidly 1990). On loamy soils in Canada, jack pines are naturally succeeded by paper birch, and results from our experiment site suggest this succession pattern in the high-dosage HCWAB plots. Indeed, the high overall survival among planted paper birch wildings supports a more widespread use of the species for boreal tailings reforestation. The survival of aspen remained below 20% regardless of substrate treatment, opposing our initial hypothesis and suggesting this species may be less suitable for tailings reforestation.
Substrate ion supply
The supply of bioavailable nutrients measured by ion exchange resin probes did not exhibit significant dose-specific responses from the HCWAB; however, some important plant nutrients were consistently higher in all treated plots compared to the controls. Specifically, the average supply rates of P, Fe, Cu, and Zn were highest in HCWAB-treated plots. Recent reports on wood ash applied to soils have described an initial rise in substrate and foliar P due to increased soil pH and phosphate-solubilizing microorganisms (Omil et al. 2013). We similarly observed higher foliar P levels in amended plots, which may be a consequence of the initial low-P status of the site. Prior experiments in boreal forests have tested wood ash additions at dosages of 4–8 t/ha and reported increased supply of important plant nutrients in amended substrates relative to controls (Deighton et al. 2021; Bieser and Thomas 2019; Noyce et al. 2016). On historic gold mine tailings in the boreal, higher HCWAB dosages (> 9 t/ha) resulted in significant increases in substrate availability of Ca, P, Zn, and K, as well as a decrease in substrate Fe (Williams and Thomas 2023b). These results are compatible with the dissolution of salts and release of carbonates from compounds on the wood ash surficial layers (Ludwig et al. 2002). Recent studies have established that wood ash’s effects on soil nutrient availability relate to direct nutrient inputs, as well as induced changes in soil pH and microbial activity (Demeyer et al. 2001). Indeed, lower solubility of wood ash P has been observed on substrates with higher initial pH (Park et al. 2005), and this likely explains the relatively minor increases on the neutral pH tailings at our experiment site.
The supply of total soil nitrogen (TN) and nitrate ions measured was lower in HCWAB-amended plots, with lowest values measured at the highest HCWAB dosage. Nitrogen in wood feedstock is converted to gaseous forms during pyrolysis (Winter et al. 1999; Pitman 2006), resulting in a high-C, low-N product. Kameyama et al. (2016) found that nitrate sorption by biochars in soils was highest for wood chars produced at high temperatures (above 800 °C). Our results corroborate these findings, as well as those from HCWAB application on N-limited historic mine tailings (Williams and Thomas 2023b) and on a boreal clearcut site (Bieser and Thomas 2019). Our field results also underscore observations made by Saarsalmi et al. (2010); in N-limited ecosystems, char-amended substrates may require additional N input. Other reports, such as those by Ferreiro et al. (2011) and Sifton et al. (2022), have proposed combining application of wood ash or biochars with N-fixing companion plants to maximize substrate supply available for trees. However, there is need for additional field studies to test this approach on N-limited mine tailings.
Applying HCWAB for the purpose of soil nutrient and pH improvements may also introduce excess toxic elements into the soil and plant ecosystem, and this could increase the likelihood of phytotoxicity or transfer to the animal food chain. Tailings amended with HCWAB from our experiment site did not display significant increases in phytotoxic elements (i.e., Al, B, Pb, S, and Cd). These findings agree with a number of prior incubation and field studies on wood ash to northern forest soils (Perkiömäki et al. 2003; Pugliese et al. 2014; Saarsalmi et al. 2004). Our previous field study on historic tailings found significant increases in available Zn and Pb in HCWAB-amended plots, but only at higher application rates of 30 t/ha (Williams and Thomas 2023b). Maintained or reduced availability of PTEs following wood ash application to substrates has been attributed to the ash’s pH-neutralizing effect (Augusto et al. 2008); however, further field experiments are required to understand whether PTEs from wood ash may be released into the soil system at higher application rates or on highly acidic/acid-generating mine tailings.
Nutrient and PTE uptake in tree tissues
We did not detect significant HCWAB addition effects on tree tissue nutrient levels; however, the concentration of K, Mg, P, Fe, and Zn was marginally higher in foliage samples from amended plots compared to the control samples, and Fe levels increased in branch tissues (Table 3, Fig. 7). A number of prior studies have likewise found little effect of wood ash additions on foliar nutrients (e.g., Saarsalami et al. 2004; Omil et al. 2013); however, a meta-analysis of 28 trials found that wood ash amendments on mineral soils significantly increased foliar K and P in early years after application (Augusto et al. 2008). The increase in tissue Fe observed on site is similar to that reported in Pinus laevigata planted on biochar-amended tailings by Ramírez-Zamora et al. (2022).
We did not find significant HCWAB effects on PTE tissue concentrations for either needle or branch samples, though certain elements of concern—La, As, Mn, Ba, Cu, and Mo—were detected at marginally higher levels in tissues from all amended plots relative to those from control tailings. Maximum concentrations of La, As, Mn, Ba, and Cu were measured in the 12.8 t/ha plot samples, whereas highest amounts of Mo were found in the samples from 6.4 t/ha treatments. Compared to other wood ashes, those produced from wood gasification often contain substantially elevated levels of dangerous metal/loids, such as Cd (Augusto et al. 2008), Pb, and As (Lebrun et al. 2017), as well as micronutrients that may be present in excess, such as Mn and Ba (Bélanger et al. 2021). However, few field studies have reported on phytoabsorption of PTEs from wood ash amendments. In boreal forest plantations, wood ash additions led to no significant changes in Mn and Ba needle concentrations in jack pine (Bélanger et al. 2021); declines in conifer needle Mn levels as a response to wood ash have also been observed (Saarsalmi et al. 2004). Arsenic is a particular concern, and the bioavailability of As ions in substrates increases with higher pH (Beesley et al. 2011; Lebrun et al. 2017). In a previous field trial on historical tailings, we detected increased As levels in jack pine needles, but only at the highest HCWAB dosage tested (30 t/ha) (Williams and Thomas 2023b). In the present study, slightly elevated As levels in needle/leaf tissues from HCWAB addition treatments were not statistically significant (Fig. S2, Table 2). Results to date suggest that tree tissue accumulation of As is likely a result of high dosage or repeated wood ash applications, or from HCWAB used concurrent to alkalinizing treatments such as liming.
Effects of volunteer vegetation
We observed significant increases in volunteer vegetation cover (Williams and Thomas 2023a) and a rise in soil TC levels in substrates on the higher dosage plots (12.8 t/ha and 19.1 t/ha). The large cover increase was correlated with reduced tree survival and growth (Fig. 8a, b). Our results thus suggest that the rapid, early response of volunteer vegetation amplified resource competition in treated plots and ultimately diminished the benefit of HCWAB for the planted trees. Reports from other field experiments in the boreal describe similar responses in volunteer vegetation, with particularly important effects in nitrogen-fixing species, attributed to higher soil available K in treated plots (Bieser and Thomas 2019; van de Voorde et al. 2014). Prior studies have found competition from select nitrogen-fixing species to be especially detrimental for jack pine survivorship (Barette et al. 2022; Robinson et al. 2014), though N-fixing nurse species such as alder can also have facilitative effects on jack pine growth in some cases (Thiffault and Hébert 2017). In the present experiment, we measured notable positive responses to HCWAB of overall vegetation cover and nitrogen-fixing legumes, including increases in the competitive, invasive species Melilotus spp., Medicago sativa, and Crepis tectorum (Williams and Thomas 2023a).
HCWAB application strategies
Use of native vegetation cover for mine tailings restoration has been associated with improvements in substrate structure, microporosity, and fertility (Tordoff et al. 2000). However, early growth of competitive understory plants may slow forest succession on tailings sites (Franklin et al. 2012; Macdonald et al. 2015), including those in the Canadian boreal (Guittonny-Larchevêque et al. 2016). The HCWAB applied on our experiment site introduced important plant nutrients, but these were likely rapidly taken up by understory ruderal vegetation and thus diverted away from the planted trees. Considerations of complex ecosystem-wide interactions and possible contraindications for HCWAB application should be a focus of future tailings reforestation research. Applying wood ash just prior to tree seeding or planting facilitates an even ash distribution and may minimize the risk of physical damage to young transplants (Hannam et al. 2018), but may not result in significant nutrient acquisition by trees (Stupak et al. 2016), suggesting a possible strategy of repeated applications.
Applying HCWAB together with N-fixing nurse plants may compensate for the low available nitrogen typically observed in soil with biochar (Sifton et al. 2022). N-fixing plants can enhance N status of proximate soils and surrounding vegetation, including planted trees (Munroe and Isaac 2014); biochar can then potentially enhance facilitative interactions by retaining fixed N in mixed-species systems (Thomas et al. 2019). The co-planting of N-fixing vegetation and tree saplings facilitate beneficial interactions, particularly when native companion species are prioritized over non-native, competitive plants (Sifton et al. 2022). In the context of tailings restoration, nitrogen is among the most limiting soil nutrients, and prior studies have noted relatively strong performance of N-fixing plants on tailings (e.g., Cross et al. 2021); however, the strategic use of native N-fixing companion plants for tailings reforestation appears not to have been explored.
The lack of HCWAB effects on foliar elements in tree samples from our site suggests wood ash biochars could be used at dosages below 20 t/ha without adverse accumulation in trees or any detrimental food chain exposure. However, the early increased supply of available nutrients and PTEs measured on amended plots, albeit marginal, may indicate accumulation in herbaceous vegetation. As is the case with other types of biochar, HCWAB is likely to sorb or otherwise immobilize PTEs while influencing substrate permeability and hydraulic conductivity (Beesley et al. 2011). There is thus a need for further trials that investigate more comprehensively the fate of PTEs on HCWAB-amended mine tailings, with the goal of developing ameliorative and adaptive long-term reclamation strategies.
Conclusion
The experimental trial described addresses a critical knowledge gap regarding in situ effects of HCWAB additions for remediation of sand-capped thickened tailings, this being the prevalent modern practice. Over two growing seasons, the survival of planted wildings was highest on tailings amended with 6.4–12.8 t/ha of HCWAB, and final tree growth increased slightly but steadily with HCWAB rate. The supply of important plant nutrients, including P, K, Zn, and Mg, was highest in the mid-level HCWAB dosage range, while no significant treatment responses were observed for potentially toxic elements. Of the four native tree species planted, birch survived at significantly higher rates independent of the HCWAB treatment and transplanted saplings with the lowest initial aboveground tree biomass survived at higher rates. Tree performance was negatively correlated with volunteer vegetation cover, which increased at the higher HCWAB dosage plots, consistent with increased HCWAB additions exacerbating competition. Findings from this field trial demonstrate the value of low to mid-level HCWAB treatment on mine tailings for planted tree performance and supply of important plant nutrients. Further investigations are required to establish optimal timing of HCWAB application and tree planting, and potential use of co-amendments and companion herbaceous species that would enhance facilitative rather than competitive effects.
Data Availability
Data for this article can be found in the University of Toronto data repository located at: https://doi.org/10.5683/SP3/UFAQ5C.
References
Augusto L, Bakker MR, Meredieu C (2008) Wood ash applications to temperate forest ecosystems – potential benefits and drawbacks. Plant Soil 306:181–198
Barette D, Marchand P, Nguena HLN, Guittonny M (2022) The effects of agronomic herbaceous plants on the soil structure of gold mine tailings and the establishment of boreal forest tree seedlings. Water Air Soil Pollut 233:16. https://doi.org/10.1007/s11270-021-05466-9
Beesley L, Marmiroli M (2011) The immobilization and retention of soluble arsenic, cadmium and zinc by biochar. Environ Pollut 159:474–480
Beesley L, Moreno-Jiménez E, Gomez-Eyles JL, Harris E, Robinson B, Sizmur T (2011) A review of biochars’ potential role in the remediation and restoration of contaminated soils. Environ Pollut 159:3269–3282
Bélanger N, Ponce GO, Brais S (2021) Contrasted growth response of hybrid larch (Larix × marschlinsii), jack pine (Pinus banksiana) and white spruce (Picea glauca) to wood ash application in northwestern Quebec. Canada Iforest 14:155–165
Benjamini Y, Hochberg Y (1995) Controlling the false discovery rate: a practical and powerful approach to multiple testing. J Roy Stat Soc B 57:289–300
Biederman LA, Harpole WS (2013) Biochar and its effects on plant productivity and nutrient cycling: a meta-analysis. GCB Bioenergy 5:202–214. https://doi.org/10.1111/gcbb.12037
Bieser JMH, Thomas SC (2019) Biochar and high-carbon wood ash effects on soil and vegetation in a boreal clearcut. Can J for Res 49:1124–1134
Bond-Lamberty B, Wang C, Gower ST (2002) Aboveground and belowground biomass and sapwood area allometric equations for six boreal tree species of northern Manitoba. Can J for Res 32:1441–1450
Bondu R, Cloutier V, Rosa E, Benzaazoua M (2017) Mobility and speciation of geogenic arsenic in bedrock groundwater from the Canadian Shield in western Québec, Canada. Sci Total Environ 574:509–519
Borgegard S, Rydin H (1989) Biomass, root penetration and heavy metal uptake in birch in a soil cover over copper tailings. J Appl Ecol 26:585–595
Brais S, Bélanger N, Guillemette T (2015) Wood ash and N fertilization in the Canadian boreal forest: soil properties and response of jack pine and black spruce. For Ecol Manage 348:1–14
Natural Resources Canada (NRCan) (2016) Improving automation and equipment, Natural Resources Canada. http://www.nrcan.gc.ca/mining-materials/green-mining/18278. Accessed 15 September 2023
Environment Canada (2023) Canadian climate normal 1981–2021 station data, Pickle Lake A Ontario. Government of Canada. https://climate.weather.gc.ca/climate_data/. Accessed 15 Jan 2023
Chisholm RA, Muller-Landau HC, Rahman KA et al (2013) Scale-dependent relationships between tree species richness and ecosystem function in forests. J Ecol 101:1214–1224
Clapham WM, Zibilske LM (1992) Wood ash as a liming amendment. Commun Soil Sci Plant Anal 23:1209–1227
Cross AT, Ivanov D, Stevens JC, Sadler R, Zhong H, Lambers H, Dixon KW (2021) Nitrogen limitation and calcifuge plant strategies constrain the establishment of native vegetation on magnetite mine tailings. Plant Soil 461:181–201
Deighton HD, Watmough SA, Basiliko N, Hazlett PW, Reid CR, Gorgolewski A (2021) Trace metal biogeochemical responses following wood ash addition in a northern hardwood forest. Can J for Res 51:817–833
Demeyer A, Voundi Nkana JC, Verloo MG (2001) Characteristics of wood ash and influence on soil properties and nutrient uptake: an overview. Biores Technol 77:287–295
DeVolder PS, Brown SL, Hesterberg D, Pandya K (2003) Metal bioavailability and speciation in a wetland tailings repository amended with biosolids compost, wood ash, and sulfate. J Environ Qual 32:851–864
Emilson CE, Bélanger N, Brais S, Chisholm CE, Diochon A, Joseph R, Markham J, Morris D, Van Rees K, Rutherford M, Venier LA, Hazlett PW (2019) Short-term growth response of jack pine and spruce spp. to wood ash amendment across Canada. GCB Bioenergy 12:158–167
Fellet G, Marchiol L, Delle Vedove G, Peressotti A (2011) Application of biochar on mine tailings: effects and perspectives for land reclamation. Chemosphere 83:1262–1267
Ferreiro A, Merino A, Díñaz N, Piñeiro J (2011) Improving the effectiveness of wood-ash fertilization in mixed mountain pastures. Grass Forage Sci 66:337–350
Foy CD, Chaney RL, White MC (1978) The physiology of metal toxicity in plants. Ann Rev Plant Physiol 29:511–566
Franklin JA, Zipper CE, Burger JA, Skousen JG, Jacobs DF (2012) Influence of herbaceous ground cover on forest restoration of eastern US coal surface mines. New for 43:905–924
Gale NV, Thomas SC (2021) Spatial heterogeneity in soil pyrogenic carbon mediates tree growth and physiology following wildfire. J Ecol 109:1479–1490
Gill KS, Malhi SS, Lupwayi NZ (2015) Wood ash improved soil properties and crop yield for nine years and saved fertilizer. J Agricult Sci 7(12). https://doi.org/10.5539/jas.v7n12p72
Grimond L, Rivest D, Bilodeau-Gauthier S, Khlifa R, Elferjani R, Bélanger N (2023) Novel soil reconstruction leads to successful afforestation of a former asbestos mine in southern Quebec, Canada. New Forests 123. https://doi.org/10.1007/s11056-023-09989-3
Guittonny-Larchevêque M, Meddeb Y, Barrette D (2016) Can graminoids used for mine tailings revegetation improve substrate structure? Botany 94:1053–1061
Hannam KD, Venier L, Allen D, Deschamps C, Hope E, Jull M, Kwiaton M, McKenney D, Rutherford PM, Hazlett PW (2018) Wood ash as a soil amendment in Canadian forests: what are the barriers to utilization? Can J for Res 48:442–450
Hannam KD, Deschamps C, Kwiaton M, Venier L, Hazlett PW (2016) Regulations and guideline for the use of wood ash as a soil amendment in Canadian forests. Canadian Forest Service Information Report GLC-X-18. Natural Resources Canada, Canadian Forest Service. http://cfs.nrcan.gc.ca/pubwarehouse/pdfs/37781.pdf. Accessed 20 Oct 2023
Hansen M, Bang-Andreasen T, Sorensen H, Ingerslev M (2017) Micro vertical changes in soil pH and base cations over time after application of wood ash on forest soil. For Ecol Manage 406:274–280
Hope ES, McKenney DW, Allen DJ, Pedlar JH (2017) A cost analysis of bioenergy-generated ash disposal options in Canada. Can J for Res 47:1222–1231. https://doi.org/10.1139/cjfr-2016-0524
Hothorn T, Westfall P, Heiberger RM, Schuetzenmeister A, Scheibe S (2023) Package “multcomp.” Simultaneous inference in general parametric models, Project for Statistical Computing, Vienna, Austria
Huotari N, Tillman-Sutela E, Moilanen M, Laiho R (2015) Recycling of ash–for the good of the environment? For Ecol Manage 348:226–240
Hytönen J (2003) Effects of wood, peat and coal ash fertilization on Scots pine foliar nutrient concentrations and growth no afforested former agricultural peat soil. Silva Fennica 37:219–234
International Biochar Initiative (IBI) (2015) Standardized product definition and product testing guidelines for biochar that is used in soil (IBI Biochar Standards) Version 2.1. https://biochar-international.org/characterizationstandard/. Accessed 15 January 2020
Jacobson S, Lundström H, Nordlund S, Sikström U, Pettersson F (2014) Is tree growth in boreal coniferous stands on mineral soils affected by the addition of wood ash? Scan J for Res 29:675–685
James AK, Helle SS, Thring RW, Sarolia GS, Rutherford PM (2014) Characterization of inorganic elements in woody biomass bottom ash from a fixed-bed combustion system, a downdraft gasifier and a wood pellet burner by fractionation. Energy Environ Res 4:85–94
Joseph S, Cowie AL, Van Zwieten L, Bolan N, Budai A, Buss W, Cayuela ML et al (2021) How biochar works, and when it doesn’t: a review of mechanisms controlling soil and plant responses to biochar. GCB Bioenergy 13:1731–1764
Kahl JS, Fernandez IJ, Rustad LE, Peckenham J (1996) Threshold application rates of wood ash to an acidic forest soil. J Environ Qual 25:220–227
Kam S, Girard J, Hmidi N, Mao Y, Longo S (2011) Thickened tailings disposal at Musselwhite Mine. In: Jewell R, Fourie AB (eds) Paste 2011: proceedings of the 14th international seminar on paste and thickened tailings. Australian Centre for Geomechanics, Perth, Australia, pp 225–236. https://doi.org/10.36487/ACG_rep/1104_21_Kam
Kameyama K, Miyamoto T, Iwata Y, Shiono T (2016) Influences of feedstock and pyrolysis temperature on the nitrate adsorption of biochar. Soil Science and Plant Nutrition 62:180–184
Kuttner BG, Thomas SC (2017) Interactive effects of biochar and an organic dust suppressant for revegetation and erosion control with herbaceous seed mixtures and willow cuttings. Restor Ecol 25:367–375
Kuuluvainen T, Nummi P (2023) Strategies for the ecological restoration of the boreal forest facing climate change. In: Girona MM, Morin H, Gauthier S, Bergeron Y (eds) Boreal forests in the face of climate change. Adv Global Change Res 74. Springer, Cham. https://doi.org/10.1007/978-3-031-15988-6_17
Larchevêque M, Desrochers A, Bussière B, Cimon D (2014) Planting trees in soils above non-acid-generating wastes of a boreal gold mine. Écoscience 2:217–231
Lebrun M, Macri C, Miard F, Hattab-Hambli N, Motelica-Heino M, Morabito D, Bourgerie S (2017) Effect of biochar amendments on As and Pb mobility and phytoavailability in contaminated mine technosols phytoremediated by Salix. J Geochem Explor 182:149–156
Lebrun M, Van Pouke R, Miard F, Scippa GS, Bourgerie S, Morabito D, Tack FMG (2019) Effects of carbon-based material and redmuds on metal(loid) immobilization and growth of Salix dasyclados Wimm. on a former mine Technosol contaminated by arsenic and lead. Land Degrad Dev 32:467–481
Lehmann J, Joseph S (2015) Biochar for environmental management: an introduction. In: Lehmann J, Joseph S (eds) Biochar for environmental management: science, technology and implementation, 2nd edn. Earthscan, London, pp 1–14
Liu H, Teng Y, Zheng N, Liu L, Yue W, Zhai Y, Yang J (2022) Field experiments of phyto-stabilization, biochar-stabilization, and their coupled stabilization of soil heavy metal contamination around a copper mine tailing site. Inner Mongolia Minerals 12:702
Ludwig B, Rumpf S, Mindrup M, Meiwes KJ, Khanna PK (2002) Effects of lime and wood ash on soil-solution chemistry and nutritional status of a pine stand in northern Germany. Scad J for Res 17:225–237
Macdonald SE, Lanhausser SM, Skousen J, Franklin J, Frouz J, Hall S, Jacobs DF, Quideau S (2015) Forest restoration following surface mining disturbance: challenges and solutions. New for 46:703–732
Marsh C, Blankinship JC, Hurteau MD (2023) Effects of nurse shrubs and biochar on planted conifer seedling survival and growth in a high-severity burn patch in New Mexico, USA. For Ecol Manage 537:120971
Mendez MO, Maier RM (2007) Phytoremediation of mine tailings in temperate and arid environments. Rev Environ Sci Biotechnol 7:47–59
Mohan D, Pittman CU Jr, Bricka M, Smith F, Yancey B, Mohammad J, Steele PH, Alexandre-Franco MF, Gómez-Serrano V, Gong H (2007) Sorption of arsenic, cadmium and lead by chars produced from fast pyrolysis of wood and bark during bio-oil production. J Colloid Interface Sci 310:57–73
Moilanen M, Silfverberg K, Hokkanen TJ (2002) Effects of wood-ash on the tree growth, vegetation and substrate quality of a drained mire: a case study. For Ecol Manage 171:321–338
Mosseler A, Major JE, Labrecque M (2014) Growth and survival of seven native willow species on highly disturbed coal mine site in eastern Canada. Can J for Res 44:340–349
Munroe JW, Isaac ME (2014) N2-fixing trees and the transfer of fixed-N for sustainable agroforestry: a review. Agron Sustain Dev 34:417–427
Nelson NO, Agudelo SC, Yuan W, Gan J (2011) Nitrogen and phosphorus availability in biochar-amended soils. Soil Sci 176:218–226
Norström SH, Bylund D, Vestin JLK, Lundström US (2012) Initial effects of wood ash application to soil and soil solution chemistry in a small, boreal catchment. Geoderma 187–188:85–93
Noyce GL, Fulthorpe R, Gorgolewski A, Hazlett P, Tran H, Basiliko N (2016) Soil microbial responses to wood ash addition and forest fire in managed Ontario forests. Appl Soil Ecol 107:368–380
Omil B, Pineiro V, Merino A (2013) Soil and tree responses to the application of wood ash containing charcoal in two soils with contracting properties. For Ecol Manage 295:199–212
Osman KT (2012) Chemical properties of soil. In: Soils. Springer, Dordrecht, pp 97–111. https://doi.org/10.1007/978-94-007-5663-2_8
Park BB, Yanai RD, Sahmn JM, Lee DK, Abrahamson LP (2005) Wood ash effects on plant and soil in a willow bioenergy plantation. Biomass Bioenerg 28:355–365
Perkiömäki J, Kiikkilä O, Moilanen M, Issakainen J, Tervahauta A, Fritze H (2003) Cadmium-containing wood ash in a pine forest: effects on humus microflora and cadmium concentration in mushrooms, berries, and needles. Can J for Res 33:2443–2451
Peterson EB, Peterson NM (1996) Ecology and silviculture of trembling aspen. Forestry Chronicle 5:28–44
Pitman RM (2006) Wood ash use in forestry – a review of the environmental impacts. Forestry 79:563–588
Pugliese S, Jones T, Preston MD, Hazlett P, Tran H, Basiliko N (2014) Wood ash as a forest soil amendment: the role of boiler and soil type on soil property response. Can J Soil Sci 94:621–634
R Core Team (2020) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Ramírez-Zamora J, Mussali-Galante P, Rodríguez A, Castrejón-Godínez ML, Valencia-Cuevas L, Tovar-Sánchez E (2022) Assisted phytostabilization of mine-tailings with Prosopis laevigata (Fabaceae) and biochar. Plants 11:3441
Reid C, Watmough SA (2014) Evaluating the effects of liming and wood-ash treatment on forest ecosystems through systematic meta-analysis. Can J for Res 44:867–885
Reid C, Bécaert V, Aubertin M, Rosenbaum RK, Deschênes L (2009) Life cycle assessment of mine tailings management in Canada. J Clean Prod 17:471–479
Robinson DE, Wagner RG, Bell FW, Swanton CJ (2014) Photosynthesis, nitrogen-use efficiency, and water-use efficiency of jack pine seedlings in competition with four boreal forest plant species. Can J for Res 31:2014–2025
Román-Dañobeytia F, Cabanillas F, Lefebvre D, Farfan J, Alferez J, Polo-Villanueva F, Llacsahuanga J, Vega CM, Velasquez M, Corvera R, Condori E, Ascorra C, Fernandez LE, Silman MR (2021) Survival and early growth of 51 tropical tree species in areas degraded by artisanal gold mining in the Peruvian Amazon. Ecol Eng 159:106097
Rudolph PO (1958) Reaction to competition. In: Service USF (ed) Silvical characteristics of jack pine (Pinus banksiana). United States Department of Agriculture, Washington DC, pp 21–22
Rudolph TD, Laidly PR (1990) Pinus banksiana lamb. Burns RM, Honkala BH (tech coords) Silvics of North America: conifers, 2nd edn. United States Department of Agriculture, Washington DC, pp 280–293
Saarsalmi A, Mälkönen E, Kukkola M (2004) Effect of wood ash fertilization on soil chemical properties and stand nutrient status and growth of some coniferous stands in Finland. Scand J for Res 19:217–233
Saarsalmi A, Smolander A, Kukkola M, Arola M (2010) Effect of wood ash and nitrogen fertilization on soil chemical properties, soil microbial processes, and stand growth in two coniferous stands in Finland. Plant Soil 331:329–340
Salam MMA, Mohsin M, Kaipiainen E, Villa A, Kuittinen S, Pulkkinen P, Pelkonen P, Pappinen A (2019) Biomass growth variation and phytoextraction potential of four Salix varieties grown in contaminated soil amended with lime and wood ash. Int J Phytorem 21:1329–1340
Sifton MA, Lim P, Smith SM, Thomas SC (2022) Interactive effects of biochar and N-fixing companion plants on growth and physiology of Acer saccharinum. Urban Forestry & Urban Greening 74:127652
Simms P (2017) 2013 Colloquium of the Canadian Geotechnical Society: geotechnical and geoenvironmental behaviour of high-density tailings. Can Geotech J 54:455–468
Solla-Gullón F, Santalla M, Rodríguez-Soalleiro RJ, Merino A (2006) Nutritional status and growth of a young Pseudotsuga menziesii plantation in a temperate region after application of wood-bark ash. For Ecol Manage 237:312–321
Solla-Gullón F, Santalla M, Pérez-Cruzado C, Merino A (2008) Response of Pinus radiata seedlings to application of mixed wood-bark ash at planting in a temperate region: nutrition and growth. For Ecol Manage 255:3873–3884
Stupak I, Asikainen A, Röser D, Pasanen K (2016) Review of recommendations for forest energy harvesting and wood ash recycling. In: Röser D, Asikainen A, Raulund-Rasmussen K, Stupak I (eds) Sustainable use of forest biomass for energy, Managing forest ecosystems, vol. 12. Springer, Dordrecht, pp 155–196. https://doi.org/10.1007/978-1-4020-5054-1_7
Thiffault N, Hébert F (2017) Mechanical site preparation and nurse plant facilitation for the restoration of subarctic forest ecosystems. Can J for Res 47:926–934. https://doi.org/10.1139/cjfr-2016-0448
Thomas SC, Gale N (2015) Biochar and forest restoration: a review and meta-analysis of tree growth responses. New for 46:931–946
Thomas SC, Halim MA, Sujeeun GNV, L, (2019) Biochar enhancement of facilitation effects in agroforestry: early growth and physiological responses in a maize-leucaena model system. Agrofor Syst 93:2213–2225
Tordoff GM, Baker JM, Willis AJ (2000) Current approaches to the revegetation and reclamation of metalliferous mine wastes. Chemosphere 41:219–228
Trippe KM, Manning VA, Reardon CL, Klein AM, Weidman C, Ducey TF, Novak JM, Watts DW, Rushmiller H, Spokas KA, Ippolito JA, Johnson MG (2021) Phytostabilization of acidic mine tailings with biochar, biosolids, lime, and locally-sourced microbial inoculum: do amendment mixtures influence plant growth, tailing chemistry, and microbial composition? Appl Soil Ecol 165:103962
Unger YL, Fernandez IJ (1990) The short-term effects of wood-ash amendment on forest soils. Water Air Soil Pollut 49:299–314
van de Voorde TFJ, Bezemer TM, Van Groenigen JW, Jeffery S, Mommer L (2014) Soil biochar amendment in a nature restoration area: effects on plant productivity and community composition. Ecol Appl 24:1167–1177. https://doi.org/10.1890/13-0578.1
Wang S, Mulligan CN (2006) Occurrence of arsenic contamination in Canada: sources, behavior and distribution. Sci Total Environ 366:701–721
Wei Z, Yin G, Wan L, Li G (2016) A case study on a geotechnical investigation of drainage methods for heightening a tailings dam. Environ Earth Sci 75:106
Williams JM, Thomas SC (2023a) Effects of high-carbon wood ash biochar on volunteer vegetation establishment and community composition on metal mine tailings. Restor Ecol 31:e13861
Williams JM, Thomas SC (2023b) High-carbon wood ash biochar for mine tailings restoration: a field assessment of planted tree performance and metals uptake. Sci Total Environ 901:165861
Winter F, Wartha C, Hofbauer H (1999) NO and N2O formation during the combustion of wood, straw, malt waste and peat. Biores Technol 70:39–49
Xu S, Eisenhauer N, Ferlian O, Zhang J, Zhou G, Lu X, Liu C, Zhang D (2020) Species richness promotes ecosystem carbon storage: evidence from biodiversity-ecosystem functioning experiments. Proc R Soc B 287:20202063
Ye ZH, Shu WS, Zhang ZQ, Lan CY, Wong MH (2002) Evaluation of major constraints to revegetation of lead/zinc mine tailings using bioassay techniques. Chemosphere 47:1103–1111
Zhang W, Xu F, Zwiazek JJ (2015) Responses of jack pine (Pinus banksiana) seedlings to root zone pH and calcium. Environ Exp Bot 111:32–34
Acknowledgements
The authors thank D Achircano, P Pajunen, S Matson, and M Deans from the Newmont Musselwhite Mine for field and administrative assistance in support of the study. The authors would also like to thank F Williams for providing guidance throughout writing and data analysis processes.
Funding
This project was funded by grants from the Natural Sciences and Engineering Research Council of Canada. Wood ash used for this field experiment was generously donated by Wood Ash Industries Inc. and Brian Coghlan.
Author information
Authors and Affiliations
Contributions
Both authors conceived, designed, and installed the field experiment, analyzed the data, and edited the manuscript. JM Williams wrote and formatted the manuscript, as well as conducted laboratory analyses. Both authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethical approval
This manuscript has not been submitted to more than one journal for simultaneous consideration.
Consent to participate
All authors agree with the content of the submission, and all agree to continue to support the follow-up work.
Consent for publication
This manuscript has not been submitted or published in other journals, and the authors agree to consent to publish.
Competing interests
The authors declare no competing interests.
Additional information
Responsible Editor: Zhihong Xu
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Highlights
• Tree survival was highest on tailings at 6.4–12.8 t/ha wood ash biochar rates.
• Substrate supply of important plant nutrients was higher in amended plots.
• Wood ash did not result in significant increases in substrate or tree tissue contaminants.
• Early volunteer vegetation cover increased with wood ash, introducing resource competition for trees.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Williams, J.M., Thomas, S.C. High-carbon wood ash biochar enhances native tree survival and growth on sand-capped mine tailings. Environ Sci Pollut Res 31, 43874–43895 (2024). https://doi.org/10.1007/s11356-024-33962-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-024-33962-y