
Abstract
Coeur d’Alene Lake (the Lake) has received significant contamination from legacy mining. Aquatic macrophytes provide important ecosystem services, such as food or habitat, but also have the ability to accumulate contaminants. We examined contaminants (arsenic, cadmium, copper, lead, and zinc) and other analytes (e.g., iron, phosphorus, and total Kjeldahl nitrogen (TKN)) in macrophytes from the Lake. Macrophytes were collected in the Lake from the uncontaminated southern end to the outlet of the Coeur d’Alene River (main contaminant source) located northward and mid lake. Most analytes showed significant north to south trends (Kendall’s tau p ≤ 0.015). Concentrations of cadmium (18.2 ± 12.1), copper (13.0 ± 6.6), lead (195 ± 193), and zinc (1128 ± 523) were highest in macrophytes near the Coeur d’Alene River outlet (mean ± standard deviation in mg/kg dry biomass). Conversely, aluminum, iron, phosphorus, and TKN were highest in macrophytes from the south, potentially related to the Lake’s trophic gradient. Generalized additive modelling confirmed latitudinal trends, but revealed that longitude and depth were also important predictors of analyte concentration (40–95% deviance explained for contaminants). We used sediment and soil screening benchmarks to calculate toxicity quotients. Quotients were used to assess potential toxicity to macrophyte associated biota and delineate where macrophyte concentrations exceeded local background concentrations. Exceedances (toxicity quotient > one) of background levels by macrophyte concentrations were highest for zinc (86%), followed by cadmium (84%), lead (23%), and arsenic (5%).
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Lakes act as natural settling basins for materials carried through their watersheds. Coeur d’Alene Lake, located in northwestern USA, is no exception and has received significant amounts of metal(loid)s contaminated sediment from legacy mining activities originating in the Coeur d’Alene River watershed (Zinsser 2020; Langman et al. 2020). The Bunker Hill mining complex located adjacent to the Coeur d’Alene River was placed on the National Priorities List in 1983 (USEPA 2021) and has been described as a Superfund “megasite” with contamination spanning an approximate 241 km long corridor (Gustavson et al. 2007). Remedial activities began after Superfund listing, however significant contamination still remains within the watershed because of the large extent and complex nature of its distribution (NRC 2005). Contaminated sediments were spread widely by the Coeur d’Alene River into adjacent wetlands, Coeur d’Alene Lake, and downstream into the Spokane River (USEPA 2002, 2012).
Aquatic plants growing within contaminated lakes and wetlands have the capacity to concentrate metal(loid)s into biomass from both sediment and water (Xing et al. 2013). The ability to collect contaminants in biomass has important implications for metals cycling (Jackson 1998) and creates risk of toxic exposure for biota living in association with this vegetation, e.g., herbivore dietary exposure (Désy et al. 2002). Metals sequestration by aquatic vegetation has even been used as a remediation technique to extract contaminants from various media (Xing et al. 2013; Newete and Byrne 2016). Aquatic macrophytes can also serve as bioindicators of metal(loid)s pollution because of their ability to accumulate contaminants (Singh et al. 2016; Ceschin et al. 2021).
In this study, we examined the spatial distribution of mining related contaminants and nutrient concentrations in biomass of submerged aquatic vegetation collected from Southern Coeur d’Alene Lake, hereafter referred to as “the Lake.” We also compared biomass concentrations to ecologically relevant thresholds (benchmarks) to screen for the most contaminated sites. Results from this work have important implications for the management of nonnative vegetation such as Myriophyllum spicatum (Scofield et al. 2021). Myriophyllum spicatum (Eurasian watermilfoil) is actively managed in the Lake to reduce its negative impacts, e.g. reduced lakefront property values and displacement of native vegetation (Liao et al. 2016). Mechanical harvesting is one such management approach, but collection of contaminated biomass can require careful disposal (Newete and Byrne 2016) and delineation of contaminated macrophytes in the Lake has not been completed. Additionally, macrophyte biomass concentrations should help inform the potential of contaminant exposure to biota associated with this vegetation, such as exposure during feeding activities (Sample et al. 2014). Resident and migratory waterfowl visiting the Lake have experienced significant toxicity for decades, which occurs from incidental ingestion of lead contaminated sediment while feeding (Spears et al. 2007). Our study will help determine sites where macrophyte contamination persists and has the potential to enter the food web of the Lake.
Methods
Study area
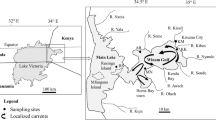
We collected aquatic macrophytes from 94 sites between the outlet of the Coeur d’Alene River and the southern boundary (Chatcolet Lake) of the Lake (Fig. 1). Sampling was confined to waters within the Coeur d’Alene Tribe’s reservation boundary and where M. spicatum has been managed (CDAT and Avista Corp. 2017). At summer pool elevation (649.8 m, NAVD88), the Lake has a surface area and maximum depth of 129 km2 and 64 m, respectively (Wood and Beckwith 2008). The Lake’s watershed covers 9690 km2 and is drained mostly by the Coeur d’Alene and St. Joe rivers, which account for 39% and 47% of the total watershed, respectively (Wood and Beckwith 2008). Most contaminant metals in the Lake originate from the Coeur d’Alene River watershed (Zinsser 2020).

Map of the study area, major rivers, and the major contamination source (Bunker Hill Box)
Vegetation sampling
We selected sites based on a stratified random design, where the study area was divided into nine 1 km wide strata from north to south. Within each stratum, ten random samples were selected from areas known to grow submerged macrophytes. Some littoral areas of the Lake do not support macrophytes, because they are steep or rocky. Vegetated areas were determined from a sonar survey conducted in July 2013 (AquaTechnex 2013) using consumer grade hydroacoustic equipment and the proprietary BioBase algorithm (Torso et al. 2020). Sites were randomly selected from areas with macrophytes occupying ≥ 50% of the water column as determined by sonar. We sampled sites from the southern extent of the sampling area first and progressed northward. This was done to limit cross contamination of metals between sites based upon prior sediment sampling (Horowitz et al. 1993, 1995) where sediments from the southern end of the Lake were less contaminated than near the outlet of the Coeur d’Alene River.
We navigated to sites with a Lowrance Point-1 GPS antennae and collected vegetation with a rake on a 5 m pole. Sampling depths were estimated from 0.25 m increment marks on the rake pole. Rake snagged vegetation was placed into a clean plastic bin and any debris was removed from the sample. A random portion of stems and leaves were selected and thoroughly rinsed with lake water. This sample was then placed into a sealable plastic bag. Samples represent a composite of above sediment vegetation collected at a site. We chose to use above sediment composites, because our primary objective was focused on delineating spatial differences in concentrations of “manageable” macrophyte biomass.
Aquatic vegetation management tools such as mechanical harvesting or contact herbicides (i.e., diquat) only crop the above sediment biomass. Nonnative species like M. spicatum typically grow interspersed with other plants (Hansen et al. 2013; Torso et al. 2020) and analyte concentrations derived from composite samples represent a real-world concentrations of manageable biomass. Additionally, site-specific factors can have a greater influence on biomass metal concentrations relative to varying bioaccumulation ability across species (Jackson 1998). Previous work in the Coeur d’Alene Lake watershed appears to support this idea that species differences (e.g., Elodea canadensis and M. spicatum) in metal concentrations of manageable biomass may be negligible relative to site differences (Scofield et al. 2021).
Bagged samples were immediately placed on ice in a cooler and transported to an accredited analytical laboratory within one day of collection. All samples were collected and handled with clean nitrile gloves, being changed frequently between sites. Chain of custody records were recorded for all samples. Field replicates were collected at four sites to assess variability attributable to sampling and laboratory methodology. Replicate variability per analyte was reported as the relative percent difference (USEPA 2014).
Laboratory analyses
Plant biomass was analyzed by an accredited laboratory for concentrations of aluminum, arsenic, cadmium, copper, iron, total Kjeldahl nitrogen (TKN), lead, manganese, phosphorus, and zinc. All analyte concentrations were determined with inductively coupled plasma-optical emission spectrometry via method 6010D (USEPA 2014) except for TKN. Total Kjeldahl nitrogen was determined by colorimetry using method 351.2 (USEPA 1993). Prior to analysis, sample biomass was dried to a constant mass at 60 °C, homogenized with a stainless steel Wiley mill, and heat digested using method 3050B (USEPA 1996) for all analytes except TKN. The TKN digestion is described in method 351.2 (USEPA 1993). Analyte concentrations along with detection and reporting limits are reported as dry mass (Table 1). All internal laboratory quality assurance and quality control measures were met as per the reported analytical methods. The raw data along with any data qualifiers can be downloaded from the data repository listed in the “Data and code availability” section.
Statistical analyses
Data analyses were preformed using R statistical software v.4.2.0 (R Core Team 2022) and tidyverse packages. All data, scripts, and package citations used in analyses can be found at the repository listed in the “Data and code availability” section. The analytical laboratory qualified some of the reported concentration data, because it was either between the detection and reporting limits, or was below a detection limit. We treated concentrations between the detection and reporting limits the same as results above the reporting limit.
There was only one nondetect from an arsenic result, and we imputed a value using the ROS method from the NADA package in this case (Helsel 2012). We also removed one sample from the analyses based on an unusually high copper concentration (50.5 mg/kg). The sample showed high leverage during modelling, with a Cook’s distance > 0.5. We assumed the reported concentration was erroneous or the sample was contaminated during handling since it originated from the southernmost sampling stratum.
Latitudinal gradients in analyte concentrations were examined graphically and with Kendall’s tau calculated by the NADA package. Latitude versus concentration figures were created with the ggplot2 package and used a generalized additive modelling (GAM) smoother to show the slope and strength (default 95% confidence interval) of the relationship between variables. The latitudinal gradients in analyte concentrations from macrophyte biomass were also assessed in relation to previous sediment mapping (Horowitz et al. 1993). We used the stars package to extract sediment raster values at macrophyte sampling sites. The degree of association between spatially overlapping macrophyte and sediment concentrations were assessed with Spearman’s rho along with plotting in ggplot2.
An aquatic macrophyte anthropogenic disturbance index was used to qualitatively describe the macrophyte community across latitude via sampling strata. The disturbance index was developed as a bioassessment tool and indicates lake trophic state as well as anthropogenic disturbance based upon macrophyte tolerances (Mikulyuk et al. 2017). Macrophyte relative frequencies were calculated as the percent of a given species divided by the sum of species observed over the entire study area or stratum.
We also used GAMs to assess the strength of spatial distribution as a predictor of analyte concentration (Wood 2017; Bergbusch et al. 2021), but without assumptions of relationship shape, e.g., monotonic as needed by Kendall’s tau. Analyte concentrations were modelled individually as functions of two-dimensional smoothers of longitude/latitude and a one dimensional smoother for sample depth with the mgcv package. We assumed a gamma distribution for the concentration data and used the “log” link function (Pedersen et al. 2019). The spatial smoothers used the Duchon spline basis and the depth smoothers used the default thin plate regression spline basis. Smoothing parameters were estimated using the “REML” method. The gam.check function was used to ensure an adequate number of basis functions. The gratia package was used to assess model diagnostics and fit. The GAM results were visualized with the ggplot2, sf, and ggspatial packages. Significant model terms were reported based on the computed F statistic at α = 0.05.
Toxicity quotients
We chose several ecologically relevant thresholds to assess the degree of contamination in macrophyte biomass. We calculated toxicity quotients (Sample et al. 2014) by dividing a biomass concentration by a threshold (benchmark), where quotients > 1 indicate the potential for toxicity. Explicit benchmarks for macrophytes have not been developed extensively (Lewis et al. 2001; Ceschin et al. 2021). However, because macrophytes are consumed by a variety of organisms and decomposed biomass becomes incorporated into sediments, we selected a range of sediment/soil screening values to gauge exposure potential to organisms associated with sediments and macrophytes (MacDonald et al. 2000; Spears et al. 2007; Scofield et al. 2021).
Cleanup actions have been ongoing in the Coeur d’Alene River watershed directed by USEPA through the Superfund program (Gustavson et al. 2007; USEPA 2021). These efforts have resulted in cleanup guidance dependent on the receptor in question, e.g. human health, ecological receptor (USEPA 2002, 2012). From these benchmarks, we selected thresholds spanning a range of sensitivities in invertebrates, birds, and mammals (i.e., ecologic receptors) associated with palustrine, lacustrine, and riverine soil/sediments (Table 2).
Additionally, we wanted to delineate where vegetation becomes contaminated enough to warrant special consideration concerning invasive species (e.g., M. spicatum) management. Mechanical harvesting has been used to remove M. spicatum, from the southernmost end (e.g., Chatcolet Lake) of the Lake. However, harvesting has not been implemented in the main Lake basin where significant amounts of contaminated sediment reside (Horowitz et al. 1993, 1995). Mechanically harvested vegetation is sometimes used in compost (Quilliam et al. 2015; Bartodziej et al. 2017). This study was intended to evaluate where in the Lake macrophytes become too contaminated (Shayler et al. 2009) to be used as compost feedstock and biomass may require special handling and disposal (Newete and Byrne 2016). Benchmarks used to evaluate the suitability of harvested biomass as compost were generally more conservative, being lower than most ecological receptor benchmarks since they are concerned with protecting human health.
Results
Latitudinal gradients
Analyte concentrations from macrophyte biomass ranged widely (e.g., mean cadmium ranged over two orders of magnitude) and some analytes showed distinct latitudinal trends (Table 3). Concentrations of cadmium, copper, lead, and zinc were highest at the northern most sites, being nearest to the contamination source (i.e., Coeur d’Alene River outlet). Kendall’s tau slopes for these contaminants were all positive and significant with p < 0.001 (Fig. 2). Arsenic concentrations were highest at northernmost sites, but the southernmost sites also had higher arsenic concentrations. Aluminum, iron, phosphorus, and TKN all had significant and negative Kendall’s tau slopes, indicating increasing concentrations from north to south, in contrast to cadmium, copper, lead, and zinc. Concentrations of aluminum, arsenic, and TKN as a function of latitude may not be monotonic as indicated by the GAM smoothers (Fig. 2) and Kendall’s tau may not be appropriate for modelling these analytes.

Latitudinal gradient of analyte concentrations in macrophyte biomass. Kendall’s tau slope and associated p values are also reported. The solid lines show a generalized additive modelling smoother by latitude for each analyte with a 95% confidence interval (shaded area). The red horizontal dashed lines show the lower basin 90th percentile sediment-soil background benchmark for contaminants of concern (USEPA 2002)
Cadmium, copper, lead, and zinc concentrations in macrophyte biomass were positively correlated with sediment concentrations (Spearman’s rho = 0.50–0.78, p < 0.001). Aluminum, iron, and manganese concentrations in biomass showed no correlation with sediment concentrations (Spearman’s rho = − 0.13–0.01, p ≥ 0.23). Arsenic in biomass was negatively correlated with sediment concentrations, but the relationship was weak (Spearman’s rho = − 0.23, p = 0.033). The strengths and direction of association between sediment and biomass concentrations were verified with plotting (Online Resource 1). Associations between sediment and biomass concentrations of phosphorus and TKN could not be assessed, because these analytes were not determined in sediments.
Macrophytes collected from the southernmost sites tended to be in shallower water (Table 3). The most common taxa across all sites were Elodea canadensis (23%), Potamogeton pusillus (16%), P. amplifolius (12%), M. spicatum (12%), P. richardsonii (8%), Ceratophyllum demersum (6%), and P. robbinsii (5%). The anthropogenic disturbance index calculated from species composition and tolerances indicated that sensitive species were more frequently found at some northern strata (i.e., clb, chp, and brb). Southerly sites had higher percentages of species that are moderately (“general”) tolerant to eutrophication and shoreline development.
Generalized additive modelling
In addition to the latitudinal gradient, the GAM results indicated that longitudinal position and macrophyte depth were also important in explaining analyte concentrations (Online Resource 2). Modelling showed that concentrations of lead (95%), cadmium (95%), zinc (92%), copper (67%), aluminum (53%), and arsenic (40%) were strongly to moderately predicted by spatial position (deviance explained %), where the longitude/latitude smoothers for these analytes were all significant (p < 0.001). Spatial position weakly predicted TKN (26%) and phosphorus concentrations (23%), but the longitude/latitude smoothers for these analytes were still significant (p ≤ 0.021). The GAM modelling performed worst for manganese (7.8%) and iron (7.5%), indicating weak relationships between concentration and spatial position or depth. Field replicates showed high variability (relative percent differences > 50) in manganese and iron concentrations, which likely contributed to poor model performance in these analytes (Table 1).
Spatial patterns in analyte concentrations were complex and nonlinear over space and depth; however, some general patterns arose (Fig. 3). The northwestern shoreline of the study area tended to have higher concentrations of cadmium, copper, lead, and zinc. Conversely, the northeastern shoreline that was sparsely populated with macrophytes had relatively lower concentrations of these contaminants. Aluminum, arsenic, and TKN were higher in the south, but arsenic concentrations were higher in the northwestern corner of the study area. The depth smoother was significant in the models of arsenic, cadmium, copper, lead, and zinc (p ≤ 0.019), where analyte concentration increased with macrophyte collection depth. Depth was not a significant predictor of aluminum, iron, manganese, phosphorus, or TKN concentrations.

Generalized additive model effects of spatial position and depth on analyte concentration in macrophyte biomass. The upper panel shows the 2-D smoother for the longitude/latitude mean effect centered at zero. The lower panel shows the 1-D smoother for the mean depth effect centered at zero. The dashed lines around the depth smoother shows a 95% confidence interval. The approximate strength of a particular smoother in predicting analyte concentration is indicated by the p value
Toxicity quotients
Toxicity quotients ranged widely depending on the contaminant and benchmark (Fig. 4). In arsenic, benchmark exceedances (i.e., quotients > 1) ranged from 0 to 15%. For cadmium, six of the nine benchmarks had exceedances in at least half the samples. Copper concentrations did not exceed any of the benchmarks examined. For lead, exceedances ranged from 0 to 28% across all benchmarks. However, in one instance, 87% of the lead concentrations were above the INOAEL benchmark. Zinc had the most exceedances relative to the other contaminants of concern, with exceedances ranging from 57 to 100%.

Toxicity quotients calculated from concentrations of contaminants in macrophyte biomass from Coeur d’Alene Lake for various benchmarks. The vertical dashed line shows the threshold where an observed concentration exceeds a benchmark indicating the potential for toxic exposure. Benchmark superscripts indicate citations: aMacDonald et al. (2000), bUSEPA (2002), cUSEPA (2012), and dShayler et al. (2009)
Given the range of exceedances, we choose to evaluate the spatial distribution of analyte concentrations based on benchmarks that had low, moderate, and high exceedance counts. These were the LOAEL, the PEC, and the LB90 benchmarks (Fig. 4). Exceedances occurred most frequently in zinc, followed by cadmium, lead, and arsenic for this subset of benchmarks.
Spatially, the distribution of toxicity quotients reflected the GAM longitude/latitude smoother effect, where the highest quotients were observed in the northwest corner of the study area and along the western shoreline (Fig. 5). For the lead LOAEL benchmark, only one sample had a quotient > 1 and was located in Cottonwood Bay directly across from the Coeur d’Alene River outlet. Zinc exceedances of the LOAEL benchmark extended much further south with a total of 53 samples having toxicity quotients > 1. For the LB90 benchmark most samples had toxicity quotients > 1 for both zinc and cadmium (n = 80, n = 78, respectively). Fewer lead and arsenic concentrations exceeded the LB90 benchmarks with 21 and 5 samples, respectively, having toxicity quotients > 1. Toxicity quotients for the PEC benchmarks were intermediate between the LOAEL and LB90 benchmarks, having between 0 (arsenic) and 58 (zinc) samples with quotients > 1.

The spatial distribution of toxicity quotients (TQ) calculated from concentrations of contaminants in macrophyte biomass from Coeur d’Alene Lake. Three benchmarks (BM) were selected to represent receptor groups with varying sensitivity to contaminants. The population lowest observed adverse effect level (LOAEL) is protective of populations of sediment associated birds and mammals (USEPA 2002, 2012). The probable effect concentration (PEC) was intended for sediment living invertebrates (MacDonald et al. 2000). The lower basin 90th percentile sediment-soil background (LB90) (USEPA 2002) benchmark was used to delineate where macrophyte biomass collected via mechanical harvesting for invasive species control would require careful disposal and harvested biomass would not be suitable for compost
We used the LB90 benchmark to identify areas where macrophyte biomass collected via mechanical harvesting would require careful disposal. Cadmium and zinc had the highest mean LB90 toxicity quotients and spanned the widest latitudinal range (Table 4). Mean quotients for both these analytes were > 1 for all strata except the southernmost stratum. Lead toxicity quotients > 1 were latitudinally restricted to the three northernmost strata. For arsenic, all mean toxicity quotients were < 1. The mean arsenic quotients were the only contaminant to not have a clear north to south gradient being higher at both the northernmost and southernmost strata.
Discussion
Spatial patterns
All analytes (except manganese and iron) showed that spatial position was an important predictor of concentration in macrophytes from Southern Coeur d’Alene Lake. All contaminants of concern except arsenic had increasing concentrations with proximity to the Coeur d’Alene River outlet, which was also reflected in the correlations between macrophyte and sediment concentrations (Horowitz et al. 1993, 1995). Cadmium and zinc concentrations reached further south relative to lead, which likely results from differing transport mechanisms in the Lake for these elements. Lead is most often associated with particulates moved during high energy runoff events, whereas cadmium and zinc are mostly observed in dissolved form within the Lake (Zinsser 2020).
For rooted submerged macrophytes, sediments are often a primary reservoir from which metals and nutrients are drawn into biomass (Jackson 1998). Presumably, the observed macrophyte concentrations from this study are more indicative of sediment conditions rather than water concentrations or species accumulation differences. However, additional sampling would be required to better define the relative importance of the water and sediment contaminant reservoirs for each species. Previous work in the Lake’s watershed showed that differences in contaminant concentrations between M. spicatum and E. canadensis were indistinguishable, but differences between sites were measureable (Scofield et al. 2021). Additionally, species accumulation differences likely cannot be easily leveraged (i.e., selectively remove hyperaccumulator biomass) in a natural lake setting, because invasive macrophytes usually grow interspersed amongst native vegetation (Hansen et al. 2013), hence the focus of this study on composite macrophyte concentrations. Currently, invasive macrophytes cannot be individually removed from the plant bed via mechanical weed harvesting and more selective control techniques (e.g., diver hand pulling) have been made impractical by the wide distribution of M. spicatum in the Lake (CDAT and Avista Corp. 2017).
The spatial patterns in macrophyte aluminum, TKN, and phosphorus were essentially reversed of those observed in cadmium, copper, lead, and zinc, where concentrations increased with distance from the Coeur d’Alene River outlet. Higher southern phosphorus and TKN concentrations may reflect higher nutrient delivery to Southern Coeur d’Alene Lake from the St. Joe River and internal nutrient cycling during thermal stratification (Wood and Beckwith 2008). The higher percentages of more tolerant macrophyte species at southern sites as indicated by the anthropogenic disturbance index appears to corroborate the Lake’s reported trophic gradient. Sensitive macrophyte species in the Lake may also be negatively impacted by annual lake-level drawdown in winter (Mikulyuk et al. 2017; Carmignani and Roy 2021). Macrophytes in the southernmost waters of the Lake may be disproportionately influenced by winter lake-level drawdown because of extensive shallow littoral areas relative to the steeper and deeper shorelines of the main Lake basin (Torso et al. 2020; Scofield et al. 2021).
The Lake’s trophic gradient may also influence the distribution of redox-sensitive arsenic and iron in macrophytes across the study area. Water column anoxia is more common in southern waters of the Lake during summer thermal stratification (Wood and Beckwith 2008), and these conditions can lead to intermediate sediment redox conditions (between − 150 and + 200 mV) which can increase metal desorption from ligands (Jackson 1998). A Coeur d’Alene Lake sediment core experiment showed that release of arsenic, cadmium, and iron into porewater increased substantially under anoxia after two weeks of incubation (Langman et al. 2020). Presumably, higher concentrations of arsenic and iron in porewater could then be drawn into macrophyte biomass at relatively higher levels.
The “U”-shaped latitudinal distribution of arsenic in macrophyte biomass was unexpected, but perhaps this redox mechanism driven by the lake trophic gradient could explain higher arsenic in the southernmost stratum. The shallower morphometry of the southern end of the study area may also contribute to higher arsenic concentrations in macrophyte biomass. Shallow lakes can reportedly have higher arsenic concentrations in biota than deeper lakes with comparable contamination (Hull et al. 2021). In shallow lakes the entire water column can be warmer and mixed which can result in more arsenic exposure to littoral biota, versus less exposure to biota of deeper lakes, because summer thermal stratification isolates the more contaminated hypolimnion from biota (Hull et al. 2021).
Aluminum concentrations in macrophyte biomass were also higher across southern sampling sites. Aluminum uptake by macrophytes is not well understood but is reportedly best described by pH, where low pH increases aluminum accumulation (Gensemer and Playle 1999). The north to south lake trophic gradient (Wood and Beckwith 2008) may lead to lower sediment pH through accumulation of carbon dioxide in bottom waters during summer stratification (Jónasson et al. 1974). The southernmost sampling strata have extensive macrophyte beds compared to the relatively thin bands of macrophytes that parallel the shoreline of northern strata (AquaTechnex 2013). Higher macrophyte biomass in addition to more pelagic carbon production across southern sites may enrich sediments with carbon that could also lower pH, presumably enhancing aluminum uptake.
Macrophyte collection depth was also important in describing analyte concentrations, but only for the contaminants of concern where plants collected from deeper waters tended to have higher contaminant concentrations. Contaminants in sediments are often associated with the finest fraction of its constituents because of higher ligand content and associated binding capacities (Jackson 1998). We suspect that plant biomass collected from shallower waters are more wave swept and likely have proportionally less fine sediment than deeper littoral waters (Benoy and Kalff 1999). Wave energy can limit macrophyte establishment in the breakwaters, but where macrophytes establish they can create a positive feedback loop by slowing water currents and collecting fine sediments that carry nutrient and metal associated particles (Madsen et al. 2001; Mebane et al. 2014). The contaminants and nutrients in settled sediments then become available for uptake into macrophyte biomass.
Toxicity quotients
While reporting toxicity quotients from several benchmarks may be redundant, we felt it necessary to include comparisons across multiple local and national benchmarks to put the observed macrophyte concentrations into context. For the ecological receptors (sediment associated invertebrates, mammals, and birds) most exceedances were attributed to zinc, cadmium, and lead concentrations, which suggest the potential for toxic exposure to macrophyte associated biota (Désy et al. 2002; Spears et al. 2007). The major assumption with this analysis is that receptor exposure via contaminated macrophytes would be similar to that of contaminated sediment or soil. While this approach may be adequate for screening purposes, we caution the reader against making generalizations about actual receptor exposure. Careful modeling accounting for site, species, and analyte specific factors would be necessary to estimate exposure via contaminated macrophytes (Sample et al. 2014), but the reported macrophyte concentrations could provide useful information on dietary inputs in future modeling efforts of this kind.
Coeur d’Alene Lake waterfowl exposure to lead from diet is reportedly less important than incidental ingestion of lead from sediment (Spears et al. 2007). The USEPA (2002) has set a cleanup level of 530 mg/kg lead in sediment and soil for ongoing remediation in the Coeur d’Alene River watershed, and the reported LOAEL toxicity quotients show this lead benchmark. Lead is used as an indicator metal in the Coeur d’Alene River watershed, because it is often collocated with other contaminants and helps provide operational clarity in prioritizing where remedial activities should occur (USEPA 2012). Fortunately, most of the observed lead concentrations in macrophytes from the current study were below the LOAEL benchmark. However, just over half of the macrophyte zinc concentrations were above the LOAEL benchmark, indicating receptors of this benchmark may experience more toxicity from zinc relative to lead assuming high consumption rates of zinc contaminated macrophytes.
Macrophytes are capable of accumulating contaminants like zinc and cadmium at high concentrations (Baldantoni et al. 2005; Xing et al. 2013). The ability of macrophytes to hyperaccumulate zinc and cadmium likely has important food web implications in the Lake. Given zinc and cadmium’s more labile nature in the Lake (Zinsser 2020), the spatial extent of potential toxicity may span a wider area relative to lead. Lead in macrophytes more frequently exceeded the PEC benchmark collected from floodplain lakes with close proximity to the Coeur d’Alene River compared to a site in the Lake’s main basin (Scofield et al. 2021). Conversely, zinc and cadmium were often highest in macrophytes collected from sites in the Lake’s main basin relative to samples from the Coeur d’Alene River floodplain lakes, which presumably is related to the more labile nature of zinc and cadmium relative to lead (Scofield et al. 2021).
The spatial extent of exceedances in composting benchmarks were greater than exceedances of the ecological benchmarks, especially for zinc and cadmium. Given zinc and cadmium’s wider reach into southern waters of the Lake, we suggest that mechanical harvesting only be used to control nuisance macrophytes in the southernmost areas of the Lake. Alternatively, if harvesting were to occur in the more contaminated areas, the collected biomass should be disposed of carefully, perhaps landfilled if contaminant concentrations are high enough to warrant hauling and disposal costs. Additionally, we would recommend monitoring for contaminants of concern in harvested biomass from southern areas of the Lake, if biomass is to be used in compost for agricultural activities. Using the LB90 benchmark appears to be in good agreement with guidance suggested for safe composting from New York State (Shayler et al. 2009), e.g., NYSOIL benchmark.
Nutrient removal potential
Cultural eutrophication of lakes is a pervasive problem (Abell et al. 2022) and is especially important in Coeur d’Alene Lake given the contaminated sediments deposited throughout much of the Lake (Horowitz et al. 1993, 1995). Maintaining high dissolved oxygen in the bottom waters of the Lake is the central management goal of the Coeur d’Alene Lake Management Plan (LMP), where high dissolved oxygen above the sediment water interface acts as an “oxic cap” to reduce the solubility and movement of loosely bound metals into the overlying lake water (IDEQ and CDAT 2009). To keep bottom waters well oxygenated the LMP aims to limit watershed nutrient loading that leads to higher algal production, which then increases microbial respiration and consumption of dissolved oxygen in the Lake’s hypolimnion during summer stratification (Langman et al. 2020).
In addition to reducing the negative impacts created by surface growing invasive macrophytes, mechanical harvesting of troublesome biomass also serves as a nutrient removal tool (Quilliam et al. 2015; Bartodziej et al. 2017). Macrophyte biomass contain significant amounts of nutrients, particularly phosphorus and nitrogen, which are often limiting nutrients for nuisance algae (Abell et al. 2022). Given the difficultly in controlling nonpoint source nutrient loading to lakes (Osgood 2017; Abell et al. 2022), we would argue that harvesting macrophyte biomass presents an opportunity to remove a nontrivial amount of nutrients from the Lake, while simultaneously lessening nuisance biomass. Using mean nutrient concentrations from the southernmost Lake stratum and the Chatcolet Lake peak macrophyte biomass (1871 g/m2) reported by Torso et al. (2020), we estimated that a total (dry mass) of 278 kg of phosphorus and 1372 kg of TKN could be removed by harvesting half of the cuttable biomass in a typical treatment area (9.1 ha).
However, we are not advocating for the widespread removal of littoral vegetation, since macrophytes play a critical role in lake ecosystem health (Abell et al. 2022). Rather we suggest that targeted harvesting of invasive plants could provide relief from nuisance biomass and a means to remove some nutrients from the Lake. Additionally, mechanical harvesting should not be used in areas with small invasive species populations given their tendency to spread via fragmentation (Quilliam et al. 2015). Since invasive watermilfoils (M. spicatum and its hybrid) are well established and widespread in Coeur d’Alene Lake (Liao et al. 2016; CDAT and Avista Corp. 2017) we do not expect such targeted harvesting to create new infestations. However, the aquatic macrophyte community of the Lake should be closely monitored to verify this assumption when harvesting is implemented.
Conclusions
Concentrations of cadmium, copper, lead, and zinc in above sediment macrophyte biomass showed distinct spatial distributions, where the most contaminated biomass was found near the Coeur d’Alene River outlet (main contamination source) and along the western shoreline of Southern Coeur d’Alene Lake. Arsenic concentrations in macrophyte biomass diverged from this pattern, being higher in both samples from near the contamination source and at the southern end of the study area (Chatcolet Lake). Concentrations of arsenic, cadmium, copper, lead, and zinc also tended to be higher in macrophyte biomass collected from deeper waters.
The spatial pattern of nutrient (phosphorus and total Kjeldahl nitrogen) and aluminum concentrations in biomass were reversed from the contaminant metals, being generally higher in macrophyte biomass from the southern end of the study area. Iron and manganese concentrations were much more evenly distributed in macrophyte biomass.
Numerous screening level benchmarks have been developed for assessing contamination in sediment and soil. Since macrophyte specific benchmarks have not been widely developed, we used these sediment and soil benchmarks to gauge the extent of contamination in macrophyte biomass. Toxicity quotients calculated from ecological receptor (i.e., sediment associated organisms) based benchmarks exceeded one less frequently compared to background level benchmarks. Concentrations of zinc in macrophyte biomass most frequently exceeded benchmarks followed by cadmium and then lead concentrations. Copper and arsenic concentrations rarely exceeded benchmarks.
Invasive macrophyte control via mechanical harvesting should be limited to areas with contaminant concentrations below background level benchmarks unless careful disposal of cut biomass can be implemented. Additionally, in these “clean” areas it would be possible to remove a nontrivial amount of nutrients with mechanical harvesting of macrophyte biomass as a measure to slow cultural eutrophication.
Data and code availability
The software description, data, and scripts used in the reported analyses are available at the following data repository: https://doi.org/10.5281/zenodo.7768213.
References
Abell JM, Özkundakci D, Hamilton DP, Reeves P (2022) Restoring shallow lakes impaired by eutrophication: Approaches, outcomes, and challenges. Crit Rev Environ Sci Technol 52:1199–1246. https://doi.org/10.1080/10643389.2020.1854564
AquaTechnex (2013) 2013 Coeur d’Alene Lake Eurasian Milfoil control program task two. Mapping report, Bellingham, WA
Baldantoni D, Maisto G, Bartoli G, Alfani A (2005) Analyses of three native aquatic plant species to assess spatial gradients of lake trace element contamination. Aquat Bot 83:48–60. https://doi.org/10.1016/j.aquabot.2005.05.006
Bartodziej WM, Blood SL, Pilgrim K (2017) Aquatic plant harvesting: an economical phosphorus removal tool in an urban shallow lake. J Aquat Plant Manag 55:26–34
Benoy GA, Kalff J (1999) Sediment accumulation and Pb burdens in submerged macrophyte beds. Limnol Oceanogr 44:1081–1090. https://doi.org/10.4319/lo.1999.44.4.1081
Bergbusch NT, Hayes NM, Simpson GL, Leavitt PR (2021) Unexpected shift from phytoplankton to periphyton in eutrophic streams due to wastewater influx. Limnol Oceanogr 66:2745–2761. https://doi.org/10.1002/lno.11786
Carmignani JR, Roy AH (2021) Annual winter water-level drawdowns influence physical habitat structure and macrophytes in Massachusetts, USA, lakes. Ecosphere 12:e03442. https://doi.org/10.1002/ecs2.3442
[CDAT and Avista Corp.] Coeur d’Alene Tribe, Avista Corporation (2017) Coeur d’Alene Reservation aquatic weed management plan. In: 4(e) condition no. 7 Spokane River hydroelectric project FERC project no. 2545
Ceschin S, Bellini A, Scalici M (2021) Aquatic plants and ecotoxicological assessment in freshwater ecosystems: a review. Environ Sci Pollut Res 28:4975–4988. https://doi.org/10.1007/s11356-020-11496-3
Désy JC, Amyot M, Pinel-Alloul B, Campbell PGC (2002) Relating cadmium concentrations in three macrophyte-associated freshwater invertebrates to those in macrophytes, water and sediments. Environ Pollut 120:759–769. https://doi.org/10.1016/S0269-7491(02)00174-4
Gensemer RW, Playle RC (1999) The bioavailability and toxicity of aluminum in aquatic environments. Crit Rev Environ Sci Technol 29:315–450. https://doi.org/10.1080/10643389991259245
Gustavson KE, Barnthouse LW, Brierley CL et al (2007) Superfund and mining megasites. Environ Sci Technol 41:2667–2672. https://doi.org/10.1021/es0725091
Hansen GJA, Zanden MJV, Blum MJ et al (2013) Commonly rare and rarely common: comparing population abundance of invasive and native aquatic species. PloS One 8:e77415. https://doi.org/10.1371/journal.pone.0077415
Helsel DR (2012) Statistics for censored environmental data using Minitab and R, 2nd edn. John Wiley & Sons, Hoboken, New Jersey
Horowitz AJ, Elrick KA, Robbins JA, Cook RB (1993) The effect of mining and related activities on the sediment-trace element geochemistry of Lake Coeur d’Alene. U.S. Geological Survey; Earth Science Information Center, Idaho, U.S.A. Part II, subsurface sediments. Open-File Report 93-656.
Horowitz AJ, Elrick KA, Robbins JA, Cook RB (1995) A summary of the effects of mining and related activities on the sediment-trace element geochemistry of Lake Coeur d’Alene, Idaho, USA. J Geochem Explor 52:135–144. https://doi.org/10.1016/0375-6742(94)00041-9
Hull EA, Barajas M, Burkart KA et al (2021) Human health risk from consumption of aquatic species in arsenic-contaminated shallow urban lakes. Sci Total Environ 770:145318. https://doi.org/10.1016/j.scitotenv.2021.145318
[IDEQ and CDAT] Idaho Department of Environmental Quality, Coeur d’Alene Tribe (2009) Coeur d’Alene lake management plan. Coeur d’Alene, Idaho
Jackson LJ (1998) Paradigms of metal accumulation in rooted aquatic vascular plants. Sci Total Environ 219:223–231. https://doi.org/10.1016/S0048-9697(98)00231-9
Jónasson PM, Lastein E, Rebsdorf A (1974) Production, insolation, and nutrient budget of eutrophic Lake Esrom. Oikos 25:255–277. https://doi.org/10.2307/3543944
Langman JB, Ali JD, Child AW et al (2020) Sulfur species, bonding environment, and metal mobilization in mining-impacted lake sediments: column experiments replicating seasonal anoxia and deposition of algal detritus. Minerals 10:849. https://doi.org/10.3390/min10100849
Lewis MA, Weber DE, Stanley RS, Moore JC (2001) The relevance of rooted vascular plants as indicators of estuarine sediment quality. Arch Environ Contam Toxicol 40:25–34. https://doi.org/10.1007/s002440010145
Liao FH, Wilhelm FM, Solomon M (2016) The effects of ambient water quality and Eurasian watermilfoil on lakefront property values in the Coeur d’Alene area of northern Idaho, USA. Sustainability 8:44. https://doi.org/10.3390/su8010044
MacDonald DD, Ingersoll CG, Berger TA (2000) Development and evaluation of consensus-based sediment quality guidelines for freshwater ecosystems. Arch Environ Contam Toxicol 39:20–31. https://doi.org/10.1007/s002440010075
Madsen JD, Chambers PA, James WF et al (2001) The interaction between water movement, sediment dynamics and submersed macrophytes. Hydrobiologia 444:71–84. https://doi.org/10.1023/A:1017520800568
Mebane CA, Simon NS, Maret TR (2014) Linking nutrient enrichment and streamflow to macrophytes in agricultural streams. Hydrobiologia 722:143–158. https://doi.org/10.1007/s10750-013-1693-4
Mikulyuk A, Barton M, Hauxwell J et al (2017) A macrophyte bioassessment approach linking taxon-specific tolerance and abundance in north temperate lakes. J Environ Manage 199:172–180. https://doi.org/10.1016/j.jenvman.2017.05.012
Newete SW, Byrne MJ (2016) The capacity of aquatic macrophytes for phytoremediation and their disposal with specific reference to water hyacinth. Environ Sci Pollut Res 23:10630–10643. https://doi.org/10.1016/j.jenvman.2015.01.046
[NRC] National Research Council (2005) Superfund and mining megasites: lessons from the Coeur d’Alene River Basin. National Academies Press, Washington, D.C.
Osgood RA (2017) Inadequacy of best management practices for restoring eutrophic lakes in the United States: guidance for policy and practice. Inland Waters 7:401–407. https://doi.org/10.1080/20442041.2017.1368881
Pedersen EJ, Miller DL, Simpson GL, Ross N (2019) Hierarchical generalized additive models in ecology: an introduction with mgcv. PeerJ 7:e6876. https://doi.org/10.7717/peerj.6876
Quilliam RS, van Niekerk MA, Chadwick DR et al (2015) Can macrophyte harvesting from eutrophic water close the loop on nutrient loss from agricultural land? J Environ Manage 152:210–217. https://doi.org/10.1016/j.jenvman.2015.01.046
Sample BE, Schlekat C, Spurgeon DJ et al (2014) Recommendations to improve wildlife exposure estimation for development of soil screening and cleanup values. Integr Environ Assess Manag 10:372–387. https://doi.org/10.1002/ieam.1482
Scofield BD, Torso K, Fields SF, Chess DW (2021) Contaminant metal concentrations in three species of aquatic macrophytes from the Coeur d’Alene Lake basin, USA. Environ Monit Assess 193:683. https://doi.org/10.1007/s10661-021-09488-y
Shayler H, McBride M, Harrison E (2009) Guide to soil testing and interpreting results. Cornell Waste Management Institute, Ithaca, New York
Singh NK, Raghubanshi AS, Upadhyay AK, Rai UN (2016) Arsenic and other heavy metal accumulation in plants and algae growing naturally in contaminated area of West Bengal, India. Ecotoxicol Environ Saf 130:224–233. https://doi.org/10.1016/j.ecoenv.2016.04.024
Spears BL, Hansen JA, Audet DJ (2007) Blood lead concentrations in waterfowl utilizing Lake Coeur d’Alene, Idaho. Arch Environ Contam Toxicol 52:121–128. https://doi.org/10.1007/s00244-006-0061-z
Torso K, Scofield BD, Chess DW (2020) Variations in aquatic macrophyte phenology across three temperate lakes in the Coeur d’Alene Basin. Aquat Bot 162:103209. https://doi.org/10.1016/j.aquabot.2020.103209
[USEPA] US Environmental Protection Agency, Region 10 (2021) Fifth five-year review report for Bunker Hill mining and metallurgical complex superfund facility. Seattle, Washington
Wood SN (2017) Generalized additive models: an introduction with R, 2nd edn. Chapman and Hall/CRC, New York
Wood MS, Beckwith MA (2008) Coeur d’Alene Lake, Idaho: Insights gained from limnological studies of 1991-92 and 2004-06. Geological Survey (U.S.)
Xing W, Wu H, Hao B et al (2013) Bioaccumulation of heavy metals by submerged macrophytes: looking for hyperaccumulators in eutrophic lakes. Environ Sci Technol 47:4695–4703. https://doi.org/10.1021/es303923w
Zinsser LM (2020) Trends in concentration, loads, and sources of trace metals and nutrients in the Spokane River Watershed, northern Idaho, water years 1990–2018. U.S. Geological Survey, Reston, Virginia
R Core Team (2022) R: A language and environment for statistical computing
[USEPA] US Environmental Protection Agency (2002) The Bunker Hill mining and metallurgical complex: operable unit 3, record of decision. Region 10
[USEPA] US Environmental Protection Agency (2012) Interim record of decision (ROD) amendment, Upper Basin of the Coeur d’Alene River. Bunker Hill mining and metallurgical complex superfund site. Region 10
[USEPA] US Environmental Protection Agency (2014) Method 6010D (SW-846): Inductively coupled plasma-atomic emission spectrometry, revision 4, Washington, D.C.
[USEPA] US Environmental Protection Agency (1993) Method 351.2, Revision 2.0: Determination of total Kjeldahl nitrogen by semi-automated colorimetry, Cincinnati, Ohio
[USEPA] US Environmental Protection Agency (1996) Method 3050B: Acid digestion of sediments, sludges and soils
Acknowledgements
Firstly, we thank the Schitsu’umsh whose support made this work possible. We also thank Darren Lantzer of Tshimakain Creek Laboratories and SVL Analytical, Inc. for processing samples and consultation on analytical results. Lastly, we are grateful to Rebecca Stevens, Laura Laumatia, and the anonymous reviewers who provided critical reviews that significantly improved the manuscript.
Funding
This work was funded by the Coeur d’Alene Tribe’s Water Resources Program and a grant from the US Bureau of Reclamation (R17AP00237-002).
Author information
Authors and Affiliations
Contributions
Ben D. Scofield: conceptualization, data curation, formal analysis, investigation, methodology, writing—original draft, and writing—review and editing. Scott F. Fields: funding acquisition, project administration, supervision, and writing—review and editing. Dale W. Chess: conceptualization, methodology, supervision, and writing—review and editing.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Responsible Editor: Christian Gagnon
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Scofield, B.D., Fields, S.F. & Chess, D.W. Aquatic macrophytes show distinct spatial trends in contaminant metal and nutrient concentrations in Coeur d’Alene Lake, USA. Environ Sci Pollut Res 30, 66610–66624 (2023). https://doi.org/10.1007/s11356-023-27211-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-023-27211-x