
Abstract
High biomass energy plants are a promising alternative to hyperaccumulators for the remediation of heavy metals (HMs). Arundo donax L. (A. donax) is a rapidly growing rhizomatous grass with high biomass production. However, the feasibility of using A. donax for the phytoremediation of combined mercury (Hg) and cadmium (Cd) pollution under neutral conditions is unclear. In this study, a hydroponic experiment was performed to investigate the impact of Hg–Cd stress on the growth and physiological properties of A. donax and HMs accumulation and transformation. Either single Hg or Cd stress slightly enhanced stem height, fresh biomass, and chlorophyll content, whereas combined Hg–Cd stress reduced these parameters. Furthermore, combined Hg–Cd stress increased the leaf content of malondialdehyde in A. donax, indicating that the combined pollution aggravated oxidative stress in A. donax. Hg volatilization was observed during the 10-day experiment, implying that a portion of the Hg2+ was transformed into Hg0 by A. donax. The bioaccumulation factor (BAF) values of A. donax were far greater than 1 for both Hg and Cd, whereas the translocation factor (TF) values were less than 1, indicating that phytovolatilization and phytostabilization rather than phytoextraction contributed to the remediation of Hg and Cd by A. donax. The solution pH decreased at the beginning of the experiment, suggesting that acidic root exudates of A. donax facilitated the accumulation and transformation of Hg under neutral conditions. Overall, the effects of Cd on Hg accumulation and transformation by A. donax followed the rule of “low promotion and high inhibition.” This study demonstrates that A. donax is a potential candidate for the phytoremediation of combined Hg–Cd pollution under neutral conditions.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Mercury (Hg) is a toxic heavy metal (HM), which is introduced to the environment through natural processes and anthropogenic activities (Wang et al. 2021). Hg pollution has become a global environmental problem nowadays. For example, in the Wanshan Mercury Mine District, which is located in the Pacific Rim Mercury Mineralization Zone and is the largest historic Hg-producing district in Guizhou Province, China, long-term mining and smelting activities have produced approximately 125.8 million tons of mine waste (Yin et al. 2016), and the contents of Hg in the atmosphere, water, and soil all far exceed background values in the province (Li et al. 2008; Zhang et al. 2010). Hg can be accumulated by aquatic organisms, plants, and human beings, posing a significant threat to ecosystem function and human health (Barone et al. 2021; Lominchar et al. 2019; Raj et al. 2020a, b; Yabanli et al. 2014; Yozukmaz et al. 2018). Feasible remediation technologies for Hg contamination are urgently needed.
Current remediation strategies for heavy metal (HM)-polluted soils include physicochemical and biological techniques. Among biological techniques, phytoremediation is an eco-friendly and low-cost technology that utilizes plants to absorb pollutants from contaminated water and soil. However, most hyperaccumulators have shallow root systems and low biomass production, and attention is increasingly turning toward fast-growing energy plants with high biomass production, such as Miscanthus × giganteus, Sorghum dochna, Ricinus communis L., Jatropha curcas, and Arundo donax L. var.versicolor Kunth (A. donax) (Pandey et al. 2016). These plants are HM tolerant and highly adaptable. After remediation, the aerial portions of the plants are generally used as biofuels to meet energy demands. Therefore, energy plants can provide not only biomass yield but also ecosystem services for pollution remediation. This study focuses on A. donax, a tall perennial rhizomatous grass with fast growth, high biomass, high adaptability, and strong tolerance to various HMs, such as Pb, Cd, Mn, Cr, Hg, As, Zn, and Cu (Cristaldi et al. 2020; Liu et al. 2019; Zhang et al. 2021).
In general, the mechanisms of phytoremediation can be classified into five main pathways: plant extraction, plant degradation, plant stabilization, plant volatilization, and plant filtration. HMs are absorbed by plant roots and transported to aboveground shoots or leaves via the xylem (Zhang et al. 2021). Hg generally exists in elemental (Hg0), ionic (Hg2+), hydroxide (Hg(OH)2), sulfide (HgS), and methyl (MeHg) forms in the environment, and transformations between these forms can occur under certain conditions. Hg2+ is the predominantly bioavailable species for plant uptake (Heaton et al. 2005; Raj and Maiti 2020) and is readily adsorbed by plant roots. However, only a small proportion of this Hg2+ transported to the aerial parts of the plant; thus, it predominantly accumulates in roots (Chen and Yang 2012). In general, acidic conditions facilitate HM accumulation in plants because most HMs are transformed into insoluble forms under neutral and weakly alkaline conditions. However, in a field trial study in the Guizhou mining area, a karst mountainous region with a soil pH around 7.0, we found that the content of Hg in the aerial parts of A. donax could reach 4.16 mg kg−1, with a yield of approximately 4.0 t (dry weight) ha−1 year−1 under neutral conditions. How does A. donax accumulate, translocate, and transform Hg from soil neutral conditions?
Coexisting HMs can synergistically, additively, or antagonistically affect HMs accumulation and translocation in plants. For example, Zn increases the uptake, translocation, and accumulation of Cd by Brassica juncea (Du et al. 2020; Kutrowska et al. 2017). Conversely, As decreases the accumulation and translocation of Zn in Pteris vittata (Zhang et al. 2014). Hg inhibits the uptake and translocation of As but enhances the toxicity of As(III) in rice seedlings (Ren et al. 2014). The interaction effects of coexisting HMs depend on many factors, such as plant genotypes and ecotypes and metal concentrations and types (Abid et al. 2019). Hg and Cd are main group IIB elements and thus have similar chemical properties. In fact, Cd is frequently found in Hg ore bodies and usually coexists with Hg in mercury mining areas (Chang 2019). However, little information is available about the effect of Cd on the accumulation, translocation, and transformation of Hg in plants.
HMs uptake and accumulation negatively affect plant physiological processes, including plant growth, photosynthesis, and the antioxidant system (Chen and Yang 2012; Pu et al. 2019; Raj and Maiti 2020). Plant cells have developed mechanisms to resist HMs stress. Superoxide dismutase (SOD) is an important antioxidant enzyme that can eliminate reactive oxygen species (ROS) induced by metal stress to prevent excessive oxidation of membrane lipids. Glutathione (GSH), the main non-protein mercaptan in plants, is an indicator of plant antioxidant capacity. GSH can also effectively eliminate ROS, lipid peroxides (LOOHs), and other harmful molecules to maintain normal cell growth and development (Mwamba et al. 2016). However, the effects of Hg–Cd stress on the growth and physiological properties of plants, including A. donax, are unclear.
To address these gaps, a hydroponic experiment was performed at pH 7.0 in this study to fulfill the following objectives: (i) understand the effects of combined Hg–Cd stress on the growth and physiological properties of A. donax; (ii) determine the accumulation, translocation, and transformation characteristics of Hg in A. donax under neutral conditions in the presence or absence of Cd; and (iii) propose feasible mechanisms for the phytoremediation of Hg by A. donax.
Materials and methods
Experimental design
Seedlings of A. donax were collected from the seedling breeding base of our research group in Nanjing, Jiangsu Province, China. Healthy A. donax seedlings of equal size were selected, purged, and cultured in Hoagland’s nutrient solution. After acclimation for 2 weeks, the vigorous seedlings were transferred to glass pots containing 400 mL of 0.25-strength Hoagland’s nutrient solution at pH 7.0. The outside of the glass pot was wrapped with aluminum foil to protect against light.
To understand the Hg accumulation and translocation capabilities of A. donax under combined Hg–Cd stress conditions, nine treatments in which the seedlings were exposed to nutrient solution containing Hg(NO3)2 (0, 0.5 or 2 mg·L−1) and/or Cd(NO3)2 (0, 0.5 or 2 mg·L−1) were established: ① CK; ② Hg1: 0.5 mg L−1 Hg(NO3)2; ③ Hg2: 2 mg L−1 Hg(NO3)2; ④ Cd1: 0.5 mg L−1 Cd(NO3)2; ⑤ Cd2: 2 mg L−1 Cd(NO3)2; ⑥ Hg1Cd1: 0.5 mg L−1 Hg(NO3)2 + 0.5 mg L−1 Cd(NO3)2; ⑦Hg2Cd2: 2 mg L−1 Hg(NO3)2 + 2 mg L−1 Cd(NO3)2; ⑧Hg1Cd2: 0.5 mg L−1 Hg(NO3)2 + 2 mg L−1 Cd(NO3)2; ⑨ Hg2Cd1: 2 mg L−1 Hg(NO3)2 + 0.5 mg L−1 Cd(NO3)2. The solution pH was maintained at 7.0 by adjustment with 0.01 M NaOH or HCl.
The phytovolatilization of Hg was quantified by enclosing the plant in a gastight compartment (Fig. 1). The experimental unit consisted of one air pump, one gastight volatilization chamber, and a Hg trap. The Hg vapor released from the plant was trapped in 150 mL of 5% KMnO4 dissolved in 1 M H2SO4. The plant was placed in a plant growth chamber at an average temperature regime of 25/15 °C (day/night) without supplementary illumination for 10 days. Each treatment had three replicates.

Experimental apparatus used to capture volatilized Hg from Arundo donax L. A Air inlet; B gastight shoot compartment (4.5 L); C Arundo donax L. seedling; D air outlet; E glass pot; F permanganate acid trap
After the 10-day hydroponic experiment, the A. donax seedlings were harvested, washed thoroughly with ultrapure water, and separated into roots and aerial parts. The principal root length, stem height, and fresh weight of each plant were determined, and the roots and aerial parts from each treatment were freeze dried and ground to powder for total Hg and Cd analyses.
Physiological and biochemical measurements
The chlorophyll content of the leaves was determined using a SPAD-502 instrument (Konica Minolta, Japan). The relative chlorophyll content was determined from the dual-wavelength optical absorbance (at 620 and 940 nm) of the leaf samples. The average value of 5 selected points on each leaf was calculated to reflect the chlorophyll content of the whole leaf (Sun et al. 2021).
Malondialdehyde (MDA) content was measured using the thiobarbituric acid method (Jiang et al. 2022). Fresh leaves (∼ 0.50 g) were crushed in a mortar, and 2 mL of 0.05 M (pH 7.8) phosphate buffer was added. The liquid mixture was centrifuged at 4500 rpm for 10 min, and the supernatant (MDA extract solution) was retained. Next, 2 mL of the MDA extract solution and 3 mL of 5% trichloroacetic acid solution were heated in a boiling water bath for 10 min and centrifuged at 4500 rpm for 10 min. The MDA content was determined by measuring the absorbance at 532 nm and 600 nm in a spectrophotometer (Shanghai Precision Instrument Co., Ltd., model v-5800).
SOD activity was assayed following Shah et al. (Shah and Nahakpam 2012). Fresh leaves (∼ 0.30 g) were homogenized in 5 mL of ice-cold 50 mM phosphate buffer at pH 6.5 and centrifuged at 10,000 g for 20 min at 0 to 4 °C. The absorbance of the supernatant was used for the measurement of SOD activities, which was determined in an ELISA reader (Beijing Perlong New Technology Co., Ltd., DNM-9602) at 450 nm.
GSH content was determined according to the method of Xun et al. (Xun et al. 2017). Briefly, leaf samples (∼ 0.50 g) were homogenized in 5% trichloroacetic acid and centrifuged at 15,000 rpm for 10 min. Two milliliters of the supernatant were combined with 2.6 mL of 150 mM NaH2PO4 buffer and 0.2 mL of 5,5′-dithiobis-(2-nitrobenzoic acid) (DTNB), and GSH content was determined by measuring the absorbance of the mixture at 412 nm in a spectrophotometer (Shanghai Precision Instrument Co., Ltd., model v-5800). Three individual replicates were performed for each plant sample.
Hg and Cd analysis
The total Hg concentration in the plant was determined following the method of Zhu et al. (Zhu et al. 2021). Roots or aerial parts (∼ 0.20 g) were mixed with 5 mL of HNO3 and 3 mL of H2O2 in a digestion vessel, heated at 140 °C for 4 h and diluted to 25 mL with ultrapure water. The total Hg concentration in the permanganate acid trap was assayed as follows: 5 mL of solution from the trap was mixed with 0.5 mL of HNO3 and 0.3 mL of H2O2 in a digestion tube, heated in a boiling water bath for 1 h and diluted to 10 mL. The Hg concentration in the digestion was determined by atomic fluorescence spectrophotometry (Model AFS-930 Jitian, Beijing, China).
The total Cd concentration in the plant was determined using the method of Zhou et al. (Zhou et al. 2020). Briefly, plant samples (∼ 0.20 g) were digested with 6 mL of HNO3 and 2 mL of H2O2 in a digestion vessel heated at 105 °C for 7 h and then diluted to 25 mL with ultrapure water. The Cd concentration in the digestion was determined by atomic absorption spectrophotometry (Varian SpectrAA 240Z or 220FS; Varian, Palo Alto, CA).
Replicates, blanks, and reference materials GBW10048 (celery) were included for quality control throughout the analytical procedure. About 95.7–101% of Cd and 94.6–98.5% of Hg in GBW10048 were recovered.
The phytoextraction ability of A. donax was assessed using the bioaccumulation factor (BAF), which is the ratio of the concentration of metals in the plant to the concentration in the environment. When the BAF is greater than 1, the plant is considered as an accumulator. The translocation factor (TF), which is the ratio of the metal concentration in the aerial parts to the metal concentration in the roots, was used to evaluate the ability of the plant to transfer metals from the roots to the aerial parts (stems and leaves).
Data analysis
Results obtained for the A. donax experiment were expressed as mean ± Standard Deviation (SD). Differences were tested for significance by analysis of variance (ANOVA). Subsequent multiple comparisons among means were examined by the least significance difference (LSD) test (p < 0.05) in SPSS 20.0. All graphics were generated using Origin 8.0.
Results and discussion
Effects of Hg and Cd on plant growth and biomass
No symptoms of toxicity in A. donax, such as leaf fall, necrosis, or chlorosis, were observed in any of the treatments during the experiment (Table 1). When A. donax was exposed to single Hg or Cd, the stem height, fresh biomass, and chlorophyll content increased as the solution concentration of Hg or Cd increased from 0.5 to 2.0 mg L−1, suggesting that A. donax is highly tolerant of Hg and Cd and that single Hg or Cd stress may even stimulate growth. These findings are consistent with those of Cristaldi et al. (Cristaldi et al. 2020). The growth-promoting effect of single Hg or Cd exposure on A. donax may be due to the hormesis effect, which has been observed in many plants (Ozturk et al. 2010). However, compared with the control, the roots of A. donax exposed to single Hg or Cd were 34.4–45.4% or 9.8–16.9% shorter, respectively. In general, roots are the main interface for ion exchange between plants and the environmental medium, and they are the most sensitive organ to the stress of HMs in the environment. Hg or Cd stress may have inhibited mitosis in root tip meristematic cells, leading to chromosomal aberrations and reduced root growth due to asymmetry among the root epidermis, cortex cells, and cell gap (He et al. 2010). The negative effects of Hg stress on root growth were much greater than those of Cd stress.
Compared to the single Hg or Cd stress treatments, the combined Hg–Cd stress treatments significantly reduced the stem height, root length, and fresh biomass of A. donax, indicating that the combination aggravated the toxic effects of these HMs on plant growth. These results are similar to those of Ren et al. (2014). Co-exposure to Hg and Cd increases the toxicity of these HMs via effects on the plant cell membrane and transporters that reduce nutrient transport and inhibit plant growth (Cargnelutti et al. 2006). With the exception of the Hg1Cd1 treatment, all combined Hg and Cd treatments significantly reduced A. donax leaf chlorophyll content compared with the control and single Hg or Cd stress treatments, consistent with a previous report by Pu et al. (2019). These findings suggest that Hg and Cd co-exposure within a certain concentration range can promote chlorophyll generation in A. donax leaves in accordance with the rule of “low promotion and high inhibition”; that is, low concentrations have positive effects, while high concentrations have negative effects (Guo et al. 2019).
Accumulation of Hg and Cd in A. donax
Hg accumulation characteristics
The contents of Hg in the roots and aerial parts of A. donax are shown in Fig. 2. The Hg content ranged from 12.47 ± 2.54 to 35.57 ± 1.76 in the roots and from 7.72 ± 2.20 to 26.27 ± 4.65 mg kg−1 in the aerial parts of A. donax, indicating that the roots of A. donax had a much higher capacity for Hg accumulation than the aerial parts. Furthermore, the accumulation of Hg in A. donax roots and aerial parts increased with increasing Hg concentration from 0.5 to 2.0 mg L−1 in both the single Hg and combined stress treatments. These results indicate that A. donax has high Hg accumulation efficiency, in accordance with previous studies (Chen and Yang 2012). Roots are the main pathway for HM absorption by plants, and the well-developed root systems of A. donax can release large amounts of root exudates to facilitate HM accumulation and translocation. Previous studies have shown that HMs are readily adsorbed by and primarily accumulate in roots, with only 1.20 to 3.80% transported aboveground (Chen and Yang 2012; Liu et al. 2020; Ren et al. 2014). The process of translocation from roots to aerial parts generally involves several steps, including symplastic absorption in roots, root sequestration, xylem loading, and storage in leaf epidermal and mesophyll cells. In this study, the accumulation of Hg in the aerial parts accounted for 32.98–44.27% of the total Hg accumulation by A. donax, which indicates that A. donax has rather high translocation efficiency. A. donax is rich in cellulose and has hollow stems, which may facilitate the translocation of Hg from the roots to the aerial parts by xylem loading (Zhang et al. 2021).

Mass fractions of Hg in the aerial parts and roots of Arundo donax L. in the different treatments (bars with different letters are significantly different at p < 0.05)
The most striking result was the generation of volatile Hg, as shown in Fig. 3. Volatile Hg was detected in all Hg stress treatments at the end of the experiment. The mass of volatile Hg ranged from 1.81 ± 0.36 in Hg1 (0.5 mg L−1 Hg) to 899.90 ± 97.73 ng in Hg2Cd1 (2 mg L−1 Hg and 0.5 mg L−1 Cd). Thus, A. donax can accumulate Hg from the environment and convert ionic Hg into gaseous Hg. Photochemical reduction has been proposed as an important route of transformation of Hg2+ to Hg0 (Ma et al. 2015), but Hg reduction and volatilization in plants can also be mediated by enzymatic reduction (Haque et al. 2010). The glass pots were protected from light by aluminum foil throughout the experiment, thus excluding photochemical reduction as the mechanism of volatile Hg generation. Consequently, the formation of volatile Hg in our experiment can be attributed to enzymatic reduction.

Mass of volatile Hg captured from Arundo donax L. in the different treatments (bars with different letters are significantly different at p < 0.05)
Effect of Cd accumulation on Hg uptake by A. donax
As shown in Fig. 4, A. donax also accumulated Cd. The concentration of Cd ranged from 3.15 ± 0.94 to 22.74 ± 0.84 mg kg−1 in the roots and from 1.25 ± 0.19 to 7.74 ± 1.83 mg kg−1 in the aerial parts. Similar to Hg, the accumulation of Cd was much higher in the roots than in the aerial parts of A. donax, and increasing the solution concentration of Cd enhanced the accumulation of Cd in both the roots and aerial parts of A. donax. However, the accumulation and translocation efficiency of Cd were much lower than those of Hg, indicating that A. donax prefers to take up Hg rather than Cd. These findings are also consistent with those of Cristaldi et al. (2020).

Mass fractions of Cd in the aerial parts and roots of Arundo donax L. in the different treatments (bars with different letters are significantly different at p < 0.05)
To further confirm the effect of Cd on Hg uptake, accumulation, and transformation in A. donax, four different combination treatments (Hg1Cd1, Hg2Cd2, Hg1Cd2, and Hg2Cd1) were analyzed in this study. As shown in Fig. 2, the presence of Cd enhanced the accumulation of Hg in the roots and aerial parts of A. donax under both low and high Hg stress. Furthermore, the accumulation of Hg in roots increased as the concentration of Cd increased from 0.5 to 2.0 mg L−1 under both low and high Hg stress (Fig. 2). Increasing the Cd concentration also promoted the accumulation of Hg in the aerial parts under high Hg stress but not low Hg stress.
In addition, the presence of Cd impacted Hg volatilization by A. donax. Under low Hg stress, the mass of volatile Hg increased slightly as the concentration of Cd increased (Fig. 3). Under high Hg stress, a low concentration of Cd promoted Hg volatilization, whereas a high Cd concentration significantly inhibited the Hg volatilization process (Fig. 3). Similar “low promotion and high inhibition” effects of coexisting HMs on Hg volatilization in plants have been observed in previous studies (Guo et al. 2019) and have been attributed to changes in plant enzyme activities induced by the combined stress. Another possibility is that coexisting HMs compete for HM transporters in roots (Du et al. 2020), thus inhibiting their accumulation and translocation compared with single HM stress. Saifullah et al. (2014) reported that phytochelatins (PCs) induced by Cd stress in roots bind Cd2+ and prevent Cd translocation from the roots to the aerial parts. However, when Zn2+ and Cd2+ are both present, Zn2+ competes with Cd2+ for PC binding, thus decreasing Cd-PC formation and promoting Cd translocation in plants. Zn, Cd, and Hg are all main group IIB elements and thus have similar accumulation and translocation properties in plants. Therefore, in A. donax, Hg stress likely induces the production of PCs to bind Hg2+, but the formation of Hg-PCs is reduced in the presence of Cd, which competes with Hg for PC binding. As a result, Cd promotes Hg accumulation and translocation in A. donax.
Changes in pH during the experiment
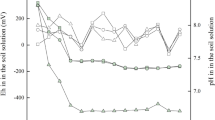
In general, Hg2+ is the most bioavailable form of Hg for uptake and accumulation in plants, but Hg2+ is susceptible to inert precipitate formation when the environmental pH is ≥ 7.0. To determine whether A. donax can accumulate Hg under neutral conditions, the pH of the culture solution was adjusted to 7.0 at the beginning of the experiment, and the pH value in the control treatment was monitored daily. As shown in Fig. 5, the solution pH decreased from 7.0 to 6.1 ± 0.07 on day 1 but increased from 6.1 ± 0.07 on day 2 to 7.4 ± 0.3 on day 10. Plants typically release large amounts of root exudates such as organic acids, amino acids, and fatty acids, which can decrease solution pH (Shi et al. 2011), enhance HM activity, and stimulate HM accumulation in plants (Ubeynarayana et al. 2021). A. donax is a perennial plant with a well-developed root system that can release acidic exudates during growth, resulting in a decrease in solution pH. In addition to acidic root exudates, plants can release alkaline root exudates such as alkaloids. Arundamine, donaxine, and N,N-dimethyltryptamine have been identified as the major alkaloids released by A. donax roots (Khuzhaev et al. 1994). Cano-Ruiz et al. (2021) also reported that soil pH increased significantly after planting A. donax on farmland for 2 years. In the present study, the release of acidic root exudates initially decreased the solution pH and promoted the uptake and accumulation of Hg by A. donax under neutral and weakly alkaline conditions. During the remainder of the incubation period, the generation of alkaline root exudates increased the solution pH.

pH in Hoagland’s nutrient solution after planting Arundo donax L
Effects of Hg and Cd on the physiological properties of A. donax
Antioxidant reactions are an important means of detoxification for plants under HM stress. To maintain normal physiological activities, plants generate a series of reactive oxygen species (ROS) in response to HM stress, such as hydrogen peroxide (H2O2), hydroxyl radicals (OH·), and superoxide anions (O2−). However, ROS can also lead to lipid peroxidation of cell membranes, damage to individual organelles, and plant death (Xun et al. 2017). MDA, SOD, and GSH are frequently used as indicators of antioxidant levels in plants under HM stress. As shown in Table 2, compared with the control, both single and combined Hg and Cd stress significantly increased A. donax leaf MDA content. Furthermore, the MDA content increased as the concentrations of coexisting Hg and Cd increased, in agreement with previous studies (Jiang et al. 2022; Mao et al. 2022). Single Hg stress resulted in slightly higher MDA content than single Cd stress, indicating that the toxicity of Hg in A. donax is higher than that of Cd. MDA is the final product of lipid peroxidation in plants, and its content usually reflects the degree of oxidative stress and the level of tolerance under adverse conditions. Our results indicate that combined Hg-Cd stress intensifies membrane lipid peroxidation in A. donax.
Leaf SOD activity was significantly higher in A. donax under either single or combined Hg and Cd stress than in the control treatment. Under single Hg or Cd stress, SOD activity increased with increasing Hg or Cd concentration; by contrast, SOD activity decreased with increasing Hg and Cd concentrations in the combination treatments. SOD is an important antioxidant enzyme that can eliminate ROS to prevent overoxidation of membrane lipids. In general, increases in HM stress induce elevated SOD content to eliminate ROS and alleviate plant damage (Kou et al. 2022). Compared with the control or single Hg or Cd stress, coexisting Hg and Cd stress aggravated the damage to the antioxidant system of A. donax and decreased SOD activity, consistent with the results of Xun et al. [21].
Compared with the control, all treatments increased GSH content. Furthermore, either single or combined Hg stress induced higher GSH content than Cd stress. These results indicate that GSH and SOD play similar roles in resisting oxidative damage. GSH is the main non-protein mercaptan in plants and is used as an indicator of plant antioxidant capacity (Abid et al. 2019). GSH can effectively eliminate ROS, lipid peroxides (LOOHs), and other harmful molecules to maintain normal cell growth and development. GSH is also a precursor of PCs, which chelate HMs to form complexes for transport to the vacuole for storage (Dago et al. 2014).
Bioaccumulation and translocation abilities of A. donax
The BAF indicates the ability of the plant to accumulate HMs from the environment. As shown in Table 3, the BAFHg of the roots and aerial parts of A. donax ranged from 16.66 ± 2.71 to 37.18 ± 2.90 and from 9.24 ± 1.22 to 18.56 ± 0.79, respectively. The BAFCd of the roots and aerial parts of A. donax ranged from 5.69 ± 0.40 to 13.60 ± 4.44 and from 1.17 ± 0.05 to 3.93 ± 0.40, respectively. The BAFHg and BAFCd were both far greater than 1, suggesting that A. donax efficiently accumulates Hg and Cd, primarily in the roots (Chen and Yang 2012). The BAFHg decreased as the Hg concentration increased. The BAFCd increased with the concentration of Cd under single stress but decreased with increasing Cd concentration under combined stress. Similarly, Xun et al. (Xun et al. 2017) reported that the BAFHg of Cyrtomium macrophyllum decreased as the soil concentration of Hg increased. In addition, the BAFHg of A. donax was higher than the BAFCd (Table 3), further indicating that the ability of A. donax to accumulate Hg is greater than its ability to accumulate Cd.
The TF indicates the efficiency of translocation of HMs from the roots to the aerial part of the plant (Raj et al. 2020a, b). As shown in Table 3, the TFHg and TFCd of A. donax were less than 1 in all treatments; furthermore, the TFHg was higher than the TFCd, in accordance with the results of Cano-Ruiz et al. (2020). The TFHg and TFCd values suggest that phytostabilization rather than phytoextraction should be considered one of the main phytoremediation pathways of A. donax. Given the formation of volatile Hg, A. donax appears to accumulate and translocate Hg from the roots to the aerial parts and transform Hg2+ into volatile Hg. This process might reduce the toxicity of Hg in the plant and result in a high TF value. In fact, the TFHg was highest in the Hg2Cd1 treatment, whereas the TFCd was highest in the Cd1 treatment, further confirming previous reports that Cd is less toxic to plants than Hg (Cristaldi et al. 2020).
Conclusion
In this study, single Hg or Cd stress had no obvious effect on A. donax growth, whereas Hg–Cd combined stress significantly inhibited A. donax growth. The acidic root exudates released from A. donax at the beginning of the incubation period quickly decreased the solution pH, which facilitated the accumulation of Hg and Cd in A. donax. A. donax exhibited high efficiency in accumulating and translocating Hg and Cd, and the accumulation of Hg and Cd was higher in the roots than in the aerial parts. Furthermore, Hg was accumulated and translocated by A. donax more readily than Cd. Even in the absence of illumination, Hg2+ was transformed into volatile Hg by A. donax. Cd exhibited a “low promotion and high inhibition” effect on the accumulation, translocation, and transformation of Hg by A. donax. The increases in leaf SOD activity and GSH content indicated high resistance of A. donax to both single and combined Hg and Cd stress. Based on the TF values and the formation of volatile Hg, phytostabilization and phytovolatilization should be considered the main phytoremediation pathways for the removal of Hg by A. donax. In summary, A. donax is a potential candidate for the phytoremediation of combined Hg–Cd pollution even under neutral conditions because of its high accumulation ability and high tolerance to Hg and Cd.
Data availability
The datasets used or analyzed during the current study are available from the corresponding author on reasonable request.
References
Abid R, Manzoor M, De Oliveira LM, da Silva E, Rathinasabapathi B, Rensing C, Mahmood S, Liu X, Ma LQ (2019) Interactive effects of As, Cd and Zn on their uptake and oxidative stress in As-hyperaccumulator Pteris vittata. Environ Pollut 248:756–762
Barone G, Storelli A, Meleleo D, Dambrosio A, Garofalo R, Busco A, Storelli MM (2021) Levels of mercury, methylmercury and selenium in fish: insights into children food safety. Toxics 9(2):39
Cano-Ruiz J, Galea MR, Amoros MC, Alonso J, Mauri PV, Lobo MC (2020) Assessing Arundo donax L. in vitro-tolerance for phytoremediation purposes. Chemosphere 252:126576
Cano-Ruiz J, Plaza A, Pinilla P, Mauri PV, Lobo MC (2021) Valorization of treated sewage sludge for Arundo donax production in a field experiment. Ind Crop Prod 170:113702
Cargnelutti D, Tabaldi LA, Spanevello RM, Jucoski GdO, Battisti V, Redin M, Blanco Linares CE, Dressler VL, de Moraes Flores EM, Nicoloso FT, Morsch VM, Chitolina Schetinger MR (2006) Mercury toxicity induces oxidative stress in growing cucumber seedlings. Chemosphere 65:999–1006
Chang H (2019) Spatial distribution and source identification of cadmium in typical polluted areas of Guizhou province: a case study of Wanshan mercury mine. Dissertation, China University of Geosciences (Beijing) (in Chinese)
Chen J, Yang ZM (2012) Mercury toxicity, molecular response and tolerance in higher plants. Biometals 25:847–857
Cristaldi A, Conti GO, Cosentino SL, Mauromicale G, Copat C, Grasso A, Zuccarello P, Fiore M, Restuccia C, Ferrante M (2020) Phytoremediation potential of Arundo donax (Giant Reed) in contaminated soil by heavy metals. Environ Res 185:109427
Dago A, Gonzalez I, Arino C, Martinez-Coronado A, Higueras P, Manuel Diaz-Cruz J, Esteban M (2014) Evaluation of mercury sress in plants from the Almaden mining district by analysis of phytochelatins and their Hg complexes. Environ Sci Technol 48:6256–6263
Du J, Zeng J, Ming X, He Q, Tao Q, Jiang M, Gao S, Li X, Lei T, Pan Y, Chen Q, Liu S, Yu X (2020) The presence of zinc reduced cadmium uptake and translocation in Cosmos bipinnatus seedlings under cadmium/zinc combined stress. Plant Physio Bioch 151:223–232
Guo JM, Yang JX, Yang J, Chen TB, Li HE, Xu TB, Zhou XY, Ye Y, Yu B (2019) Interaction of Cd and Zn affecting the root morphology and accumulation of heavy metals in Sedum aizoon. Environ Sci 40(1):470–479 (in Chinese)
Haque S, Zeyaullah M, Nabi G, Srivastava PS, Ali A (2010) Transgenic tobacco plant expressing environmental E. coli merA gene for enhanced volatilization of ionic mercury. J Microbiol Biotechn 20:917–924
He JY, Ren YF, Yan YP, Zhu C, Jiang DA (2010) Impacts of cadmium stress on the growth of rice seeding and division of their root tip cells. Acta Pedol Sin 47(1):138–144 (in Chinese)
Heaton ACP, Rugh CL, Wang NJ, Meagher RB (2005) Physiological responses of transgenic merA-tobacco (Nicotiana tabacum) to foliar and root mercury exposure. Water Air Soil Pollut 161:137–155
Jiang N, Li ZA, Yang JM, Zu YQ (2022) Responses of antioxidant enzymes and key resistant substances in perennial ryegrass (Lolium perenne L.) to cadmium and arsenic stresses. BMC Plant Biol 22:145
Khuzhaev VU, Aripova SF, Shakirov RS (1994) Dynamics of Arundo-donax alkaloids accumulation. Chem Nat Compd 5:637–638
Kou M, Xiong J, Li M, Wang M, Tan W (2022) Interactive effects of Cd and Pb on the photosynthesis efficiency and antioxidant defense system of Capsicum annuum L. B Environ Contam Tox 108:917–925
Kutrowska A, Malecka A, Piechalak A, Masiakowski W, Hanc A, Baralkiewicz D, Andrzejewska B, Zbierska J, Tomaszewska B (2017) Effects of binary metal combinations on zinc, copper, cadmium and lead uptake and distribution in Brassica juncea. J Trace Elem Med Bio 44:32–39
Li P, Feng X, Shang L, Qiu G, Meng B, Liang P, Zhang H (2008) Mercury pollution from artisanal mercury mining in Tongren, Guizhou, China. Appl Geochem 23:2055–2064
Liu YN, Xiao XY, Guo ZH (2019) Identification of indicators of giant reed (Arundo donax L.) ecotypes for phytoremediation of metal-contaminated soil in a non-ferrous mining and smelting area in southern China. Ecol Indic 101:249–260
Liu ZC, Chen BN, Wang LA, Urbanovich O, Nagorskaya L, Li X, Tang L (2020) A review on phytoremediation of mercury contaminated soils. J Hazard Mater 400:123138
Lominchar MA, Sierra MJ, Jimenez-Moreno M, Guirado M, Martin-Doimeadios RCR, Millan R (2019) Mercury species accumulation and distribution in Typha domingensis under real field conditions (Almaden, Spain). Environ Sci Pollut Res 26:3138–3144
Ma X, Liu RH, Wei L, Wang Y, Liu SX (2015) Effect of ultraviolet on the photoproduction of dissolved gaseous mercury in coastal seawater during winter. China Environ Sci 35(11):3462–3467 (in Chinese)
Mao YQ, Liu WG, Yang XD, Chang YB, Yang T, Wang XY, Chen YG (2022) Syntrichia caninervis adapt to mercury stress by altering submicrostructure and physiological properties in the Gurbantunggut Desert. Sci Rep-UK 12:11717
Mwamba TM, Ali S, Ali B, Lwalaba JL, Liu H, Farooq MA, Shou J, Zhou W (2016) Interactive effects of cadmium and copper on metal accumulation, oxidative stress, and mineral composition in Brassica napus. Int J Environ Sci Te 13:2163–2174
Ozturk F, Duman F, Leblebici Z, Temizgul R (2010) Arsenic accumulation and biological responses of watercress (Nasturtium officinale R. Br.) exposed to arsenite. Environ Exp Bot 69:167–174
Pandey VC, Bajpai O, Singh N (2016) Energy crops in sustainable phytoremediation. Renew Sust Energ Rev 54:58–73
Pu G, Zeng D, Mo L, Liao J, Xu G, Huang Y, Zhang C (2019) Cadmium accumulation and its effects on physiological characteristics of Arundo donax L. in a simulated wetland. Global Nest J 21:423–429
Raj D, Maiti SK (2020) Sources, bioaccumulation, health risks and remediation of potentially toxic metal(loid)s (As, Cd, Cr, Pb and Hg): an epitomised review. Environ Monit Assess 192(2):108
Raj D, Kumar A, Maiti SK (2020a) Brassica juncea (L.) Czern. (Indian mustard): a putative plant species to facilitate the phytoremediation of mercury contaminated soils. Int J Phytoremediat 22:733–744
Raj D, Kumar A, Maiti SK (2020b) Mercury remediation potential of Brassica juncea (L.) Czern. for clean-up of flyash contaminated sites. Chemosphere 248:125857
Ren JH, Sun HH, Wang SF, Luo J, Ma LNQ (2014) Interactive effects of mercury and arsenic on their uptake, speciation and toxicity in rice seedling. Chemosphere 117:737–744
Saifullah SN, Bibi S, Ahmad M, Ok YS (2014) Effectiveness of zinc application to minimize cadmium toxicity and accumulation in wheat (Triticum aestivum L.). Environ Earth Sci 71:1663–1672
Shah K, Nahakpam S (2012) Heat exposure alters the expression of SOD, POD, APX and CAT isozymes and mitigates low cadmium toxicity in seedlings of sensitive and tolerant rice cultivars. Plant Physiol Biochem 57:106–113
Shi SJ, Richardson AE, O’Callaghan M, DeAngelis KM, Jones EE, Stewart A, Firestone MK, Condron LM (2011) Effects of selected root exudate components on soil bacterial communities. Fems Microbiol Ecol 77:600–610
Sun JH, Yang L, Yang XT, Wei J, Li LT, Guo EH, Kong YH (2021) Using spectral reflectance to estimate the leaf chlorophyll content of maize inoculated with Arbuscular mycorrhizal fungi under water stress. Front Plant Sci 12:646173
Ubeynarayana N, Jeyakumar P, Bishop P, Pereira RC, Anderson CWN (2021) Effect of soil cadmium on root organic acid secretion by forage crops. Environ Pollut 268:115839
Wang B, Chen M, Ding L, Zhao Y, Man Y, Feng L, Li P, Zhang L, Feng X (2021) Fish, rice, and human hair mercury concentrations and health risks in typical Hg-contaminated areas and fish-rich areas, China. Environ Int 154:106561
Xun Y, Feng L, Li Y, Dong H (2017) Mercury accumulation plant Cyrtomium macrophyllum and its potential for phytoremediation of mercury polluted sites. Chemosphere 189:161–170
Yabanli M, Yozukmaz A, Sel F (2014) Heavy metal accumulation in the leaves, stem and root of the invasive submerged macrophyte myriophyllum spicatum L. (haloragaceae): an example of Kadin Creek (Mugla, Turkey). Braz Arch Biol Techn 57:434–440
Yin RS, Gu CH, Feng XB, Hurley JP, Krabbenhoft DP, Lepak RF, Zhu W, Zheng LR, Hu TD (2016) Distribution and geochemical speciation of soil mercury in Wanshan Hg mine: effects of cultivation. Geoderma 272:32–38
Yozukmaz A, Yabanli M, Sel F (2018) Heavy metal bioaccumulation in Enteromorpha intestinalis, (L.) Nees, a macrophytic algae: the example of Kadin Creek (Western Anatolia). Braz Arch Biol Techn 61:18160777
Zhang H, Feng X, Larssen T, Shang L, Vogt RD, Rothenberg SE, Li P, Zhang H, Lin Y (2010) Fractionation, distribution and transport of mercury in rivers and tributaries around Wanshan Hg mining district, Guizhou province, southwestern China: part 1-Total mercury. Appl Geochem 25:633–641
Zhang CJ, Sale PWG, Doronila AI, Clark GJ, Livesay C, Tang CX (2014) Australian native plant species Carpobrotus rossii (Haw.) Schwantes shows the potential of cadmium phytoremediation. Environ Sci Pollut Res 21:9843–9851
Zhang D, Jiang QW, Liang DY, Huang SX, Liao JX (2021) The potential application of giant reed (Arundo donax) in ecological remediation. Front Env Sci 9:652367
Zhou JW, Li Z, Liu MS, Yu HM, Wu LH, Huang F, Luo YM, Christie P (2020) Cadmium isotopic fractionation in the soil-plant system during repeated phytoextraction with a cadmium hyperaccumulating plant species. Environ Sci Technol 54:13598–13609
Zhu HX, Teng Y, Wang XM, Zhao L, Ren WJ, Luo YM, Christie P (2021) Changes in clover rhizosphere microbial community and diazotrophs in mercury-contaminated soils. Sci Total Environ 767:145473
Funding
This research was funded by the National Key Research and Development Program of China (2019YFC1803705) and the Major Projects of the National Natural Science Foundation of China (41991335).
Author information
Authors and Affiliations
Contributions
Xiuhua Li and Ling Zhao wrote the paper; Ying Teng, Yongming Luo, and Qiguo Zhao conceived and designed the experiments; Xiuhua Li and Ying Teng analyzed the data; Xiuhua Li performed the experiments and collected the data.
Corresponding author
Ethics declarations
Ethics approval
Not applicable.
Consent to participate
All the authors approved of participating in this publication.
Consent for publication
All authors approved of participating in this manuscript.
Competing interests
The authors declare no competing interests.
Additional information
Responsible Editor: Gangrong Shi
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Li, X., Zhao, L., Teng, Y. et al. Effects of cadmium on mercury accumulation and transformation by Arundo donax L.. Environ Sci Pollut Res 30, 62461–62469 (2023). https://doi.org/10.1007/s11356-023-26516-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-023-26516-1