
Abstract
Feeding 9 billion by 2050 is one of major challenges for researchers. Use of diversified crops, nonconventional water resources and rehabilitation of marginal lands are alternate options to produce more food to face climate change projections. Adaptation to climate change through climate smart agriculture practices, agroecology activities, and crop-based management packages can help transform the marginal lands from environmental burdens into productive and economic blocks. This review discusses the recent advancements on specialty group of alternate crops (oil seeds, legumes, cereals, medicinal, lignocellulose, and fruit crops) which can adapt in the marginal environments. Availability of alternate water resources (saline water, treated wastewater) for irrigation cannot be omitted. Crop diversification systems involving drought and salt-tolerant crops are likely to be the key to future agricultural and economic growth in the regions where salt-affected soils exist and/or saline aquifers are pumped for irrigation. These systems may tackle three main tasks: sustainable management of land resources and enhancement of per unit productivity; intensification of agroecological practices to increase soil fertility; and improving productivity of marginal lands for diversified climate smart crops. This review explores various aspects of marginal lands and selection of tolerant crop genotypes, crop diversification, and agroecological practices to maximize benefits.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
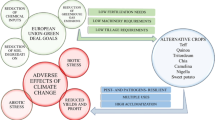
The global human being population will cross 9 billion by 2050, but the food production has not been significantly increased at the same rate (FAO 2011a, b). It is, therefore, imperative to enhance the food production by 44 million tons per year for the next 40 years (Tester and Langridge 2010) that equals to a 38% more than the historical trends in production. The world arable land is rapidly decreasing due to land degradation, urbanization, salinity, and drought and flooding. These entire factors contribute towards creation of challenge and causing hinders to achieve the UN development goals of sustainability. The climate change scenario is further exacerbated this challenge and make it an acute problem. The drought, as an important component of climate change scenario, should have to be tackled with all available conventional and non-conventional water resources (Setter and Waters 2003). Salinity is a big problem causing land degradation and ecosystem functioning and affecting agricultural productivity all over the world (Fig. 1; Flowers et al. 1997; Munns and Tester 2008; Hussain et al. 2015). This situation is worse in arid and semiarid countries and in coastal regions where a significant portion of the land has been affected due to salinity (Pitman and Läuchli 2002). The main reason behind the scene is that mostly these areas are receiving significantly less rainfall, and farmers are using more saline water in these degraded marginal lands to irrigate their crops (Malash et al. 2008). The land degradation situation is very bad in central Asian states because more than 50% land has already been converted into unfertile marginal land because of waterlogging and continuous use of low quality highly saline–sodic water for irrigation (Kijne 2005; Qadir et al. 2008).

a and b Water logging and salt crusts on the top soil limit crop production in arid, semi-arid regions, and salt-degraded marginal land of Umm Ul Queen, UAE
In this scenario, a significant association between different components of crop production chain (environment, agriculture and food production, agroecology) exists that all together form the natural environment. This means that to taking care of the quality of nature and natural resources are not only a civilization requirement but also a prerequisite for the operation of agricultural production and ultimately food security. Each agricultural activity requires firstly biophysical means such as suitable land, water, and climate and secondly socioeconomic conditions such as credit, infrastructure, inputs, and markets (DeClerk et al. 2012). Approximately 80% of African population live in the rural sector, and their state of food security depends directly on agricultural production or indirectly through providing for agricultural labor (Tomich et al. 1995), whereas food security is affected by a myriad of factors including poverty, incomes, and unemployment. Furthermore, environmental change will have significant impacts on household food security through extreme weather events that will have direct and indirect negative effects on household food security (IPCC 2001). The marginal lands can also contribute to food security and poverty reduction, but it depends upon several agroecosystem components such as maintain soil fertility through organic fertilizers, judicial, and safe use of nonconventional water resources (desalinated water, treated wastewater, rainwater, diseases, and pest control). Access to food is more than ever a question of interest. Recent increases in world grain prices have added to the claims that we are facing a global food crisis. Alarming population growth, natural resources degradation, unfavorable climatic conditions, and decrease in agricultural research have contributed to recent shortages in food crops and added weight to calls to increase the supply of agricultural commodities.
In this manuscript, recent publications and advances on climate resilient crops from major crops groups viz. oil seed crops (safflower, rapeseed/mustard, soybean, maize, desert gourd); food legumes (cowpea, faba bean, soybean, chickpea, sesbania, amaranth); nonlegume grain crops (barley, quinoa, amaranth, teff, sorghum, millet, triticale); medicinal crops (moringa, chia); lignocellulosic crops (perennial crops, grasses, sorghum, triticale, barley); and fruit crops (date palm, olive, phalsa, jambolan, guava Indian jujube, Indian gooseberry, karanda) have been considered. In the second part of the review with the need for pragmatic and empirical data, we focused on the agrobiodiversity perspectives of marginal lands and crop yield reduction following exposure to major abiotic constraints (drought, salinity) with the aim to improve our understanding on general topics such as crop agronomy (brief botanical description, food/nutritional value), adaptation strategies, and potential marginal areas for their cultivation. The specific objective of this review is to collect in a unique manuscript the last update on salt and drought tolerant crops and their adaptation strategies in marginal environments to provide detailed information that can contribute towards poverty alleviation, food security, and environmental sustainability in degraded areas.
Methodology
For this review article, we followed the PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) guidelines. The study mainly focused on literature navigation from 1964 to 2020. The literature synthesis involved arid and semiarid regions with a main focus on studies conducted in the West Asia and North Africa (WANA) region categorized among the “highly saline-degraded marginal lands” (Hussain et al. 2019; Rodríguez et al. 2020a, b; Lyu and Xu 2020). The articles highlighting relationship (correlations and causations) between salinity and drought impact and plant tolerance potential were selected.
A systematic literature search on salinity and drought as important growth limiting factors, and on diversification of crops and crop varieties in respect to their capacity of salinity and drought adaptation/tolerance thus suitable for cultivation in marginal environments, was done. Four databases viz. Google Scholar, Scopus, Web of Science and Centre for Agriculture and Bioscience International (CABI) were used. The selection of these data bases depends upon large articles collection and their widely availability in PRISMA systematic reviews. However, CABI database is more focused on plant biology, agriculture, and environmental science research.
We determined keywords addressing the following topics: (1) oil seed crops, (2) legume crops, (3) cereal grain crops, (4) medicinal crops, (5) lignocelluloses crops, (6) and fruit crops. We revised literature from the abovementioned data bases during 2018–2019 and used wildcards (*) to account for various word spellings. We identified 1037 articles and added 126 articles through the references section of the retrieved articles. We added 27 additional records from previous knowledge and from a recently published systematic review of the nutritional drivers of food selection (Hussain et al. 2019).
The second part of the review was focused on agrobiodiversity and on its usefulness in marginal areas targeting mainly North Africa and West Asia, farmers’ perception, and impact of biodiversity on crop yield and physiological attributes, especially plant growth, biochemical, and yield traits. Contingency tests were used to evaluate the effects (positive or negative) of salinity and drought on crops for finding a relationship between crop growth and yield threshold, under these abiotic stresses.
Results
Following the identification, screening and eligibility phases, we identified 425 out of 1190 articles that fit our selection criteria. From these, 125 duplicates were deleted. Meanwhile, a total of 300 full texts were reviewed in detail and assessed for inclusion in this systematic review.
The lack of consistent keywords used in research related to morphological characteristics and physiological aspects necessitated the broad selection of keywords utilized. Naturally, a broader search resulted in many hits; several of which were irrelevant leading to the large number of articles excluded. Examples of some of the article topics considered irrelevant for this systematic review included salinity, drought, and heavy metal–related ecosystem impact on growth, yield, and quality of date fruit flesh, GIS spatial assessments of land use for date cultivation, and any discussion of biochar production from different plant parts (leaves, stem, bark, flesh, and pits), and date-waste products.
Discussion
Food security, marginal lands, and agricultural productivity
Food is the main element necessary to get nutrition to maintain growth, health, and development and is the main component for economic development (Lisa et al. 2006). In a broad sense, food security exists when “all people, at all times, have physical, social, and economic access to sufficient, safe and nutritious food that meets their dietary needs and food preferences to live an active and healthy life” (FAO 2001). However, food insecurity occurs when crop production system in under stress and situation will become verse. Therefore, sufficient food is not available for everyone or it is not utilized in a proper way. In developing countries and in sub-Saharan Africa, the situation is at alarming stage because of rapid population growth, soil degradation, nonavailability of good quality irrigation water and irrigation system, and change in the climate.
The present world food grain production is not adequate to feed the huge population (FAO 2011a, b). A lot of population is suffering from food insecurity and malnutrition. Agriculture is the main way of getting food, and farmers are the main workers to cultivate the soil and produce food, oil seeds, and cash crops. In sub-Saharan Africa, nonadoption of new technologies has further lagged that of Asia. Efficient, profitable, and sustainable development of marginal lands and supporting smallholder farmers in the Africa and Asia has a great potential to contribute food security in these regions. This may be achieved by lowering the dependency on other regions and increasing self-sufficiency through breeding of salt and drought crop genotypes that have better potential to survive in marginal environment due the presence of resistant genes (Fig. 2).

Schematic flow chart showing the processes to develop new crop cultivars to meet the challenges of crop production for the next future. MAS, marker-assisted selection
Marginal environment: an untapped potential
Marginal lands are the “margins of cultivation” (Tang et al. 2010) and include the “poorest land which can be remuneratively operated under given price, cost, and other conditions” (Tang et al. 2010). To fulfill the projected demand in 2050, the food production must be increased equivalent to that of the Indian subcontinent (Phalan et al. 2014).
Different types of unproductive lands can be included in the category of marginal lands. However, nutrient-poor sandy soils, salt-degraded and poor soil properties, and lands with bas quality brackish water, unproductive soils that are not suitable for any kind of vegetation are also included in this list. The typical examples of marginal lands include contaminated lands (heavy metals, salts), degraded eroded soils, and soils used for industrial and municipal wastes dumping, unproductive fallow agricultural lands (Smith et al. 2013; Tilman et al. 2006; Nixon et al. 2001; Table 1). Agricultural systems are in a state of transition to meet evolving challenges in many regions of the world, increasing global population, and degrading and depleting natural resources. This entire situation is further complicated with climate change and requires innovative solution for proper management of rapidly increasing areas of marginal lands and saline water resources. Environments can be marginal either as far as the biophysical aspects are concerned or in terms of their capacity to sustain socioeconomic activities (or of course both). An extreme example of the former is the Empty Quarter of the Arabian Peninsula, where in the absence of water, there is practically no biotic activity (Al-Dakheel et al. 2015). An example of the latter is the “slash and burn” farming systems in tropical rainforests, which yield low returns to farmers, but when undertaken through traditional farming practices ends up with different areas at different stages of a natural regeneration process. These two examples may appear marginal from one lens but sustainable from another. For example, a barren desert of sand dunes supports flora and fauna that is sustainable in the sense of being adapted to low water availability, while a low productivity farming system in the tropical rainforest mimics the local ecosystem in terms of regenerating nutrient cycles, and supporting biodiversity. Marginal areas are therefore an integral component of the overall ecosystem of humans, plants, and animals interacting with land, water, and climate variables (Al-Dakheel and Hussain 2016). The continuum of natural resources from high potential regions to marginal zones is affected by the interaction among its components, which are constantly in a state of change. Some productive areas can be reduced into marginal resources because of poor management, such as the case of salinization of irrigated lands in Central Asia, Iraq, and Pakistan (Bianchi et al. 2006). Others are examples of marginal regions being transformed into production zones through smart investments (such as tile drainage in the Egyptian delta), drip irrigation (throughout the Middle East and North Africa), and treated urban wastewater reuse (UAE, Jordan, Tunisia), all of which involved the appropriate application of science and technology through substantial investments of public and private financial resources. These examples have expanded irrigation on large tracks of semiarid zones and desert regions in South Asia, North Africa, and Central Asia. The agroindustrial approach has focused on how to cost-effectively convert marginal environments to favorable ones through a combination of technology innovations and capital investments. Typically, better soil and water management techniques, together with innovations in technology applications, and innovative agronomic practices, have brought about this transformation. The agroecological approach by contrast explicitly incorporates natural resource management concerns in the analysis of marginal environments. It is in a better position to incorporate risks caused by climate change, unsustainable natural resource usage in assessing the sustainability of outcomes.
Food security, poverty, and marginalized farmers
Food insecurity prevails among the poor. To focus on food security, therefore, means to focus on poor people. The poverty is highest in Africa, followed by India and eastern Asia. Several thousands of resource poor farmers depend upon marginal lands and have scarce freshwater resources. The hunger is closely associated to the improvement and bringing the marginalized lands under cultivation because this is the big issue in developing world. However, farmers from these marginal lands are the main backbone of the world food security (Chappell and LaValle 2011).
Special attention to the high potential zones may have caused imbalanced resource allocation at the expense of regions with poorly endowed natural resources. The impact of these agroindustrial on marginal areas have been generally minimal—evidence points to resource poor farmers not having gained much from the green revolution. However, this sizable group of about 1.4 billion people lives and works in an underlying biophysical environment is often inherently fragile to support the economic needs of growing populations in a sustainable manner. Although they are huge in terms of number, their poverty and the small-scale nature of their activities mean that marginal farmers exert little market power. Moreover, their lack of mobilization (and the obstacles to this) means that their numbers rarely translate into political voice. The communities within which they reside are typically represented in parliament by urban dwellers and/or medium–large scale agricultural producers. Furthermore, as a group they have limited or no voice even within decentralized administrative systems. Women farmers (i.e., majority of marginal farmers in sub-Saharan Africa) may also be subject to various forms of social exclusion and political marginalization. Lack of influence in centers of power is a characteristic feature of marginal farmers and one that has led to years of policy neglect.
Role of agroecology in the crop diversification for sustainable development
The vulnerability of agroecosystems
Due to climate change cascades and geographical expansion, the monocultures have significantly increased through devotion of single crop to a piece of land and cultivation of the same crop over year-to-years. In this regard, rice, wheat, maize, and potatoes roughly accounted for 60% of the food grain source while animals provide 90% of all protein source (Vigouroux 2011). During the twentieth century, 60–70% of the total land area in USA was devoted to bean (2–3 varieties), area under potato was 72% with 3–4 varieties, and 2–3 varieties of cotton were planted. Due to increasing demand for food and fuel and climate change crises, the role played by agroecological practices and services provided by them (ecological and socioeconomic) are well recognized (Altieri 2004; De Schutter 2010). Several researchers documented that modern agriculture can be vulnerable to climate change (human or natural factor) and can led to drastic reduction in crop yield, globally, and in marginal environment, particularly.
Due to climatic perturbations, drought has drastically affected 26 states of USA causing significant reduction in crop yield on an area of 55% (1 billion hectares) of the total area. The severe crop losses due to destruction of heavy monsoon rains in 2011 flood in Pakistan caused a significant reduction of planted crops, trees, and ultimately destroying 2.4 million hectares and mortality to > 450,000 livestock that resulted in huge economic losses (2.9 billion dollars) (IPCC 2014).
Various agricultural practices such as adaptation of monoculture of biofuel crops are responsible for severe insect-pests outbreak because change in agrobiodiversity can lead to elimination of natural enemies of insects/pests. The monoculture ecosystem developed in the different states of USA such as biofuel crops has significant impact on landscape diversity that reduced (24%) biocontrol service (due to decrease in natural enemies supply to soybean field. According to reports of Landis et al. (2008), soybean production was decreased in the respective states, and producers had suffered an estimated cost of $58 million per year.
The ecological role of biodiversity in agroecosystems
In agroecosystem, the species diversity plays an important role to make different species less resilient against various degree and types of environmental shocks. Protecting the species against environmental fluctuations enhancing the capacity of different component of ecosystem (support to more than one species/component, in case one species will fail) might act as buffer against ecosystem failure (Cabell and Oelofse 2012).
Enhancing agrobiodiversity to reduce vulnerability
Different agroecological practices might help to maintain a healthy ecosystem which in turn will enhance the base of plant protection, health, productivity, yield stability, and soil health, in this regard, diversification at either or both species and genetic level. This include examples of polyculture, variety mixtures at different levels (landscape or field), e.g., agroforestry, integrated crop-livestock interaction, hedgerows, and corridors. The farmers might see these options suitable for best implementation of a successful strategy for a sustainable agroecosystem.
Adaptation of diversifying cropping systems may help, in marginal environment, to reduce the incidence of insect, pest, and diseases attack that will lead to low crop damage and higher yield (Altieri 2004). In a study conducted by Zhu et al. (2000), in China, evidenced that the farmers who planted four different varieties of rice (> 3000 ha), there were 44% less blast incidence, and 89% higher yield than other famers field where single variety of rice was cultivated. The Napier grass and leguminous silver leaf (Desmodium) planted between rows of maize demonstrated an excellent repelling crops for borers and ticks and to control Striga (a parasitic weed) as compared with maize monoculture. Furthermore, leguminous silver leaf can increase N-fixation soil fertility and crop yield (15–20%) (Khan et al. 2010). Biodiversity plays a positive role in stability of agroecosystem, and it will be vital under future climate change scenarios (Altieri 2004). Diversity in agroecosystem will also help to buffer beside high temperature, drought episodes, and rainfall. It may also affect the crop growth yield due to differential responses from different crop plants against the environmental perturbations (Altieri and Koohafkan 2013).
Crop diversification for marginal environment
Crop diversification represents an option for marginal lands by providing economic benefits to farmers and helping at the same time the environment conservation through the improvement of soil physical properties. Different drought-, salt-, and heavy metal–tolerant crops have been screened, selected, and developed at various agriculture research centers around the world for promoting the rehabilitation of marginal lands. These crops include abiotic stress tolerant genotypes of maize, safflower, quiona, pearl millet, sorghum, barley, perennial grasses, mustard, Sesbania, and triticale. These crops showed significant salt tolerance potential and yield stability under low to high salinity that indicates their adaptation to marginal environment (Table 2). These salt and drought crops have less water requirement but wider uses as food, feed, and industrial. These properties make them promising candidate for the diversification of production systems enhancing their economic value (Al-Dakheel et al. 2015; Hussain et al. 2015; Al-Dakheel and Hussain 2016). Meanwhile developing new crop genotypes that can produce higher yield with fewer inputs or that can increase yield stability and sustainable management of all the components of production system in marginal lands are of paramount importance (Fig. 3). These crop varieties should be highly tolerant to drought, salinity, high temperature, and strongly resistant to insects, pest, and pathogens (Cooper et al. 2014). Crop diversification might occur at field and landscape scale and should include several forms of innovative practices such as agroforestry, integrated crop–livestock interaction, legume-cereal intercropping, relay cropping, perennial forage crops, etc. This indicates a variety of option that farmers can adapt to cope with climate change scenario and to combat yield loss due to continuous monocropping.

Crop production system management components for sustainable development of nutrient poor marginal sandy desert soils; a Soil preparation and installation of irrigation pipes with drippers, b spreading of agrel sheet after seed sowing to protect the seed from birds eating, c profound germination and seedling growth of different crops in the field, d using appropriate net technology to protect the mature crop from birds attack
Nowadays agriculture production systems in the marginal lands need to adapt to the new climate and associated factors. Adaptation is an important stage that will help to cope with climate change severity and its impact on crop production. However, during the previous experimental work and projects conducted in West Asia and North Africa, it was observed that different types of adaptation strategies should have used. In this way, modifying different agronomic practices like changing sowing dates, introducing new stress tolerant crop genotypes, improving irrigation practices, and using non-traditional water resources will help to maintain a sustainable agroecosystem (Hussain and Al-Dakheel 2015). However, several attributes like agroecosystem diversification, integrated crop–livestock interaction, soil organic amendments, and water management will lead to enhance the general characteristics of agrobiodiversity with durable benefits.
Oil seed crops
Safflower
Safflower is an important oil seed crop mainly cultivated for highly nutritive oil (32–40%) has great genotypic and phenotypic plasticity in wide range of environments, as winter and summer crops. The seeds of different safflower varieties are enriched with vitamins (thiamine and β-carotene), essential nutrient elements, bioactive compounds, oil contents, 35-50%, and α, β, and γ tocopherols (Camas et al. 2007; Velasco et al. 2005; Khalid et al. 2017). Safflower seed oil possesses linoleic acid and tocopherol which are important polyunsaturated fatty acids (Han et al. 2009). These molecules showed several pharmacological and health perspectives. Khalid et al. (2017) showed several therapeutic properties of safflower seed oil such as atherosclerosis, skin disorders, bone-problems, and menopause.
Safflower can be cultivated in saline and marginal arid lands and is more successful than other oil seed crops (Kaya 2009a, b; Hussain et al., 2016). It salt tolerant and therefore is a promising oilseed crop for arid climate (Kar et al. 2007a, b; Hussain and Al-Dakheel 2018). Safflower can have great ability to withstand drought stress as it has very less water requirement compared with other oilseed crops. However, drought stress at reproductive phase can significantly reduce the seed yield of safflower (Table 3). For instance, drought stress at flowering and grain filling can reduce the yield by 35–50% (Istanbulluoglu 2009) and 50–62%, respectively (Istanbulluoglu et al. 2009). However, heavy irrigation rate reduced the yield by 7 and 15% in winter and summer planted safflower at early growth and flower initiation stages, respectively. Movahhedy-Dehnavy et al. (2009) reported that drought stress decreased the seed yield but also reduced the seed concentration as 12 and 49% decrease in oil concentration. However, drought stress at grain filling did not influence the oil concentration; rather, a small increase in oil concentration was observed when drought was imposed during grain filling stage (Table 5). Safflower genotypes and soil type also influence the crop performance under drought stressed condition. Santos et al. (2017) tested four safflower genotypes in sand and clay soil and found that IAPAR and IMA-4409 genotypes were relatively drought tolerant than other tested genotypes. Singh et al. (2016) demonstrated that safflower plants receiving 1–4 times less water produced 45.9, 33.6, 22.9, and 12.3% less yield than control.
In salinity tolerance evaluation trials with < 265 safflower genotypes, Fraj et al. (2013a, b, c) reported that 52 genotypes had shown salinity tolerance potential in pot culture study and were further selected for field trials. In the field trial, Fraj et al. (2013a, b, c) showed salinity tolerance potential of several hundred genotypes and screened out 20 most promising ones that were higher yielder at medium and high salinity. Safflower can be grown on marginal soils as recently Hussain and Al-Dakheel (2018) identified two salt tolerant safflower cultivars (PI248836 and PI167390) which are salt tolerant and high yield producer under salt stress. Yeilaghi et al. (2012) evaluated 64 safflower genotypes and found great variation (7.73–55.9%) among the tested genotypes for seed yield under salt stress (12 dS m-1). The extent of yield reduction was low in salt tolerant genotypes (7.7–28%), while the decrease was > 40% in salt sensitive genotypes. Furthermore, genotype Darab1 produced the maximum oil yield of 1.2 and 0.85 t ha-1 under normal and saline conditions respectively (Table 5). Moreover, due to its ability to withstand suboptimal climate and soil condition, its cultivation has expanded on marginal land (Hussain et al. 2015). Safflower is drought tolerant crop (Lovelli et al. 2007) with a deep root system which can extract water from a depth of 1.6 m (Hojati et al. 2011; Singh et al. 2016) extend up to 1.6 m thus has the ability to grow well on and dry and marginal soils. Moreover, cultivation of drought tolerant safflower cultivar and irrigation management can improve the safflower productivity on water deficit environment and marginal land (Singh et al. 2016). Influence of salinity and drought stresses on safflower is in Tables 3 and 4.
Safflower can be a prospective crop for edible oil purposes under climate change scenario on marginal land as it has less water requirement and can tolerate moderate salinity and drought to some extent (Velasco et al. 2005; Hussain and Al-Dakheel 2018; Han et al. 2009; Istanbulluoglu et al. 2009; Santos et al., 2017; Fraj et al. 2013a, b, c; Yeilaghi et al. 2012). However, there is need to optimize agronomic practices for safflower production for adaptation and to improve its performance on marginal soils. Moreover, water productivity (crop per drop) and irrigation management (using drip irrigation) can help to reduce the yield losses caused by terminal drought.
Rapeseed/mustard
The seeds of mustard (Brassica juncea L.) crop are a good source of edible oil while tender leaves can be used for cooking the food. Rao et al. (2013b) conducted a field experiment on selected genotypes (from a global collection of 100 accessions) and reported that yield of 5 genotypes were higher than rest of the genotypes. Seed yield was highest (3.04 t ha-1) in genotype ATC 93142 followed by followed by genotypes ATC 93358 (2.90 t ha-1) and ATC 93337 (2.89 t ha-1), respectively. In another study, Oplinger et al. (1991) demonstrated that mustard yield varied between 900 and 1200 kg ha-1 while it was yielded 2.5–3.0 t ha-1 under favorable growing conditions. Chauhan et al. (2007) screened the 14 genotypes of Indian mustard (Brassica juncea L.) across two locations under semiarid and irrigated conditions. They reported that yield of mustard ranged from 0.85 to 1.69 and 1.02 to 2.78 under semiarid and irrigated conditions. Among the tested genotypes, PSR-20, PRO-97024, JMMWR-941, IS-1787, PCR-7, RC-1446, and RH-819 produced more yield with under moisture stress and had relatively low drought susceptibility index. The genotypes with low drought susceptibility index for seed yield can be used in breeding programs for development of drought tolerant mustard genotypes. Moreover, mustard is most tolerant among brassica species as B. juncea showed least reduction in morphological traits with higher osmolyte accumulation and better K+/Na+ ratio (Kumar et al. 2009). The saline water (EC 7.48 dS m−1) irrigation to mustard brassica reduced the crop yield by 12.0%, while application of canal water and saline water alternatively and as mixed irrigation reduced the yield by 4.97 and 8.0% respectively. The ability of mustard to withstand drought and salt stress with least reduction in yield compared with other crops of brassica family make it suitable candidate crop to be grown in marginal growth environment.
Soybean
Soybean (Glycine max (L.) Merr.) is a high valued legume crop rich in protein (40–42%) and oil contents (18–22%) (Robert 1986). According to a report of FAO (Food and Agriculture Organization) (2003), it is a major source of valuable human protein. The area and production of soybean has increased substantially since 1961–2007 with an annual growth of 4.6% (Masuda and Goldsmith (2009). Soybean is cultivated in several countries; USA is a major producer and is followed by Brazil, Argentina, China, and India (Rodríguez-Navarro et al. 2011). It can be grown on a range of soils, therefore, can be used as potential crop to improve crop diversification on marginal lands. It is moderately salt tolerant (threshold 5 dS m-1), and a decline in yield was observed in soil having salinity > 5 dS m-1 (Ashraf 1994). Miransari and Smith (2007) reported a decline of 5.3 and 29.7% in soybean yield at 640 and 1280 kg/ha NaCl respectively (Table 5). Chen et al. (2018) reviewed the development of research on salt tolerance of soybean and reported many germplasm accessions with high salinity tolerance and genes/QTL responsible for salt tolerance in soybean, which can be used to develop salt-resistant soybean genotypes.
Soybean is, however, sensitive to drought stress at critical growth stages especially reproductive stage (Table 6). However, deep tillage before soybean cultivation in rainfed condition can reduce the yield reduction due to water stress. Moreover, soybean can improve the crop diversification on marginal lands by using it in crop rotation with cereals or nonleguminous crops, intercropping which will help in improving the soil characteristics through enhanced uptake, breaking of disease and pest cycle. Nevertheless, there is need to introduce the tolerant soybean genotypes with better agronomic practices in areas with poor soil or harsh environment to use it as potential candidate crop on marginal lands.
Maize
Maize (Zea mays L.) is native to Central America but can be successfully cultivated in Canada, Caribbean, America (Central, North and South), Russia, Andean mountains, Chile and Argentina (Ecoport 2010). Maize is an important crop that can be used as energy feed for ruminant livestock. In the cold environments, it is necessary to ensile the crops for year around livestock consumption (Brewbaker 2003). However, in arid, semi-arid and tropical regions, the harvest of maize at different times is always helpful to maintain green forage supply to the ruminants. In some regions, farmers also practice the supply of completely maize plant to the ruminants that is a source of nutritive green fodder year around. In water scare regions of West Asia and North Africa, maize crop is an alternate source of fodder for livestock for smallholder farmers (Potter 2016). Maize is a high energy producer that overcomes several other fodder crops from tropical and subtropical regions and is 40% below in digestibility (Brewbaker 2003). Moreover, maize is not a labor-intensive crop because it should be harvested once after 3 months compared with other forage crops that must be harvested almost monthly (Methu et al., 2006).
Desert gourd
Desert gourd (Citrullus colocynthis (L.) Schrad.) is a xerophytic perennial creeper native to the Mediterranean basin, the Middle East, and South Asia. This is known for its high seed oil content (Bande et al. 2012). The plant often grows as wild in sandy soils covering large areas and surviving under hyperarid Arabian desert conditions with less than 50-mm annual precipitation. The plant can grow in coastal habitat with brackish water (Qasim et al. 2011). The desert gourd can be chosen as a nonfood biodiesel feedstock crop and the possibility for its cultivation in both arid and biophysically marginal arid lands because it occurs naturally in such habitats (Menon et al. 2016). When blended with petroleum, biodiesel from the desert gourd is known to exhibit performance parameters like that of Jatropha (Jatropha curcas L.)—another nonfood biodiesel crop (Mathur et al. 2012). The blends also have lower smoke opacity. The natural distribution of desert gourd suggests that compared with Jatropha, it could be more suitable for growing on lands generally inhospitable to produce crops of any kind or otherwise yield a profit. In a study conducted in sandy desert, Menon et al. (2016) evaluated seed yield of different accessions of desert gourd. The seed yield ranged from 0.47 to 14.95 t ha-1; the highest seed yield was harvested in KMK 1 (14.95 t ha-1) followed by RMS 227 (12.37 t ha-1) and RMS 244 (11.63 t ha-1). However, the oil yield varied between 0.07 and 3.44 t ha-1, and that was the highest in genotype RMS 228 (3.44 t ha-1), followed by RMS 244 (2.28 t ha-1). Interestingly, oil from desert gourd reportedly has lower viscosity (Pal et al. 2010), which could be of great advantage in terms of its potential as a biodiesel feedstock. Root and callus extracts of desert gourd have antimicrobial (Gurudeeban et al. 2010), antiinflammatory (Rajamanickam et al. 2010), antidiabetic (Gurudeeban and Ramanathan 2010), and antioxidant (Gurudeeban et al. 2010) properties.
Legume crops
Cowpea
Cowpea (Vigna unguiculata (L.) Walp.) is an important protein crop that possess a well-developed root system. This is annual plant and can grow, develop, and reproduce in the warmer climates. Some species are erect, and others are climbers. Both leaves and seeds are highly nutritive while seeds are mostly cooked food. Several farmers used the plant as a forage for livestock in Asia and North Africa (Modi and Mabhaudhi 2013). Cowpea is a big source of vegetal protein and is a balance food for poor population in several countries (El-Jasser 2011). Cowpea can be grown as intercrop with many cereal crops like sorghum, maize, and millet in South Asia and North Africa region (Cook et al. 2005a, b). Madamba et al. (2006a, b) reported that through proper soil management and agronomic practices, obtained 0.5 t ha-1 fodder yield. Several varieties of cowpea that has dual purpose usage can provide both grain and fodder especially in Africa (Tarawali et al. 1997a, b). The dual purpose nature of cowpea makes it an ideal candidate crop for food security in North Africa, especially under climate change scenario. Mullen et al. (2003a, b) reported the global grain yield of cowpea ranged between 1.5 and 2 t ha-1. However, in another study, Rao et al. (2013b) grain yield higher than 2 t ha−1was recorded. Cowpea can be grown on marginal land as it can grow successfully on salt affected and water deficit soil condition. The salt and drought tolerant genotypes can produce significant yield on saline areas. Taffouo et al. (2009) conducted a study on 18 cowpea genotypes and demonstrated that yield reduction was less (9–24%) in salt-tolerant genotypes Melakh(9.3%),Tsacre (21.9%), and Garoua GG(24%), while the extent of decline was > 50% in salt-sensitive genotypes, i.e., IT97K-573-1-1 (60.3%), IT04K-227-2 (56.6%), Mouride (56.5%), and Mouola PG (53.2%) (Tables 5 and 6). Likewise, cowpea genotypes exhibit great genetic diversity under drought stress. Ishiyaku and Aliyu 2013 screened 22 cowpea genotypes for drought tolerance indices. They demonstrated that genotype IT93K-452-1 and IT98K-412-13 with drought resistance index (DRI) were drought tolerant. Recently, Belko et al. (2014) tested 30 short and medium duration cowpea cultivars for their DRI and geometric mean productivity (GMP). They found that among short duration genotypes IT85F-3139 (1.6 t ha-1), IT93K-693-2 (1.48), IT97K-499-39(1.31), KVx-61-1(1.37), Mouride (1.36) produced maximum grain yield under drought stress and normal condition and exhibited the higher DRI and GMP. Likewise, for medium duration cowpea genotypes, KVx-421-25(1.80), KVx-403 (1.72), IT93K-503-1 (1.62), IT97K-207-15 (1.59), and IT96D-610 (1.57) produced maximum grain yield under terminal drought stress and possess higher DRI and GMP (Table 6). However, lowest grain yield was recorded in IT93K-93-10 and IT95M-303 (0.53 t ha-1) among short duration genotypes, while IT95M-303 (0.63) produced fewer yield among medium duration cultivars under drought stress. The medium duration cowpea cultivar exhibited higher grain yield than short duration genotypes, and the extent of decline was 60–80% in drought sensitive genotypes while it was 40–50% in drought tolerant genotypes under terminal drought stress (Belko et al. 2014).
Cowpea can help to improve crop diversification on marginal lands reducing soil erosion, improving the microbial activity and nitrogen availability in soil. Moreover, abiotic stress tolerant seed and fodder cowpea cultivars can help in food security through sustainable grain and biomass production for humans and animals respectively. Moreover, cultivation of cowpea can break the cycle of disease and other pests and thus facilitate the cultivation of other field crops.
Faba bean
Faba bean (Vicia faba L.) is rich source of amino acids, proteins, and grown for its food and feed value. It offers ecosystem services through enhanced N fixation and cropping system diversification (Jensen et al. 2010). It can be cultivated on fertile soils but can also be grown on low fertile soils with less water. Drought is a major problem in most of the dry regions, but autumn sown crop is more resistant than spring sown. This might be due to deep root system of autumn sown crop. Moreover, it uses less water than cereals, and therefore, the carryover moisture can enhance the N uptake, growth, and grain yield of succeeding nonleguminous crop in dry land (Papastylianou et al. 1981) and semiarid conditions (Miller et al. 2002). Faba bean can tolerate moderate drought (French 1998). Faba bean survived 8 weeks after drought stress; however, there was a decline in growth, which affects the grain yield (French 1998). Limited water supply can produce comparable grain yield in faba bean (Theib et al. 2005). Al-Suhaibani (2009) found that drought stress limits the yield of faba bean. However, severe yield reduction was noticed at water level below 4000 m3 ha−1 (Table 5). Faba bean can also grow on moderately saline soils. However, salt stress exceeding from 6.5 dS m−1 may cause a significant yield reduction (Katerji et al. 2011). Yield reduction in different legumes under salinity and drought stresses is given Tables 5 and 6.
Faba bean can improve crop diversification on waterlogged soils. For instance, Solaiman et al. (2007) demonstrated that faba bean has the greatest ability to tolerate waterlogging stress than other grain legumes. Moreover, faba bean can successfully grow in semiarid areas with low-cost production and negative impact on environment (De Giorgio and Fornaro 2004). Faba bean cultivation as crop rotation and intercropping with other crops help in improving the diversity, nutrient availability and disease and pest control (Hauggaard-Nielsen et al. 2008). There is need to develop drought tolerant faba bean cultivar as it has the potential to grow as alternate crop on low-input soils.
Chickpea
Chickpea (Cicer arietinum L.) is the third most cultivated legume with high nutritious value for human consumption. Almost 72% of world production is contributed by South Asia alone (FAOSTAT, 2014). It is mostly grown in dry land areas. In south Asia, it is mostly cultivated on low fertile sandy soil. It has very less input and water requirement, therefore, can be grown successfully on soils/climate where most of the commercial crops fail. It is being cultivated on many parts of the world on saline soils (Flowers et al. 2010). Improved chickpea genotypes can help in crop diversification on poor soils. Vadez et al. (2007) screened 263 chickpea accession for salinity tolerance (Table 5; 1.9 L of 80 mM NaCl per 7.5-kg Vertisol). They reported 6-fold variation in the tested germplasm, with some of the accessions produced up to 20% higher yield than previously released salt tolerant cultivar. The tolerant genotypes could maintain higher number of filled pods under salinity stress. Among the tested genotypes desi type was more tolerant than kabuli accessions. Chickpea is mostly grown on dry land or sand soils and need very less water. However, water stress at reproductive stage can reduce the chickpea yield. Leport et al. (2006) studied the influence of drought stress at pod formation on chickpea and found that drastically reduce seed yield in desi and kabuli chickpea types (Table 6) due to increased pod abortion. However, like salinity stress tolerance, chickpea genotypes with large number of flowers and braches can produce better yield under drought-stressed condition. Moreover, kabuli chickpea type is less drought tolerant than desi type.
The chickpea genotypes with salt and drought resistance can be successfully grown on marginal land. It can adapt to poor soil fertility and can substitute cereals in rainfed and salt-affected soils. Moreover, chickpea can improve the soil health through enhanced microbial activities and nutrient availability. Irrigation management at pod formation on these soils can improve the chickpea yield.
Sesbania
Sesbania is an important nutritive legume crop that can be grown successfully under degraded saline habitat. Moreover, it has good potential for forage production in marginal lands. In a research trial, biomass yield of sesbania was reported up to 45 t ha−1 year−1 that was comparably more as compared with that obtained from alfalfa (30 t ha−1) (ICBA 2013; Sattar et al. 2002). It was also found that sesbania is more salt tolerant than alfalfa and has shown its tolerance up to 8–10 dS m−1 (Karadge and Chavan 1983), while alfalfa can tolerate salinity up to 2.0 dS m−1 (FAO 2009).
Amaranth (Amaranthus cruentus L.)
Amaranth can be cultivated in tropical and subtropical regions with warm environmental conditions (Mposi 1999a, b). It is an annual C4 crop has high protein contents in leaves as well as vitamins and mineral and dietary fiber (Andini et al. 2013). The leaves are a good source of certain minerals like riboflavin, niacin, ascorbic acid, calcium, and magnesium (Singhal and Kulkarni 1988a, b). Being a nutritional crop with potential to withstand against drought and salinity, an important crop should have been explored as a candidate crop for marginal lands (Chaudhari et al., 2009). Amaranth has great capacity to grow under water deficit condition as it can recover from severe drought spells. Therefore, it can be grown successfully on water deficit and marginal lands (Liu and Stützel 2002) Moreover, there is also genotypic variation in amaranth as Omami and Hammes (2006) tested A. tricolor and A. cruentus performance under salt and drought stress and found that A. tricolor is more salt tolerant than A. cruentus. Palada and Chang (2003a, b) demonstrated that amaranth can tolerate soil pH in the range of 4.5–8.0.
Because of rapid root and shoot growth, it is proved to be an efficient user of soil moisture (Liu and Stützel 2004a, b). Amaranthus is drought tolerant as it can survive and produce significant biomass under very severe drought condition (Table 7; Chauhan and Abugho 2013). Amaranth can be grown in dry areas successfully where most of the commercial crop fails, and it also has high yield potential. For instance, Barba de la Rosa et al. (2009) reported high yield of two genotypes Gabriela (14 22 kg/ha) and DGETA (1475 kg ha-1) of A. hypochondriacus grown on Mexican Highlands zone than maize and soybean (Table 8).
The screening and selection of salt and drought tolerant varieties of Amaranth might useful for development in marginal lands and to combat food and nutrition security (Alemayehu et al. 2015a, b). Moreover, there is need to optimize the production technology of Amaranth to evaluate it as potential alternate crop on marginal soils.
Nonlegume grain crops
Barley
Barley (Hordeum vulgare L.) is a cereal grain crop that was domesticated thousand years ago in Middle East. Barley grains has significant nutritious value due to the presence of antioxidant phytochemicals (total polyphenols, proanthocyanidins, carotenoids), dietary fiber, protein (14.4%), β-glucan (4.6%) contents, and insoluble bound phenolic acids represented 88.3%, and includes ferulic, salicylic, and gallic acids (Suriano et al. 2018). A significant portion of barley has been used as feed for poultry, cattle, camels, and sheep and for malting, brewing, and preparation for alcoholic and nonalcohol beverages.
It has shown excellent growth and adaptation in a variety of environment including marginal lands and has shown the ability to tolerate drought and moderate salinity. To evaluate a forage yield response towards the different levels of salinity, an experiment (both lab and field based) was carried out on 2300 accessions of Batini barley. The germination, seedling growth, and tillering was measured and found a large variation among each parameter which was attributed to difference in genetic makeup. They concluded that some of barley genotypes were stable and salt tolerant and should contribute to increasing barley production in arid and marginal environments (Al-Dakheel et al. 2012). In a field study, Jaradat et al. (2004a) reported that barley landraces from Oman (Batini 1, Batini 2, and Batini 5) were most salt tolerant genotypes from a global population of 234 Barley genotypes from Omani landraces. Furthermore, Jaradat et al. (2004b) found that Batini 4 was the most salt tolerant barley and possess long roots than all other genotypes. It is relatively more tolerant to salt stress than other cereals (Qiu et al. 2011) and can be grown successfully on marginal lands. For instance, soils with EC of 5 dSm−1 will be marginal for vegetable, but barley (6.8 d Sm−1salinity threshold) can be grown successfully on these soils for forage production (Maas, 1990). Wild barely and its domesticated species are important feed crop in fertile crescent and covers ~ 5 million ha or drought stresses, low input marginal land (Newton et al., 2011).
Barley is mostly cultivated in semiarid or rainfed areas. It can produce good yield on water deficit soils. However, drought stress on critical growth stages like flowering and grain filling can severely reduce the barley yield (Table 9). Mansour et al. (2017) evaluated the performance of barley genotypes against drought tolerance and found that drought tolerant barley genotypes produce more biomass and grain yield (4.97 t ha−1 at 482 mm) with higher water use efficiency (WUE) at low water supply compared with drought-sensitive genotypes at higher irrigation level (3.51 t ha−1 at 561 mm) (Table 10). Moreover, barley can be grown successfully on water limited soils by better irrigation management (drip irrigation) and cultivation of drought tolerant barley cultivars (Mansour et al. 2017) and by application of irrigation water at critical stages like booting and heading. Moreover, barley can be grown in crop rotation with oats and winter rye on marginal soils (low fertility soils) (Ellmer 2008). Furthermore, yield reduction due to impact of salinity and drought are demonstrated in Tables 9 and 10. Barley can be used in crop rotation as it can help in controlling the biotic stresses (disease cycle, pest weed pressure) and also gave good biomass and grain yield under suboptimal growth condition. Moreover, barley genotypes with high tolerance against abiotic stresses need to be developed as it has potential to produce significant yield on marginal lands. It has been cultivated successfully on marginal lands and has shown adaptation for biomass and grain yield in different regions across Middle East, North Africa, East Asia, West Asia, and Mediterranean basins (Al-Dakheel et al. 2012; Jaradat et al. 2004a; Qiu et al. 2011; Saade et al. 2016; Mansour et al. 2017).
Quinoa
Quinoa is an important nutritive grain crop that has shown its potential to grow and produce good and stable yield in different marginal environments. Different genotypes of quinoa have been selected following long-term screening against salinity and drought stress and possess positive attributes in a wider global context (Jacobsen 2003a, b). Previously, quinoa crop has demonstrated high tolerance in extreme soil environment and proved as an industrial crop because it is a multipurpose cereal grain crop with a lot of benefits regarding health perspectives. Quinoa seeds are rich in methionine, lysine, and threonine (amino acids) as the other cereals and legumes are lack of these ones (Repo-Carrasco et al. 2003a, b). Because of high nutrition properties, FAO called the year 2013 as “Year of Quinoa”, and it also identified as a major crop for rehabilitation of marginal lands for future food security (Bazile et al. 2015a, b). In a 3-year study, Rao and Shahid (2012a, b, c) found that quinoa demonstrated good yield potential (range 53.86–359.86 g m2 (2007–2008) and 3.32–258.42 g m2), and adaptation for marginal lands of United Arab Emirates. The yield reported in these studies also indicates a wide genetic diversity under UAE conditions.
Quinoa is attracting the global attention due to the limitation of freshwater use, and its high tolerance to different abiotic stresses. Quinoa has also shown good performance in marginal lands of several Central Asia and MENA countries. These countries have nutrient poor sandy soil and saline water resources. In a pilot scale experiment, different quinoa genotypes were evaluated against forage and seed yield under agroecological conditions of Uzbekistan, Tajikistan, and Kyrgyzstan. The average seed yield varied in the range of 294 g m−2 (Q3) and 557 g m−2 (Q5) in Uzbekistan. However, in Tajikistan, yield was in the range of 147 g m−2 (Q2) to 336 g m−2 (Q1) (ICBA 2015). In FAO’s multicountry quinoa trial, conducted during 2014; the seed yield was varied significantly across different countries and locations within the countries. In a multicountry quinoa yield trial, Dost (2015a, b) found that highest yield (7.50 t ha−1) was recorded in Lebanon, followed by Egypt (3.87 t ha−1), while the lowest yield was recorded in Mauritania (0.23 t ha−1). The average seed yield was in the range of 0.11–0.96 t ha−1 (Iraq), 0.24–1.90 t ha−1 (Yemen), 0.41–3.87 t ha−1 (Egypt), 1.50–7.50 t ha−1 (Lebanon), 0.16– 1.56 t ha−1 (Iran), and 0.03–0.23 t ha−1 (Mauritania). Rao et al. (2013a, b) reported that quinoa yield was higher (> 2 t ha-1) in two accessions Ames 13761 and NSL 106398 that was comparatively higher as compared to those reported by Bhargava et al. (2007a, b). Yield reduction is quinoa is very low even at very high salt stress (Table 7), and some genotypes of quinoa produce higher yield at 40 dS m−1 than control plants (Yazar et al. 2015a, b). Likewise, it has great ability to withstand drought stress (Table 8).
The above results indicate that optimization of production technology on marginal soils, irrigated with low-quality saline water is necessary in order to screen out suitable genotypes from a global collection that can survive in the harsh saline environment and should have a stable and acceptable yield potential. Therefore, tolerant quinoa genotypes should have to be developed as a candidate crop for marginal lands in order to evaluate their yield potential for food security. Quinoa has significant potential to adapt to several climatic environments and can tolerate to some extent drought, salinity, heavy metals, ultraviolet radiations, high temperature, and flooding and other marginal lands. This will facilitate its adaptation under climate change scenario.
Teff
Teff [Eragrostis tef (Zuccagni) Trotter] is a tropical cereal, and its cultivation is mainly confined to Ethiopia and Africa where it is used in food and beverages. The grain of teff is may be smallest carbohydrate rich kernel with high mineral contents and fiber and gluten free (Arendt and Zannini 2013). Due to its gluten-free status, it is becoming popular as wheat substitute for people having coeliac disease (Zannini et al. 2012). It is a low-risk crop can be grown on marginal soils as it has can grow under harsh climatic conditions and diverse ecological surrounding where other cereal crops fail (Arendt and Zannini 2013). Teff can be grown from sea level to high altitude of up to 3000 m above sea level under various rainfall conditions. However, it produces more yields in areas receiving 300 mm (Ketema 1993). Moreover, drought stress at grain-filling stage can limit the grain yield of teff. Shiferaw et al. (2012) studied the influence of drought stress at grain-filling stage in 18 teff genotypes and found a decrease of 42-70% in grain yield. Asfaw and Dano (2011), screened 10 accessions and five varieties of teff for salinity tolerance and found that accession 237186 and variety DZ-cr-37 were salt tolerant, while varieties DZ-Cr-358 and DZ-01-1681 and accession 236514 were salt sensitive. They further reported an increase in grain yield at 2 and 4 dS/m in DZ-Cr-37, 229747, and 237131. However, grain yield was substantially reduced at 8 dS/m in salt sensitive and salt intermediate genotypes. Recently Abraha et al. (2016) tested 144 teff genotypes under optimum and drought stressed conditioned, and they selected DZ-Cr-37, DZ-Cr-385, DZ-01-2053, HO-Cr-136, Dabbi, 207832, Shawa-Gemerra, and Zagure genotypes as drought tolerant due to their early maturity and high yield under moisture deficient condition while genotypes (DZ-Cr-387, DZ-01-787, DZ-01-3186, 9432, 9403, 9415, 205917, 205896, 215678, 213237, Jano, Kaye-Agachew, Purpurea, Kaye-Murri, and Dschanger) produced high yield with increased lodging resistance under normal condition. These genotypes can be used to develop teff cultivar with higher yield under drought stressed condition. In many studies, teff variety DZ-Cr-37 has been used in low rain fall areas which are susceptible to terminal drought as it is proposed to be drought tolerant with high adaptability under diverse climatic condition (Ayele et al. 2001; Cannarozzi et al. 2014; Abraha et al. 2016). Development of salt and drought tolerant genotypes that have capacity to adapt the marginal lands might help to improve the crop diversification on marginal lands.
Sorghum
Sorghum, members of the grass family, is native to Africa, and it is considered an important nutritional crop. Sorghum is now cultivated over the world mainly in developing countries such as China, Central and South America, Africa, South Africa, India, and Indonesia (Mann et al. 1983) (Fig. 4c). Sorghum genus includes several annual and perennial species (approx. 3000). The most important one is Sorghum bicolor (L), Moench, sin. Sorghum vulgare Pers, with Sorghum bicolor var. eusorghum or grain sorghum, Sorghum b. var. technicum or broom sorghum, (c) Sorghum B. var. saccharatum or sweet sorghum and (d) Sorghum b. var. sudanese, or feed sorghum varieties. In Africa and India, the grain is used as food, because it is, like other cereals, a good source of starch, sugars, protein, and phenols that confer it not only nutritional quality but also important beneficial effects on human health. Because of the slow release of sugars than other cereals decreasing detrimental effects in diabetic diseases, the phenolic acids and flavonoids have important anticarcinogenic and antioxidant properties (Awika and Rooney 2004). Being a gluten-free cereal, sorghum importance is growing in all the developed countries in where the occurrence of celiac disease (CD) is sharply rising. The incidence of CD was a little bit higher (from 1:22 to 1:39), for the first- and second-degree relatives of CD patients and 1:56 for patients having either gastrointestinal symptoms or a disorder associated with CD, in USA (Accomando and Cataldo 2004). In Europe, the prevalence of CD is 1:100, and this number is set to rise. Sorghum is a good source of crude fat (3%) and vitamins D, E, and K, B-complex. Sorghum showed significant quantity of thiamin, riboflavin, and niacin. Sorghum has several nutraceutical properties, and its use in food industry for producing malted and distilled beverages (beer, low-alcohol drinks) and popped grains.

Diversified and specialty group of crops (oil seeds, legumes, cereals, medicinal crops, lignocellulose, and fruit crops) that has the capacity of adaptation to tolerate salinity and drought and are suitable for cultivation in marginal environments with proper agronomic and management practices
Sorghum is in fact a plant with a well-developed root system that growing longer than 1 m explains its inclined resistance to thermic stress compared with others cereal crops. Sorghum has adaptation potential to cultivate in arid and semiarid areas where annual rainfall is around 100 mm (Assefa et al. 2010). Sorghum pollen can tolerate higher temperature (45–50 °C). The crop can be cultivated in sandy and nutrient poor soils (pH range 4.5–8.5) because of its low fertilizer requirements for growth, development, and reproductive stage. Sorghum is nowadays commercially in both developed and developing countries (FAOSTAT 2012). Intensive sorghum systems are highly mechanized, use hybrid seed and fertilizers, and are either no-till with herbicide or mechanized tillage. Sorghum yields range from 3 to 5 metric tons per ha under intensive cultivation, as compared with more extensive smallholder systems which average 0.5 to 1.0 metric tons per ha (Clay 2004). Nevertheless, the tendency to grow sorghum on marginal and heavily sloped lands does pose some environmental risks—including soil degradation and erosion—that can be mitigated through the adoption of best practices. Early sorghum hybrids exhibited a 40% yield advantage over open pollinated varieties (Duvick 1999). In South Africa to some extent, this intensification has already occurred.
Sorghum is a multipurpose industrial crop that has shown high tolerance against salinity and is very suitable for saline and marginal lands (Rao et al. 2015). Sorghum, a C4 crop, is normally a tall annual grass with adventitious root system has been proved to be useful for grain as well as forage production (Harada et al. 2000). In USA, different varieties of sweet sorghum can be used as fodders and development of hybrid fodder. Once established, the sweet sorghum has shown the salinity tolerance potential (Cook et al. 2005a, b) while other workers also described the sorghum as highly salt-tolerant crop plant (Fahmy et al. 2010). Balole and Legwaila (2006) reported that forage sorghum produced an average of 20 t ha−1that might be reached at 75 t ha−1under favorable growth conditions. Sorghum should have to be promoted in water scare regions because it needs less water during growth period and to reach to reproductive stage. Thus, sorghum is very suitable in drought prone area and areas with declining aquifers (Rao et al. 2015). In a long-term screening, selection, and development of salt tolerant genotypes, Krishnamurthy et al. (2007) screened seven genotypes that showed tolerance against salinity under field conditions. Influences of salinity and drought on the yield of sorghum genotypes are shown in Tables 11 and 12.
In India, 75% of sorghum is already of high-yielding varieties (Pray and Nagarajan, 2009). But despite the increasing global awareness of plant genetic resources and links to food security, sorghum is relatively neglected in scientific research, agricultural programs, and policies (Garí 2002). In South Africa, sorghum is grown either as a rainy season crop or as a postrainy season crop. The highest sorghum yields are achieved with the rainy season crop (around 1 ton/ha) due in large part to abundant water access—sorghum will also tolerate poorly drained soils and can survive temporary waterlogging during the rainy season (Fageria 2011). Drought still poses a threat to rainy season sorghum, however, particularly in years of midseason drought. The postrainy season crop is even more drought constrained as the crop is dependent upon postrainy season stored soil moisture (Murty et al., 2007). In addition to direct impacts on the plant, drought can lead to reduced nutrient uptake and make sorghum more susceptible to pests (Assefa et al. 2010). Efforts to overcome water constraints on sorghum production in smallholder systems focus on improved water management, planting timing, and using diverse and drought-resistant varieties. In both SA and SSA, optimizing planting dates for sorghum to prevent water stress during water-sensitive growth periods is another key strategy to mitigate sorghum yield reductions (Assefa et al. 2010). Singh et al. (2016) identified late sowing and low water-holding capacity of shallow soils as the key impediments to expanded sorghum productivity in India. Depending on the local situation, making maximum efforts to prepare land early and plant the crop as early as possible with the first rains can boost crop production. It also considered a good strategy to use local varieties that are often better adapted to drought conditions than commercial varieties. However, local varieties are often lower yielding and typically do not perform as well under optimal growing conditions (Yadav, 2010). Sorghum researches have been focused on plant breeding for drought tolerance and for early maturation (Assefa et al. 2010). While poor soil fertility is a significant constraint to sorghum production, very few smallholder farmers in SSA and SA use fertilizer (and even more rarely are inorganic fertilizers applied on sorghum and millet plots). Moisture deficiencies during crop growth inhibit nutrient uptake, making fertilizer application even less beneficial and economical. Clay (2004) reported that while some smallholder sorghum farmers have invested in hybrid improved seeds such as in India, fertilizer use remains uncommon. In contrast, in developed countries—and in some intensive commercial operations in South Asia—major increases in dryland yield have been attributed to increased fertilizer use and hybrid seed advancement (Assefa and Staggenborg 2010). Similar to other crops, soil quality, and sorghum yields are higher in sorghum production systems where plant foliage remains in the field as a mulch following harvest—practice relatively more common when the crop is grown for grain rather than silage (Meyer et al. 1999). While mulching can mitigate the nutrient-depleting effects of repeated cropping and short fallows, households often prefer to use mulch for building material, fuel, and fodder (Wezel, 2000). Cover crops, crop rotation, and continuous farming along with green manure can further reduce the fertilizer and water requirements of sorghum crops (Clay 2004) or both sorghum production, conservation till age and no-till agricultural practices have been shown to reduce erosion by up to 80% (Meyer et al. 1999). Sorghum plant matter is particularly beneficial to soil if it is chopped at the time of harvesting. Weeds are a primary constraint to sorghum production (Estep et al. 2011; Waddington et al. 2010). Pests and diseases caused especially by fungi, viruses, bacteria, and nematodes sorely limit the sorghum production (Clay 2004). Grain molds lead to important decrease in sorghum grain yield and quality, especially in areas in which ameliorated cultivars have been used. The grain mold problem is due to the arrival of late rains after grain fill. A wide variety of herbicides and pesticides are also used on sorghum crops in intensive production systems, especially in SA (Clay 2004; Khan et al. 2000). Literature on environmental impacts of pest management practices specific to sorghum production is limited up to now. Herbicides and pesticides regularly used on intensive sorghum crops have been shown to cause harm to surrounding ecosystems (Clay 2004; Ragnarsdottir 2000; Kamrin 1997), but few data are available for sorghum yet.
Sorghum is a cereal that could be easily adapted to stress conditions reducing only in part its growth and nutritive properties mainly in respect to the cultivar used. The different cultivars adopt different mechanism for salt stress tolerance increasing the production and the accumulation of organic solutes. Sorghum is really appreciated for its nutritional properties and agronomic advantages mainly in arid and semiarid environments. Sorghum grains are an important source of gluten-free flour making it suitable to hundreds of gluten intolerant populations. Sorghum flour combined with flours from other cereals and legumes can be an important opportunity for ameliorating its nutritional value. Food processing represents also an economic benefit for this cereal. Research on how different processing can affect sorghum nutritional properties could help spread consume of sorghum as food.
Millets
Millets are highly variable small grain annual crops cultivated (Kothari et al. 2005) in temperate, subtropical, and tropical regions with low-input supply and mostly grown in harsh environments (salinity, low soil fertility, drought, heat, and chilling stresses). They play important role in ensuring food security to poor people in semiarid tropics due to low cultivation of major food crops owing to poor soil fertility and low precipitation (Das and Rakshit 2016). Pearl millet, finger millet, proso millet, and foxtail millet are the most cultivated species used as food, feed, and fuel (Hithamani and Srinivasan 2014) (Fig. 4d). All millets other than sorghum and pearl millets are called small or minor millets. Pearl millet represents more than half of global millet production. Proso millet is used as a feed for birds in developed countries and as food in parts of Asia. The foxtail millet is more utilized as food in China and in Europe. The other species (aia, kodo and small millet, fonios, and teff) are instead more used locally in small countries as food grains. The different species differ from one another, due to their physical characteristics, growth duration, quality, and adaptation to soil and climate. For the nutrient and protein content, millet can be considered equivalent or better than wheat, rice, corn, and sorghum. Millets are enriched in proteins, vitamins especially vitamin A, and oil content that is better than maize grain oil (Nambier et al., 2011). As compared with other cereals, it is less susceptible to insects and pests. Additionally, millets are considered the most suitable crops for agriculture and food security on lands with low fertility. Millets are able to easily grown on low fertility and sandy soils in which other cereal crops produce low yields (Amadou et al. 2013; Changmei and Dorothy 2014). Millet is gaining attention in designing the modern foods because it is a gluten-free cereal which is rich in polyphenols and other biological active compounds with beneficial effects on human health. Due to the increased awareness regarding its health promoting compounds, and its ability to grow in adverse environmental conditions (drought, salinity, and high temperature), millet represents a good alternative to traditional cereals such as wheat and rice to be cultivated mainly in marginal lands. Millet has nutritive properties equal or superior to the major cereals (Leder 2004), and its nutritional properties change in respect to the variety to the climatic conditions and the soil on which it is grown. The average carbohydrates content of millets varies from 56.88 to 72.97 g/100 g, the minor amount was found in barnyard millet. The average protein content of all the millets range between 10 and 11%, except for finger millet, in which protein ranges from 4.76 to 11.70 g/100 g. Proso millet is the variety that contains the highest amount of proteins. The lipid content in all millets are similar to the other cereals; finger millet contains the lowest lipid amount while pearl millet has the highest amount of lipid. Little millet (Panicum sumatrense) and kodo millet (Paspalum scrobiculatum) contain the highest dietary fiber (785% higher than rice and wheat); this makes millets a food suitable for diabetic people (Chandel et al. 2014). Millets contain significantly higher amount of calcium and mineral than other cereals (Pontieri et al. 2014). Barnyard millet and pearl millet are richer in iron than the other millet varieties and other cereals. Millets contain also high amount of β-carotene and B vitamins especially riboflavin, niacin, and folic acid. Millets contain also a great amount of flavonoids and phenolic compounds (Chandrasekara and Shahidi 2011). Millet also possesses different secondary metabolites that have shown therapeutic and pharmacological potential. Several millet genotypes showed antioxidant, antimutagenic, antiestrogenic, antiinflammatory, and antiviral effects (Devi et al. 2014). Total antioxidant capacity of finger, little, foxtail, and proso millets resulted high for their high content of total carotenoid and tocopherol (Dykes and Rooney 2006). Millet is able to grow in presence of salinity but at different extent depending on the salt concentration and millet variety. Kumari and Vishnuvardhan (2013) showed a different response of 3 diverse accessions (IC426676, IC382888, and IPS 145) of Kodo millet to NaCl salinity. They demonstrated that all the 3 accessions germinated and grew at 50 mM NaCl, but increasing salinity germination percentage and seedling growth decreased. The accession’s most resistant to salinity was IC 426676 which is able to germinate and grow at 150 mM NaCl with a short decrease also in the amount of total protein. The production of biomass and grains in field in presence of salinity depended on the genotypes and accessions. Khan et al. (2000) examined 8 varieties of millet demonstrating that at 8 dS/m, only a reduction of 20% in all the studied varieties was observed. Increasing salinity a progressive decrease in biomass was detected. The production of grain decreased increasing salinity. The greatest decrease was observed at 16 dS/m, but for up to 8 dS/m, a 20% reduction was observed. Although, in general, millets are able to grow better than cereals in semiarid environments, drought or inadequate moisture represents a threat affecting productivity. Studies in pearl millet showed that drought caused yield loss in respect to the variety and the duration of the stress. Pearl millet productivity and growth were less affected by drought conditions instead little millet productivity was decreased by drought stress conditions. The effects of salinity and drought on yield of millet are shown in Tables 13 and 14.
Millet adapt to diverse intensity of drought and salinity from seed germination to growth. Many species are able to germinate and grow on arid lands also in extreme drought and salinity conditions without modifying their nutritional values. Millet environmental adaptability determines the numerous plants per unit of area, allowing the extension of this plant to drier and salinity areas.
Triticale
Triticale is a human made hybrid cereal grain, obtained from crossing wheat and rye (Zhu 2018) (Fig. 4e, f); it is self-pollinating (similar to wheat) and not cross pollinating (like rye). It combines the best properties of both its parents, inheriting the most of wheat’s qualities important for food products, and the most of rye’s properties which confer resistance to disease, tolerance to drought, hardiness, and adaptability to “hard” soils. The first varieties had spring wheats as parents easily killed by low temperatures. Subsequently, winter wheats have been used as parents, producing varieties with good winter resistance. For instance, recent publications evidenced that spring triticale is a vital drought tolerant crop in North Africa and play a significant role in alleviating poverty for many needy families in some developing countries. Forage yield was reported in the range of 5.6–10.9 t ha−1 and grain yield between 2.2 and 5.6 t ha−1 (ICBA 2013). After a long-term salinity trails conducted over years at ICBA, in order to identify salt tolerant accession among a large population, the accession PI 429166 had excellent grain yield and dry matter production. The highly productive winter-type triticale can instead be successful cultivated in cold and wet environment of Northern Europe.
Triticale is grown on about 3 million ha in the world (FAO, 2003), and it is widely used as forage in farming system even if triticale grain contains vitamins, mineral nutrients, and proteins in concentration similar to wheat and rye. For these reasons, triticale should be highly recommended as food, mainly because both spring and winter varieties on average contain 10% of proteins, 56% of carbohydrates 2.8% of crude fibers, 4% of free sugars, and 1.6% of ash, representing a valid alternative to wheat and rye mainly in area where these two species have difficult to grow. Triticale is grown in areas with an annual average rainfall ranging from 300 to 900 mm. Very little triticale is irrigated. Triticale for grain is generally sown in autumn and harvested in summer time. Triticale was able to grow under salinity and drought stress conditions and numerous works evidenced that triticale lines were better than wheat cultivars for resistance to drought taking in account grain yield and majority of physiological traits. Giunta et al. (1993) confirmed that triticale genotypes were more drought tolerant than wheat and there was slight reduction in its grain yield. Wheat production decreased significantly (by 25, 54, and 87%) under drought stress, while triticale slightly or nonsignificantly decreased its yield (by 8%) in comparison with the irrigated control, evidencing a greater drought tolerance of triticale attributable to the greater ability of its roots to absorb water from soil. Akbarian et al. (2011) found an inverse and significant relationship between grain yield loss due to drought stress and proline content. These findings provided evidences on the key role of proline in inducing tolerance to drought stress.
Proline was also evaluated in 18 lines of triticale under salinity (16 dS m−1), and the results showed an inverse and significant relationship between grain yield loss under salinity and proline content confirming its key role in alleviating abiotic stress in plants (Salehi and Arzani 2014). Akgun et al. (2011) evidenced that high salinity levels (25 dS m−1) decreased seed germination (− 50%) of 5 triticale genotypes obtained from the CIMMYT while increased proline levels. Kaydan and Yagmur 2008 evaluated seed germination and seedling growth of triticosecale Witm. cv. Presto under osmotic stresses caused by water and NaCl. Their results evidenced that water stress, at the same osmotic potential of salt, was more detrimental than salinity compared. They compared different varieties having different seed size and highlighted that seed with greater size resisted more to salinity than seeds with smaller size. Lirong et al. (2016) tested eight varieties of triticale for drought resistance using 20% polyethylene glycol 6000. Their results showed that the seedling fresh weight of all varieties decreased, MDA content increased, proline content, and soluble sugar content increased with water stress. Soluble protein contents raised in some varieties while decreased in some others. The activity of SOD increased, and POD, CAT activity variations were different in all the 8 varieties. After a comprehensive analysis, we can conclude that the drought resistance depended mainly on the varieties and less on the entity of stress. Table 15 and 16 demonstrate influence of salinity and drought on yield of Quinoa, Amaranth, Chia, and Teff.
Medicinal Crops
Chia
Chia (Salvia hispanica L.,) is an ancient food rich in ω-3/-6 fatty acids, insoluble fiber, protein, minerals, and antioxidants (Orona-Tamayo et al. 2017). Its seed contain antibiotics phenolics and terpenoids that make it medicinally important plant (Baricevic and Bartol 2000). Chia is commercially grown in Bolivia and Paraguay; nonetheless, during last 15 years, its area also expanded in Argentina, Australia, and Mexico due to high economic return (8-12000 USD/ton) (Peperkamp 2014). The increase in area under Chia is due to its ability to produce significant yield on marginal lands and low-input condition. Under low-input condition, it can produce around 600 kg/ha, with some growers have reported upto1200 kg ha−1 under suboptimal growth condition (Coates 2011). However, in normal soil with optimal input supply 2500 kg ha−1 has also been reported (Cahill 2003; Ullah et al. 2015).
Chia can be good alternative crop under marginal growth conditions as it can grow on semiarid (Bochicchio et al. 2015) and acidic soils (Muñoz et al. 2013). It performs best on sand soils and is adapted to low nutrient supply (Yeboah et al. 2013). However, it has less resistant to salinity (Heuer et al. 2002) as reduction of 69, 76.1, and 89.8% in biomass of chia was observed at 20, 40, and 60 mM salt stress respectively (Table 8). It can produce good yield even under high summer temperature and drought condition (Win et al. 2018) as Chia planted in spring March and April produced 32.2 and 27.9% less yield than may planted crop. Baginsky et al. (2016) studied influence of climate on chia production in three distinct regions and found that Chia produced maximum yield (> 2900 kg ha-1) under desert condition of Valle de Azapa (VA) and Canchones (CH). Chia is sensitive to low temperature as can cause significant yield reduction. Tables 8 and 9 demonstrate influence of salinity and drought on yield of quinoa, amaranth, chia, and teff. Chia is a good option for crop diversification on dry, sandy soil, and high temperature environment; it got attention in recent years. Development of salt sand chilling resistant chia genotypes will help to improve the crop diversification on marginal land, as it is very low input requiring crop and can grow under limited water supply and withstand high temperature.
Moringa
Moringa (family Moringaceae) is an Old-World dry tropical plant genus with 13 species that has shown great potential in terms of food, feed and pharmacopeia. Three species of Moringa have shown great potentials to be suitable feedstock for sustainable production of biodiesel. These are Moringa oleifera (Atabani et al., 2013), Moringa stenopetala (Ejigu et al., 2010) and Moringa peregrina (Salaheldeen et al., 2015). In the UAE, two Moringa tree species are growing the exotic M. oleifera which is cultivated as ornamental plants in the UAE and the native M. peregrina, which is growing naturally in the mountains of the UAE. Several colleagues reported that M. oleifera has significant nutritional benefits and is a proven source of animal forages. Meanwhile, it is quite suitable for cultivation to nutrient poor saline and marginal lands with low water and nutrient availability (Nouman et al., 2014). Ayerza, (2011) indicated that M. oleifera has oil productivity of 580 kg ha−1. This indicates this M. oleifera could be cultivated economically in subtropical regions of the world (Salaheldeen et al., 2014). The whole Moringa tree is a good alternate source of pharmaceutical agents and antioxidant and has nutritional feedback.
It has wood that is good for firewood and charcoal and also resists termites. Al-Kahtani, (1995) reported 12.4% moisture, 53.9% oil, 23.2% protein, 12.1% dietary fibers, 17.5% carbohydrate, and 2.6% ash. However, the oil content of the exotic M. oleifera seeds was recorded as 34.8% (Anwar and Rashid 2007) in the dry seeds. Polyunsaturated fatty acids, amino acids, and sterols can be obtained from M. peregrina seeds (Al-Dabbas 2010). Salaheldeen et al. (2015) converted the crude oil of M. peregrina to biodiesel by the transesterification reaction, catalyzed by potassium hydroxide and got high ester content (97.79%). Sharma et al. (2009) have reported that Moringa oil exhibited the highest thermooxidative stability, compared with other vegetable oils, such as Jatropha oil, cottonseed oil, canola oil, and sunflower oil (Sharma et al. 2009).
Lignocellulosics crops
Perennial forage grass
Cenchrus ciliaris L. is an important forage plant that can be grown in drought and saline affected marginal lands. It can be cultivated in different arid and semiarid areas of Asia, Africa, and Australia (Arshadullah et al. 2011). It is C4 plant and is valued for palatable forage and intermittent grazing. Al-Dakheel et al. (2015) reported that the dry matter yield varies from 24.5 to 73.0 t ha−1 at 15 dS m−1). However, the biomass yield varies from 26.3 to 65.8 t ha−1among the top five genotypes at 15 dS m−1. These screen and selected accessions of C. ciliaris are a good forage source for animals in arid and semiarid regions. Meanwhile, Al-Dakheel and Hussain (2015) demonstrated that genotype Grif 1619 was the highest dry matter yielding accession at 15 dS m−1.
Fruit crops
Some fruit crops have shown promising potential and adaptation under marginal environment. The prominent fruit crops that can be grown in saline and marginal lands include date palm (Phoenix dactylifera L.), olive (Olea europaea L.), phalsa (Grewia asiatica L.), chicle (Manilkara zapota (L.) P. Royen), guava (Psidium guajava L.), jambolan (Syzygium cumini L.), Indian jujube (Ziziphus mauritiana Lam.), Indian gooseberry (Phyllanthus emblica L.), and karanda (Carissa carandas L.) (Maas and Hoffman, 1977; Qureshi and Barrett-Lennard, 1998). The performance of two important fruit trees (date palm and olives) are summarized here.
Date palm
Date palm (Phoenix dactylifera L.), is an important nutritive fruit crop, rich in minerals, nutrients, and carbohydrates. The fruit tree has the capacity to tolerate harsh climate, marginal environment drought, high temperature, and salinity (12.8 dS m–1) (Ramoliya and Pandey 2003). According to the study of Aljuburi (1992), the Lulu cultivar was more sensitive than other three tested varieties (Khalas, Barhee, Boman) at different salinity levels (0, 0.6, 1.2, and 1.8%). According to reports of Kurup et al. (2009), date palm can tolerate the salinity up to a certain level of threshold. Ramoliya and Pandey (2003) demonstrated that date palm is tolerant to salinity up till 12.8 dS m–1. Meanwhile, several seed germination, growth, and physiological features showed tolerance against salt stress in the range of 4.3–12.8 dS m–1 while higher salinity reported to be lethal at different growth stages. In seedling growth bioassays, the variety Nakhla hamra showed enhancement in epicotyl length, root growth, and proline contents under drought condition imposed by PEG. While other variety Tijib showed more tolerance against salinity based on physiological and biochemical indicator (Djibril et al. 2005). Al Kharusi et al. (2017) evaluated salinity tolerance potential among 10 date palm seedlings and subjected them to 240 mM salt stress. They showed that photosynthesis, electrolyte leakage, and the shoot K+/Na+ ratio were reduced in the susceptible cultivars while Manoma and Umsila were reported as resistance cultivars. Date palm varieties, Deglet Noor’, and ‘Medjool showed poor performance against salinity (24000–520 ppm) and their growth; leaf development was highly decreased under salt stress (Furr and Ream 1968). From research trials of Aljuburi (1992), seedling growth, development, and leaf elongation were inhibited at all levels of salinity (0, 0.6, 1.2, and 1.8%) in different date varieties. He concluded that Lulu variety was more tolerant to salinity as compared with other varieties.
Several plasticity traits such as photosynthesis, gas exchange measurements, chlorophyll a/b ratio, ribulose 1,5-bisphosphate activity, ionic accumulation in leaves especially Na+ and K+, and electrolyte leakage might be used as stress biomarkers to elucidate the salinity in date palm seedlings (Youssef and Awad 2008). They found that seedlings accumulated a significant amount of salts in their foliage parts following salinity treatments. Al-Wali et al. (2011) documented that tissue culture based Khalas variety was significantly affected in terms of seedling growth and development, and response was different in pots added with or without mulch. The mechanism of salinity tolerance in date palm is not well clear. However, Alrasbi et al. (2010), reported that date plants have capacity to control ion homeostasis through restricted entry and exit of Na+, Cl-, through maintain the K+ internal cellular content. According to research trials of Alrasbi et al. (2010), Khalas, Khunaizy, and Abunarinjah varieties showed normal growth following irrigation with saline water at 9 dS m−1. However, they observed 50% reduction in growth parameters at 18 dS m−1 salinity. The mechanism of salt tolerance was Na and Cl in the leaves through counteracting with K.
Olives
Olive tree has shown promising salt tolerance potential and researchers have pointed out salt exclusion as a major phenomenon behind this story (Gucci and Tattini, 1997). Some varieties showed moderate salinity tolerance while certain others were highly salt tolerant. In the saline environment of Northern Chile, Sotomayer et al. (1990) demonstrated excellent yield following irrigation with saline water (13 dS m−1). The truck growth seemed to have relation with age. A decrease of 30% in 2000 and 55% in 2001 as compared with 1999 was observed; salinity tolerance potential decreased with the passage of time (Aragues et al. 2005). Furthermore, Fernandez and Moreno (1999) demonstrated that olive is sensitive to waterlogging and should be preferentially cultivated in sandy and well-drained soils. Chartzoulakis (2005) showed the salt tolerance in olives depended upon the cultivars and prevention of salt translocation was the main mechanism involved in this context. Meanwhile, osmotic adjustment, stomatal closure, and leaf abscission appeared to play a role. The fruit weight and oil content decreased under salt stress.
Conclusion and future research needs
Selection and potential use of tolerant genotypes with proven adaptation capacity to nutrient poor marginal lands provides good opportunity to achieve stable yield. Crop diversification, innovative high-throughput phenotyping strategies, and sustainable agriculture based on drought and salt-tolerant plant species are likely to be the key factor for maintaining/improving food security in marginal environments. Crop diversification is a very cost-effective option to improve the productivity of marginal lands with high economic return. The joint research strategies could promote the development, improvement, and use of diversified crops such as cereals, oil seed crops, forages, and legumes to shift from subsistence farming to climate smart agriculture. We can conclude that cereals, oil seed crops, forages, and legumes, should have equal importance when crop diversification will be adapted for increasing sustainable agriculture. Different strategies in terms of high throughput technologies, genomic resources, genes associated with stress responses have to be recruited to generate stress tolerant diversified crops. Crops improvement and sustainable production systems need to be integrated in order to address more efficiently future demands of food production presently exacerbated by global climate changes.
In future, the linkages between farmers, researchers, policy makers, and community workers needs to be strengthen in order to achieve the maximum benefits from the use of nonconventional water resources and crop diversification. In this context, it is of primary importance to foster the use of drought and salt tolerant genotypes, agroecological principles, plant biodiversity, and nonconventional water resources to speed up the process of sustainable development on the marginal lands. Farmers and research scientist should have to continue dialogues at different platforms to gain maximum output from crop diversification. Meanwhile, better utilization of agrobiodiversity, environmental resources, and providing climate smart solutions to the farmer’s current problems should be on the top of the agenda at the regional and national level. Providing better options to farmers to grow alternate crops will enhance their capacity to adapt to the climate change agriculture perspectives. Inclusion of medicinal and industrial crops (oil seed and lignocellulose crops) in the current cropping system will stabilize the farmer’s income in a sustainable way and opportunity to better utilization of available natural resources in a judicial way.
References
Abraha MT, Shimelis H, Laing M, Assefa K (2016) Performance of tef [Eragrostistef (Zucc.) Trotter] genotypes for yield and yield components under drought-stressed and non-stressed conditions. Crop Sci 56(4):1799–1806
Accomando S, Cataldo F (2004) The global village of celiac disease. Dig Liver Dis 36(7):492–498
Agarwal S, Kumar A, Singh PK (2016) Influence of saline water irrigation on crude protein and amino acids in two genotypes of finger millet (Eleusine coracana Gaertn.). Int J Agric Sci Res 6:261–268
Ahmed IM, Cao F, Zhang M, Chen X, Zhang G, Wu F (2013) Difference in yield and physiological features in response to drought and salinity combined stress during Anthesis in Tibetan Wild and Cultivated Barleys. PLoS ONE 8(10):e77869. https://doi.org/10.1371/journal.pone.0077869
Akbarian A, Arzani A, Salehi M, Salehi M (2011) Evaluation of triticale genotypes for terminal drought tolerance using physiological traits. Indian J Agric Sci 81(12):1110–1115
Akgun I, Kara B, Altindal D (2011) Effect of salinity (NaCl) on germination, seedling growth and nutrient uptake of different triticale genotypes. Turkish J Field Crops 16(2):225–232
Al Kharusi L, Assaha DV, Al-Yahyai R, Yaish MW (2017) Screening of date palm (Phoenix dactylifera L.) cultivars for salinity tolerance. Forests 8(4):136
Al-Dakheel AJ, Hussain MI (2015) Saving fresh water resources through cultivation of salt-tolerant forage grasses: seasonal and genotypic variations. Biosalinity News 16:10–12
Al-Dakheel AJ, Hussain MI (2016) Genotypic variation for salinity tolerance in Cenchrus ciliaris L. Front Plant Sci 7:1090
Al-Dakheel AJ, Fraj MB, Shabbir GM, Al Gailani AQM (2012) Evaluation of Batini barley landraces from Oman and breeding lines under various irrigation salinity levels. Agric Sci Res J 2:42–50
Al-Dakheel AJ, Hussain MI, Abdul Rehman AQM (2015) Impact of irrigation water salinity on agronomical and quality attributes of Cenchrus ciliaris L. accessions. Agricultural Water Management 159:148–154. https://doi.org/10.1016/j.agwat.2015.06.014
Alemayehu FR, Bendevis MA, Jacobsen SE (2015a) The potential for utilizing the seed crop amaranth (Amaranthus spp.) in East Africa as an alternative crop to support food security and climate change mitigation. J Agron Crop Sci 201:321–329. https://doi.org/10.1111/jac.12108
Alemayehu FR, Bendevis MA, Jacobsen SE (2015b) The potential for utilizing the seed crop amaranth (Amaranthus spp.) in East Africa as an alternative crop to support food security and climate change mitigation. J Agron Crop Sci 201:321–329. https://doi.org/10.1111/jac.12108
Aljuburi HJ (1992) Effect of sodium chloride on seedling growth of four date palm varieties. Ann Arid Zone 31(4):259–262
Almodares A, Hadi MR, Dosti B (2008) The effects of salt stress on growth parameters and carbohydrates contents in sweet sorghum. Res J Environ Sci 2(4):298–304
Al-Naggar AMM, Badran AEE, El-Moghazi Mai MA (2017) Genotype and drought effects on morphological, physiological and yield traits of quinoa (Chenopodium quinoa Willd.). Asian J Adv Agric Res 3(1):1–15
Alrasbi SARN, Hussain, Schmeisky H (2010) Evaluation of the growth of date palm seedlings irrigated with saline water in the Sultanate of Oman, ISHS Acta Horticulturae. 882: IV, International Date Palm Conference
Altieri M (2004) Linking ecologists and traditional farmers in search for sustainable agriculture. Front. Ecol. Environ 2:35–42.
Altieri MA, Koohafkan P (2013) Strengthening resilience of farming systems: a key prerequisite for sustainable agricultural production. Wake up before it is too late: make agriculture truly sustainable now for food security in a changing climate. UNCTAD, TER13 Report, Geneva
Al-Dabbas MM, Ahmad RAFAT, Ajo RY, Abulaila KHALED, Akash MUHANAD, Al-Ismail KHALID (2010) Chemical composition and oil components in seeds of Moringa peregrina (Forssk) Fiori. Crop Res 40(1):2
Al-Kahtani HA (1995) Moringa peregrina (Al-Yassar or Al-Ban) seeds oil from Northwest Saudi Arabia. J King Saud Univ, 1, 31–45
Al-Suhaibani NA (2009) Influence of early water deficit on seed yield and quality of faba bean under arid environment of Saudi Arabia. Am-Eurasian J Agric Environ Sci 5(5):649–654
Al-Wali BR, Kurup SS, Alhammadi MS (2011) Response of Tissue Culture Plantlets of Date Palm (Phoenix dactylifera L) var. Khalas to Moisture Stress under the UAE Condition. M.Sc. Thesis, UAE University, Al-Ain
Amadou I, Gounga ME, Le GW (2013) Millets: nutritional composition, some health benefits and processing-a review. Emirates J Food Agric 25:501–508. https://doi.org/10.9755/ejfa.v25i7.12045
Andini R, Yoshida S, Ohsawa R (2013) Variation in protein content and amino acids in the leaves of grain, vegetable and weedy types of amaranths. Agronomy 3:391–403. https://doi.org/10.3390/agronomy3020391
Anwar F, Rashid U (2007) Physico-chemical characteristics of Moringa oleifera seeds and seed oil from a wild provenance of Pakistan. Pak. J. Bot. 39:1443–1453.
Arendt EK, Zannini E (2013) Teff. Cereal grains for the food and beverage industries. 351–369e. https://doi.org/10.1533/9780857098924.351
Arshadullah M, Malik MA, Rasheed M, Jilani G, Zahoor F, Kaleem S (2011) Seasonal and genotypic variations influence the biomass and nutritional ingredients of Cenchrus ciliaris grass forage. Int J Agric Biol 13:120–124
Asfaw KG, Dano FI (2011) Effects of salinity on yield and yield components of tef [Eragrostistef (Zucc.) Trotter] accessions and varieties. Curr Res J Biol Sci 3:289–299
Ashraf M (1994) Breeding for salinity tolerance in plants. Crit Rev Plant Sci 13:17–42
Assefa Y, Staggenborg SA (2010) Grain sorghum yield with hybrid advancement and changes in agronomic practices from 1957 through 2008. Agron J 102:703–706. https://doi.org/10.2134/agronj2009.0314 https://www.agronomy.org/publications/aj/articles/102/2/703
Assefa Y, Staggenborg SA, Prasad VPV (2010) Grain sorghum water requirement and responses to drought stress: a review. Crop Management 9. https://doi.org/10.1094/CM-2010-1109-01-RV
Atabani AE, Mahlia TMI, Masjuki HH, et al. (2013) A comparative evaluation of physical and chemical properties of biodiesel synthesized from edible and nonedible oils and study on the effect of biodiesel blending. Energy 58:296–304
Awika JM, Rooney LW (2004) Sorghum phytochemicals and their potential impact on human health. Phytochemistry 65(9):1199–1221
Ayele M, Blum A, Nguyen HT (2001) Diversity for osmotic adjustment and root depth in TEF [Eragrostistef (Zucc) Trotter]. Euphytica 121:237–249. https://doi.org/10.1023/A:1012099914738
Ayerza R (2011) Seed yield components, oil content, and fatty acid composition of two cultivars of moringa (Moringa oleifera Lam.) growing in the Arid Chaco of Argentina. Industrial Crops and Products 33:389-394
Aymen EM, Zhani K, Meriem BF, Hannachi C (2012) Seed priming for better growth and yield of safflower (Carthamus tinctorius) under saline condition. J Stress Physiol Biochem 8(3):135–143
Ayodele VI (1999) Influence of soil water stress at different physiological stages on growth and seed yield of Amaranthus species. In III International Symposium on Irrigation of Horticultural Crops 537 (pp. 767-772)
Ayres RS, Westcott DW (1976) Water quality for agriculture. In: Irrigation and Drainage Paper 29. Rome, FAO
Bafeel SO (2014) Physiological parameters of salt tolerance during germination and seedling growth of Sorghum bicolor cultivars of the same subtropical origin. Saudi J Biol Sci 21:300–304
Baginsky C, Arenas J, Escobar H, Garrido M, Valero N, Tello D, Pizarro L, Valenzuela A, Morales L, Silva H (2016) Growth and yield of chia (Salvia hispanica L.) in the Mediterranean and desert climates of Chile. Chilean J Agric Res 76(3):255–264
Balole TV, Legwaila GM (2006) Sorghum bicolor (L.) Moench. Record from Protabase. Brink M. & Belay G. (edts). PROTA (Plant Resources of Tropical Africa / Ressources végétales de l’Afrique tropicale), Wageningen, Netherlands.
Bande Y, Adam N, Azmi Y, Jamarei O (2012) Determination of selected physical properties of egusi melon (Citrullus lanatus) seeds. J Basic Appl Sci 8:257–265
Barba de la Rosa AP, Fomsgaard IS, Laursen B, Mortensen AG, Olvera-Martínez L, Silva-Sánchez C, De León-Rodríguez A (2009) Amaranth (Amaranthus hypochondriacus) as an alternative crop for sustainable food production: phenolic acids and flavonoids with potential impact on its nutraceutical quality. J Cereal Sci 49(1):117–121. https://doi.org/10.1016/j.jcs.2008.07.012
Baricevic D, Bartol T (2000) The biological/pharmacological activity of Salvia genus V., Pharmacology. In: Kintzois SE (ed) Sage: The Genus Salvia. Harwood Academic Publishers, Abingdon Marston, pp 143–184
Bassil ES, Kaffka SR (2002) Respones of safflower (Carthamus tinctorius L.) to saline soil and irrigation II. Crop response to salinity. Agric Water Manag 54:81–92
Bazile D, Bertero D, Nieto C. 2015a. State of the Art report on Quinoa around the World in 2013. Rome: Food and Agriculture Organization of the United Nations (FAO) & CIRAD (Centredecoopérationinternationale en rechercheagronomiquepourledéveloppement).
Bazile D, Bertero D, Nieto C (2015b) State of the Art report on Quinoa around the World in 2013. Rome: Food and Agriculture Organization of the United Nations (FAO) & CIRAD (Centredecoopérationinternationaleenrechercheagronomiquepourledéveloppement)
Behboudian MH, Ma Q, Turner NC, Palta JA (2001) Reactions of chickpea to water stress: yield and seed composition. J Sci Food Agric 81(13):1288–1291. https://doi.org/10.1002/jsfa.939
Belko N, Cisse N, Diop NN, Zombre G, Thiaw S, Muranaka S, Ehlers J (2014) Selection for postflowering drought resistance in short-and medium-duration cowpeas using stress tolerance indices. Crop Sci 54:25–33
Bhargava A, Shukla S, Rajan S, Ohri D (2007a) Genetic variability and interrelationship among various morphological and quality traits in quinoa (Chenopodium quinoa Willd.). Field Crop Res 101:104–116. https://doi.org/10.1007/s10722-005-3011-0
Bhargava A, Shukla S, Rajan S, Ohri D (2007b) Genetic variability and interrelationship among various morphological and quality traits in quinoa (Chenopodium quinoa Willd.). Field Crop Res 101:104–116. https://doi.org/10.1007/s10722-005-3011-0
Bianchi FIJA, Booij CJH, Tscharntke T (2006) Sustainable pest regulation in agricultural landscapes: a review on landscape composition, biodiversity and natural pest control. Proc R Soc B 273:1715–1727. https://doi.org/10.1098/rspb.2006.3530
Bochicchio R, Philips TD, Lovelli S, Labella R, Galgano F, Di Marisco A, Amato M (2015) Innovative crop productions for healthy food: the case of chia (Salvia hispanica L.). In A. Vastola (Ed.), The sustainability of agro-food and natural resource systems in the Mediterranean Basin (pp. 29 45). Springer International Publishing
Brewbaker JL (2003) Corn Production in the tropics - The Hawaii Experience. College of Tropical Agriculture and Human Resources University of Hawaii at Manoa
Cabell JF, Oelofse M (2012) An indicator framework for assessing agroecosystem resilience. Ecol Soc 17(1)
Cahill JP (2003) Ethnobotany of chia, Salvia hispanica L. (Lamiaceae). Econ Bot 57:604–618
Camas N, Cirak C, Esendal E (2007) Seed yield, oil content and fatty acids composition of safflower (Carthamus tinctorius L.) grown in northern Turkey conditions. Anadolu Tarım Bilim Derg 22:98–104
Cannarozzi G, Plaza-Wuthrich S, Esfeld K, Larti S, Wilson YS, Girma D et al (2014) Genome and transcriptome sequencing identifies breeding targets in the orphan crop tef (Eragrostistef). BMC Genomics 15:581. https://doi.org/10.1186/1471-2164-15-581
Chandel G, Kumar M, Dubey M, Kumar M (2014) Nutritional properties of minor millets: neglected cereals with potentials to combat malnutrition. Curr Sci 107(7):1109–1111
Chandrasekara A, Shahidi F (2011) Determination of antioxidant activity in free and hydrolyzed fractions of millet grains and characterization of their phenolic profiles by HPLC-DAD-ESI-MSn. J Funct Foods 3:144–158
Changmei S, Dorothy J (2014) Millet-the frugal grain. Int J Sci Res Rev 3:75–90
Chappell MJ, LaValle LA (2011) Food security and biodiversity: can we have both? Agric Hum Values 23:3–26. https://doi.org/10.1007/S10460-009-9251-4
Chaudhari PP, Patel PT, Desai LJ (2009) Effect of nitrogen management on yield, water use and nutrient uptake of grain amaranth (Amaranthus hypochodriacus) under moisture stress. Indian J Agr 54(1):69-73
Chauhan BS, Abugho SB (2013) Effect of water stress on the growth and development of Amaranthus spinosus, Leptochloachinensis, and rice. Am J Plant Sci 4(05):989
Chauhan JS, Tyagi MK, Kumar A, Nashaat NI, Singh M, Singh NB, Welham SJ (2007) Drought effects on yield and its components in Indian mustard (Brassica juncea L.). Plant Breed 126(4):399–402
Chen HT, Liu XQ, Zhang HM, Yuan XX, Gu HP, Cui XY, Chen X (2018) Advances in salinity tolerance of soybean: genetic diversity, heredity, and gene identification contribute to improving salinity tolerance. J Integr Agric 17:2215–2221. https://doi.org/10.1016/S2095-3119(17)61864-1
Clay J (2004) World agriculture and the environment. Island Press, Washington, DC
Coates W (2011) Whole and ground chia (Salvia hispanica L.) seeds, chia oil-effects on plasma lipids and fatty acids. In: Patel V, Preedy R, Watson V (eds) Nuts and seeds in health and disease prevention. Academic, San Diego, pp 309–314
Cook BG, Pengelly BC, Brown SD, Donnelly JL, Eagles DA, Franco MA, Hanson J, Mullen BF, Partridge IJ, Peters M, Schultze-Kraft R (2005a) Tropical forages: an alternative selection tool. CSIRO, DPI&F (Qld), CIAT and ILRI, Brisbane
Cook BG, Pengelly BC, Brown SD, Donnelly JL, Eagles DA, Franco MA, Hanson J, Mullen BF, Partridge IJ, Peters M, Schultze-Kraft R (2005b) Tropical forages: an alternative selection tool. CSIRO, DPI&F (Qld), CIAT and ILRI, Brisbane
Cooper M, Gho C, Leafgren R, Tang T, Messina C (2014) Breeding drought-tolerant maize hybrids for the US corn-belt: discovery to product. J Exp Bot 65:6191–6204
Das IK, Rakshit S (2016) Millets, their importance, and production constraints. Biotic Stress Resistance in Millets:3–19. https://doi.org/10.1016/b978-0-12-804549-7.00001-9
DeClerk F, Le Coq JF, Rapidel B, Beer J (2012) Ecosystem services from agriculture and agroforestry. Measurement and Payment. Earthscan, London, Washington DC
De Falco B, Fiore A, Bochicchio R, Amato M, Lanzotti V (2018) Metabolomic analysis by UAE-GC MS and antioxidant activity of Salvia hispanica (L.) seeds grown under different irrigation regimes. Ind Crop Prod 112:584–592. https://doi.org/10.1016/j.indcrop.2017.12.030
De Giorgio D, Fornaro F (2004) Tillage systems for a sustainable growth of broad bean (Viciafaba L., major) in a semiarid region of Southern Italy. In: ISCO 2004—13th International Soil Conservation Organization Conference. Conserving Soil and Water for Society: Sharing Solutions. Brisbane, July 2004, (paper no. 934), pp. 1–4
De Schutter O (2010) Report submitted by the Special Rapporteur on the right to food. Human Rights Council 16th Session. United Nations General Assembly, United Nations, New York
Devi PB, Vijayabharathi R, Sathyabama S, Malleshi NG, Priyadarisini VB (2014) Health benefits of finger millet (Eleusine coracana L.) polyphenols and dietary fiber: a review. J Food Sci Technol 51(6):1021–1040
Dost M (2015a) Field evaluation results across locations and identification of suitable quinoa varieties. In: Wrap upWorkshop of Regional Quinoa Project (TCP/RAB/3403–FAO). Food and Agriculture Organization of the United Nations, Rome
Dost M (2015b) Field evaluation results across locations and identification of suitable quinoa varieties. In: Wrap upWorkshop of Regional Quinoa Project (TCP/RAB/3403–FAO). Food and Agriculture Organization of the United Nations, Rome
Duvick DN (1999) Heterosis: feeding people and protecting natural resources. In J. G. Coors & S. Pandey (Eds.), The Genetics and Exploitation of Heterosis in Crops (pp. 19-29). Madison, WI: American Society of Agronomy, Inc., Crop Science Society of America, Inc., Soil Science Society of America, Inc. Retrieved from http://www.biotechinfo.net/heterosis_duvick1.pdf
Dykes L, Rooney LW (2006) Review sorghum and millet phenols and antioxidants. J. Cereal Sci. 44:236-251
Ecoport (2010) Ecoport database. http://www.ecoport.org
Ejigu A, Asfaw A, Licence P (2010) Moringa stenopetala seed oil as a potential feedstock for biodiesel production in Ethiopia, 316-320
El-Jasser ASH (2011) Chemical and biological properties of local cowpea seed protein grown in Gizan region. International Scholarly and Scientific Research & Innovations 5:68–75
Ellmer F (2008) Soil organic matter of a sandy soil influenced by agronomy and climate
Elshibli S, Korpelainen H (2008a) Microsatellite markers reveal high genetic diversity in date palm (Phoenix dactylifera L.) germplasm from Sudan. Genet 134:251–260
Elshibli S, Korpelainen H (2008b) Excess heterozygosity and scarce genetic differentiation in the population of Phoenix dactylifera L: human impact or ecological determinants. Plant Genetic Resources:Characterization and Utilization 7:95–104
Estep M, Van Mourik T, Muth P, Guindo D, Parzies H, Koita O, Weltzein E, Bennetzen J (2011) Genetic Diversity of a Parasitic Weed, Striga hermonthica, on Sorghum and Pearl Millet in Mali. Trop Plant Biol 4:2,91–2,98
Fageria NK (2011) Growth and mineral nutrition of field crops. CRC Press Taylor & Francis Group, Boca Raton
Fahmy AA, Youssef KM, El Shae HM (2010) Intake and nutritive value of some salt-tolerant fodder grasses for sheep under saline conditions of South Sinai, Egypt. Small Rumin Res 91:110–115. https://doi.org/10.1016/j.smallrumres.2009.11.023
FAO (2001) The State of Food Insecurity in the World. FAO, Rome
FAO (2003) Food and Agriculture Organization. http://apps.fao.org
FAO (2009) The State of Food Insecurity in the World. FAO, Rome, Rome
FAO (2011a) Why Has Africa Become a Net Food Importer? Explaining Africa Agricultural and Food Trade Deficits. Trade and markets division, Food and Agriculture Organization of the United Nations, Rome
FAO (2011b) The State of Food Insecurity of the World: How does international price volatility affect domestic economies and food security? FAO, WFP, IFAD, Rome
FAO (Food and Agriculture Organization) (2003) http://apps.fao.org.
FAOSTAT (2012) Available from FAOSTAT Web site:http://faostat.fao.org/
FAOSTAT F (2014) Food and Agriculture Organization statistical database
Flowers TJ, Gracia A, Koyama M, Yeo AR (1997) Breeding for salt tolerance in
Flowers TJ, Gaur PM, Gowda CLL, Krishnamurthy L, Srinivasan S, Siddique KHM, Turner NC, Vadez V, Varshney RK, Colmer TD (2010) Salt sensitivity in chickpea. Plant Cell Environ 33:490–509
Fraj MB, Al-Dakheel AJ, McCann IR, Shabbir GM, Rumman GA, Al Gailani AM (2013a) Selection of high yielding and stable safflower (Carthamus tinctorius L.) genotypes under salinity stress. Agric Sci Res J 3:273–283
Fraj MB, Al-Dakheel AJ, McCann IR, Shabbir GM, Rumman GA, Al GA (2013b) Selection of high yielding and stable safflower (Carthamus tinctorius L.) genotypes under salinity stress. Agric Sci Res J 3:273–283
Fraj MB, Al-Dakheel AJ, McCann IR, Shabbir GM, Rumman GA, Al Gailani AM (2013c) Selection of high yielding and stable safflower (Carthamustinctorius L.) genotypes under salinity stress. Agric Sci Res J 3:273–283
Francois LE, Bernstein L (1964) Salt tolerance of safflower. Agron J 56:38–40
Frederick JR, Camp CR, Bauer PJ (2001) Drought-stress effects on branch and mainstem seed yield and yield components of determinate soybean. Crop Sci 41(3):759–763
French RJ (1998) Effect of early water deficit on growth and development of faba bean. Proceeding of the Australian Agronomy Conference. (C.F. http://www.regional.org.au/au/asa)
Furr JR, Ream CL (1968) Salinity effects on growth and salt uptake of seedlings of the date, Phoenix dactylifera L. Proc Amer Soc Hort Sci 92:268–273
Garí JA (2002) Review of the African millet diversity. International workshop on fonio, food security and livelihood among the rural poor in West Africa. IPGRI/IFAD, Bamako, Mali, 19-22 November 2001
Gill RK, Sharma AD, Singh P, Bhulla SS (2001) Effect of various abiotic stresses on the growth, soluble sugars and water relations of sorghum seedlings grown in light and darkness. Bulg J Plant Physiol 27(1–2):72–84
Giunta F, Motzo R, Deidda M (1993) Effect of drought on yield and yield components of durum wheat and triticale in a Mediterranean environment. Field Crop Res 33(4):399–409
González A, Ayerbe L (2010) Effect of terminal water stress on leaf epicuticular wax load, residual transpiration and grain yield in barley. Euphytica 172(3):341–349
González A, Martı́n, I. and Ayerbe, L. (1999) Barley yield in water-stress conditions.: the influence of precocity, osmotic adjustment and stomatal conductance. Field Crop Res 62(1):23–34
González A, Martín I, Ayerbe L (2008) Yield and osmotic adjustment capacity of barley under terminal water-stress conditions. J Agron Crop Sci 194(2):81–91. https://doi.org/10.1111/j.1439-037x.2007.00289.x
Gurudeeban S, Ramanathan T (2010) Antidiabetic effect of Citrullus colocynthis in alloxon-induced diabetic rats. Inventi Rapid: Ethno pharmacology 1:112
Gurudeeban S, Rajamanickam E, Ramanathan T, Satyavani K (2010) Antimicrobial activity of Citrullus colocynthis in gulf of Mannar. Int J Curr Res 2:78–81
Haddadin MF (2015) Assessment of drought tolerant barley varieties under water stress. 5(2):131–137
Hammami Z, Gauffreteau A, BelhajFraj M, Sahli A, Jeuffroy MH, Rezgui S, Bergaoui K, McDonnell R, Trifa Y (2017) Predicting yield reduction in improved barley (Hordeum vulgare L.) varieties and landraces under salinity using selected tolerance traits. Field Crop Res 211:10–18
Han X, Cheng L, Zhang R, Bi J (2009) Extraction of safflower seed oil by supercritical CO2. J Food Eng 92:370–376
Harada H, Yoshimura Y, Sunaga Y, Hatanaka T (2000) Variations in nitrogen uptake and nitrate-nitrogen concentration among sorghum groups. Soil Sci. Plant Nutr 46:97-104.
Harris BN, Sadras VO, Tester M (2010) A water-centred framework to assess the effects of salinity on the growth and yield of wheat and barley. Plant Soil 336(1-2):377–389
Hauggaard-Nielsen H, Jørnsgård B, Kinane J, Jensen ES (2008) Grain legume— cereal intercropping: the practical application of diversity, competition and facilitation in arable and organic cropping systems. Renewable Agric. Food Syst 23(1):3–12
He J, Jin Y, Du YL, Wang T, Turner NC, Yang RP, Siddique KH, Li FM (2017) Genotypic variation in yield, yield components, root morphology and architecture, in soybean in relation to water and phosphorus supply. Front Plant Sci 8:1499
Heuer B, Yaniv Z, Ravina I (2002) Effect of late salinization of chia (Salvia hispanica), stock (Matthiolatricuspidata) and evening primrose (Oenothera biennis) on their oil content and quality. Ind Crop Prod 15(163):167
Hirich A, Choukr-Allah R, Jelloul A, Jacobsen SE (2014a) Quinoa (Chenopodium quinoa willd.) Seedling, water uptake and yield responses to irrigation water salinity. Acta Hortic 1054:145–152. https://doi.org/10.17660/ActaHortic.2014.1054.16
Hirich A, Jelloul A, Choukr-Allah R, Jacobsen SE (2014b) Saline water irrigation of quinoa and chickpea: seedling rate, stomatal conductance and yield responses. J Agron Crop Sci 200(5):378–389
Hithamani G, Srinivasan K (2014) Effect of domestic processing on the polyphenol content and bioaccessibility in finger millet (Eleusine coracana) and pearl millet (Pennisetum glaucum). Food Chemistry 164:55–62
Hojati M, Modarres-Sanavy SAM, Karimi M, Ghanati F (2011) Responses ofgrowth and antioxidant systems in Carthamustinctorius L. under water deficitstress. Acta Physiol Plant 33:105–112
Hussain MI, Al-Dakheel AJ (2015) Using alternate water resources for cultivation of salt tolerant perennial grasses under marginal environment. TROPENTAG, Management of Land use systems for enhanced food security-conflicts, controversies and resolutions, Berlin, Germany, September 16–18
Hussain MI, Al-Dakheel AJ (2018) Effect of salinity stress on phenotypic plasticity, yield stability, and signature of stable isotopes of carbon and nitrogen in safflower. Environ Sci Pollut Res 25(24):23685–23694. https://doi.org/10.1007/s11356-018-2442-z
Hussain MI, Lyra DA, Nikos N, Farooq M. Ahmad N (2015) Salt and drought stresses in safflower: a review. Agron Sustain Dev 36:1.
Hussain MI, Lyra DA, Farooq M, Nikoloudakis N, Khalid N (2016) Salt and drought stresses in safflower: a review. Agron Sustain Dev 36:4
Hussain MI, Muscolo A, Farooq M, Ahmad W (2019) Sustainable use and management of non-conventional water resources for rehabilitation of marginal lands in arid and semiarid environments. Agric. Water Manag. 221:462–476.
ICBA (2013) Biosalinity News. Newsletter for International Center for Biosaline Agriculture 14:3
ICBA (2015) ICBA Annual Report 2015. International Center for Biosaline Agriculture, Dubai
IPCC (2001) Climate Change 2001: the scientific basis. In: Houghton JT, Ding Y, Griggs DJ, Noguer M, Van der Linden PJ, Dai X, Maskell K, Johnson CA (eds) Contribution of Working Group I to the Third Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, 881 pp
IPCC (2014) Climate Change 2014: impacts, adaptation, and vulnerability. Part A: Global and Sectoral Aspects. In: Field CB, Barros VR, Dokken DJ, Mach KJ, Mastrandrea MD, Bilir TE, Chatterjee M, Ebi KL, Estrada YO, Genova RC, Girma B, Kissel ES, Levy AN, MacCracken S, Mastrandrea PR, White LL (eds) Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, 1132 pp
Incekaya Ç, Yazar A (2016) SALTMED model performance for quinoa irrigated with fresh and saline water in a Mediterranean environment. Irrig Drain 65(1):29–37
Ishiyaku MF, Aliyu H (2013) Field evaluation of cowpea genotypes for drought tolerance and striga resistance in the dry Savanna of the North-West Nigeria. Int J Plant Breed Genet 7:47–56
Istanbulluoglu A (2009) Effects of irrigation regimes on yield and water productivity of safflower (Carthamustinctorius L.) under Mediterranean climatic conditions. Agric Water Manag 96(12):1792–1798
Istanbulluoglu A, Gocmen E, Gezer E, Pasa C, Konukcu F (2009) Effects of water stress at different development stages on yield and water productivity of winter and summer safflower (Carthamustinctorius L.). Agric Water Manag 96(10):1429–1434. https://doi.org/10.1016/j.agwat.2009.04.00
Jacobsen SE (2003a) The worldwide potential for quinoa (Chenopodium quinoa Willd.). Food Rev Int 19:167–177. https://doi.org/10.1081/FRI-120018883
Jacobsen SE (2003b) The worldwide potential for quinoa (Chenopodium quinoa Willd.). Food Rev Int 19:167–177. https://doi.org/10.1081/FRI-120018883
Jaradat AA, Shahid M, Al Maskri AY (2004a) Genetic diversity in the Batini barley landrace from Oman: I. Spike and seed quantitative and qualitative traits. Crop Sci 44:304–315
Jaradat AA, Shahid M, Al Maskri AY (2004b) Genetic diversity in the Batini barley landrace from Oman: I. Spike and seed quantitative and qualitative traits. Crop Science 44:304–315
Jensen ES, Peoples MB, Hauggaard-Nielsen H. (2010) Faba bean in cropping systems. Field Crop Res 115(3):203–216. https://doi.org/10.1016/j.fcr.2009.10.008
Kamrin MA (1997) Pesticide profiles: toxicity, environmental impact, and fate. CRC Press LLC, Boca Raton
Kar G, Kumar A, Martha M (2007a) Water use efficiency and crop coefficients of dry season oilseed crops. Agric Water Manag 87:73–82. https://doi.org/10.1016/j.agwat.2006.06.002
Kar G, Kumar A, Martha M (2007b) Water use efficiency and crop coefficients of dry season oilseed crops. Agric Water Manage 87:73–82. https://doi.org/10.1016/j.agwat.2006.06.002
Karadge BA, Chavan PD (1983) Physiological studies in salinity tolerance of Sesbania aculeata POIR. Biol Plant 25:412–418. https://doi.org/10.1007/BF02903138
Katerji N, Mastrorilli M, Lahmer FZ, Maalouf F, Oweis T (2011) Faba bean productivity in saline–drought conditions. Eur J Agron 35(1):2–12
Kausar A, Ashraf MYM, Niaz M (2014) Some physiological and genetic determinants of salt tolerance in Sorghum (Sorghum Bicolor (L.) Moench): Biomass production and nitrogen metabolism. Pak J Bot 46(2):515–519
Kaya MD (2009a) The role of hull in germination and salinity tolerance in some sunflower (Helianthus annuus L.) cultivars. Afr J Biotechnol 8(4):597–600
Kaya MD (2009b) The role of hull in germination and salinity tolerance in some sunflower (Helianthus annuus L.) cultivars. Afr J Biotechnol 8:597–600
Kaydan D, Yagmur M (2008) Germination, seedling growth and relative water content of shoot in different seed sizes of triticale under osmotic stress of water and NaCl. Afr. J. Biotechnol 7:2862
Ketema S (1993) Phenotypic variations in tef (Eragrostistef) germplasm- morphological and agronomic traits. A catalon technical manual No. 6. Institute of Agricultural Research. Addis Ababa, Ethiopia
Khalid N, Khan RS, Hussain MI, Farooq M, Ahmad A, Ahmad I (2017) A comprehensive characterisation of safflower oil for its potential applications as a bioactive food ingredient-a review. Trends Food Sci Technol 66:176–186
Khan MS, Zaidi A, Ahemad M, Oves M, Wani PA (2010) Plant growth promotion by phosphate solubilizing fungi–current perspective. Arch. Agron. Soil Sci. 56(1):73–98
Khan ZR, Pickett JA, Berg JVD, Wadhams LJ, Woodcock CM (2000) Exploiting chemical ecology and species diversity: stem borer and Striga control for maize and sorghum in Africa. Pest Manag Sci 56(11):957–962
Kijne JW (2005) Towards a strategy for feasible investment in drainage for Aral Sea basin. GRID - Magazine of the IPTRID Network (FAO/United Kingdom), pp 13-15
Kothari SL, Kumar S, Vishnoi RK, Kothari A, Watanabe KN (2005) Applications of biotechnology for improvement of millet crops: review of progress and future prospects. Plant Biotechnol 22:81–88. https://doi.org/10.5511/plantbiotechnology.22.81
Krishnamurthy L, Serraj R, Hash CT, Dakheel AJ, Reddy BVS (2007) Screening sorghum genotypes for salinity tolerant biomass production. Euphytica 156:15–24. https://doi.org/10.1007/s10681-006-9343-9
Kumar G, Purty RS, Sharma MP, Singla-Pareek SL, Pareek A (2009) Physiological responses among Brassica species under salinity stress show strong correlation with transcript abundance for SOS pathway-related genes. J. Plant Physiol. 166: 507–520.Kurup S.S., Hedar Y., Al Daheri M., et al. (2009). Morpho-physiological evaluation and RAPD marker assisted characterization of date palm varieties for salinity tolerance. J Food Agric Environ 7:503–507
Kumari R, Vishnuvardhan Z, Babu K (2013) A study on effect of NaCl stress on Kodomillet (Paspalum scrobiculatum) during germination stage. Ann. Plant Sci 2:388–394
Landis DA, Gardiner MM, van der Werf W, Swinton SM (2008) Increasing corn for biofuel production reduces biocontrol services in agricultural landscapes. P Natl Acad Sci 105(51):20552-20557
Leder I (2004) Sorghum and Millets. Cultivated plants, primarily as food sources. In: Gyargy F (ed) Encyclopedia of life support systems. UNESCO, Eolss Publishers, Oxford
Leport L, Turner NC, Davies SL, Siddique KHM (2006) Variation in pod production and abortion among chickpea cultivars under terminal drought. Eur J Agron 24(3):236–246
Lirong B, Liran S, Xiaoli G, Xiaona Z. 2016. Effects of water stress on physiological characteristics of different genotypes of triticale and rye seedlings. Crops 2016-04.
Lisa LC, Alderman H, Aduayom D (2006) Food insecurity in sub-Saharan Africa: new estimates from household expenditure surveys. Research Report, 146. Volume Research Report. Washington, DC: 146, International Food Policy Research Institute (IFPRI).
Liu F, Stützel H (2002) Leaf water relations of vegetable amaranth (Amaranthus spp.) in response to soil drying. Eur J Agron 16:137–150
Liu F, Stützel H (2004a) Biomass partitioning, specific leaf area, and water use efficiency of vegetable amaranth (Amaranthus spp.) in response to drought stress. Scientia Horticulturae 102:15–27. https://doi.org/10.1016/j.scienta.2003.11.014
Liu F, Stützel H (2004b) Biomass partitioning, specific leaf area, and water use efficiency of vegetable amaranth (Amaranthus spp.) in response to drought stress. Scientia Horticulturae 102:15–27. https://doi.org/10.1016/j.scienta.2003.11.014
Lombani M, Arzani A (2011) Morpho-physiological traits associated with terminal drought-stress tolerance in triticale and wheat. Agron Res 9(1–2):315–329
Lovelli S, Perniola M, Ferrara A, Di Tommaso T (2007) Yield response factor towater (Ky) and water use efficiency of Carthamustinctorius L. and Solanum melongena L. Agric. Water Manage 92:73–80
Lyu C, Xu Z (2020) Crop production changes and the impact of Grain for Green program in the Loess Plateau of China. J Arid Land:1–11
Maas EV (1990) Crop salt tolerance. In: Agricultural Salinity Assessment and Management Ed. K K Tanji, ASCEManuals & Reports on Engineering No. 71, pp. 262–304 ASCE. New York
Maas EV, Grattan SR (1999) Crop yields as affected by salinity. In: Skaggs RW, van Schilfgaarde J (eds) Agricultural Drainage. ASA-CSSA-SSSA, Madison, pp 55–108
Madamba R, Grubben GJH, Asante IK, Akromah R (2006a) Vigna unguiculata (L.) Walp. Record from Protabase. Brink M & Belay G (Editors). PROTA (Plant Resources of Tropical Africa / Ressources végétales de l’Afrique tropicale), Wageningen, Netherlands
Madamba R, Grubben GJH, Asante IK, Akromah R (2006b) Vigna unguiculata (L.) Walp. Record from Protabase. Brink M & Belay G (Editors). PROTA (Plant Resources of Tropical Africa / Ressourcesvégétales de l’Afriquetropicale), Wageningen, Netherlands
Mahmood K (2011) Salinity tolerance in barley (Hordeum vulgare L.): effects of varying NaCl, K?/Na? and NaHCO3 levels on cultivars differing in tolerance. Pak J Bot 43(3):1651–1654
Malash N, Flowers T, Ragab R (2008) Effect of irrigation methods, manage ment and salinity of irrigation water on tomato yield, soil moisture and salinity distribution. Irrig.Sci. 26:313–323. https://doi.org/10.1007/s00271-007-0095-7
Maleki P, Bahrami HA, Saadat S, Sharifi F, Dehghany F, Salehi M (2018) Salinity threshold value of Quinoa (Chenopodium quinoa Willd.) at various growth stages and the appropriate irrigation method by saline water. Commun Soil Sci Plant Anal 49(15):1815–1825. https://doi.org/10.1080/00103624.2018.1474917
Malik KA, Aslam Z, Naqvi M (1986) Kallar grass: a plant for saline land. Nuclear Institute for Agriculture and Biology, Faisalabad
Mann JA, Kimber CT, Miller FR (1983) The origin and early cultivation of sorghums in Africa. Texas Agricultural Experiment Station, Texas A&M University System, USA., Pages: 21
Mansour E, Abdul-Hamid MI, Yasin MT, Qabil N, Attia A (2017) Identifying drought-tolerant genotypes of barley and their responses to various irrigation levels in a Mediterranean environment. Agric Water Manag 194:58–67
Masuda T, Goldsmith PD (2009) World soybean production: area harvested, yield, and longterm projections. Int Food Agribus Manag Rev 12:143–161
Mathur Y, Poonia M, Pandel U, Singh R (2012) Performance and emission characteristics of diesel engine using low concentration thumba oil diesel blends. Int J Wind Renew Energy 1:108–113
Menon K, Sood N, Rao NK (2016) Study of morpho-agronomic diversity and oil content in desert gourd (Citrullus colocynthis (L.) Schrad.). Aust J Crop Sci 10:1000–1006
Methu JN, Kiruiro EM, Abat N (2006) Your feed shortage problems. Use maize forage. KARI resource center, Nairobi
Meyer LD, Dabney SM, Murphree CE, Harmon WC, Grissinger EH (1999) Crop production systems to control erosion and reduce runoff from upland silty soils. Trans ASAE 42(6):1645–1652
Miller PR, Waddington J, McDonald CL, Derksen DA (2002) Cropping sequence affects wheat productivity on the semiarid northern Great Plains. Can J Plant Sci 82:307–318
Miransari M, Smith DL (2007) Overcoming the stressful effects of salinity and acidity on soybean nodulation and yields using signal molecule Genistein under field conditions. J Plant Nutr 30(12):1967–1992
Modi AT, Mabhaudhi T (2013) Water use and drought tolerance of selected traditional and indigenous crops. Water Research Commision of South Africa: Pretoria, South Africa
Movahhedy-Dehnavy M, Modarres-Sanavy SAM, Mokhtassi-Bidgoli A (2009) Foliar application of zinc and manganese improves seed yield and quality of safflower (Carthamustinctorius L.) grown under water deficit stress. Ind Crop Prod 30(1):82–92. https://doi.org/10.1016/j.indcrop.2009.02.004
Mposi MS (1999a) Vegetable amaranth improvement for South Africa: a preliminary investigation. The Australian New Crops Newsletter 11:8–14
Mposi MS (1999b) Vegetable amaranth improvement for South Africa: a preliminary investigation. Australian New Crops Newslett 11:8–14
Mullen CL, Holland JF, Heuke L (2003a) AGFACTS: Cowpea, lablab and pigeon pea. Available online: http://www.dpi.nsw.gov.au/_data/assets/pdf_file/0006/157 488/cowpea-lablab-pigeon-pea.pdf
Mullen CL, Holland JF, Heuke L. (2003b) AGFACTS: Cowpea, lablab and pigeon pea. Available online: http://www.dpi.nsw.gov.au/_data/assets/pdf_file/0006/157 488/cowpea-lablab-pigeon-pea.pdf
Munns R, Tester M (2008) Mechanismsofsalinitytolerance. AnnuRev PlantBiol 59:651–681. https://doi.org/10.1146/annurev.arplant.59.032607.092911
Muñoz LA, Cobos A, Diaz O, Aguilera JM (2013) Chia seed (Salvia hispanica): an ancient grain and a new functional food. Food Rev Int 29(394):408
Murty MVR, Piara Singh, Wani SP, Khairwal IS, Srinivas K (2007) Yield gap analysis of sorghum and pearl millet in India using simulation modeling. Global Theme on Agroecosystems Report no. 37. International Crops Research Institute for the Semi-Arid Tropics, Patancheru 502324, Andhra Pradesh, India, p 82
Newton AC, Akar T, Baresel JP, Bebeli PJ, Bettencourt E, Bladenopoulos KV, Czembor JH, Fasoula DA, Katsiotis A, Koutis K, Koutsika-Sotiriou M (2011) Cereal landraces for sustainable agriculture. In Sustainable Agriculture Volume 2 (pp. 147-186). Springer, Dordrecht
Nixon DJ, Stephens W, Tyrrel SF, Brierley ED (2001) The potential for short rotation energy forestry on restored landfill caps. Bioresource Technology 77:237–245. https://doi.org/10.1016/S0960-8524(00)00081-X
Nouman W, Basra SMA, Yasmeen A, Gull T, Hussain SB, Zubair M, Gul R (2014) Seed priming improves the emergence potential, growth and antioxidant system of Moringa oleifera under saline conditions. Plant Growth Regul. 73:267–278
O'Donnell NH, Lindberg BM, Neale AD, Hamilla JD, Blomstedt CK, Roslyn MG (2013) Effects of PEG-induced osmotic stress on growth and dhurrin levels of forage sorghum. Plant Physiol 73:83–92
Omami EN, Hammes PS (2006) Interactive effects of salinity and water stress on growth, leaf water relations, and gas exchange in amaranth (Amaranthusspp.). N Z J Crop Hortic Sci 34(1):33–44
Oplinger ES, Oelke EA, Putnam DH, Kelling, KA, Kaminski AR, Teynor TM, Doll JD, Durgan BR (1991) Mustard. In: Alternative Field Crops Manual, University of Wisconsin-Exension, Cooperative Extension. https://www.hort.purdue.edu/newcrop/afcm/mustard.html
Orona-Tamayo D, Valverde ME, Paredes-López O (2017) Chia—The New Golden Seed for the 21st Century. Sustainable Protein Sources, 265–281
Pakar N, Pirasteh-Anosheh H, Emam Y, Pessarakli M (2016) Barley growth, yield, antioxidant enzymes and ions accumulation affected by Pgrs under salinity stress. J Plant Nutr 39:1372–1379. https://doi.org/10.1080/01904167.2016.1143498
Pal A, Kachhwaha SS, Maji S, Babu MKG (2010) Thumba (Citrullus colocynthis) seed oil: a sustainable source of renewable energy for biodiesel production. J Sci Ind Res 69:384–389
Palada M, Chang L (2003a) Suggested cultural practices for vegetable amaranth. The World Vegetable Centre (AVRDC), International Cooperators’ Fact Sheet: Shanhua, Taiwan. 3–552
Palada M, Chang L (2003b) Suggested cultural practices for vegetable amaranth. The World Vegetable Centre (AVRDC), International Cooperators’ Fact Sheet: Shanhua, Taiwan. 3–552.
Papastylianou I, Puckridge DW, Carter ED (1981) Nitrogen nutrition of cereals in a short-term rotation. I Single season treatments as a source of nitrogen for subsequent cereal crops. Aust J Agric Res 32(5):703–712
Peperkamp M (2014) Chia from Bolivia: a modern super seed in a classic pork cycle. CBI Marked Intelligence (pp. 1 15). Netherland: Ministry of Foreign Affairs. ,www.cbi.eu/market-information/grains-pulses/chia/
Phalan B, Green R, Balmford A (2014) Closing yield gaps: perils and possibilities for biodiversity conservation. Philosophical Trans Royal Soc B: Biol Sci 369:20120285
Pitman MG, Läuchli A (2002)“Globalimpactofsalinityandagricultural ecosystems,”in Salinity:Environment—Plants—Molecules, edsA.LäuchliandU. Lüttge (Dordrecht:Kluwer),3–20
Pontieri P, Troisi J, Fiore RD, Bean SR, Roemer E, Boffa A, Giudice AD, Pizzolante G, Alifano P, Giudice LD (2014) Mineral contents in grains of seven food grade sorghum hybrids grown in Mediterranean environment. J Crop Sci 8(11):1550–1559
Potter L (2016) How can the people’s sovereignty be achieved in the oil palm sector? Is the plantation model shifting in favour of smallholders. Land and development in Indonesia. Searching for the people’s sovereignty 315–342
Pray C E, Nagaraja L (2009) Pearl millet and sorghum improvement in India (Vol. 919). Intl Food Policy Res Inst
Pulvento C, Riccardi M, Lavini A, D'andria R, Ragab R (2013) SALTMED model to simulate yield and dry matter for quinoa crop and soil moisture content under different irrigation strategies in south Italy. Irrig Drain 62(2):229–238
Qadir M, Tubeileh A, Akhtar J, Larbi A, Minhas PS, Khan MA (2008) Productivity enhancement of salt-affected environments through crop diversification. Land Degrad Dev 19:429–453. https://doi.org/10.1002/ldr.853
Qasim M, Gulzar S, Khan MA (2011) Halophytes as medicinal plants. In: Ozturk M, Mermut AR, Celik A (eds) Urbanization, land use, land degradation and environment. Daya Publishing House, Turkey
Qiu L, Wu DZ, Ali S, Cai SG, Dai F, Jin X, Wu F, Zhang G (2011) Evaluation of salinity tolerance and analysis of allelic function of HvHKT1 and HvHKT2 in Tibetan wild barley. Theor Appl Genet 122:695–703. https://doi.org/10.1007/s00122-010-1479-2
Radhouane L (2007) Response of Tunisian autochthonous pearl millet (Pennisetum glaucum (L.) R. Br.) to drought stress induced by polyethylene glycol (PEG) 6000. Afr J Biotechnol 6(9):1102–1105
Ragnarsdottir KV (2000) Environmental fate and toxicology of organophosphate pesticides. J Geol Soc 157:859–876
Rahman MM, Mostafiz SB, Paatero JV, Lahdelma R (2014) Extension of energy crops on surplus agricultural lands: a potentially viable option in developing countries while fossil fuel reserves are diminishing. Renew Sustain Energy Rev 29:108–119
Raimondi G, Rouphael Y, Di Stasio E, Napolitano F, Clemente G, Maiello R et al (2017) Evaluation of Salvia hispanica performance under increasing salt stress conditions. Acta Hortic 1170:703–708
Rajamanickam E, Gurudeeban S, Ramanathan T, Satyavani K (2010) Evaluation of anti inflammatory activity of Citrullus colocynthis. Int J Curr Res 2:67–69
Ramoliya PJ, Pandey AN (2003) Soil salinity and water status affect growth of Phoenix dactylifera seedlings. New Zealand Journal of Crop and Horticultural Science 31(4):345–353
Rao NK, Shahid M (2012a) Quinoa–a promising new crop for the Arabian Peninsula. Am-Eurasian J Agric Environ Sci 12:1350–1355. https://doi.org/10.5829/idosi.aejaes.2012.12.10.1823
Rao NK, Shahid M (2012b) Quinoa–a promising new crop for the Arabian Peninsula. Am-Eurasian J Agric Environ Sci 12:1350–1355. https://doi.org/10.5829/idosi.aejaes.2012.12.10.1823
Rao NK, Shahid M (2012c) Quinoa–a promising new crop for the Arabian Peninsula. American-EurasianJ AgricEnvironSci 12:1350–1355
Rao NK, Rahman KU, Shoaib. (2013a) Quinoa – prospects as an alternative crop for salt-affected areas. 3rd International Conference on Neglected and Under-utilized Species, 25-27 September 2013, At Accra, Ghana
Rao NK, Rahman KU, Shoaib (2013b) Quinoa – prospects as an alternative crop for salt-affected areas. 3rd International Conference on Neglected and Under-utilized Species, 25-27 September 2013, At Accra, Ghana
Rao PS, Kumar CG, Prakasham RS, Rao AU, Reddy BVS (2015) Sweet sorghum: breeding and bioproducts. In: Cruz VMV, Dierig DA (eds) Industrial Crops 9: 1-28. Springer, New York. https://doi.org/10.1007/978-1-4939-1447-0_1
Razzaghi F, Ahmadi SH, Adolf VI, Jensen CR, Jacobsen SE, Andersen MN (2011) Water relations and transpiration of quinoa (Chenopodium quinoa Willd.) under salinity and soildrying. J Agron Crop Sci 197:348–360
Repo-Carrasco R, Espinoza C, Jacobsen SE (2003a) Nutritional value and use of the Andean crops quinoa (Chenopodium quinoa) and Kañiwa (Chenopodium pallidicaule). Food Rev Int 19:179–189. https://doi.org/10.1081/FRI-120018884
Repo-Carrasco R, Espinoza C, Jacobsen SE (2003b) Nutritional value and use of the Andean crops quinoa (Chenopodium quinoa) and Kañiwa (Chenopodium pallidicaule). Food Rev Int 19:179–189. https://doi.org/10.1081/FRI-120018884
Robert JW (1986) The soybean solution: meeting world food needs. NIT-College of Agriculture, University of Illinois at Urbana, Champaign, USA, 1 Bulletin, pp. 4–27
Rodríguez JP, Rahman H, Thushar S, Singh RK (2020a) Healthy and resilient cereals and pseudo-cereals for marginal agriculture: molecular advances for improving nutrient bioavailability. Front Genet 11
Rodríguez JP, Rahman H, Thushar S, Singh RK (2020b) Healthy and resilient cereals and pseudo-cereals for marginal agriculture: molecular advances for improving nutrient bioavailability. Front Genet, 11
Rodríguez-Navarro DN, Oliver IM, Contreras MA, Ruiz-Sainz JE (2011) Soybean interactions with soil microbes, agronomical and molecular aspects. Agron Sustain Dev 31:173–190
Roy RC, Sagar A, Tajkia JE, Razzak MDA, Zakir Hossain AKM (2018) Effect of salt stress on growth of sorghum germplasms at vegetative stage. J Bangladesh Agril Univ 16(1):67–72
Royo C, Abaza M, Blanco R, García del Moral LF (2000) Triticale grain growth and morphometry as affected by drought stress, late sowing and simulated drought stress. Funct Plant Biol 27(11):1051–1059
Saade S, Maurer A, Shahid M, Oakey H, Schmöckel SM, Negrão S, Pillen K, Tester M (2016) Yield-related salinity tolerance traits identified in a nested association mapping (NAM) population of wild barley. Sci Rep 6:32586
Sadeghipour O, Abbasi S (2012) Soybean response to drought and seed inoculation. World Appl. Sci J 17:55–60
Sailaja K, Sujatha B (2013) Impact of salt stress (NaCl) on pigments, phenols and flavonoids in C4 (sorghum bicolor) and C3 (Oryza sativa) cultivars. Int J Biol Pharm Res 4(5):361–367
Salaheldeen M, Aroua MK, Mariod AA, Cheng SF, Abdelrahman MA (2014) An evaluation of Moringa peregrina seeds as a source for biofuel. Ind. Crops Prod 61:49–61
Salaheldeen M, Aroua MK, Mariod AA, Cheng SF, Abdelrahman MA, Atabani AE (2015) Physicochemical characterization and thermal behavior of biodiesel and biodiesel–diesel blends derived from crude Moringa peregrina seed oil. Energy Convers. Manage. 92:535–542
Salehi M, Arzani A (2013) Grain quality traits in triticale influenced by field salinity stress. AJCS 7(5):580–587
Salehi M, Arzani A (2014) Evaluation of triticale genotypes for salt tolerance using physiological traits 2014. Emir J Food Agric 26(3):277–283
Samarah NH (2005) Effects of drought stress on growth and yield of barley. Agron Sustain 25(1):145–149
Samarah NH, Alqudah AM, Amayreh JA, McAndrews GM (2009) The effect of late-terminal drought stress on yield components of four barley cultivars. J Agron Crop Sci 195(6):427–441. https://doi.org/10.1111/j.1439-037x.2009.00387
Santos RF, Bassegio, D, de Almeida Silva M (2017) Productivity and production components of safflower genotypes affected by irrigation at phenological stages. Agric. Water Manag. 186:66–74
Sattar H, Mehrazi M, Awartani M, Awad AR (2002) Alfalfa crop water requirement study in the United Arab Emirates. Emirates J Agric Res 4:12–24
Schilling RK, Marschner P, Shavrukov Y, Berger B, Tester M, Roy SJ, Plett DC (2014) Expression of the Arabidopsis vacuolar H+-pyrophosphatase gene (AVP 1) improves the shoot biomass of transgenic barley and increases grain yield in a saline field. Plant Biotechnol J 12(3):378–386
Setter T, Waters I (2003) Review of prospects for germplasm improvement for waterlogging tolerance in wheat, barley and oats. Plant Soil 253:1–34. https://doi.org/10.1023/A:1024573305997
Shanazari M, Golkar P, Maibody AMM (2018) Effects of drought stress on some agronomic and bio-physiological traits of Trtiticum aestivum, Triticale, and Tritipyrum genotypes. Archives of Agronomy and Soil Science 64:2005–2018. https://doi.org/10.1080/03650340.2018.1472377
Sharma B, Rashid U, Anwar F, Erhan S (2009) Lubricant properties of Moringa oil using thermal and tribological techniques. J Therm Anal Calori 96:999–1008
Shiferaw, B., Kassie, M., Jaleta, M., Yirga, C. (2014). Adoption of improved wheat varieties and impacts on household food security in Ethiopia. Food Policy 44:272-284
Singh S, Angadi SV, Grover K, Begna S, Auld D (2016) Drought response and yield formation of spring safflower under different water regimes in the semiarid Southern High Plains. Agric Water Manag 163:354–362
Singhal RS, Kulkarni PR (1988a) Review: amaranths-an underutilized resource. Int J Food Sci Technol 23:125–139. https://doi.org/10.1111/j.1365-2621.1988.tb00559.x
Singhal RS, Kulkarni PR (1988b) Review: amaranths-an underutilized resource. Int J Food Sci Technol 23:125–139. https://doi.org/10.1111/j.1365-2621.1988.tb00559.x
Smith LS, Thelen KD, MacDonald SJ (2013) Yield and quality analyses of bioenergy crops grown on a regulatory brownfield. Biomass and Bioenergy 49:123–130. https://doi.org/10.1016/j.biombioe.2012.12.017
Solaiman Z, Colmer TD, Loss SP, Thomson BD, Siddique KHM (2007) Growth responses of cool-season grain legumes to transient waterlogging. Aust J Agric Res 58(5):406–412
Sun Y, Niu G, Osuna P, Zhao L, Ganjegunte G, Peterson G, Peralta-Videa JR, Gardea-Torresdey JL (2014) Variability in salt tolerance of sorghum bicolor L. Agricultural Science 2. Issue 1:09–21
Suriano S, Iannucci A, Codianni P, Fares C, Russo M, Pecchioni N, Marciello U, Savino M (2018) Phenolic acids profile, nutritional and phytochemical compounds, antioxidant properties in colored barley grown in southern Italy. Food Res Int 113:221–233
Taffouo VD, Kouamou JK, Ngalangue LMT, Ndjeudji BAN, Akoa A (2009) Effects of salinity stress on growth, ions partitioning and yield of some cowpea (Vigna unguiculata L. Walp.) cultivars. Int. J. Bot 5(2):135–143
Tang Y, Xie JS, Geng S (2010) Marginal land-based biomass energy production in China. J Integr Plant Biol 52:112–121. https://doi.org/10.1111/j.1744-7909.2010.00903.x
Tarawali SA, Singh BB, Peters M, Blade SF (1997a) Cowpea haulms as fodder. In: Singh BB (ed) Advances in Cowpea Research, IITA
Tarawali SA, Singh BB, Peters M, Blade SF (1997b) Cowpea haulms as fodder. In: Singh BB (ed) Advances in Cowpea Research, IITA
Tester M, Langridge P (2010) Breeding technologies to increase crop production in a changing world. Science 327:818–822. https://doi.org/10.1126/science.1183700
Theib O, Ahmed H, Mustafa MP (2005) Faba bean productivity under rainfed and supplementing irrigation in northern Syria. Agric Water Management 73 N(1):57–72
Tilman D, Hill J, Lehman C (2006) Carbon-negative biofuels from low-input high-diversity grassland biomass. Science 314:1598–1600. https://doi.org/10.1126/science.1133306
Tomich TP, Kilby P, Johnston BF (1995) Transforming agrarian economies: opportunities seized, opportunities missed. Cornell University Press, USA
Ullah R, Nadeem M, Khalique A, Imran M, Mehmood S, Javid A, Hussain J (2015) Nutritional and therapeutic perspectives of chia (Salvia hispanica L.): a review. J Food Sci Technol 1:9. https://doi.org/10.1007/s13197-015-1967-0
Vadez V, Krishnamurthy L, Serraj R, Gaur PM, Upadhyaya HD, Hoisington DA, Siddique KHM (2007) Large variation in salinity tolerance in chickpea is explained by differences in sensitivity at the reproductive stage. Field Crop Res 104(1-3):123–129. https://doi.org/10.1016/j.fcr.2007.05.014
Velasco L, Pérez-Vich B, Fernández-Martínez JM (2005) Identification and genetic characterization of a safflower mutant with a modified tocopherol profile. Plant Breed 124:459–463
Vigouroux Y, Barnaud A, Scarcelli N, Thuillet AC (2011) Biodiversity, evolution and adaptation of cultivated crops. Comptes rendus biologies 334:450–457
Vijayalakshmi T, Varalaxmi Y, Jainender S, Yadav SK, Vanaja M, Jyothilakshmi N, Maheswari M (2012) Physiological and biochemical basis of water-deficit stress tolerance in pearl millet hybrid and parents. Am J Plant Sci 3:1730–1740
Waddington SR, Li X, Dixon J, Hyman G, De Vicente MC (2010) Getting the focus right: production constraints for six major food crops in Asian and African farming systems. Food Security 2(1):27–48
Wezel A (2000) Scattered shrubs in pearl millet fields in semiarid Niger: Effect on millet production. Agrofor. Syst. 48:219–228
Win AN, Xue Y, Chen B, Liao F, Chen F, Yin N, Mei F, Wang B, Shi X, He Y, Chai Y (2018) Chia (Salvia hispanica) experiment at a 30° N site in Sichuan Basin, China. Ciência Rural 48(9):e20180105. https://doi.org/10.1590/0103-8478cr20180105
Yadav PC, Sadhu AC, Swarnkar P, Pate MR (2010) Effect of integrated nitrogen management on forage yield of multicut sorghum, available nitrogen and microbial count in the soil. J. Indian Soc Soil Sci. 58:303–308
Yazar A, Incekaya Ç, Sezen SM, Jacobsen SE (2015a) Saline water irrigation of quinoa (Chenopodium quinoa) under Mediterranean conditions. Crop Pasture Sci 66(10):993–1002
Yazar A, Incekaya Ç, Sezen SM, Jacobsen SE (2015b) Saline water irrigation of quinoa (Chenopodium quinoa) under Mediterranean conditions. Crop Pasture Sci 66(10):993–1002
Yeboah S, OwusuDanquah E, Lamptey J, Mochiah M, Lamptey S, Oteng-Darko P et al (2013) Influence of planting methods and density on performance of chia (Salvia hispanica) and its suitability as an oilseed plant. Agric Sci 2(14):26
Yeilaghi H, Arzani A, Ghaderian M, Fotovat R, Feizi M, Pourdad SS (2012) Effect of salinity on seed oil content and fatty acid composition of safflower (Carthamus tinctorius L.) genotypes. Food Chem 130:618–625
Yousfi S, Serret MD, Voltas J, Araus JL (2010) Effect of salinity and water stress during the reproductive stage on growth, ion concentrations, ∆13C, and d15N of durum wheat and related amphiploids. J Exp Bot 61(13):3529–3542
Youssef T, Awad MA (2008) Mechanisms of Enhancing Photosynthetic Gas Exchange in Date Palm Seedlings (Phoenix dactylifera L.) under Salinity Stress by a 5- Aminolevulinic Acid-based Fertilizer. J Plant Growth Regulation 27(1):1–9
Zannini E, Jones JM, Renzetti S, Arendt EK (2012) Functional replacements for gluten. Annu Rev Food Sci Technol 3:227–245
Zhang F, Yu J, Johnston CR, Wang Y, Zhu Y, Lu F, Zhang Z, Zou J (2015) Seed Priming with Polyethylene Glycol Induces Physiological Changes in Sorghum (Sorghum bicolor L. Moench) Seedlings under Suboptimal Soil Moisture Environments. PLoS ONE 10(10):e0140620. https://doi.org/10.1371/journal.pone.0140620
Zhu F (2018) Triticale: Nutritional composition and food uses. Food Chem 241:468-479
Zhu Y, Chen H, Fan J, Wang Y, Li Y, Chen J, Fan J, Yang S, Hu L, Leung H, Mew TW (2000) Genetic diversity and disease control in rice. Nature 406(6797):718-722
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Responsible editor: Kitae Baek
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Hussain, M.I., Farooq, M., Muscolo, A. et al. Crop diversification and saline water irrigation as potential strategies to save freshwater resources and reclamation of marginal soils—a review. Environ Sci Pollut Res 27, 28695–28729 (2020). https://doi.org/10.1007/s11356-020-09111-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-020-09111-6