
Abstract
This study evaluated the ameliorative potential of grape seed extract (GSE) against Ehrlich solid tumor (EST)–induced hepatic tissue alterations in mice. The control group was infused with physiological saline. The second group received GSE (50 mg/kg day by day orally) for 2 weeks. The third group was subcutaneously injected with 2.5 million of EST cells. The fourth group was injected with EST cells and treated with GSE extract simultaneously. The fifth group was injected with EST cells and kept for 2 weeks until the appearance of a solid tumor, then treated with GSE for 2 weeks. The phytochemical analysis of GSE revealed the presence of total phenols (17.442 mg GAE/g) and total flavonoid (6.687 mg CE/g) with antioxidant activity of 81.506 mg TE/g DPPH. The Ehrlich solid tumor significantly raised the activities of ALT, AST, and ALP; the level of alpha fetoprotein (AFP) in serum; and the protein expressions of hepatic proliferating cell nuclear antigen (PCNA) and tumor suppressor protein (P53), as well as induced DNA damage and pathological alterations in liver tissue. However, it significantly reduced serum albumin and total protein levels. In contrast, the co- or post-treatment of EST-bearing mice with GSE reduced the activities of ALT, AST, and ALP; the level AFP in serum; and hepatic P53 and PCNA protein expressions. In addition, it reduced EST-induced hepatic DNA damage and pathological alterations, while it increased serum albumin and total protein levels. This study suggested that GSE is a potent hepatoprotective agent and both co- and post-treatment of EST-bearing mice with GSE almost had the same effects.
Graphical abstract

Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Malignant tumor is the most dreadful disease found among individuals and the biggest single reason for death in humans. It is a class of illnesses characterized by the uncontrolled development of cells that become invasive Wang et al. (2018). Tumors are initiated due to DNA abnormalities of the affected cells leading to uncontrolled cell division and subsequently increases in tissue mass. Much chemotherapy activates apoptosis through chemical or physical damage of DNA (Knijnenburg et al. 2018). Most cancer therapies are chemotherapy and radiation that kill cancer cells by inducing apoptosis in any case and, consequently, influence the patient’s life Evan and Vousden (2001).
The malignant tumor of the breast is the predominant kind of tumor and thought to be the second reason for mortalities in ladies (Siegel et al. 2014). An Ehrlich solid tumor (EST) is an unconstrained murine mammary adenocarcinoma that reproduced the malignancy of the breast (Mishra et al. 2018; El-Masry et al. 2019). EST has been utilized as a transplantable tumor model for simple examination of the antineoplastic impacts of various chemical substances (Kabel et al. 2013). However, it can induce oxidative stress and hepatotoxicity (Aldubayan et al. 2019; El-Masry et al. 2020).
Numerous plants and their products that have significant antioxidant activities can be used as supportive treatments during cancer therapy (Alotaibi et al. 2021; Altwaijry et al. 2020, 2021). Grape seeds contain many nutrients and active ingredients such as proteins, sugars, minerals, salts, flavonoids, coumarins, and tannins (Kim et al. 2006). In addition, it is a rich source of polyphenolic substances, including proanthocyanidins (89%), dimmers (6.6%), trimers (5.0%), tetramers (2.9%), and oligomers (74.8%) (Meeran et al. 2009; Bagchi et al. 2014; Schelz et al. 2016). Also, they are rich in epicatechin, anthocyanins, catechins, and epicatechin-3-o-gallate. Thus, grape seed extract (GSE) has powerful antioxidant properties (Moosavi et al. 2016) empowering to be utilized for therapy of numerous sicknesses, for example, Alzheimer’s disease Fouad and Rizk (2019) and gonad cancer (Ozpinar et al. 2017), and used as an anti-mutagenic, antiviral (Kim et al. 2006), anti-inflammatory, and anti-cancerogenic agent, as well as reduces mitochondrial damage and inhibits cell apoptosis (Zhou and Raffoul 2012; Tousson et al. 2018). Furthermore, resveratrol, an important polyphenolic compound present in grapes, has been shown to ameliorate the adverse effects of stroke on patients (Fodor et al. 2018) and oxidative stress induced by zinc oxide nanoparticles in Oreochromis niloticus and fipronil in albino rats, respectively (Abdel-Daim et al. 2019; AlBasher et al. 2020). Also, it has renal and neuroprotective effects via its antioxidant and anti-inflammatory activities (Ibrahim et al. 2018; Rahman et al. 2020). Therefore, this study evaluated the ameliorative potential of grape seed extract against Ehrlich solid tumor–prompted hepatic tissue alterations in mice.
Materials and methods
Grape seed extract
Grape seed extract, commercially known as Gervital capsules, 150 mg, was obtained from the Arab company for pharmaceuticals and medicinal plants (MEPACO-MEDIFOOD), Anshas El Raml, Sharqeya, Egypt.
Quantification of the total phenolic and flavonoid contents in grape seed extract
The amount of the total phenolic compounds of grape seed extract (GSE) was quantified according to Saggu et al. (2014) using Folin–Ciocalteu. Briefly 500 μl of GSE was added to 250 μl of Folin–Ciocalteu reagent, mixed, and left for 5 min for oxidation then neutralized by the addition of 1.25 ml of 20% Na2CO3 for 40 min. The optical density was measured at 725 nm wavelength, and the amount of the total phenolic compounds was quantified by using the gallic acid calibration curve and expressed as micrograms of gallic acid equivalent (GAE) per gram of the extract.
The total flavonoid content was determined according to Oyouni et al. (2019). Briefly, 500 μl of GSE was added to 250 μl of 5% NaNO2 and mixed for 6 min, then 2.5 ml of a 10% AlCl3 was added and left for 7 min. Finally, 1.25 ml of 1 M NaOH was added and the mixture was centrifuged at 5000g for 10 min. The optical density of the supernatant was measured at 510 nm wavelength. The total flavonoid amount was expressed as micrograms of catechin equivalent (CE) per gram of the extract.
Determination of the antioxidant activities of grape seed extract
The antioxidant activities of GSE were determined using the DPPH free radical scavenging assay. The ultimate 1,1-diphenyl-2-picryl-hydrazyl (DPPH) concentration was 50 μM in a 3.0 mL final reaction volume, and the absorbance was measured after 60 min at wavelength 517 nm against methanol. The percent inhibition of the DPPH free radical was calculated following the equation described by Choi et al. (2002). The antioxidant activity was estimated by Trolox’s calibration curve and expressed as milligrams of Trolox equivalent (mg TE/g).
Determination of phenolic acid profile
The phenolic acid profile of GSE was determined according to Oyouni et al. (2018) as follows: 20 mL of 2 M NaOH was added to 1 g of the extract in a conical flask, and the flask was flushed with N2. The flask with its content was shacked at room temperature for 4 h. Six molar of HCl was used to adjust the pH to 2, then the mixture was centrifuged for 10 min at 5000 rpm and the supernatant was harvested. The phenolic compounds were extracted with 50 mL ethyl ether and ethyl acetate 1:1 twice, and the organic phase was evaporated at 45 °C and the remaining was diluted in 2 mL of methanol. HPLC analysis was performed according to Kontogianni et al. (2013) and Akela et al. (2018) using Agilent Technologies 1100 series liquid chromatograph equipped with an auto sampler and a diode-array detector.
Reagents
Kits utilized for the analysis of the activities of alanine aminotransferase (ALT) and aspartate aminotransferase (AST) in serum were bought from Biodiagnostic Company, Egypt. While those used for measuring the levels of serum albumin and total protein were bought from Diamond Company, Egypt. The serum alkaline phosphatase (ALP) activity was determined using the diagnostic kit from BioMérieux Co, France. Alpha fetoprotein (AFP) was quantified by the automated quantitative enzyme-linked fluorescent assay (ELFA) (Biomerieux, Marcy-L’Etoile, France).
Animals
One hundred adult female Swiss albino mice of 22–25 g body weight were obtained from the Egyptian Organization for Biological Products and Vaccines (Cairo, Egypt). The mice were allowed water and commercial standard diet ad libitum.
Induction of the Ehrlich solid tumor
Mice with Ehrlich ascites carcinoma (EAC) (Fig. 1a) were obtained from the Egyptian National Cancer Institute (NCI; Cairo University, Egypt). The solid tumor was initiated by injecting 2.5 × 106 cells/mouse subcutaneously in the left thigh of the lower limb of female mice (Fig. 1b). The tumor was kept up in the mice by week after week subcutaneous injection of 2.5 × 106 cells/mouse (Fig. 1c).

Chromatograms of mixed standard solution of gallic acid, catachine, chlorgenic acid, syringic acid, vanillic acid, caffeic acid, ferulic acid, protochatchuic acid, rosmarinic acid, quercetin, and kaempferol
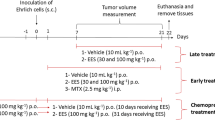
Experimental design
Following one week of acclimatization, 100 female mice were randomly allocated into 5 experimental groups, with 20 mice each.
Control group
Mice were subcutaneously infused with normal saline.
Grape seed extract (GSE) group
Mice received GSE (50 mg/kg body weight) orally day by day, for 2 weeks (Li et al. 2015).
Ehrlich solid tumor (EST) group
Mice were infused subcutaneously with 2.5 × 106 cells/mouse diluted in physiological saline Mansour and Anis (2010).
Ehrlich solid tumor and grape seed extract (EST and GSE) group
Mice were subcutaneously infused with 2.5 × 106 cells/mouse to initiate tumor and treated simultaneously with GSE (50 mg/kg body weight) orally day by day for 2 weeks.
Ehrlich solid tumor then grape seed (EST then GSE) group
Mice were subcutaneously infused with 2.5 × 106 cells/mouse to initiate tumor and left for 2 weeks until the appearance of the solid tumor then treated with GSE (50 mg/kg body weight) orally day by day for another 2 weeks.
Blood and tissue sampling
By the end of the experiment, the mice were anesthetized by intraperitoneal injection of sodium pentobarbital then scarified by decapitation. Blood samples were collected without anticoagulant, left to clot, and centrifuged to get sera that were kept at – 20 °C till used.
Livers were cut into two parts. One part was used for the comet assay while the other one was kept in 10% neutral-buffered formalin for histopathological and immunohistochemical examinations.
Biochemical assays
The activities of serum AST and ALT were analyzed calorimetrically as indicated by Reitman and Frankel (1957). The activity of serum ALP was analyzed following Belfield and Goldberg (1971). Total protein and albumin serum levels were estimated according to Doumas et al. (1977) and Bowers and Wong (1980), respectively. The serum AFP level was estimated according to Jang et al. (2016).
Comet assay
One gram of each liver sample was added to 1 mL of super-cold phosphate-buffered saline (PBS), mixed by stirring for 5 min, and then filtered. One hundred microliters of cell suspension was blended with 600 μL of low-melting agarose (0.8% in PBS) and 100 μL of this combination was spread on pre-coated slides. At that point, the slides were submerged in lysis buffer (0.045 M TBE, pH 8.4, containing 2.5% sodium dodecyl sulfate (SDS)) for 15 min and put in an electrophoresis chamber containing the TBE buffer without SDS and stained with ethidium bromide 20 μg/mL at 4 °C. The electrophoresis was carried out at 2 V/cm for 2 min and 100 mA. The migration patterns of DNA fragments of 100 cells from each group were assessed with a fluorescence microscope. The comet tail lengths were estimated from the center of the nucleus to the furthest limit of the tail with × 40 for checking and estimating the size of the comet. For visualization of DNA damage, the observations were made for GelRed-stained DNA by using a fluorescent microscope (× 40). Comet 5 image analysis software created by Kinetic Imaging, Ltd. (Liverpool, UK) connected to a CCD camera was utilized to survey the quantitative and subjective degree of DNA damage in cells by estimating the length of DNA movement and the percentage of moved DNA. Finally, the program determined the tail moment. Generally, 100 randomly selected cells were analyzed per sample according to Dhawan et al. (2009) and Abd Eldaim et al. (2019).
Histopathological investigation
Fixed liver samples were dehydrated and embedded in molten paraffin. Finally, samples were cut into 7-μm-thick sections, mounted on clean slides, and processed for hematoxylin and eosin staining Bancroft and Cook (1994). The severity of the pathological lesions was evaluated by modified semiquantitative scoring system (− indicated no alterations, + indicated mild alterations, ++ indicated moderate alterations, and +++ indicated severe alterations) (Chen et al. 2008).
Immunohistochemical examination
Protein expression of PCNA and P53 in the liver samples was detected in the deparaffinized sections by avidin–biotin–peroxidase immunohistochemical technique (Elite–ABC, Vector Laboratories, CA, USA) using PCNA monoclonal antibody (dilution 1:100; DAKO Japan Co., Tokyo, Japan) according to Tousson et al. (2011) and P53 monoclonal antibody (dilution 1:200 DAKO Japan Co, Ltd, Tokyo, Japan) according to Van der Loos et al. (2010) and Abd Eldaim et al. (2019). All stained slides (PCNA and P53) were checked using an Olympus microscope, and photos were captured by a digital camera (Canon Power Shot A620). The expression of PCNA and P53 was expressed as the percentage of positive stained cells per total 1000 counted cells in about 10 high-power fields. Negative = 0%; positive cells, weak and mild less than 5%, for both P53 and PCNA; positive cells, moderate = 8% for P53 and 17% for PCNA; positive cells, strong = 18% for P53 and 30% for PCNA.
Statistical analysis
Data were expressed as mean ± standard error of the mean (SEM). Data were analyzed by one-way analysis of variance (ANOVA) followed by least significant difference (LSD) tests to assess the significance of differences among groups. Differences at p < 0.05 were considered statistically significant. The analysis was done by using the Statistical Package for the Social Sciences (SPSS software version 16).
Results
The total phenolic and flavonoid contents and the antioxidant activity of grape seed extract
The phytochemical analysis of GSE revealed the presence of total phenols at a concentration of 17.442 mg GAE/g and total flavonoid at a concentration of 6.687 mg CE/g with antioxidant activity of 81.506 mg TE/g DPPH (Table 1). Screening of the phenolic acids and flavenoids of GSE by HPLC revealed the presence of gallic acid, protochatchuic acid, catachine, chlorgenic acid, caffeic acid, syringic acid, vanillic acid, ferulic acid, rosmarinic acid, quercetin, and kaempferol (Fig. 1 and Table 2)
Grape seed extract reduced EST-induced mortality in mice
Ehrlich solid tumor led to 20% mortality in EST-bearing mice. However, treatment of mice bearing EST with GSE simultaneously or after the development of EST decreased the mortality rate to 10% (Table 3). Conversely, GSE-treated mice were healthy and no clinical signs of toxicity and no mortality were detected throughout the experimental period.
Grape seed extract decreased the EST development in mice
Figure 2 reveals that treatment of EST-bearing mice (Fig. 2(D)) with GSE either co- or post-treatment decreased the size of EST (Fig. 2(E and F)).

Induction of Ehrlich solid tumor. A: mouse bearing Ehrlich ascites carcinoma (EAC). B: female mouse injected with EAC in the left thigh of the lower limb. C: developed EST. D, E, and F: developed Ehrlich solid tumor (EST) of EST (D), EST and GSE (E), and EST then GSE (F) groups
Grape seed extract modulated EST alterations in liver function biomarkers
Table 4 demonstrates the impacts of EST and/or GSE on liver function biomarkers. Ehrlich solid tumor induced a significant (p < 0.05) increase in the activities of serum ALT, AST, and ALP and the level of serum AFP. However, it significantly reduced total protein and albumin serum levels compared to the normal control mice. In contrast, administration of GSE either simultaneously with inoculation of EAC or after the appearance of EST reduced the increase in the activities of serum AST, ALT, and ALP, and the level of AFP, while it diminished the EST-prompted decrease in serum total protein compared to mice with EST (Table 4). The co- and post-treatment of EST-bearing mice with GSE almost had the same effects on the liver function biomarkers. Treatments of mice with GSE did not cause significant changes in the activities and levels of liver function biomarkers (Table 4).
Grape seed extract ameliorated EST-initiated liver DNA damage
Table 5 and Fig. 3 illustrated the effects of EST and/or GSE on DNA of hepatic tissue. Ehrlich solid tumor significantly elevated hepatic tissue DNA damage (p < 0.05) indicated by expanded tail length, tail DNA%, and tail moment compared to the normal control mice. However, administration of GSE either simultaneously with the inoculation of EST or after the appearance of EST with GSE modulated EST-prompted hepatic tissue DNA damage compared to EST-bearing mice. The co- and post-treatment of EST-bearing mice with GSE almost had the same effects on hepatic tissue DNA (Table 5 and Fig. 3).

Comet assay for assessment of hepatic tissue DNA damage: G1, control group; G2, GSE group; G3, EST group; G4, EST and GSE group; G5, EST then GSE group
Grape seeds extract protected mice hepatic tissue from EST-induced histopathological alterations
Figure 4 illustrates the histopathological alteration in the hepatic tissues of all groups. Hepatic tissue sections of the control and GSE groups revealed normal hepatic architecture (Fig. 4a, b). However, sections of hepatic tissues of EST-bearing mice showed severely injured hepatocytes, focal necrosis, and decreased Kupffer cells, with cellular anaplasia, pleomorphism, and anisocytosis (Fig. 4c, d). In contrast, hepatic tissue sections of EST-bearing mice co-treated with GSE showed mild injured hepatocytes with moderate cellular infiltration (Fig. 4e). Liver sections of post-treated EST with GSE showed moderate vacuolated hepatocytes (Fig. 4f).

Photomicrographs of mice liver sections of all groups stained with H & E. Hepatic tissue sections of the control and GSE groups (a and b) showed normal hepatic architecture with normal central veins (CV). Liver section of mice bearing EST (c and d) showed severely injured hepatocytes, focal necrosis (arrows), and cellular anaplasia, pleomorphism, and anisocytosis (arrow heads). Hepatic tissue sections of GSE and EST groups (e) showed mild injured hepatocytes with moderate cellular infiltration (arrows). Liver sections of the EST then GSE group (f) showed moderate vacuolated hepatocytes
Grape seed extract reduced EST-induced PCNA protein expression in hepatic tissue
PCNA protein expression in the liver sections of all experimental groups is presented in Fig. 5. Liver sections of the control and GSE-treated mice showed negative or mild expression for PCNA protein in hepatocyte nuclei (Fig. 5a, b). Conversely, liver sections of the EST group showed strong PCNA protein expression (Fig. 5c, d). PCNA expression in the liver sections was significantly increased in EST-bearing mice compared to the control group (Fig. 5g). Liver sections of the co-treated group demonstrated moderate expression for PCNA protein and mild reactions of the post-treated group (Fig. 5e, f).

Photomicrographs of hepatic tissue sections of all groups stained with PCNA-ir. Hepatocyte nuclei of the control and GSE groups showed faint reactions for PCNA-ir (arrows) (a and b). Hepatic tissues of the EST group showed strong protein expression (arrows) for PCNA (c and d). Hepatic tissues of the EST and GSE groups revealed moderate protein expression (arrows) for PCNA (e). Hepatic tissue sections of the EST then GSE group showed mild protein expression (arrows) for PCNA (f)
Grape seed extract reduced EST-increased P53 protein expression in hepatic tissues
The expression of P53 protein in the hepatic tissue sections of all experimental groups is shown in Fig. 6. Hepatic tissue sections of the control and GSE-treated mice showed negative or mild P53 protein expression (Fig. 6a, b). Liver sections of the EST group showed strong P53 protein expression (Fig. 6c, d).The protein expression of P53 in the hepatic tissues of the EST group was significantly elevated compared to that of the control group (Fig. 6g). Liver sections of the EST and GSE groups showed moderate expression for P53 protein and mild expression in the EST then GSE group (Fig. 6e–g).

Photomicrographs of hepatic tissue sections of all groups stained with P53-ir. Faint P53-ir reactions (arrows) were detected in the hepatic tissue sections of the control and GSE groups (a and b). Strong protein expression (arrows) for P53 was detected in the hepatic tissue sections of the EST group (c and d). Hepatic tissue sections of the EST and GSE groups had moderate protein expression for P53 (arrows) (e). Hepatic tissue sections of the EST then GSE group showed mild protein expression for P53 (arrows) (f)
Discussion
Ehrlich carcinoma is comparable to human cancer as it is undifferentiated, has a quick development rate, and is more delicate to chemotherapies (Kabel et al. 2013). The function of vital organs of animals can be inversely influenced by the development of a tumor. The finding of this study indicated that Ehrlich solid tumor induced hepatic dysfunction represented by an increase of serum ALT, AST, and ALP activities and a decrease of serum total protein and albumin levels. Our findings agree with that of Gupta et al. (2004) and Sakr et al. (2011) who found that EST increases the activities of AST, ALT, and ALP in serum. In addition, Abd Eldaim et al. (2021) indicated that Ehrlich ascites carcinoma (EAC) raises the activities of serum ALT, AST, and ALP in mice and Salem et al. (2011) showed a reduction in serum levels of total protein and albumin of mice bearing EAC. Alpha fetoprotein is one of the tumor markers that can occur principally in patients with liver malignant tumors, hepatocellular carcinoma, or benign liver diseases Lazarevich (2000). Our results were matched with those of Aldubayan et al. (2019) and Abd Eldaim et al. (2021) who indicated a significant increase of serum AFP levels in mice bearing EAC. The EST-induced alteration of liver function biomarkers might be attributed to the hypoxia in liver periventricular areas Biscoveanu and Hasinski (2000), which altered hepatic tissue structure and function as the histopathological investigations of the hepatic tissues of EST-bearing mice revealed hepatotoxicity that was manifested by the alteration of hepatic tissue architectures. These findings were in line with those of Chakraborty et al. (2007) and Tousson et al. (2020) who demonstrated the infiltration of Ehrlich tumor cells in the internal organs, which may result from proliferation and migration of tumor cells that leads to aggregations of inflammatory cells and mitochondrial destruction. The possible reason for EST-induced alterations in hepatic tissue structure and function is the induction of oxidative stress in the livers of EST-bearing mice as it has been reported that EST can induce oxidative stress and hepatotoxicity (Aldubayan et al. 2019; El-Masry et al. 2020).
Tumor suppressor protein (P53) is a transcription factor implicated in the regulation of the cell cycle, expression of genes, aging, apoptosis, and suppression of cancer (Haris 1996; DeVita et al. 2011). It stops the cell cycle at G1 and G2 in case of DNA damage, allowing the activation of DNA-repairing proteins. In case of unrepaired damage, the Bax gene is activated leading to apoptosis Haris (1996). PCNA proteins are the standard markers for the proliferation of cells; thus, they can be used to assess the growth of the cell population. PCNA proteins have been recognized as significant members in DNA replication during ordinary and neoplastic cell division (Juríková et al. 2016). It constructs a ring around the DNA to encourage and control its replication Stoimenov and Helleday (2009). Immunohistochemical examination showed a strong expression of apoptotic proteins, P53, and proliferation marker, PCNA, in the livers of mice bearing EST indicating that EST tumors have a high proliferation rate. These findings were parallel with those of Tousson et al. (2020) who found that EAC reduces the liver antioxidant system while it induces P53 protein expression in the hepatic tissue of mice. Furthermore, Nakopoulou et al. (1995) reported strong P53 and PCNA protein expression in hepatocellular carcinoma.
Concerning the hepatoprotective impacts of GSE against EST-prompted liver injury, the results of this study revealed that the treatment of mice bearing EST with GSE modulates EST-altered liver function. These results agreed with those of El-Sayed et al. (2014) who reported that proanthocyanidins, one of the active compounds of grape seed extract, protect the liver against acetaminophen-induced hepatic damage. In addition, Kandemir et al. (2012) stated that grape seed extract has potent antioxidant activities that protect rabbits’ livers against cisplatin-induced toxicity. Further, Shin and Moon (2010) found that grape seeds and skin inhibit dimethylnitrosamine-induced hepatic injury in rats and subsequently inhibit the increment of the activities of serum AST and ALT and AFP serum level. Tsai et al. (1994) stated that the level of serum AFP has a direct relationship with the serum AST, ALT, and ALP activities in most of the liver diseases such as hepatitis C virus inducing hepatic cirrhosis (Fattovich et al. 1997) and acute and chronic hepatitis Yao et al. (2007). However, Yao (2003) indicated a sharp decrease in the serum level of AFP following treatment of patients suffering from chronic hepatitis B virus infection and cirrhosis with lamivudine. This improvement in liver function of mice bearing EST treated with GSE can be explained by the protective effect of GSE against EST-induced liver damage as the histopathological examination of hepatic tissue samples of GSE-treated groups showed an improvement in injured hepatocytes. These finding was parallel with that reported by Sun et al. (2004) who demonstrated that grape containing polyphenols prevents ethanol-induced liver injury. Also, administration of tamoxifen-intoxicated rats with grape seeds reduced liver toxicity and injury (El-Beshbishy et al. 2010). In addition, proanthocyanidin and vitamin E protect rat liver against doxorubicin-induced hepatotoxicity Al-Sowayan and Kishore (2012). The hepatoprotective potentials of GSE against EST-induced hepatic injury in our study may be attributed to phenolic and flavonoid compounds present in GSE and include gallic acid, protochatchuic acid, catachine, quercetin, and kaempferol (Tables 1 and 2) as the flavonoids and phenolic compounds have antioxidant properties (Moosavi et al. 2016). In addition, catachine, gallic acid, quercetin, and kaempferol have antioxidant and hepatoprotective activities against carbon tetrachloride- and thioacetamide-induced hepatotoxicity in rats (Liu et al. 2015; Qureshi et al. 2019; Mousa et al. 2019).
Finally, our findings indicated that treatment of mice bearing EST with GSE decreased P53 and PCNA protein expression in the hepatic tissues as it has been shown that GSE protects the human liver cells against cancer chemotherapeutic drug–induced cytotoxicity through modulation of the expression of regulatory genes of the cell cycle and apoptosis such as bcl-2, P53, and c-myc (Al-Rasheed et al. 2018). Further, GSE supplementation reduces the number of colonic cells expressing PCNA and extracellular signal–regulated kinases 1 and 2 (ERK1/2) signaling Yang et al. (2015). In addition, GSE decreases the proliferation of colorectal cancer cells, enhances caspase-3 activity, and subsequently increases apoptosis (Nomoto et al. 2004). Also, GSE decreases cell proliferation in the intestinal tract of APCmin/mice Velmurugan et al. (2010) and azoxymethane-induced Fischer 344 rats Johansson and Hansson (2013).
Conclusion
Ehrlich solid tumor altered the liver function and increased DNA damage, P53 and PCNA protein expression in hepatic tissue with marked histopathological alterations. However, treatment of mice bearing EST with GSE either simultaneously or after the development of EST ameliorated EST-altered liver functions and structure and P53 and PCNA protein expression via its antioxidant activities. Both co- and post-treatment of EST-bearing mice with GSE almost had the same effects. This study suggested that GSE is a potent hepatoprotective agent.
Data availability
All data used in this study are included in this published article.
References
Abd Eldaim MA, Tousson E, El Sayed IE, Awd WM (2019) Ameliorative effects of Saussurea lappa root aqueous extract against Ethephon-induced reproductive toxicity in male rats. Environ Toxicol 34(2):150–159
Abd Eldaim MA, Tousson E, El Sayed IE, Abd Elmaksoud AZ, Ahmed AS (2021) Ameliorative effects of 9-diaminoacridine derivative against Ehrlich ascites carcinoma–induced hepatorenal injury in mice. Environ Sci Pollut Res. https://doi.org/10.1007/s11356-020-11857-y
Abdel-Daim MM, Eissa IAM, Abdeen A, Abdel-Latif HMR, Ismail M, Dawood MAO, Hassan AM (2019) Lycopene and resveratrol ameliorate zinc oxide nanoparticles-induced oxidative stress in Nile tilapia, Oreochromis niloticus. Environ Toxicol Pharmacol 69:44–50
Akela MA, El Atrash AM, El Kilany MI, Tousson E (2018) Qualitative and quantitative characterization of biologically active compounds of Rosemary (Rosmarinus officinalis) leaf extract. Journal of Advanced Trends in Basic and Applied Science 2(1):59–64
AlBasher G, Abdel-Daim MM, Almeer R, Ibrahim KA, Hamza RZ, Bungau S, Aleya L (2020) Synergistic antioxidant effects of resveratrol and curcumin against fipronil-triggered oxidative damage in male albino rats. Environ Sci Pollut Res Int 27(6):6505–6514
Aldubayan MA, Elgharabawy RM, Ahmed AS, Tousson E (2019) Antineoplastic activity and curative role of avenanthramides against the growth of Ehrlich solid tumors in mice. Oxidative Med Cell Longev 5162687:12
Alotaibi B, Tousson E, El-Masry TA, Altwaijry N, Saleh A (2021) Ehrlich ascites carcinoma as model for studying the cardiac protective effects of curcumin nanoparticles against cardiac damage in female mice. Environ Toxicol 36(1):105–113
Al-Rasheed N, El-Masry T, Tousson E, Hassan H, Al-Ghadeer A (2018) Hepatic protective effect of grape seed proanthocyanidin extract against Gleevec-induced apoptosis, liver injury and Ki67 alterations in rats. Braz J Pharm Sci 54(2):e17391. https://doi.org/10.1590/s2175-97902018000217391
Al-Sowayan NS, Kishore U (2012) Prophylactic efficacy of a combination of proanthocyanidin and vitamin e on hepatotoxicity induced by doxorubicin in rats. Int Res J of Pharmaceuticals 2:161–169
Altwaijry N, El-Masry TA, Alotaibi B, Tousson E, Saleh A (2020) Therapeutic effects of rocket seeds (Eruca sativa L.) against testicular toxicity and oxidative stress caused by silver nanoparticles injection in rats. Environ Toxicol 35(9):952–960
Altwaijry N, El-Masry TA, Alotaibi BS, Tousson E, Alodhayani AA, El-Morshedy K, Elmaghed NA, Sayed AE, Saleh A (2021) Potential therapeutic effects of avenanthramide-C against lung toxicity caused by silver nanoparticles injection in rats. Pak J Pharm Sci 34(1):337–343
Bagchi D, Swaroop A, Preuss HG, Bagchin M (2014) Free radical scavenging, antioxidant and cancer chemoprevention by grape seed proanthocyanidin: an overview. Mutat Res 768:69–73
Bancroft JD, Cook HC (1994) Manual of histological techniques and their diagnostic application. Churchill Livingstone, Edinburgh, London, New York, Tokyo, pp 23–26
Belfield A, Goldberg DM (1971) Revised assay for serum phenyl phosphatase activity using 4-amino-antipyrine. Enzyme 12(5):561–573
Biscoveanu M, Hasinski S (2000) Abnormal results of liver function test in patients with Grave’s disease. Endocr Pract 6:367–369
Bowers LD, Wong ET (1980) Kinetic serum creatinine assays. II. A critical evaluation and review. Clin Chem 26:555–561
Chakraborty T, Bhuniya D, Chatterjee M, Rahaman M, Singha D, Chatterjee BN, Datta S, Rana A, Samanta K, Srivastawa S, Maitra SK, Chatterjee M (2007) Acanthus ilicifolius plant extract prevents DNA alteration in a transplantable Ehrlich ascites carcinoma-bearing murine model. World J Gastroenterol 13(48):6538–6548
Chen TM, Subeq YM, Lee RP, Chiou TW, Hsu BG (2008) Single dose intravenous thioacetamide administration as a model of acute liver damage in rats. Int J Exp Pathol 89:223–231
Choi CW, Kim SC, Hwang SS, Choi BK, Ahn HJ, Lee MY, Park SH, Kim SK (2002) Antioxidant activity and free radical scavenging capacity between Korean medicinal plants and flavonoids by assay-guided comparison. Plant Sci 163:1161–1168
DeVita VT, Jr TS, Lawrance SA Rosenberg (2011) Cancer: principles and practice of oncology, 9th Edition. Philadelphia, Lippincott Williams & Wilkins, a Wolters Kluwer Business.
Dhawan A, Bajpayee M, Parmar D (2009) Comet assay: a reliable tool for the assessment of DNA damage in different models. Cell Biol Toxicol 25:5–32
Doumas BT, Watson WA, Biggs HG (1977) Albumin standards and the measurement of serum albumin with bromocresol green. Clin Chim Acta 31:87–96
El-Beshbishy H, Mohamadin AM, Nagy AA, Abdel-Naim AB (2010) Amelioration of tamoxifen-induced liver injury in rats by grape seed extract, black seed extract and curcumin. Indian J Exp Biol 48(3):280–288
El-Masry TA, Al-Shaalan NH, Tousson E, Buabeid M, Alyousef AM (2019) The therapeutic and antineoplastic effects of vitamin B17 against the growth of solid-form Ehrlich tumours and the associated changes in oxidative stress, DNA damage, apoptosis and proliferation in mice. Pak J Pharm Sci 32(6):2801–2810
El-Masry T, Al-Shaalan N, Tousson E, Buabeid M, Al-Ghadeer A (2020) Potential therapy of vitamin B17 against Ehrlich solid tumor induced changes in Interferon gamma, Nuclear factor kappa B, DNA fragmentation, p53, Bcl2, survivin, VEGF and TNF-α expressions in mice. Pak J Pharm Sci 33(1):393–401
El-Sayed ME, Mansour AM, Nady M (2014) Protective effects of red wine polyphenols and grape-seed proanthocyanidin extract on acetaminophen-induced liver injury. Int J Pharm Sci Res 5(11):782–789
Evan G, Vousden K (2001) Proliferation, cell cycle and apoptosis in cancer. Nature 411:6835. https://doi.org/10.1038/2F35077213
Fattovich G, Giustina G, Degos F, Tremolada F, Diodati G, Almasio P, Nevens F, Solinas A, Mura D, Brouwer JT, Thomas H, Njapoum C, Casarin C, Bonetti P, Fuschi P, Basho J, Tocco A, Bhalla A, Galassini R, Noventa F, Schalm SW, Realdi G (1997) Morbidity and mortality in compensated cirrhosis type C: a retrospective follow-up study of 384 patients. Gastroenterology 112:463–472
Fodor K, Tit DM, Pasca B, Bustea C, Uivarosan D, Endres L, Iovan C, Abdel-Daim MM, Bungau S (2018) Long-term resveratrol supplementation as a secondary prophylaxis for stroke. Oxidative Med Cell Longev 18:4147320
Fouad GI, Rizk MZ (2019) Possible neuromodulating role of different grape (Vitis vinifera L.) derived polyphenols against Alzheimer’s dementia: treatment and mechanisms. Bull Natl Res Cent 43:108
Gupta M, Mazumder UK, Kumar RS, Sivakumar T, Vamsi M (2004) Antitumor activity and antioxidant status of Caesalpinia bonducella against Ehrlich ascites carcinoma in Swiss albino mice. J Pharmacol Sci 94:177–184
Haris CC (1996) Structure and function of the p53 tumor suppressor gene: clues for rational cancer therapeutic strategies. J Natl Cancer Inst 88(20):1442–1455
Ibrahim A, Al-Hizab FA, Abushouk AI, Abdel-Daim MM (2018) Nephroprotective effects of benzyl isothiocyanate and resveratrol against cisplatin-induced oxidative stress and inflammation. Front Pharmacol 9:1268
Jang ES, Jeong S, Kim JW, Choi YS, Leissner P, Brechot C (2016) Diagnostic performance of alpha-fetoprotein, protein induced by vitamin K absence, osteopontin, Dickkopf-1 and its combinations for hepatocellular carcinoma. PLoS One 11(3):e0151069
Johansson ME, Hansson GC (2013) Mucus and the goblet cell. Dig Dis 31:305–309
Juríková M, Danihel L, Polák S, Varga I (2016) Ki67, PCNA, and MCM proteins: markers of proliferation in the diagnosis of breast cancer. Acta Histochem 118(5):544–552
Kabel AM, Abdel-Rahman MN, El-Sisi A, Haleem MS, Ezzat NM, El Rashidy MA (2013) Effect of atorvastatin and methotrexate on solid Ehrlich tumor. Eur J Pharmacol 713(13):47–53
Kandemir F, Benzer E, Ozdemir N (2012) Protective antioxidant effects of grape seed extract in a cisplatin-induced hepatotoxicity model in rabbits. Rev Med Vet 163(11):539–545
Kim SY, Jeong SM, Park WP, Nam KC, Ahn DU, Lee SC (2006) Effect of heating conditions of grape seeds on the antioxidant activity of grape seed extracts. Food Chem 97:472–479
Knijnenburg TA, Wang L, Zimmermann MT, Chambwe N, Gao G et al (2018) Genomic and molecular landscape of dna damage repair deficiency across the cancer genome atlas. Cell Rep 23(1):239–254
Kontogianni VG, Tomic G, Nikolic I, Nerantzaki AA, Sayyad N, Stosic-Grujicic S, Stojanovic I, Gerothanassis IP, Tzakos AG (2013) Phytochemical profile of Rosmarinus officinalis and Salvia officinalis extracts and correlation to their antioxidant and antiproliferative activity. Food Chem 136:120–129
Lazarevich NL (2000) Biochem Mosc 65(1):117–133
Li SG, DingYS NQ, Xu SZ, Pang LJ, Ma RL, Jing MX, Feng GL, Liu JM, Guo SX (2015) Grape seed proanthocyanidin extract alleviates arsenic-induced oxidative reproductive toxicity in male mice. Biomed. Environ Sci 28:272–280
Liu J, Lu JF, Wen XY, Kan J, Jin CH (2015) Antioxidant and protective effect of inulin and catechin grafted inulin against CCl4-induced liver injury. Int J Biol Macromol 72:1479–1484
Mansour SZ, Anis LM (2010) Possible effect of 5, 6–dimethyle– 4 isothiocyanatethieno[ 2,3– d] pyrimidine and / or irradiation on Ehrlich carcinoma in mice. J Radical Res 3(2):599–618
Meeran SM, Vaid M, Punathil T, Katiyar SK (2009) Dietary grape seed proanthocyanidins inhibit 12-O-tetradecanoyl phorbol-13-acetate-caused skin tumor promotion in 7, 12-dimethylbenz[a]anthracene-initiated mouse skin, which is associated with the inhibition of inflammatory responses. Carcinogenesis 30:520–528
Mishra S, Tamta AK, Sarikhani M, Desingu PA, Kizkekra S, Pandit A, Kumar S, Khan S, Raghavan SC, Sundaresan NR (2018) Subcutaneous Ehrlich ascites carcinoma mice model for studying cancer-induced cardiomyopathy. Sci Rep 8:5599. https://doi.org/10.1038/s41598-018-23669-9
Moosavi F, Hosseini R, Saso L, Firuzi O (2016) Modulation of neurotrophic signaling pathways by polyphenols. Drug Des Devel Ther 10:23–42
Mousa AA, El-Gansh HAI, Eldaim MAA, Mohamed MAE, Morsi AH, El Sabagh HS (2019) Protective effect of Moringa oleifera leaves ethanolic extract against thioacetamide-induced hepatotoxicity in rats via modulation of cellular antioxidant, apoptotic and inflammatory markers. Environ Sci Pollut Res Int 26(31):32488–32504
Nakopoulou L, Janinis J, Giannopoulou I, Lazaris AC, Koureas A, Zacharoulis D (1995) Immunohistochemical expression of p53 protein and proliferating cell nuclear antigen in hepatocellular carcinoma. Pathol Res Pract 191(12):1208–1213
Nomoto H, Iigo M, Hamada H, Kojima S, Tsuda H (2004) Chemoprevention of colorectal cancer by grape seed proanthocyanidin is accompanied by a decrease in proliferation and increase in apoptosis. Nutr Cancer 49(1):81
Oyouni AA, Saggu S, Tousson E, Rehman H (2018) Immunosuppressant drug tacrolimus induced mitochondrial nephrotoxicity, modified PCNA and Bcl-2 expression attenuated by Ocimum basilicum L. in CD1 mice. Toxicol Rep 5:687–694
Oyouni AA, Saggu S, Tousson E, Mohan A, Farasani A (2019) Mitochondrial nephrotoxicity induced by tacrolimus (FK-506) and modulatory effects of Bacopa monnieri (Farafakh) of Tabuk Region. Pharm Res 11(1):20–24
Ozpinar H, Ozpinar N, Karakus S (2017) The effects of Erzincan grape (Vitis vinifera spp., Cimin) and Benzothiazol on a Caenorhabditis elegans organism model. Pharmacogn Mag 13:S380–S384
Qureshi SA, Jahan M, Lateef T, Ahmed D, Rais S, Azmi MB (2019) Presence of gallic acid and rutin improve the hepatoprotective strength of Withania coagulans. Pak J Pharm Sci 32:301–308
Rahman MH, Akter R, Bhattacharya T, Abdel-Daim MM, Alkahtani S, Arafah MW, Al-Johani NS, Alhoshani NM, Alkeraishan N, Alhenaky A, Abd-Elkader OH, El-Seedi HR, Kaushik D, Mittal V (2020) Resveratrol and neuroprotection: impact and its therapeutic potential in Alzheimer’s disease. Front Pharmacol 11:619024
Reitman S, Frankel S (1957) A colorimetric method for the determination of serum glutamic oxaloacetic and glutamic pyruvic transaminases. Am J Clin Pathol 28:56e63
Saggu S, Sakeran MI, Zidan N, Tousson E, Mohan A, Rehman H (2014) Ameliorating effect of chicory (Chichorium intybus L.) fruit extract against 4-tert-octylphenol induced liver injury and oxidative stress in male rats. Food Chem Toxicol 72:138–146
Sakr SA, Badr OM, Abd-Eltawab HM (2011) Ameliorative effect of saffron extract on mice bearing solid tumors. ISESCO J Science Technol 12(22):60–70
Salem FS, Badr MT, Neamat-Allah AE (2011) Biochemical and pathological studies on the effects of levamisole and chlorambucil on Ehrlich ascites carcinoma-bearing mice. Vet Ital 47(1):89–95
Schelz Z, Ocsovszki I, Bózsity N, Hohmann J, Zupkó I (2016) Antiproliferative effects of various furanoacridones isolated from Ruta graveolens on human breast cancer cell lines. Anticancer Res 36(6):2751–2758
Shin M, Moon J (2010) Effect of dietary supplementation of grape skin and seeds on liver fibrosis induced by dimethylnitrosamine in rats. Nutr Res Pract 4(5):369–374
Siegel R, DeSantis C, Jemal A (2014) Colorectal cancer statistics, 2014. CA Cancer J Clin 64(2):104–117
Stoimenov I, Helleday T (2009) PCNA on the crossroad of cancer. Biochem Soc Trans 37(3):605–613. https://doi.org/10.1042/BST0370605
Tousson E, Ali EM, Ibrahim W, Mansour MA (2011) Proliferating cell nuclear antigen as a molecular biomarker for spermatogenesis in PTU-induced hypothyroidism of rats. Reprod Sci 18(7):679–686
Tousson E, Elgharabawy RM, Elmasry TA (2018) Grape seed proanthocyanidin ameliorates cardiac toxicity induced by boldenone undecylenate through inhibition of NADPH oxidase and reduction in the expression of NOX2 and NOX4. Oxidative Med Cell Longev 9434385:12
Tousson E, Hafez E, Gazia MM, Salem SB, Mutar TF (2020) Hepatic ameliorative role of vitamin B17 against Ehrlich ascites carcinoma–induced liver toxicity. Environ Sci Pollut Res 27:9236–9246
TSAI JF, Chang WY, JENG J, HO M, Lin ZY, Tsai JH (1994) Hepatitis B and C virus infection as risk factors for liver cirrhosis and cirrhotic hepatocellular carcinoma: a case-control study. Liver14:98–102
Van der Loos CM, Meijer-Jorna LB, Broekmans ME, Ploegmakers HP, Teeling P, de Boer OJ, van der Wal ACJ (2010) Anti-human vascular endothelial growth factor (VEGF) antibody selection for immunohistochemical staining of proliferating blood vessels. J Histochem Cytochem 58:109–118
Velmurugan B, Singh RP, Kaul N, Agarwal R, Agarwal C (2010) Dietary feeding of grape seed extract prevents intestinal tumorigenesis in APCmin/ mice. Neoplasia 12:95–102
Wang X, Mo F, Bo H, Xiao L, Chen G, Zeng P, Huang YN, Lei Z, Yuan WJ, Chen ZH (2018) Upregulated expression of long non-coding RNA, LINC00460, suppresses proliferation of colorectal cancer. J Cancer 9:2834–2843. https://doi.org/10.7150/jca.26046
Yang G, Yansong X, Hanying Z, Du M, Zhu M-J (2015) Favourable effects of grape seed extract on intestinal epithelial differentiation and barrier function in IL10-deficient mice. British J Nutrition 114(01):1–9
Yao FY (2003) Dramatic reduction of the alpha-fetoprotein level after lamivudine treatment of patients with chronic hepatitis B virus infection and cirrhosis. J Clin Gastroenterol 36(5):440–442
Yao DF, Dong Z, Yao M (2007) Specific markers in hepatocellular carcinoma. Hepatobiliary Pancreat Dis Int 6:241–247
Zhou K, Raffoul JJ (2012) Potential anticancer properties of grape antioxidants. J Oncol:803294. https://doi.org/10.1155/2012/803294,8
Acknowledgements
The authors thank Taif University for the financial support of Taif University Researchers Supporting Project (TURSP-2020/09), Taif University, Taif, Saudi Arabia.
Funding
This study was supported by the Taif University Researchers Supporting Project (TURSP-2020/09), Taif University, Taif, Saudi Arabia.
Author information
Authors and Affiliations
Contributions
Abd Eldaim MA, El Sayed IE, and Tousson E proposed the idea and the experimental design; Elsharkawy HN and Soliman MM conducted the experiment and wrote the draft of the manuscript with the help of El Sayed IE and Abdel Aleem AH; Abd Eldaim MA, Tousson E, and El Sayed IE supervised the implementation of the experiment and analyzed the results; Tousson E and Elsharkawy HN performed the histopathological examination. All authors discussed the results and contributed to finalize the manuscript.
Corresponding author
Ethics declarations
Ethical approval
Rearing and treatment of mice and all experimental procedures were conducted according to the guide for animal use that was approved by the Institutional Animal Care and Use Committee (IACUC-SCI-TU-0041), Faculty of Science, Tanta University.
Consent to participate
Not applicable
Conflict of interest
The authors declare no competing interests.
Additional information
Responsible Editor: Mohamed M. Abdel-Daim
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Abd Eldaim, M.A., Tousson, E., Soliman, M.M. et al. Grape seed extract ameliorated Ehrlich solid tumor-induced hepatic tissue and DNA damage with reduction of PCNA and P53 protein expression in mice. Environ Sci Pollut Res 28, 44226–44238 (2021). https://doi.org/10.1007/s11356-021-13904-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-021-13904-8