
Abstract
The aim of this study was to evaluate and compare the effects on biochemical parameters and organosomatic indices in the freshwater bivalve Diplodon chilensis exposed to a glyphosate-based formulation under direct and dietary exposures (4 mg a.p./L). After 1, 7, and 14 days of exposure, reduced glutathione (GSH) and thiobarbituric acid reactive substances (TBARS) levels and the activities of glutathione-S- transferase (GST), superoxide dismutase (SOD), and catalase (CAT) were evaluated in the gills and digestive gland. The hepatosomatic (HSI) and branchiosomatic (BSI) indices were also analyzed. Direct and dietary glyphosate-based formulation exposure altered the redox homeostasis in the gills and digestive gland throughout the experimental time, inducing the detoxification response (GST), the antioxidant defenses (SOD, CAT, GSH), and causing lipid peroxidation. After 14 days of exposure, the HSI and BSI increased significantly (43% and 157%, respectively) only in the bivalves under direct exposure. Greater changes in the biochemical parameters were induced by the dietary exposure than by the direct exposure. Furthermore, the gills presented an earlier response compared to the digestive gland. These results suggested that direct and dietary exposure to a glyphosate-based formulation induced oxidative stress in the gills and digestive glands of D. chilensis. Thus, the presence of glyphosate-based formulations in aquatic ecosystems could represent a risk for filter-feeding organisms like bivalves.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Nowadays, agricultural production is closely linked to the use of agrochemicals. In Argentina, glyphosate is the most widely used herbicide, applied in the fallow period for weed control in glyphosate-resistance transgenic crops such as soybean, cotton, and corn (Vara 2004). Different commercial formulations of glyphosate are used in the field; in particular, the Glifosato Atanor® mixture with the addition of the surfactant Impacto® is widely used in soybean fields in the Pampas region of Argentina (Romero et al. 2011; Iummato et al. 2019).
Glyphosate is a systemic, broad-spectrum, post-emergent herbicide. Its primary mechanism of action is the inhibition of the enzyme 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS), disrupting aromatic amino acid biosynthesis, which leads to decreased protein synthesis and, eventually, the death of plants (Duke 1988). EPSPS is present in bacteria, plants, and some fungi and algae, but not in animals (Richards et al. 2006), so glyphosate is generally considered harmless for animals (Mensah et al. 2015). However, adverse health effects have been documented in animals exposed to different glyphosate-based formulations and oxidative stress has been proposed as a possible mechanism of action in several aquatic animals such as bivalves (Abdel-Nabi et al. 2007, Dos Santos and Martínez 2014), fish (Nwani et al. 2013; Li et al. 2017; Ma et al. 2019), and tadpoles (Costa et al. 2008; Riaño et al. 2020).
Oxidative stress occurs when an imbalance is triggered between prooxidant compounds (e.g., reactive oxygen species, ROS) and antioxidant defenses such as reduced glutathione (GSH) and the enzymes superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx). In an oxidative stress condition, the prooxidant species cause oxidative damage to lipids and other macromolecules, which finally damages the cells (Livingstone 1993; Ríos de Molina 2003) and soft tissues of the exposed organisms (Valavanidis et al. 2006). Since there is evidence that several glyphosate-based formulations are able to induce oxidative stress in aquatic organisms, different parameters related to oxidative damage and antioxidant responses are used as biomarkers (Torres et al. 2008; Gomes et al. 2017). In addition, another biomarker that can be analyzed in bivalves and other aquatic animals exposed to contaminants is the detoxifying enzyme glutathione-S-transferase (GST) (Chahouri et al. 2023). Finally, organosomatic indices are used as indicators of the physiological status of bivalves and other organisms. These indices can be altered under stress conditions (Cartier et al. 2004; Touahri et al. 2016).
Although glyphosate is mostly applied to crops, it can reach water bodies in different ways (runoff, drainage, air drift) and has a half-life in water of 7 to 142 days (Annett et al. 2014). Thus, glyphosate has been detected in many surface water systems worldwide. In Argentina, Lutri et al. (2020), Peruzzo et al. (2008), and Avigliano and Schenone (2015) have reported maximum glyphosate levels of 0.16, 0.7, and 1.6 mg/L, respectively, in streams and rivers of the Pampas region and the Misiones province. Additionally, Ronco et al. (2008) reported high concentrations, within a range from 1.8 to 10.9 mg/L of glyphosate, in streams of the Pampas region, and Sasal et al. (2017) found a maximum concentration of 105 mg/L in water bodies near agricultural or forestry production sites in the Entre Ríos province. Other countries also reported high concentrations of glyphosate in water, such as China, Colombia, and Portugal, with maximum concentrations of 15.21, 2.77, and 2.46 mg/L, respectively (Brovini et al. 2021; Fan et al. 2022).
Aquatic organisms are not only directly exposed to contaminants that are present in water bodies, but also are indirectly exposed to contaminants present in their food. In particular, algae are the basis of many aquatic trophic chains and algal cells can bioconcentrate contaminants adsorbed in their cell walls (Okay et al. 2000), thereby transferring them to organisms higher up in the trophic chain (Torres et al. 2008). Filter-feeding organisms, such as bivalves (clams, oysters, and mussels), are likely exposed to contaminants due to their sessile lifestyle and feeding habits, so they can bioaccumulate xenobiotics in their tissues (Okay et al. 2000; Hanana et al. 2012). Thus, bivalves are considered excellent pollution biomonitors (Chahouri et al. 2023). In water bodies, bivalves are exposed to contaminants that are present in the water column, and the gill is the first organ to come into contact with them. Different contaminants can cross the gill tissue by passive diffusion, depending on their molecular weight and physico-chemical characteristics, enter the circulatory system, and then disperse to different organs (Katagi 2010). Bivalves are also exposed to contaminants present in their habitat through food. In this case, contaminants first come into contact with the digestive system and then enter the circulatory system (Katagi 2010). Therefore, these two types of exposure to pollutants, direct and dietary, can trigger different responses in bivalves. The evaluation of the effects of both types of exposure provides more integrated information on the impact of contaminants on bivalves that are subjected to a polluted environment.
Several studies have revealed adverse effects of glyphosate and its formulations on bivalves, such as damage to the gills and digestive gland, increase in somatic indices, and biochemical alterations (e.g., oxidative stress) (Abdel-Nabi et al. 2007; Bringolf et al. 2007; Hanana et al. 2012; Iummato et al. 2013; Mottier et al. 2013; Zomer Sandrini et al. 2013; Dos Santos and Martínez 2014; Iummato et al. 2018; Cuzziol Boccioni et al. 2021).
Diplodon chilensis is a bivalve distributed in the south of Chile and Argentina, both in pristine environments and in regions of intensive use of agrochemicals (such as the Alto Valle region of the Río Negro and Neuquén provinces, Northern Patagonia, Argentina) (Wais 1987; Abrameto et al. 2019). D. chilensis is a benthic organism that lives in the water–sediment interface of lotic and lentic waterbodies. This species is sensitive to toxic compounds and the eutrophication of waterbodies (Rocchetta et al. 2014; Yusseppone et al. 2020). Since the accumulation of nutrients and contaminants has been documented in D. chilensis, it has been proposed as a bioremediation organism (Sabatini et al. 2011; Bianchi et al. 2014).
Bivalves, as filter-feeding organisms that filter large amounts of phytoplankton in aquatic environments, face the impact of different contaminants present either in the water column or in their food. Although glyphosate is known to induce alterations in a freshwater mussel exposed through diet (Iummato et al. 2018), comparative studies on the influence of the route of exposure on the final effects have not been carried out yet. Also, there are no studies on enzymatic or oxidative stress responses associated with dietary exposure to glyphosate in freshwater bivalves. Therefore, the aim of this study was to evaluate and compare the effects of direct and dietary exposures to a glyphosate-based formulation (GBF) on a freshwater bivalve. Alterations of biochemical parameters and organosomatic indices were evaluated in bivalves exposed by direct and dietary routes over 1, 7, and 14 days. For this purpose, the oxidative stress biomarkers SOD, CAT, GSH, and lipid damage, the activity of the detoxifying enzyme GST, and the hepatosomatic and branchiosomatic indices were analyzed.
Materials and methods
Chemicals
The commercial formulation of glyphosate used in this study was Glifosato ATANOR® (48% p/v isopropylamine salt of N-phosphonomethyl glycine, Atanor, Munro, Buenos Aires, Argentina). The surfactant was alkyl aryl polyglycol ether 50% (p/v) IMPACTO® (AGROASIST S.R.L., Argentina).
Organisms
The BAFC CA4 strain of Scenedesmus vacuolatus (Chlorophyceae, Chlorophyta) came from the Culture Collection of the Laboratorio de Biología de Protistas of the Departamento de Biodiversidad y Biología Experimental that belongs to the Centro de Recursos Genéticos of the Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires (Argentina).
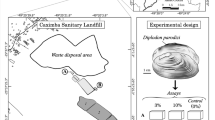
Fifty-four adult specimens of D. chilensis (Bivalvia, Hyriidae) (Gray, 1828) of 6.79 ± 0.39 cm shell length and 29.87 ± 5.16 g were collected by diving at a depth of 7 m in the Paimun Lake (39° 43′ 30.51″ S, 71° 32′ 44.89″ W), Lanín National Park, Neuquén, Argentina. Bivalves were transported to the lab, where they were acclimatized for 3 weeks in tanks with aerated dechlorinated tap water, at 20 ± 2 °C, and a 12:12 h (L:D) photoperiod regime. During the acclimatization period, bivalves were fed with S. vacuolatus twice a week (Sabatini et al. 2011). Specimens collected corresponded to adult animals from 32 to 36 years old (Rocchetta et al. 2014).
Algal cultures
Algal cultures were grown in Bold’s basal medium (BBM, Bischoff and Bold 1963) with Glifosato ATANOR® (4 mg active principle/L) and 2.5% surfactant IMPACTO® (treated cultures), or without any glyphosate or surfactant (control cultures). Cultures were incubated for 4 days (Iummato et al. 2019) at 23 ± 1 °C, under continuous agitation and illumination (80 μmol photons/m2 s). After the incubation time, the cells from the control and treated cultures were harvested, washed, and resuspended in dechlorinated water to obtain a concentrated cell suspension. The cell density used to feed bivalves was determined by cell counting in a Neubauer’s chamber, using a Leica light microscope at 400 × .
The glyphosate concentration in BBM was analytically determined at INQUIMAE–CONICET, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires by HPLC–UV chromatography after a derivatization step with FMOC-Cl (Sancho et al. 1996; Stalikas and Konidari 2001). The equipment used for the analysis was an HPLC–UV system (Jasco Analytical Instruments, Easton MD, USA) with a Microsorb C18 5 µm column, L × I.D. 250 × 4.6 mm inner diameter (Varian). The glyphosate concentration measured was 93.3 ± 2.3% of the nominal value. The exposure concentration used in this study was within the range of glyphosate levels found in some water bodies in Argentina (Ronco et al. 2008; Avigliano and Schenone 2015), Canada, and the USA (World Health Organization 2005; Canadian Council of Ministers of the Environment 2012).
Filtration rate
A filtration rate experiment was carried out to evaluate the maximum cell concentration at which D. chilensis could filter control or treat S. vacuolatus cells at a similar rate (Sabatini et al. 2011). Bivalves were placed individually in beakers with 400 mL of dechlorinated tap water with constant aeration, at 20 ± 1 °C (3 replicates per treatment, 9 containers in total). After 48 h of acclimatization without food, when the bivalves had the valves open, control S. vacuolatus cells at a final concentration of 2 × 105 cells/mL were added to beakers 1 to 6, while the same density of treated S. vacuolatus cells was added to beakers 7 to 9. In addition, a concentrated GBF solution (Glifosato ATANOR® with 2.5% surfactant IMPACTO) was added to beakers 4 to 6, in sufficient quantity to reach a concentration of 4 mg active principle (a.p.)/L. Samples were taken at 0, 1, 2, 3, 4, and 24 h after the start of the experiment, and cell density was determined by direct counting. The filtration rate for each bivalve was expressed as L/h per dry soft tissue mass (g) and was calculated according to Jorgensen (1990). Dry soft tissue mass was measured after drying the soft tissues for 48 h at 60 °C until constant mass.
Experimental design
After the acclimatization period, 3 groups of 15 animals each were weighed and placed in 1-L glass containers with 400 mL of aerated dechlorinated tap water (1 individual per container). Control bivalves were fed with control S. vacuolatus cells (cultured without GBF). Direct glyphosate-based formulation exposure bivalves (dirGEB) were fed with control S. vacuolatus cells, and GBF was added to the tap water for a final concentration of 4 mg a.p/L. Dietary glyphosate-based formulation exposure bivalves (dietGEB) received treated S. vacuolatus cells (Iummato et al. 2018). Each group of bivalves was fed with 1.8 × 105 algal cells/mL for 24 h, twice a week, for 2 weeks. Water was replaced completely each time before adding the algae. For the dirGEB group, water renewal included 4 mg a.p./L GBF. Five individuals of each group (control bivalves, dirGEB, and dietGEB) were sacrificed at the end of each experimental period (1, 7, and 14 days), and their body mass, shell length, height, and width were recorded. Soft bodies were weighed, and the gills and digestive glands were removed and stored at − 80 °C until biochemical analysis. The branchiosomatic (BSI) and hepatosomatic (HSI) indices, defined as the organ wet mass/total mass of the organism, were calculated.
The experiment was performed following the “National Research Council’s Guide for the Care and Use of Laboratory Animals” and the “ARRIVE guidelines.”
Sample preparation
Gills and digestive glands were homogenized with 0.154 M KCl (1:5 w/v) containing protease inhibitors (0.2 mM benzamidine and 0.5 mM phenylmethylsulfonyl fluoride). Homogenates were centrifuged at 11,000 × g for 30 min, and supernatants were used for the determination of enzymatic activities, GSH content, and lipid peroxidation levels. All procedures were carried out at 4 °C. Total soluble protein content was determined according to Bradford (1976), using bovine serum albumin as standard.
Lipid peroxidation
Determination of lipid peroxidation levels was carried out by the thiobarbituric acid reactive substances (TBARS) technique (Buege and Aust 1978). TBARS content was estimated as malondialdehyde (MDA) equivalents, using the extinction coefficient of the MDA–thiobarbituric acid complex (156/mM cm). The results were calculated as nmol TBARS per mg protein and were expressed as % with respect to the control bivalves.
Additional details regarding the procedure for determining TBARS content can be found in the Supplementary Material.
Reduced glutathione content (GSH)
GSH content was estimated following the Anderson (1985) method. Briefly, an aliquot of the supernatant was deproteinized with 5% sulfosalicylic acid, centrifuged, and used as a source of endogenous GSH. The concentration of GSH was assessed through its reaction with 5,5′-Dithiobis (2-nitrobenzoic acid) (DTNB), measured by the absorbance at 412 nm. The results were calculated as nmol GSH per mg protein and were expressed as % with respect to the control bivalves.
Additional details regarding the procedure for determining GSH content can be found in the Supplementary Material.
Enzymatic activities
Superoxide dismutase (SOD, EC 1.15.1.1) activity was recorded following the Beauchamp and Fridovich (1971) method. This procedure is based on the inhibition of the photochemical reduction of nitro blue tretrazolium (NBT). One SOD unit represented the amount of enzyme necessary to inhibit the NBT reduction rate by 50%. The results were calculated as units of SOD per mg protein and were expressed as % with respect to the control bivalves.
Catalase (CAT, EC1.11.1.6) activity was measured by monitoring the decomposition of hydrogen peroxide spectrophotometrically at 240 nm (Aebi 1984), using an extinction coefficient of 40/M cm. One CAT unit was defined as the enzyme necessary to decompose 1 mmol of H2O2 per minute. The results were calculated as CAT units per mg protein and were expressed as % with respect to the control bivalves.
Glutathione-S-transferase (GST, EC1.11.1.9) activity was recorded following the Habig et al. (1974) method, making use of 1-chloro-2,4-dinitrobenzene (CDNB) as substrate. One unit of GST was defined as the enzyme quantity required to catalyzing the formation of 1 μmol of GS-DNB per minute. The results were calculated as units of GST per mg protein and were expressed as % with respect to the control bivalves.
Additional details on the procedures for determining enzyme activities are provided in the Supplementary Material.
Statistical analysis
The effects of glyphosate-based formulation and exposure time on the biochemical parameters and the hepatosomatic and branchiosomatic indices were tested by a two-way analysis of variance (ANOVA) followed by a Tukey post hoc test. Normality and homogeneity of variances were tested by Lilliefors and Bartlett’s tests, respectively (Sokal and Rohlf 1999). GraphPad Prism 6 and Statistica 8 software were used for statistical analyses. The correlation matrix was obtained by the Pearson correlation method, using GraphPad Prism 6. Principal component analysis (PCA) was performed using R Project software, with TBARS, SOD, CAT, GSH, GST, HSI, and BSI studied in the gills and digestive gland as active variables. The treatment (control, dietary exposure, and direct exposure) was employed as a supplementary qualitative variable. The ellipsoids of the PCA’s graphics were defined by 95% confidence.
Results
There was no mortality in any of the groups of bivalves throughout the experimental period. The number of male and female bivalves in each experimental group was similar.
Filtration rate
No statistically significant differences in the filtration rate between the groups (p > 0.05) were recorded. The filtration rate of D. chilensis was 0.90 ± 0.18 L/h g for bivalves fed with S. vacuolatus control cells without GBF in water, 0.90 ± 0.30 L/h g for bivalves fed with S. vacuolatus control cells with GBF in water, and 0.89 ± 0.30 L/h g for bivalves fed with GBF treated S. vacuolatus cells.
Hepatosomatic and branchiosomatic indices
The BSI of the dirGEB showed a significant increase compared to the controls and dietGEB (157%, p < 0.001) after 14 days of exposure (Table 1). The HSI of the dirGEB showed a significant increase (43%, p < 0.05) compared to the control bivalves after 14 days of exposure. No significant differences were observed either in shell length or total weight between all the bivalves analyzed (Table 1).
Biochemical parameters
Gills
After 1 and 7 days of exposure, the content of TBARS of the dietGEB gills was significantly higher compared to that of the controls (p < 0.05) and dirGEB (p < 0.01). After 14 days of exposure, the dirGEB gills evidenced a significantly higher TBARS content than the control (p < 0.05) and dietGEB gills (p < 0.05) (Fig. 1A). The SOD activity of the dirGEB gills did not show significant differences from that of the controls at any of the exposure times, while SOD activity of the dietGEB gills showed a significant increase compared to the controls and dirGEB after 1 (p < 0.05 and p < 0.05, respectively) and 14 days (p < 0.001 and p < 0.05, respectively) of exposure (Fig. 1B). Gills CAT activity of the dietGEB and the dirGEB only showed a significant increase compared to the controls (p < 0.001 and p < 0.05, respectively) after 1 day of exposure (Fig. 1C). GST activity of the dietGEB gills showed a significant increase compared to the controls (p < 0.001) and dirGEB (p < 0.01) after 7 days of exposure, whereas GST activity in the dirGEB gills significantly increased compared to the controls (p < 0.001) and dietGEB (p < 0.01) after 14 days of exposure (Fig. 1D). The GSH content of the dirGEB gills did not show significant differences from that of the controls at any of the exposure times, while GSH content of the dietGEB gills showed a significant increase compared to the control bivalves (p < 0.001) and dirGEB (p < 0.001) after 7 days of exposure (Fig. 1E).

Lipid peroxidation (TBARS) level, superoxide dismutase (SOD), catalase (CAT) and glutathione-S-transferase (GST) activities, and glutathione reduced (GSH) content in the gills (A–E) and digestive gland (F–J) of D. chilensis exposed to GBF by dietary and direct exposure at different times. Data are expressed as mean (% respect to control) ± SD (n = 5). Different letters indicate significant differences (p < 0.05)
Digestive gland
The TBARS content of the dietGEB digestive gland was significantly elevated compared to that of the controls (p < 0.001) and dirGEB (p < 0.05) after 14 days of exposure (Fig. 1F). The TBARS content in the dirGEB digestive gland did not show significant differences from that of the controls at any of the exposure times. After 7 and 14 days of treatment, dirGEB showed a significant increase in digestive gland SOD activity compared to controls (p < 0.01 and p < 0.001, respectively), while dietGEB showed a significant increase compared to controls (p < 0.001) and dirGEB (p < 0.001) after 14 days of exposure (Fig. 1G). CAT activity of the dirGEB digestive gland did not show significant differences from that of the controls at any exposure time. However, CAT activity of the dietGEB digestive gland showed a significant increase compared to the controls and dirGEB after 7 (p < 0.05 and p < 0.01, respectively) and 14 days (p < 0.05 and p < 0.01, respectively) of exposure (Fig. 1H). The GST activity of the digestive gland of dirGEB showed a significant increase compared to the controls only after 7 days of exposure (p < 0.01). This activity of dietGEB showed a significant increase compared to controls after 7 (p < 0.05 and) and 14 days of treatment (p < 0.001) and compared to dirGEB after 14 days of exposure (p < 0.001 and) (Fig. 1I). The GSH content of the digestive gland of dietGEB and dirGEB showed a significant increase compared to the controls (p < 0.01 and p < 0.01, respectively) after 7 days of exposure (Fig. 1J).
Multivariate analyses
Correlation matrix
After 1 day of exposure, in the gills, TBARS correlated positively with SOD (p < 0.05) and CAT (p < 0.001) and correlated negatively with GSH (p < 0.05), while SOD correlated positively with CAT (p < 0.05). In the digestive gland, GSH correlated positively with SOD (p < 0.05) and negatively with CAT (p < 0.05) (Table 2).
After 7 days of exposure, in the gills, CAT correlated positively with BSI (p < 0.01) and GSH correlated positively with GST (p < 0.05). In the digestive gland, GSH correlated positively with SOD (p < 0.001) and GST (p < 0.01), and negatively with HSI (p < 0.05), while SOD correlated positively with GST (p < 0.01) (Table 2).
After 14 days of exposure, in the gills, GST correlated positively with TBARS (p < 0.01) and negatively with GSH (p < 0.05). In the digestive gland, SOD correlated positively with CAT (p < 0.05) and with GST (p < 0.05) (Table 2).
Principal component analysis (PCA)
The first three principal components of the PCA explained 67.44%, 69.51%, and 75.31% of the cumulative variance after 1, 7, and 14 days of exposure, respectively (Table 3).
After 1 day of exposure, the 95% confidence ellipsoids overlapped in the tridimensional plane formed by the components of the PCA, indicating no differences between the treatments and the control groups, considering all the studied variables simultaneously (Fig. 2A).

Principal component analysis —95% confidence ellipsoids corresponding to the control, dietary, and direct exposure after 1 (A), 7 (B), and 14 (C) days of exposure
After 7 days of exposure, the confidence ellipsoids corresponding to direct exposure and control overlapped, indicating no differences between them. The ellipsoid of the dietary exposure is separated in the space, indicating differences between the control and direct exposure, considering all the studied variables simultaneously (Fig. 2B).
After 14 days of exposure, the three confidence ellipsoids were distant among them in the plane formed by the components of the PCA, indicating differences between the treatments and between the treatments and the control (Fig. 2C).
Discussion
In Argentina, the intensive use of glyphosate-based formulations (GBFs) leads to its progressive accumulation in the environment. Thus, in aquatic ecosystems, filter-feeding organisms may come in contact with this herbicide through the surrounding water and/or through the food they eat. In this study, the effects of these two ways of exposure to a GBF were evaluated in the Argentinean native freshwater bivalve D. chilensis. The bivalves were exposed to a GBF in the water column (dirGEB) or through S. vacuolatus microalgae contaminated with GBF as food (dietGEB). In a previous study, we showed that S. vacuolatus displayed biochemical alterations (including oxidative stress responses) and disturbances to cell structure when exposed for 96 h to 4 mg a.p./L of the same GBF used in this study (Iummato et al. 2019). These findings suggest that the GBF had entered the algal cells. Moreover, algae can metabolize different xenobiotics (Okay et al. 2000), so it is possible that S. vacuolatus may have metabolized the components of the glyphosate formulation into more toxic metabolites. Therefore, dietGEB may have received GBF and/or toxic metabolites and oxidative stress products from the contaminated S. vacuolatus cells through the diet.
The results obtained show that both direct and dietary GBF exposure induced biochemical alterations in the gills and digestive gland, and alterations in the organosomatic indices BSI and HSI. All analyzed biomarkers related to detoxification processes and oxidative stress (GST, TBARS, GSH, SOD, and CAT) differed significantly between treated and control bivalves throughout the experimental period and increased with the exposure time. Furthermore, gills and digestive glands can respond differently to contaminants (Limon-Pacheco and Gonsebatt 2009), and in our study, we observed different effects on these two organs depending on the type of exposure (diet or direct exposure).
In the dietGEB, the GBF and/or toxic metabolites present in the microalgal cells would have entered the digestive system first, where it/they would have been released from the microalgae by the digestive processes (Penry 2000) and then distributed to other organs, including the gills (Katagi 2010). The digestive gland performs intracellular digestion of food and has an important amount of different enzymes. Likewise, it is the main organ for xenobiotic detoxification and has highly efficient enzymatic mechanisms against ROS (Canesi et al. 2012; Dos Santos and Martínez 2014). Pesticides that enter bivalves are generally unevenly distributed between the different organs, with a higher concentration found in the digestive gland and gonads and a lower concentration in the gills and mantle (Katagi 2010). Therefore, it is likely that the digestive gland was subjected to a higher amount of toxicants than the gills. In the present study, the gills and digestive glands of the dietGEB showed different biochemical alterations depending on the treatment time. After 1 day of exposure to GBF through the diet, gills displayed higher TBARS levels as well as increased CAT and SOD activities, which were positively correlated. SOD is a metalloprotein enzyme that constitutes the first line of defense in the cellular antioxidant defense system. One of the products of its reaction is hydrogen peroxide (Limon-Pacheco and Gonsebatt 2009), which can be removed by the action of the enzyme CAT (Kohen and Nyska 2002). These two enzymes could be induced by an oxidative stress condition evidenced in this case by the increase in TBARS. In contrast to gills, after 1 day of exposure, no alterations were observed in the digestive gland parameters of the dietGEB. Probably, the concentration of ROS and/or toxic substances after 1 day of exposure did not reach the threshold for inducing lipid damage or antioxidant defenses in the digestive gland of the exposed bivalves. Moreover, at this time the gills of the dietGEB would appear to be more sensitive to GBF and/or algal metabolites and would appear to show an earlier response than the digestive gland. After 7 days of exposure, the digestive gland of dietGEB exhibited increased activities of CAT and GST and elevated GSH content, while the gills of the dietGEB showed increased GST activity and increased TBARS and GSH levels. A positive correlation was found between GST activity and GSH levels in the gills and digestive gland, probably related to the fact that increased GSH production is required for increased GST activity (Gao et al. 2018). GSH is a tripeptide considered the principal homeostatic regulator of the cellular redox state (Rios de Molina 2003) and acts as a cofactor of several antioxidant enzymes (Limon-Pacheco and Gonsebatt 2009). GSH levels are regulated by different enzymes, like GST (Peña-Llopis et al. 2002). GST plays a crucial role in the detoxification of xenobiotics, intervening in the phase II reactions of the biotransformation of xenobiotics by catalyzing their conjugation with GSH. GST can detoxify both exogenous and endogenous compounds (e.g., products of lipid peroxidation) also acting as glutathione peroxidase (Jokanović 2001; Ketterer et al. 1988). Therefore, the induction of GST activity could be related to the presence of xenobiotics in the GBF. The increases in GSH and TBARS levels and GST activity could indicate an oxidative stress condition in the gills of the dietGEB (Valavanidis et al. 2006). After 14 days of exposure, in the gills of the dietGEB, was only observed an increase in the SOD activity. While the digestive gland of the dietGEB exhibited increases in TBARS, SOD, CAT, and GST activity, there was a positive correlation between SOD and CAT and between SOD and GST. The increased antioxidant defenses and the increase in the TBARS could be indicating an imbalance in the cellular redox state, caused by an oxidative stress condition in the exposed bivalves (Valavanidis et al. 2006).
In the dirGEB, the gills would have been the first organ to come into contact with the herbicide dissolution. Gills play a fundamental role in respiration and feeding and are the main organ that interacts directly with any xenobiotics present in the water (Abdel-Nabi et al. 2007, Dos Santos and Martínez 2014). The gills are also involved in directing food to the digestive tract (Canesi et al. 2012). The entry of different low molecular weight compounds through the gills has been recorded (Fiala-Médioni et al. 1986). Therefore, glyphosate and other low molecular weight components of the GBF could enter the bivalves through the branchial epithelium of the gills. In the dirGEB, the gills and digestive gland also showed different biochemical alterations depending on the treatment time. After 1 day of exposure to the GBF, no alterations were observed in the digestive gland parameters of the dirGEB, while the gills showed an increase in CAT activity. After 7 days of exposure, no alterations were observed in the gills parameters of the dirGEB, while the digestive gland showed increases in SOD and GST activities, and in GSH content. Positive correlations were found between SOD and GST with GSH and between SOD and GST. In summary, the digestive gland showed an induction of antioxidant and detoxification responses during this period, without recorded lipid damage. Likely, the concentration of ROS and/or toxic substances after 7 days of exposure was insufficient to cause lipid damage in the digestive gland of the treated bivalves. After 14 days of exposure, the digestive gland of the dirGEB only showed an increase in SOD activity, while the gills of the dirGEB showed increases in TBARS and GST activity. The elevated GST activity was positively correlated with increased TBARS levels, suggesting a potential detoxification of lipid peroxides by GST, as proposed by Ketterer et al. (1988).
In summary, D. chilensis exposed to GBF, both directly and indirectly through the diet, showed an increase in lipid peroxidation levels, GSH concentration, and activities of antioxidant and detoxifying enzymes of the gills and digestive gland. To our knowledge, there are practically no studies that analyze the effect of dietary exposure to GBF in bivalves. Several authors have observed the induction of oxidative stress in aquatic organisms, including freshwater worms, tadpoles, and fish, upon direct exposure to glyphosate formulations (Costa et al. 2008; Contardo-Jara et al. 2009; Nwani et al. 2013; Li et al. 2017; Ma et al. 2019; Riaño et al. 2020). In bivalves, most of the studies have focused on the effects of direct exposure to glyphosate or glyphosate-based formulations through the water. Abdel-Nabi et al. (2007) and Dos Santos and Martínez (2014) have reported an increase in TBARS content and SOD activity in the gills and digestive gland of Ruditapes decussatus and Curbicula fluminea. Furthermore, investigations in our laboratory revealed that direct exposure to glyphosate acid in an outdoor microcosm led to an increase in TBARS levels in the mussel Limnoperna fortunei after 26 days of exposure (Iummato et al. 2013).
The biochemical parameters analyzed in the gills and digestive gland were differentially affected, depending on the type of exposure (dietary or direct exposure). When examining all exposure times and organs, it is apparent that the dietGEB displays a greater number of biochemical alterations in comparison to the dirGEB (as evidenced by the ANOVA). This trend was also detected in the multivariate analyses performed for the different exposure times. In the PCA analysis, no differences were observed between the treatments and the control after 1 day of exposure. However, after 7 days of exposure, the dietGEB had more affected variables than the dirGEB, as shown by the separation of the dietGEB ellipsoid from both the control and the dirGEB ellipsoids. After 14 days of exposure, both treatment ellipsoids (dirGEB and dietGEB) were separated from each other and the control, showing that GEB had a greater effect after a longer exposure time. The differences between the treatments could be caused by bioaccumulation of the GBF and/or toxic metabolites in the S. vacuolatus exposed cells (Okay et al. 2000). This would result in the dietGEB being in contact with different toxics, and/or higher concentrations of toxics, than the dirGEB. It should be noted that glyphosate bioaccumulation has been reported in other organisms exposed to glyphosate and glyphosate formulations, such as fish, oligochaetes, snails, and the clam Ruditapes decussatus, with bioconcentration factors ranging from 1.2 to 42.3 (Contardo-Jara et al. 2009; Druart et al. 2011; Canadian Council of Ministers of the Environment 2012; Hanana et al. 2012). Also, algal cells cultured with GBF could contain toxic metabolites related to oxidative stress, such as lipid peroxidation products (malondialdehyde, 4-hydroxy-2-nonenal), which are potentially harmful to bivalve tissue cells (Kalinina et al. 2014).
The analyzed organosomatic indices only showed alterations after the longest exposure time. After 14 days of exposure, an increase in the BSI of the dirGEB may be the result of inflammatory processes in the tissue (Valavanidis et al. 2006) and/or tissue hyperplasia due to tissue damage (Bianchi et al. 2014). Also, direct exposure to GBF for 14 days caused an increase in the HSI of the dirGEB. An increase in the HSI has been previously observed in frogs exposed to a GBF by Paunescu and Ponepal (2011). These authors proposed that it could be due to increased liver development, which could include an increase in the endoplasmic reticulum of liver cells to increase the production of enzymes for metabolization and detoxification of xenobiotics (Thammachoti et al. 2012). On the other hand, prawns exposed to a glyphosate formulation for 7 and 14 days showed dilatation of the endoplasmic reticulum and the Golgi complex in the hepatopancreas (Silveira Melo et al. 2019). Since the histological alterations leading to changes in organs are complex, changes in organosomatic indices require more time to manifest than changes in biochemical markers. Therefore, in the present study, the effects of GBF exposure were recorded first at the biochemical level (in the gills and digestive gland) and then in the organosomatic indices (after 14 days of exposure).
Conclusions
Bivalves can be exposed to contaminants in their environment directly and also through the food they consume. In this study, we provide evidence that a GBF has toxic effects on D. chilensis when presented both in water and in food. The results show that direct and dietary GBF exposures induced oxidative stress in the gills and the digestive gland in these bivalves and also altered the organosomatic indices. The exposure through the diet appears to have more toxic effects than the direct exposure since more biochemical alterations were recorded in the dietGEB than in the dirGEB. Moreover, we show that the toxic effects of the GBF increase over time.
The results from this study also show that the biochemical parameters TBARS levels and SOD and CAT activities could be considered early markers in D. chilensis, since the alterations at the biochemical level manifested at the shorter time of exposure to the GBF. Similarly, the gills could be considered an early response organ, as they showed biochemical alterations before the digestive gland. Therefore, the analysis of the biochemical parameters of the gills could give an early warning related to the presence of herbicide-based formulations in aquatic environments.
Data availability
The data that support the findings of this study are available from the corresponding author, upon request.
References
Abdel-Nabi IM, El-Shenawy NS, Taha IA, Moawad S (2007) Oxidative stress biomarkers and bioconcentration of Reldan and Roundup by the edible clam Ruditapes decussatus. Acta Zool Sin 53:910–920
Abrameto M, Macchi P, Solimano P (2019) Informe del estado ambiental del Río Negro. Universidad Nacional de Río Negro
Aebi H (1984) Catalase in vitro. MethodEnzymol 105:121–126
Anderson ME (1985) Determination of glutathione and glutathione disulfide in biological samples. Method Enzymol 113:548–553
Annett R, Habibi HR, Hontela A (2014) Impact of glyphosate and glyphosate-based herbicides on the freshwater environment. Review article. J ApplToxicol 34:458–479. https://doi.org/10.1002/jat.2997
Avigliano E, Schenone NF (2015) Human health risk assessment and environmental distribution of trace elements, glyphosate, fecal coliform and total coliform in Atlantic Rainforest mountain rivers (South America). Microchem J 122:149–158. https://doi.org/10.1016/j.microc.2015.05.004
Beauchamp C, Fridovich I (1971) Superoxide dismutase: improved assays and an assay applicable to acrylamide gels. Anal Biochem 44:276–286
Bianchi VG, Castro JM, Rocchetta I, Bieczynski F, Luquet CM (2014) Health status and bioremediation capacity of wild freshwater mussels (Diplodon chilensis) exposed to sewage water pollution in a glacial Patagonian lake. Fish Shellfish Immunol 37:268–277. https://doi.org/10.1016/j.fsi.2014.02.013
Bischoff HW, Bold HC (1963) Phycological studies IV. Some soil algae from Enchanted Rock and related algal species. Univ TexPubl 6318:1–95
Bradford MM (1976) A rapid and sensitive method for the quantitation of micrograms quantities utilizing the principle of protein dye binding. Anal Biochem 72:248–254
Bringolf RB, Cope WG, Mosher S, Barnhart MC, Shea D (2007) Acute and chronic toxicity of glyphosate compounds to glochidia and juveniles of Lampsilis siliquoidea (Unionidae). Environ Toxicol Chem 26:2094–2100. https://doi.org/10.1897/06-519R1.1
Brovini EM, Cardoso SJ, Quadra GR, Vilas-Boas JA, Paranaíba JR, Oliveira Pereira R, Fernandes Mendonça R (2021) Glyphosate concentrations in global freshwaters: are aquatic organisms at risk? Environ Sci Pollut Res 28:60635–60648. https://doi.org/10.1007/s11356-021-14609-8
Buege JA, Aust SD (1978) Microsomal lipid peroxidation. Method Enzymol 52:302–310
Canesi L, Ciacci C, Fabbri R, Marcomini A, Pojana G, Gallo G (2012) Bivalve molluscs as a unique target group for nanoparticle toxicity. Mar Environ Res 76:16–21. https://doi.org/10.1016/j.marenvres.2011.06.005
Cartier S, Pellerin J, Fournier M, Tamigneaux E, Girault L, Lemaire N (2004) Use of an index based on the blue mussel (Mytilus edulis and Mytilus trossulus) digestive gland weight to assess the nutritional quality of mussel farm sites. Aquaculture 241:633–654. https://doi.org/10.1016/j.aquaculture.2004.08.015
Canadian Council of Ministers of the Environment (2012) Canadian water quality guidelines for the protection of aquatic life: Glyphosate. In: Canadian environmental quality guidelines, Canadian council of ministers of the environment, Winnipeg
Chahouri A, Yacoubi B, Moukrim A, Banaoui A (2023) Bivalve molluscs as bioindicators of multiple stressors in the marine environment: recent avances. Cont Shelf Res 264 (105056). https://doi.org/10.1016/j.csr.2023.105056
Contardo-Jara V, Klingelmann E, Wiegand C (2009) Bioaccumulation of glyphosate and its formulation Roundup Ultra in Lumbricus variegatus and its effects on biotransformation and antioxidant enzymes. Environ Pollut 157:57–63. https://doi.org/10.1016/j.envpol.2008.07.027
Costa MJ, Monteiro DA, Oliveira-Neto AL, Rantin FT, Kalinin AL (2008) Oxidative stress biomarkers and heart function in bullfrog tadpoles exposed to Roundup Originals. Ecotoxicology 17:153–163. https://doi.org/10.1007/s10646-007-0178-5
Cuzziol Boccioni AP, Lajmanovich RC, Peltzer PM, Attademo AM, Martinuzzi CS (2021) Toxicity assessment at different experimental scenarios with glyphosate, chlorpyrifos and antibiotics in Rhinella arenarum (Anura: Bufonidae) tadpoles. Chemosphere 273:1–12. https://doi.org/10.1016/j.chemosphere.2020.128475
de Silveira Melo M, dos Gonçalves Santos TP, Jaramillo M, Nezzi L, Rauh Muller YM, Nazari EM (2019) Histopathological and ultrastructural indices for the assessment of glyphosate-based herbicide cytotoxicity in decapod crustacean hepatopancreas. AquatToxicol 210:207–214. https://doi.org/10.1016/j.aquatox.2019.03.007
Dos Santos CK, Martinez CBR (2014) Genotoxic and biochemical effects of atrazine and Roundup, alone and in combination, on the Asian clam Corbicula fluminea. Ecotoxicol Environ Saf 100:7–14. https://doi.org/10.1016/j.ecoenv.2013.11.014
Druart C, Millet M, Scheifler R, Delhomme O, Raeppel C, Vaufleury A (2011) Snails as indicators of pesticide drift, deposit, transfer and effects in the vineyard. Sci Total Environ 409:4280–4288. https://doi.org/10.1016/j.scitotenv.2011.07.006
Duke SO (1988) Glyphosate. In: Kearney PC, Kaufman DD (eds) Herbicides: chemistry, degradation and mode of action. Marcel Dekker, New York, pp 1–58
Fan Y, Zhang K, Zhang R, Guo G, Li H, Bai Y, Lin Y, Cai T (2022) Derivation of water quality criteria for glyphosate and its formulations to protect aquatic life in China. Environ Sci Pollut Res 29:51860–51870. https://doi.org/10.1007/s11356-022-19517-z
Fiala-Médioni A, Alayse AM, Cahet G (1986) Evidence of in situ uptake and incorporation of bicarbonate andamino acids by a hydrothermal vent mussel. J Exp Mar Biol Ecol 96:191–198. https://doi.org/10.1016/0022-0981(86)90242-X
Gao W, Pu L, Chen M, Wei J, Xin Z, Wang Y, Yao Z, Shi T, Guo C (2018) Glutathione homeostasis is significantly altered by quercetin via the Keap1/Nrf2 and MAPK signaling pathways in rats. J Clin Biochem Nutr 62(1):56–62. https://doi.org/10.3164/2Fjcbn.17-40
Gomes MP, Gingras Le Manac’h S, Hénault-Ethier L, Labrecque M, Lucotte M, Juneau P (2017) Glyphosate-dependent inhibition of photosynthesis in willow. Front Plant Sci 8:1–13. https://doi.org/10.3389/fpls.2017.00207
Habig W, Pabst M, Jakoby W (1974) Glutathione S-transferases: the first enzymatic step in mercapturic acid formation. J Biol Chem 249:7130–7139
Hanana H, Simon G, Kervarec N, Mohammadou BA, Cérantola S (2012) HRMAS NMR as a tool to studymetabolic responses in heart clam Ruditapes decussatus exposed to Roundup®. Talanta 97:425–431. https://doi.org/10.1016/j.talanta.2012.04.057
Iummato MM, Di Fiori E, Sabatini SE, Cacciatore LC, Cochón AC, Ríos de Molina MC, Juárez AB (2013) Evaluation of biochemical markers in the golden mussel Limnoperna fortunei exposed to glyphosate acid in outdoor microcosms. EcotoxicolEnvironSaf 95:123–129. https://doi.org/10.1016/j.ecoenv.2013.05.024
Iummato MM, Sabatini SE, Cacciatore LC, Cochón AC, Cataldo D, Ríos de Molina MC, Juárez AB (2018) Biochemical responses of the golden mussel Limnoperna fortunei under dietary glyphosate exposure. Ecotoxicol Environ Saf 163:69–75. https://doi.org/10.1016/j.ecoenv.2018.07.046
Iummato MM, Fassiano AB, Graziano M, Dos Santos M, Ríos de Molina MC, Juárez AB (2019) Effect of glyphosate on the growth, morphology, ultrastructure and metabolism of Scenedesmus vacuolatus. Ecotoxicol Environ Saf 172:471–479. https://doi.org/10.1016/j.ecoenv.2019.01.083
Jokanović M (2001) Biotransformation of organophosphorus compounds. Toxicology 166:139–160. https://doi.org/10.1016/s0300-483x(01)00463-2
Jorgensen CB (1990) Bivalve filter feeding: hydrodynamics, bioenergetics, physiology and ecology. Olsen and Olsen, Fredensborg
Kalinina EV, Chernov NN, Novichkova MD (2014) Role of glutathione, glutathione transferase, and glutaredoxin in regulation of redox-dependent processes. Biochem Mosc 79:1562–1583. https://doi.org/10.1134/s0006297914130082
Katagi T (2010) Bioconcentration, bioaccumulation, and metabolism of pesticides in aquatic organisms. Rev Environ Contam Toxicol 204:1–132. https://doi.org/10.1007/978-1-4419-1440-8_1
Ketterer B, Meyer DJ, Tan KH (1988) The role of glutathione transferase in the detoxication and repair of lipid and DNA hydroperoxides. In: Oxygen radicals in biology and medicine. Basic Life Sciences 49. Springer, Boston, MA. https://doi.org/10.1007/978-1-4684-5568-7_105
Kohen R, Nyska A (2002) Oxidation of biological systems: oxidative stress phenomena, antioxidants, redox reactions, and methods for their quantification. ToxicolPathol 30(6):620–650. https://doi.org/10.1080/01926230290166724
Li M-H, Ruan L-Y, Zhou J-W, Fu Y-H, Jiang L, Zhao H, Wang J-S (2017) Metabolic profiling of goldfish (Carassius auratis) after long-term glyphosate based herbicide exposure. AquatToxicol 188:159–169. https://doi.org/10.1016/j.aquatox.2017.05.004
Limon-Pacheco J, Gonsebatt ME (2009) The role of antioxidants and antioxidant-related enzymes in protective responses to environmentally induced oxidative stress: minireview. Mutat Res 674:137–147. https://doi.org/10.1016/j.mrgentox.2008.09.015
Livingstone DR (1993) Biotechnology and pollution monitoring: use of molecular biomarkers in the aquatic environment. J Chem Technol Biotechnol 57:195–211. https://doi.org/10.1002/jctb.280570302
Lutri VF, Matteoda E, Blarasin M, Aparicio V, Giacobone D, Maldonado L, Becher Quinodoz F, Cabrera A, Giuliano Albo J (2020) Hydrogeological features affecting spatial distribution of glyphosate and AMPA in groundwater and surface water in an agroecosystem. Córdoba, Argentina. Sci Total Environ 711:1–12. https://doi.org/10.1016/j.scitotenv.2019.134557
Ma J, Zhu J, Wang W, Ruan P, Rajeshkumar S, Li X (2019) Biochemical and molecular impacts of glyphosate-based herbicide on the gills of common carp. Environ Pollut 252:1288–1300. https://doi.org/10.1016/j.envpol.2019.06.040
Mensah PK, Palmer CG, Odume ON (2015) Ecotoxicology of glyphosate and glyphosate- based Herbicides-toxicity to wildlife and humans. In: Larramendy ML, Soloneski S (eds.), Toxicity and Hazard of Agrochemicals. InTech Publisher, 93–112. https://doi.org/10.5772/60767
Mottier A, Kientz-Bouchart V, Serpentini A, Lebel JM, Jha AN, Costil K (2013) Effects of glyphosate-based herbicides on embryo-larval development and metamorphosis in the Pacific oyster, Crassostrea gigas. AquatToxicol 128–129:667–678. https://doi.org/10.1016/j.aquatox.2012.12.002
Nwani CD, Nagpure NS, Kumar R, Kushwaha B, Lakra WS (2013) DNA damage and oxidative stress modulatory effects of glyphosate-based herbicide in freshwater fish, Channa punctatus. Environ Toxicol Phar 36:539–547. https://doi.org/10.1016/j.etap.2013.06.001
Okay OS, Donkin P, Peters LD, Livingstone DR (2000) The role of algae (Isochrysis galbana) enrichment on the bioaccumulation of benzo[a]pyrene and its effects on the blue mussel Mytilus edulis. Environ Pollut 110:103–113. https://doi.org/10.1016/s0269-7491(99)00282-1
Paunescu A, Ponepal CM (2011) Effect of Roundup® herbicide on physiological indices in marsh frog Pelophylax ridibundus. In: University of agronomic sciences and veterinary medicine (ed) Scientific papers series A, vol 54, Bucharest, pp 269–274
Peña-Llopis S, Ferrando MD, Peña JB (2002) Impaired glutathione redox status is associated with decreased survival in two organophosphate-poisoned marine bivalves. Chemosphere 47:485–497. https://doi.org/10.1016/s0045-6535(01)00323-x
Penry DL (2000) Digestive kinematics of suspension-feeding bivalves: modeling and measuring particle processing in the gut of Potamocorbula amurensis. Mar Ecol Prog Ser 197:181–192
Peruzzo PJ, Porta AA, Ronco AE (2008) Levels of glyphosate in surface waters, sediments and soils associated with direct sowing soybean cultivation in North pampasic region of Argentina. Environ Pollut 156:61–66. https://doi.org/10.1016/j.envpol.2008.01.015
Riaño C, Ortiz-Ruiz M, Pinto Sanchez NR, Gomez E (2020) Effect of glyphosate (Roundup Active®) on liver of tadpoles of the colombian endemic frog Dendropsophus molitor (Amphibia: Anura). Chemosphere 250:1–7. https://doi.org/10.1016/j.chemosphere.2020.126287
Richards TA, Dacks JB, Campbell SA, Blanchard JL, Foster PG, McLeod R, Roberts CW (2006) Evolutionary origins of the eukaryotic shikimate pathway: gene fusions, horizontal gene transfer, and endosymbiotic replacements. Eukaryot Cell 5(9):1517–1531. https://doi.org/10.1128/ec.00106-06
Ríos de Molina MC (2003) El estrés oxidativo y el destino celular. Química Viva 2(1):17–28
Rocchetta I, Lomovasky BJ, Yusseppone MS, Sabatini SE, Bieczynski F, Ríos de Molina MC, Luquet CM (2014) Growth, abundance, morphometric and metabolic parameters of three populations of Diplodon chilensis subject to different levels of natural and anthropogenic organic matter input in a glaciar lake of North Patagonia. Limnologica 44:72–80. https://doi.org/10.1016/j.limno.2013.06.004
Romero D, Ríos de Molina MC, Juárez AB (2011) Oxidative stress induced by a commercial glyphosate formulation in a tolerant strain of Chlorella kessleri. Ecotoxicol Environ Saf 74:741–747. https://doi.org/10.1016/j.ecoenv.2010.10.034
Ronco A, Carriquiriborde P, Natale GS, Martin M, Mugni H, Bonetto C (2008) Integrated approach for the assessment of biotech soybean pesticides impact on low order stream ecosystems of the Pampasic Region. Ecosystem ecology research trends. Nova Science Publisher Inc, New York, USA, pp 209–239
Sabatini SE, Rocchetta I, Nahabedian DE, Luquet CM, Eppis MR, Bianchi L, Ríos de Molina MC (2011) Oxidative stress and histological alterations produced by dietary copper in the freshwater bivalve Diplodon chilensis. Comp Biochem Physiol Part C 154:391–398. https://doi.org/10.1016/j.cbpc.2011.07.009
Sancho JV, Hernandez F, Lopez FJ, Hogendoorn EA, Dijkman E (1996) Rapid determination of glufosinate, glyphosate and aminomethylphosphonic acid in environmental water samples using precolumn fluorogenic labeling and coupled column liquid chromatography. J Chromatogr A 737:75–83. https://doi.org/10.1016/0021-9673(96)00071-4
Sasal MC, Wilson MG, Sione SM, Beghetto SM, Gabioud EA, Oszust JD, Paravani EV, Demonte L, Repetti MR, Bedendo DJ, Medero SL, Goette JJ, Pautasso N, Schulz GA (2017) Monitoreo de glifosato en agua superficial en Entre Ríos. La investigación acción participativa como metodología de abordaje. Revista de Investigaciones Agropecuarias 43(2):195–205
Sokal RR, Rohlf FJ (1999) Introducción a la Bioestadística. Reverté, Barcelona, España
Stalikas CD, Konidari CN (2001) Analytical methods to determine phosphonic and amino acid group containing pesticides. J Chromatogr A 907:1–19. https://doi.org/10.1016/s0021-9673(00)01009-8
Thammachoti P, Khonsue W, Kitana J, Varanusupakul P, Kitana N (2012) Morphometric and gravimetric parameters of the rice frog Fejervarya limnocharis living in areas with different agricultural activity. J Environ Prot 3:1403–1408. https://doi.org/10.4236/jep.2012.310159
Torres MA, Barros MP, Campos SCG, Pinto E, Rajamani S, Sayre RT, Colepicolo P (2008) Biochemical biomarkers in algae and marine pollution: a review. Ecotoxicol Environ Saf 71:1–15. https://doi.org/10.1016/j.ecoenv.2008.05.009
Touahri HG, Boutiba Z, Benguedda W, Shaposhnikov S (2016) Active biomonitoring of mussels Mytilus galloprovincialis with integrated use of micronucleus assay and physiological indices to assess harbor pollution. Mar Pollut Bull 110:52–64. https://doi.org/10.1016/j.marpolbul.2016.06.029
Valavanidis A, Vlahogianni T, Dassenakis M, Scoullos M (2006) Molecular biomarkers of oxidative stress in aquatic organisms in relation to toxic environmental pollutants. Ecotoxicol Environ Saf 64:178–189. https://doi.org/10.1016/j.ecoenv.2005.03.013
Vara AM (2004) Transgénicos en Argentina: más allá del boom de la soja. Revista Iberoamericana De Ciencia, Tecnología y Sociedad 1(3):101–129
Wais IR (1987) Macrozoobenthos of Negro River Basin, Argentine Patagonia. Stud Neotrop Fauna Environ 22(2):73–91. https://doi.org/10.1080/01650528709360721
World Health Organization (2005) Glyphosate and AMPA in drinking-water. WHO/WSH/03.04/97. Geneva, Switzerland
Yusseppone MS, Bianchi VA, Castro JM, Noya AT, Minaberry YS, Sabatini SE, Luquet CM, Ríos de Molina MC, Rocchetta I (2020) In situ experiment to evaluate biochemical responses in the freshwater mussel Diplodon chilensis under anthropogenic eutrophication conditions. Ecotoxicol Environ Saf 193:1–11. https://doi.org/10.1016/j.ecoenv.2020.110341
Zomer Sandrini J, Coimbra Rola R, Moreira Lopes F, Buffon HF, Marques Freitas M, Martinez C, Martins G, Rosa CE (2013) Effects of glyphosate on cholinesterase activity of the mussel Perna perna and the fish Danio rerio and Jenynsia multidentata: In vitro Studies. Aquat Toxicol 130–131:171–173. https://doi.org/10.1016/j.aquatox.2013.01.006
Acknowledgements
We thank Florencia Molina for the graphic assistance and Lucila Herbert and Carlos Luquet for the English language revision.
Funding
This work was partly supported by grants from CONICET—Argentina (Consejo Nacional de Investigaciones Científicas y Técnicas, PIP 11220130100020CO) and from Universidad de Buenos Aires—Argentina (UBACyT 20020120200176BA 451, 2020130100248BA, 20020170100359BA).
Author information
Authors and Affiliations
Contributions
All authors contributed to the work, read and approved the final manuscript, and consented to publish it. Iummato MM: conceptualization, investigation, formal analysis, writing—original draft, writing—review and amp; editing. Sabatini SE: conceptualization, investigation; Rocchetta I: conceptualization, investigation; Yusseppone MS: investigation; Ríos de Molina MC: conceptualization, investigation; writing—review and amp; editing, supervision, funding acquisition; Juarez AB: conceptualization, investigation, writing—review and amp; editing, supervision, funding acquisition.
Corresponding author
Ethics declarations
Ethical approval
The experiments of this work involved animals and were carried out following the “National Research Council’s Guide for the Care and Use of Laboratory Animals” and the “ARRIVE guidelines.”
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Conflict of interest
The authors declare no competing interests.
Additional information
Responsible Editor: Bruno Nunes
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Iummato, M.M., Sabatini, S.E., Rocchetta, I. et al. Oxidative stress in the bivalve Diplodon chilensis under direct and dietary glyphosate-based formulation exposure. Environ Sci Pollut Res 31, 23610–23622 (2024). https://doi.org/10.1007/s11356-024-32639-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-024-32639-w