
Abstract
Acrylamide (AA) is a heat-induced toxin formed during thermal processing of many commonly consumed foods, including meat products, French fries, potato crisps, bread, cereals, cookies, and coffee. There is thus potentially high dietary exposure of humans to AA, which can induce significant oxidative stress. Hesperidin (HS) and diosmin (DS) are flavone glycosides that have antioxidant properties. The aim of this study was to investigate the protective effects of HS and DS against AA toxicity. Fifty-six adult male Wistar albino rats were divided into seven groups. The first group was orally administered 0.5% (w/v) dimethyl sulfoxide (DMSO) and considered as the control group. The second and third groups were orally administered 10 mg/kg/day of HS or DS, respectively. The fourth group received 20 mg/kg/day of AA orally for 14 days. The fifth and sixth groups were given 10 mg/kg/day of HS or DS, respectively, followed by AA. The seventh group was given both HS and DS after AA administration. AA intoxication significantly (p ≤ 0.05) increased serum levels of liver function enzymes (ALT, AST, and ALP), kidney function products (urea and creatinine), oxidative DNA damage marker (OHdG), proinflammatory markers (TNF-α, IL-1β, and IL-6), lipid peroxidation marker (malondialdehyde), and nitric oxide (NO). On the other hand, it significantly (p ≤ 0.05) decreased levels of reduced glutathione (GSH) in the liver, kidney, and brain. The activities of glutathione peroxidase (GSH-Px), superoxide dismutase (SOD), and catalase (CAT) in the liver, kidney, and brain tissues were also reduced. HS and DS supplementation prevented lipid peroxidation, normalized the serum parameters altered by AA, and enhanced the tissue concentrations and activities of antioxidant biomarkers. It could be concluded that HS and DS have potent protective effects against oxidative stress, lipid peroxidation, and DNA damage induced by AA toxicity in rats.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Acrylamide, or acrylic amide (AA), is a water-soluble, odorless, white, and crystalline chemical compound. The amyloid monomers of AA have the chemical formula C3H5NO (Kim et al. 2017). AA is formed during the heat treatment of various foods, such as French fries, bread, cereals, and meat products at temperatures of 120 °C or higher (Ghasemian et al. 2014; Mottram et al. 2002; Stadler et al. 2002). AA forms ammonia by non-thermal decomposition and produces oxides of nitrogen, carbon dioxide, and carbon monoxide during thermal decomposition (Charoenpanich 2013). Moreover, it is used industrially to synthesize the polyacrylamide that is utilized in wastewater treatment, tertiary oil recovery, gel electrophoresis, paper making, and sometimes the production of dyes and other monomers (Friedman 2003). Food and Agriculture Organization (FAO)/World Health Organization (WHO) committee employed “margin of exposure (MOE)” as an approach to estimate AA risk assessment (Authority 2005; FAO 2002). Based on the this risk assessment, the MOE of AA showed a severe human health concern compared to other chemicals (Authority 2005).
The toxic effects of AA have been studied since the 1950s. In vivo, AA is biotransformed into its epoxide, glycidamide (C3H5NO2), which is a potential genotoxin with mutagenic properties both in vitro and in vivo (Calleman et al. 1990). Meanwhile, long-term exposure to AA may also cause some damage to the neurons in humans, and cumulative neurotoxicity, including cerebellar Purkinje cell death, ataxia, skeletal muscle weakness, cognitive impairment, and degeneration of distal axons and nerve terminals, has been suggested to be a potential result of occupational exposure to AA monomers (LoPachin et al. 2002). AA is absorbed through the skin and spread throughout the circulatory system, which thus contains the highest levels of AA after exposure. The levels of AA then begin to rise in some organs, especially in the kidneys, liver, and testes (Hamdy et al. 2017). It was previously observed that some flavanone glycoside derivatives have protective effects against AA-induced toxicity (Hamdy et al. 2017; Uthra et al. 2017).
Hesperidin (HS) is a flavanone glycoside that can be isolated in large amounts from citrus fruits, such as Satsuma mandarin (Citrus unshiu Marcov.), sweet orange (Citrus sinensis L.), bitter orange (Citrus aurantium L.), and lemon (Li et al. 2018) that are readily available in everyday life. Several biomedical applications of HS have been reported, such as its ability to chelate metal ions (Kuntić et al. 2014) and its potential anti-allergenic (Kim et al. 2011), anti-inflammatory (Wei et al. 2012), radio-protective (Petrova et al. 2011), anti-cancer (Yumnam et al. 2016), and antioxidant properties (Selvaraj and Pugalendi 2010), as well as it being able to suppress cyclooxygenase-2 (COX-2) gene expression (Shaban et al. 2017). Furthermore, HS reduces levels of nitrogen oxide (NO•), the superoxide anion radical (O2•−), the superoxide radical (LOO•), the hydroxyl radical (OH•), and singlet oxygen (1O2) (Maneesai et al. 2018). Moreover, it helps in the treatment of kidney damage and liver tissue damage caused by inflammation (Fouad et al. 2019; Turk et al. 2019).
Diosmin (DS) is a flavone glycoside derivative of diosmetin, which is mainly present in plants in the citrus family and herbs like Teucrium gnaphalodes L’Her. DS is made semi-synthetically through the dehydrogenation of HS (Szymański et al. 2016). DS has been demonstrated to have many pharmacological benefits in the treatment of such vascular diseases as varicose veins, venous ulcers, chronic venous insufficiency, peripheral edema, and stasis dermatitis (Jantet 2002). Moreover, it was reported that DS has an anti-hyperglycemic effect because it enhances the secretion of β-endorphin and increases the glucose uptake of tissues (Hsu et al. 2017). In addition, DS plays a significant role in delaying the formation of cataracts in diabetic rats (Wojnar et al. 2017). In combination with HS, DS has also been used as a nutritional supplement in the USA and some European countries. Moreover, it has anti-inflammatory (Tahir et al. 2013) and anti-hyperglycemic effects (Ahmed et al. 2016).
Therefore, in this study, we investigated the protective effects of HS and DS against AA-induced toxicity in rats. To do this, lipid peroxidation, DNA oxidation, and levels of antioxidant biomarkers, serum proinflammatory cytokines, and hepatic and renal injury markers in the liver, kidney, and brain tissues were evaluated.
Material and methods
Chemicals
Pure (99%) acrylamide, hesperidin, and diosmin were obtained from Sigma Chemical Company (St Louis, MO, USA). All assay kits were bought from Biodiagnostics Co. (Cairo, Egypt). Enzyme-linked immunosorbent assay (ELISA) kits for analyzing cytokines were purchased from R&D (Mannheim, Germany), while those for 8-hydroxy-2'-deoxyguanosine (8-OhdG) were purchased from Cayman Chemical Co. (MI, USA).
Experimental animals
We obtained a total of 56 adult male Wistar albino rats weighing 150 ± 10 g from the Egyptian Organization for Biological Products and Vaccines. For 7 days, the animals were acclimatized to the experimental laboratory conditions (Department of Pharmacology, Faculty of Veterinary Medicine, Suez Canal University, Egypt) while being kept at a temperature of 25 ± 2 °C and under a 12-h light/dark cycle in a well-ventilated room with moderate humidity (60 ± 5%). All the animal rearing and handling procedures in this study were approved by the Research Ethical Committee at the Faculty of Veterinary Medicine, Suez Canal University, Egypt. All possible measures were taken to alleviate any suffering and stress experienced by the animals.
Experimental design
Rats were randomly assigned to one of seven different groups, with each group representing a different treatment and containing eight animals. The first group was given 0.5% (w/v) dimethyl sulfoxide (DMSO) orally for 21 days and was considered the control group. Rats in the second and third groups were given 10 mg/kg/day of HS or DS, respectively, dissolved in 0.5% DMSO for 21 days, with this mixture administered orally by stomach gavage. Animals in the fourth group were orally administered 20 mg/kg of AA daily for 14 days, according to the recommendations of Abramsson-Zetterberg (Abramsson-Zetterberg 2003) and Zhang et al. (Zhang et al. 2013). In the fifth and sixth groups, rats were orally administered HS or DS, respectively, at the same doses as those used in the second and third groups, and then administered AA at the same dose as that used in the fourth group. Rats in the seventh group were administered both HS and DS after intoxication with AA at the same dose as that used in the fourth group.
Blood collection and serum and tissue preparation
Blood samples were collected via direct cardiac puncture. Samples were centrifuged at 3000 rpm for 15 min after coagulation under the normal laboratory conditions described earlier. Furthermore, rats were sacrificed by decapitation, and then brain, kidney, and liver samples were rapidly excised and dissected into two parts each, the first of which was fixed in 10% formalin for histopathological analysis and the second of which was used for biochemical analyses. Tissue samples (0.5 g each) were then washed with a standard cold saline solution (0.9% NaCl in distilled water), homogenized in 2.5× their volume of ice-cold 50 mM sodium phosphate buffer (pH = 7.4) and 0.1 mM EDTA (ethylenediaminetetraacetic acid) to eliminate red blood cells (RBCs) and clots. The resulting homogenates were centrifuged at 5000 rpm for 30 min, and then the supernatant was collected and stored at − 80 °C to later evaluate the content and activities of lipid peroxidation and antioxidant biomarkers in each tissue type (Fig. 1).

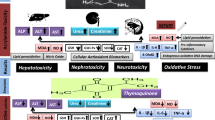
Structure, sources, and toxicity of acrylamide. Abbreviations: MDA, malondialdehyde concentration; GSH, reduced glutathione concentration; SOD, superoxide dismutase activity; CAT, catalase activity; 8-OHdG, 8-hydroxy-2'-deoxyguanosine; TNF-α, tumor necrosis factor-alpha
Serum biochemical analysis
The collected serum samples were used for the measurement of their content of renal injury markers. Creatinine levels were estimated using the method reported by Larsen (Larsen 1972), and urea levels were estimated using the method defined by Coulombe (Coulombe 1963). Moreover, the content of alkaline phosphatase (ALP) was evaluated according to the method of Tietz et al. (Tietz et al. 1983). To measure the serum levels of alanine transferase (ALT) and aspartate transferase (ALT), we used the methods illustrated by Reitman and Frankel (Reitman and Frankel 1957).
Evaluation of tissue lipid peroxidation and antioxidant enzymes’ activities
To assess lipid peroxidation, the methods illustrated by Mihara and Uchiyama (Mihara and Uchiyama 1978) were followed to measure the brain, hepatic, and renal tissues’ content of malondialdehyde (MDA). The concentration of nitric oxide (NO) in these tissues was evaluated according to the method of Green et al. (Green et al. 1982). Levels of an antioxidant marker, reduced glutathione (GSH), were measured according to the methods of Beutler et al. (Beutler et al. 1963). The activities of antioxidant enzymes, including glutathione peroxidase (GSH-Px), catalase (CAT), and superoxide dismutase (SOD), were analyzed according to the methods of Aebi (Aebi 1984), Nishikimi et al. (Nishikimi et al. 1972), and Paglia and Valentine (Paglia and Valentine 1967), respectively.
Estimation of oxidative DNA damage markers and proinflammatory cytokines
ELISA kits were used to measure the serum levels of interleukin-1β (IL-1β) and interleukin-6 (IL-6) (Glory Science Co. Ltd., Del Rio, TX, USA), as well as tumor necrosis factor-alpha (TNF-α) (BioSource International Inc., Camarillo, CA, USA). These analyses were performed while following the manufacturer’s instructions. The absorbance of each marker was read using an automatic ELISA reader at 420 nm.
Statistical analyses
All values are reported herein as the mean and the standard error of the mean (SEM) value of each measured parameter in each treatment. Data were analyzed using one-way analysis of variance (ANOVA), accompanied by Tukey’s post hoc test, to test for significant differences among treatments. A p value ≤ 0.05 was considered statistically significant. All analyses were performed using SPSS (Statistical Package for Social Sciences) software (version 22).
Results
Effects of HS and DS on tissue serum biochemical parameters in AA-intoxicated rats
The deleterious effects of AA intoxication and the preventive effects of HS and/or DS on the different serum biochemical parameters measured in this study are presented in Table 1. Our findings demonstrated that there were significantly (p ≤ 0.05) elevated serum levels of liver function enzymes (ALT 281.23%; AST 240.86%; ALP 223.62%) and kidney function enzymes (urea, 299.66%; creatinine, 981.58%) in the AA-intoxicated rats compared to those in control rats. Similarly, serum levels of the products of DNA oxidation and proinflammatory cytokines were significantly (p ≤ 0.05) increased, as there were higher levels of 8-OHdG (244.91%), TNF-α (197.66%), IL-1β (248.90%), and IL-6 (234.47%) in AA-intoxicated rats than in control rats (Fig. 2).

Protective effects of hesperidin (HS) and diosmin (DS) against acrylamide-induced changes in serum levels of various biomarkers: a 8-OHdG, 8-hydroxy-2'-deoxyguanosine; b TNF-a, tumor necrosis factor-alpha; c IL-1β, interleukin-1β; and d IL-6, interleukin 6. Data are presented as mean ± SEM values. Columns labelled with different letters are significantly different (p ≤ 0.05).
Administration of HS or DS to the AA-intoxicated rats ameliorated most of the negative effects of AA on their serum levels of ALT, AST, ALP, urea, and creatinine, as these were significantly (p ≤ 0.05) reduced in these treatments (by about 32.78, 39.2, 30.86, 39.6, and 29.76% for HS, and 36.39, 46.2, 37.57, 43.47, and 37.91% for DS, respectively) compared to those in the AA-intoxicated group. Compared with the control group, rats treated with HS and DS showed slight reductions in the serum levels of these parameters, although this reduction was not statistically significant (p > 0.05). While there was no significant differences between these two groups (AA-HS and AA-DS), the levels of these parameters were still higher in them than those in the control group. In the group of rats that were treated with both DS and HS, the serum levels of these parameters were significantly (p ≤ 0.05) reduced compared with those in the AA-DS and AA-HS groups. Compared to the control group, there was a slight, but non-significant, increase in the values of these parameters in the AA-HS-DS group.
In terms of 8-OHdG and TNF-α levels, those in AA-intoxicated rats were significantly (p ≤ 0.05) higher than those in the control group. Administration of HS or DS to the AA-intoxicated rats significantly (p ≤ 0.05) decreased the levels of 8-OHdG (HS 19.41%; DS 29.75%) and TNF-α (HS 17.14%, DS 20.44%) in them compared with those in the AA group. In the AA-HS-DS group, HS and DS completely relieved the deleterious effects of AA on the serum levels of 8-OHdG and TNF-α.
Concerning the levels of IL-1β and IL-6, these were significantly (p ≤ 0.05) increased in the AA-intoxicated group compared to those in the control group. HS and DS administration led to significant reductions (by 31.25 and 36.49%, respectively) in IL-1β levels. Moreover, the HS and DS treatments also led to significant reductions (by 36.57 and 42.75%, respectively) in IL-6 levels when they were administered to the AA-intoxicated rats. The serum levels of IL-1β and IL-6 were returned to normal values in the rats in the AA-HS-DS group.
Role of HS and DS in ameliorating the oxidative stress and production of antioxidant biomarkers induced by AA intoxication
Liver
The deleterious effects of AA and the preventive effects of HS and/or DS on hepatic lipid peroxidation (LPO) and the values of antioxidant parameters are shown in Fig. 3. AA intoxication caused a significant (p ≤ 0.05) increase in the hepatic MDA (196.60%) and NO (175.52%) content. On the other hand, it caused a significant (p ≤ 0.05) decrease in the concentration of GSH (62.5%) and in the activities of GSH-Px (76.06%), SOD (69.78%), and CAT (81.07%) compared to those in the control group.

Protective effects of hesperidin (HS) and diosmin (DS) against acrylamide-induced hepatotoxicity, assessed based on a MDA, malondialdehyde concentration; b NO, nitric oxide concentration; c GSH, reduced glutathione concentration; d GSH-Px, glutathione peroxidase activity; e SOD, superoxide dismutase activity; and f CAT, catalase activity. Data are presented as mean ± SEM values. Columns labelled with different letters are significantly different (p ≤ 0.05).
In the AA-HS and AA-DS groups, hepatic MDA and NO levels were significantly (p ≤ 0.05) decreased (AA-HS by 30.19 and 17.53%, respectively; AA-DS by 35.70 and 27.03%, respectively) compared to those in the AA-intoxicated group, but they were still higher than those in the control group, with there being no significant difference between these two groups (AA-HS and AA-DS). However, in the AA-HS-DS group, there was a complete recovery of the concentrations of MDA and NO that made them almost the same as those in the control group, with there not being any significant difference between this group and the control. In contrast, HS and/or DS administration to the AA-intoxicated group increased the concentration of GSH (by 173.37 and 183.55%, respectively) and the activities of GSH-Px (268.43 and 282.36%, respectively), SOD (220.32 and 239.34%, respectively), and CAT (246.15 and 294.87%, respectively) in their liver tissues compared to those in the AA-intoxicated group. Administration of both HS and DS helped protect the hepatic tissues against the toxic effects of AA and restored the levels of oxidative stress and antioxidant biomarkers in the liver to their normal state.
Kidney
The renal concentrations and activities of MDA, NO, and GSH, and the renal LPO parameters of the control and treated groups, are shown in Fig. 4. The renal MDA and NO concentrations were significantly (p ≤ 0.05) increased (by 206.32 and 168.52%, respectively) in the AA-intoxicated rats compared to those in the control. On the other hand, significant (p ≤ 0.05) reductions in the renal content of GSH (49.68%) and activities of GSH-Px (56.66%), SOD (70.33%), and CAT (71.62%) were observed in the AA-intoxicated rats compared with those in the control group.

Protective effects of hesperidin (HS) and diosmin (DS) against acrylamide-induced nephrotoxicity, assessed based on a MDA, malondialdehyde concentration; b NO, nitric oxide concentration; c GSH, reduced glutathione concentration; d GSH-Px, glutathione peroxidase activity; e SOD, superoxide dismutase activity; and f CAT, catalase activity. Data are presented as mean ± SEM values. Columns labelled with different letters are significantly different (p ≤ 0.05).
In the AA-HS, AA-DS, and AA-HS-DS groups, the renal MDA and NO levels were significantly (p ≤ 0.05) decreased in comparison with those in the AA-intoxicated rats (AA-HS: 23.24 and 16.14%, respectively; AA-DS: 29.19 and 22.46%, respectively; AA-HS-DS: 50.18 and 37.45%, respectively). Regarding the renal concentration of GSH and the activities of GSH-Px, SOD, and CAT, there were significant (p ≤ 0.05) increases in the values of these parameters with HS and DS supplementation (AA-HS: by 133.21, 139.34, 200.29, and 239.68%, respectively; AA-DS: by 141.59, 156.34, 217.95, and 268.25%, respectively) compared to those in the control group. In the AA-HS-DS group, all parameters reached the same levels as those measured in the control group, except for the concentration of GSH and activities of SOD and CAT, which showed only partial recoveries toward control values compared to those in the AA-intoxicated group.
Brain
The harmful effects of AA and the protective effects of HS and/or DS on brain tissue LPO and antioxidant parameters are shown in Fig. 5. Significant (p ≤ 0.05) increases in the concentrations of MDA (222.42%) and NO (193.15%) in the brain were observed. On the other hand, the concentration of GSH and the activities of GSH-Px, SOD, and CAT were significantly (p ≤ 0.05) reduced in the brain tissues of the AA-intoxicated group compared to those in the control group (by 61.04, 70.57, 72.03, and 69.66%, respectively).

Protective effects of hesperidin (HS) and diosmin (DS) against acrylamide-induced neurotoxicity, assessed based on a MDA, malondialdehyde concentration; b NO, nitric oxide concentration; c GSH, reduced glutathione concentration; d GSH-Px, glutathione peroxidase activity; e SOD, superoxide dismutase activity; and f CAT, catalase activity. Data are presented as mean ± SEM values. Columns labelled with different letters are significantly different (p ≤ 0.05).
Administration of HS or DS to AA-intoxicated rats significantly (p ≤ 0.05) reduced the concentrations of MDA and NO in their brain tissues (AA-HS: by 25.70 and 27.17%, respectively; AA-DS: by 37.00 and 34.00%, respectively). However, these treatments significantly (p ≤ 0.05) increased the concentration of GSH and the activities of GSH-Px, SOD, and CAT in the brain tissues in the AA-HS group (by 169.70, 199.83, 167.27, and 176.23%, respectively) and AA-DS group (by 178.38, 236.68, 194.48, and 201.98%, respectively) in comparison to those in the AA-intoxicated group. The treatment of the AA-intoxicated group with a combination of HS and DS maintained the concentrations and activities of the measured parameters in the brain tissues of the AA-HS-DS group within their normal (control) values.
Discussion
This was the first study to have assessed the protective role of HS and/or DS against AA-induced injuries to hepatic, renal, and brain tissues. Our results highlighted the fact that the administration of AA (at a dose of 20 mg/kg of body weight) significantly induced the inflammatory response and oxidative injury and reduced the activities of antioxidant enzymes, in the liver, kidney, and brain. On the other hand, we demonstrated that HS and DS significantly ameliorated the inflammatory response and oxidative injury induced by AA exposure. Notably, the combination of HS and DS was more effective in reducing AA-induced injuries than either HS or DS was alone, which suggests that there is a more beneficial effect when used in combination.
Reactive oxygen species (ROS) is a broad umbrella term, under which many different chemical species containing oxygen are included, such as superoxide, peroxides, alpha-oxygen, and the hydroxyl radical (Hayyan et al. 2016). It is well-known that ROS have critical roles in homeostasis and cell signaling. ROS are produced in mammals as antimicrobial agents and as normal products of cellular metabolism (Bhattacharyya et al. 2014). Nevertheless, ROS production can be increased dramatically under various types of abnormal conditions (e.g., toxicities, heat exposure, and ultraviolet radiation), which can cause significant damage to cellular structures, and thus induce so-called ROS toxicity or oxidative stress (Babizhayev 2016). This toxicity may cause the oxidation of amino acids, damage to DNA or RNA, oxidative deactivation of specific enzymes, and the oxidation of polyunsaturated fatty acids into lipids (Nita and Grzybowski 2016). Furthermore, ROS inhibit the oxidative defense systems in organelles, which leads to many diseases, such as diabetes, atherosclerosis, and cancer (Lobo et al. 2010). The current body of evidence shows that accumulation of AA induces oxidative damage in different organs of the body, including the brain, liver, and kidneys (Yousef and El-Demerdash 2006).
Our results showed that administration of AA (at a dose of 20 mg/kg of body weight) induced the inflammatory response and DNA oxidation through the elevation of levels of 8-OHdG, TNF-α, IL-1β, and IL-6 in organ tissues. Moreover, AA caused dramatic reductions in the concentration of GSH and the activities of GSH-Px, SOD, and CAT, but caused significant elevations in the concentrations of MDA and NO. In line with our findings, a recent study by Hamdy et al. (Hamdy et al. 2017) demonstrated significant increases in the serum levels of biomarkers of liver and kidney injury (such as AST, ALT, and creatinine), oxidative DNA damage markers, and proinflammatory cytokines after AA exposure in female rats. Our results were also in agreement with the findings of several other studies (Abd El-Halim and Mohamed 2012; Abdel-Daim et al. 2015; Alturfan et al. 2012; Venkatasubbaiah et al. 2014; Zhang et al. 2013), which showed that AA has deleterious effects on brain, liver, and kidney tissues because it increases the incidence of lipid peroxidation and alters antioxidant enzyme systems. Such a dramatic increase in the occurrence of lipid peroxidation, especially in the brain, liver, and kidneys, potentially has a role in changing the oxidative stress state of an organism, as was indicated by the observed reduction of the concentration of GSH and the activities of SOD, GSH-Px, and CAT in the present study. Moreover, the elevation of NO and MDA concentrations is also considered a marker of this kind of oxidative stress state alteration (Abd El-Halim and Mohamed 2012; Abdel-Daim et al. 2015; Alturfan et al. 2012; Zhang et al. 2013). Glutathione S-transferase (GST) can stimulate the conjugation of the reduced form of GSH and SOD for the detoxification and breakdown of xenobiotic molecules (Ramsay and Dilda 2014). These effects of GST and SOD have significant roles in defending against ROS and protecting the organs of the body (Birben et al. 2012). AA is known to have several delirious effects. It reduces the activity and concentration of GST and GSH, respectively. Furthermore, AA contributes to the accumulation of free radicals, NO, and superoxide, which react together to form peroxynitrite, a chemical that can damage cell membranes. Moreover, when the production of ROS exceeds the ability of the antioxidant defense system of the cell to control, oxidative stress may cause various pathological conditions (Abdel-Daim et al. 2015).
As mentioned before, HS and DS are polyphenol compounds that could prevent AA toxicity via several mechanisms. The antioxidant properties of polyphenols are mediated by their involvement in direct radical scavenging and augmenting cellular antioxidant defenses (Parhiz et al. 2015). Furthermore, like many other polyphenol compounds, HS can promote or upregulate the expression of the genes nuclear factor-erythroid 2 p45 (Nrf2) and ERK 1/2, which leads to the upregulation of HO-1 expression, which decreases intracellular concentrations of pro-oxidants and increases those of bilirubin, an endogenous antioxidant (Elavarasan et al. 2012). Nrf2 increases the levels of antioxidant enzymes in the cell, such as GST, CAT, and SOD. Thus, through its effects on ERK/Nrf2 signaling, HS increases the capacity of the cellular antioxidant defense system by inducing HO-1 expression (Chen et al. 2010). Regarding anti-inflammatory properties, it was observed herein that HS decreased the NO2 content and suppressed the expression of the iNOS protein in tissues, which resulted in the inhibition of NO production and the suppression of the activities of COX-1 and COX-2 (Sakata et al. 2003). Flavonoids have the ability to block the NF-κB and MAPK signaling pathways, which are responsible for the regulation of inflammation-induced enzymes and cytokines, such as TNF-α, IL-1β, and IL-6 (Kang et al. 2011).
In the present study, the supplementation of AA-intoxicated rats with HS or DS maintained the serum concentrations of inflammatory biomarkers and DNA oxidation markers within normal levels in their brain, liver, and kidney tissues. However, these levels were still higher than those in the control. Besides, HS and DS enhanced the activities of enzyme systems and the concentration of antioxidant molecules. It was reported that HS reduced the oxidative stress by inhibition of the expression of nicotinamide adenine dinucleotide phosphate oxidase. Additionally, HS preserved the bioavailability of NO in the body tissues (Kouhpayeh et al. 2008). Our findings match those of Mahmoud et al. (Mahmoud et al. 2012) and Sahu et al. (Sahu et al. 2013), who reported that HS had a significant effect on the reversion of many biochemical parameters, such as GSH, MDA, SOD, ROS, NO, TNF-a, and IL-6 levels, to their normal states in cisplatin-induced nephrotoxic rats and high-fat-fed, streptozotocin-induced type-2 diabetic rats. This effect can be explained by the ability of HS to suppress the production of proinflammatory cytokines and potentiate the antioxidant defense system (Mahmoud et al. 2012). Additionally, DS can scavenge free radicals and upregulate the expression of such cellular antioxidant enzymes as CAT, SOD, and GSH-Px. These findings supported those of our previous study, in which we demonstrated the protective role of DS against methotrexate-induced cytotoxicity in rats (Abdel-Daim et al. 2017). Moreover, they are also in agreement with the findings of Vasco et al. (Vasco et al. 2018), who demonstrated that there was a protective effect of HS/DS in an animal model of sepsis-induced acute kidney injury.
Notably, we found that the combination of HS and DS was more effective in reducing the severity of AA-induced injuries than either HS or DS was alone. Further studies are recommended using different doses of each of them and their combinations in order to deeply understand the type of HS-DS interaction.
Conclusions
Acrylamide-induced toxicity caused damage and injury to liver, kidney, and brain tissues through the increase of DNA oxidative damage and the reduction of the antioxidant enzyme activities. Our findings demonstrated that HS and DS significantly reduced the serum and tissues’ biochemical alterations induced by AA.
References
Abd El-Halim SS, Mohamed MM (2012) Garlic powder attenuates acrylamide-induced oxidative damage in multiple organs in rat. Journal of Applied Sciences Research 8:168–168
Abdel-Daim MM, Abd Eldaim MA, Hassan AGA (2015) Trigonella foenum-graecum ameliorates acrylamide-induced toxicity in rats: roles of oxidative stress, proinflammatory cytokines, and DNA damage. Biochemistry and Cell Biology 93:192–198
Abdel-Daim MM, Khalifa HA, Abushouk AI, Dkhil MA, Al-Quraishy SA (2017) Diosmin attenuates methotrexate-induced hepatic, renal, and cardiac injury: a biochemical and histopathological study in mice. Oxidative Medicine and Cellular Longevity 2017:1–10
Abramsson-Zetterberg L (2003) The dose-response relationship at very low doses of acrylamide is linear in the flow cytometer-based mouse micronucleus assay. Mutation research 535:215–222
Aebi H (1984) Catalase in vitro. Methods in enzymology 105:121–126
Ahmed S, Mundhe N, Borgohain M, Chowdhury L, Kwatra M, Bolshette N, Ahmed A, Lahkar M (2016) Diosmin modulates the NF-kB signal transduction pathways and downregulation of various oxidative stress markers in alloxan-induced diabetic nephropathy. Inflammation 39:1783–1797
Alturfan AA, Tozan-Beceren A, Şehirli AÖ, Demiralp E, Şener G, Omurtag GZ (2012) Resveratrol ameliorates oxidative DNA damage and protects against acrylamide-induced oxidative stress in rats. Molecular Biology Reports 39:4589–4596
Authority EFS (2005) Opinion of the Scientific Committee on a request from EFSA related to a harmonised approach for risk assessment of substances which are both genotoxic and carcinogenic. EFSA Journal 3:282
Babizhayev MA (2016) Generation of reactive oxygen species in the anterior eye segment. Synergistic codrugs of N-acetylcarnosine lubricant eye drops and mitochondria-targeted antioxidant act as a powerful therapeutic platform for the treatment of cataracts and primary open-an. BBA Clinical 6:49–68
Beutler E, Duron O, Kelly BM (1963) Improved method for the determination of blood glutathione. The Journal of laboratory and clinical medicine 61:882–888
Bhattacharyya A, Chattopadhyay R, Mitra S, Crowe SE (2014) Oxidative stress: an essential factor in the pathogenesis of gastrointestinal mucosal diseases. Physiological reviews 94:329–354
Birben E, Sahiner UM, Sackesen C, Erzurum S, Kalayci O (2012) Oxidative stress and antioxidant defense. The World Allergy Organization journal 5:9–19
Calleman CJ, Bergmark E, Costa LG (1990) Acrylamide is metabolized to glycidamide in the rat: evidence from hemoglobin adduct formation. Chem Res Toxicol 3:406–412
Charoenpanich J (2013): Removal of acrylamide by microorganisms. Applied Bioremediation - Active and Passive Approaches
Chen M, Gu H, Ye Y, Lin B, Sun L, Deng W, Zhang J, Liu J (2010) Protective effects of hesperidin against oxidative stress of tert-butyl hydroperoxide in human hepatocytes. Food and Chemical Toxicology 48:2980–2987
Coulombe F (1963) A new simple semimicro method for colorimetric determination of urea. Clinical chemistry 9:102–108
Elavarasan J, Velusamy P, Ganesan T, Ramakrishnan SK, Rajasekaran D, Periandavan K (2012) Hesperidin-mediated expression of Nrf2 and upregulation of antioxidant status in senescent rat heart. Journal of Pharmacy and Pharmacology 64:1472–1482
FAO W (2002) Health implications of acrylamide in food. FAO/WHO Joint Commission, Switzerland
Fouad AA, Abdel-Gaber SA, Abdelghany MI (2019) Hesperidin opposes the negative impact of cyclophosphamide on mice kidneys. Drug and Chemical Toxicology:1–6
Friedman M (2003) Chemistry, biochemistry, and safety of acrylamide. A review. J Agric Food Chem 51:4504–4526
Ghasemian S, Rezaei K, Abedini R, Poorazarang H, Ghaziani F (2014) Investigation of different parameters on acrylamide production in the fried beef burger using Taguchi experimental design. J Food Sci Technol 51:440–448
Green LC, Wagner DA, Glogowski J, Skipper PL, Wishnok JS, Tannenbaum SR (1982) Analysis of nitrate, nitrite, and [15N]nitrate in biological fluids. Analytical biochemistry 126:131–138
Hamdy SM, Shabaan AM, Abdel Latif AKM, Abdel-Aziz AM, Amin AM (2017) Protective effect of hesperidin and Tiger nut against acrylamide toxicity in female rats. Experimental and Toxicologic Pathology 69:580–588
Hayyan M, Hashim MA, AlNashef IM (2016) Superoxide ion: generation and chemical implications. Chemical reviews 116:3029–3085
Hsu C-C, Lin MH, Cheng JT, Wu MC (2017) Antihyperglycaemic action of diosmin, a citrus flavonoid, is induced through endogenous β-endorphin in type I-like diabetic rats. Clinical and Experimental Pharmacology and Physiology 44:549–555
Jantet G (2002) Chronic venous insufficiency: worldwide results of the RELIEF study. Reflux assEssment and quaLity of lIfe improvEment with micronized Flavonoids. Angiology 53:245–256
Kang SR, Park KI, Park HS, Lee DH, Kim JA, Nagappan A, Kim EH, Lee WS, Shin SC, Park MK, Han DY, Kim GS (2011) Anti-inflammatory effect of flavonoids isolated from Korea Citrus aurantium L. on lipopolysaccharide-induced mouse macrophage RAW 264.7 cells by blocking of nuclear factor-kappa B (NF-κB) and mitogen-activated protein kinase (MAPK) signalling pathways. Food Chemistry 129:1721–1728
Kim H, Lee SG, Rhie J (2017) Dermal and neural toxicity caused by acrylamide exposure in two Korean grouting workers: a case report. Annals of Occupational and Environmental Medicine 29:1–6
Kim S-H, Kim B-K, Lee Y-C (2011) Antiasthmatic effects of hesperidin, a potential Th2 cytokine antagonist, in a mouse model of allergic asthma. Mediators of Inflammation 2011:1–12
Kouhpayeh A, Mirkhani H, Nekooeian AA (2008) Improvement of endothelium-dependent relaxation in aorta of rat models type 1 and 2 diabetes by hespiridin. Iranian Cardiovascular Research Journal 1:200–207
Kuntić V, Brborić J, Holclajtner-Antunović I, Uskoković-Marković S (2014) Evaluating the bioactive effects of flavonoid hesperidin--a new literature data survey. Vojnosanitetski pregled 71:60–65
Larsen K (1972): Creatinine assay in the presence of protein with LKB 8600 Reaction Rate Analyser. Clinica chimica acta; international journal of clinical chemistry 38, 475-6
Li X, Hu X, Wang J, Xu W, Yi C, Ma R, Jiang H (2018): Inhibition of autophagy via activation of PI3K/Akt/mTOR pathway contributes to the protection of hesperidin against myocardial ischemia/reperfusion injury. International Journal of Molecular Medicine, 1917-1924
Lobo V, Patil A, Phatak A, Chandra N (2010) Free radicals, antioxidants and functional foods: Impact on human health. Pharmacognosy reviews 4:118–126
LoPachin RM, Ross JF, Lehning EJ (2002) Nerve terminals as the primary site of acrylamide action: a hypothesis. Neurotoxicology 23:43–59
Mahmoud AM, Ashour MB, Abdel-Moneim A, Ahmed OM (2012) Hesperidin and naringin attenuate hyperglycemia-mediated oxidative stress and proinflammatory cytokine production in high fat fed/streptozotocin-induced type 2 diabetic rats. Journal of Diabetes and its Complications 26:483–490
Maneesai P, Bunbupha S, Potue P, Berkban T, Kukongviriyapan U, Kukongviriyapan V, Prachaney P, Pakdeechote P (2018) Hesperidin prevents nitric oxide deficiency-induced cardiovascular remodeling in rats via suppressing TGF-β1 and MMPs protein expression. Nutrients 10:1549
Mihara M, Uchiyama M (1978) Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Analytical biochemistry 86:271–278
Mottram DS, Wedzicha BL, Dodson AT (2002) Acrylamide is formed in the Maillard reaction. Nature 419:448–449
Nishikimi M, Appaji Rao N, Yagi K (1972) The occurrence of superoxide anion in the reaction of reduced phenazine methosulfate and molecular oxygen. Biochemical and Biophysical Research Communications 46:849–854
Nita M, Grzybowski A (2016) The role of the reactive oxygen species and oxidative stress in the pathomechanism of the age-related ocular diseases and other pathologies of the anterior and posterior eye segments in adults. Oxidative Medicine and Cellular Longevity 2016:1–23
Paglia DE, Valentine WN (1967) Studies on the quantitative and qualitative characterization of erythrocyte glutathione peroxidase. The Journal of laboratory and clinical medicine 70:158–169
Parhiz H, Roohbakhsh A, Soltani F, Rezaee R, Iranshahi M (2015) Antioxidant and anti-inflammatory properties of the citrus flavonoids hesperidin and hesperetin: an updated review of their molecular mechanisms and experimental models. Phytotherapy Research 29:323–331
Petrova A, Davids LM, Rautenbach F, Marnewick JL (2011) Photoprotection by honeybush extracts, hesperidin and mangiferin against UVB-induced skin damage in SKH-1 mice. Journal of Photochemistry and Photobiology B: Biology 103:126–139
Ramsay EE, Dilda PJ (2014) Glutathione S-conjugates as prodrugs to target drug-resistant tumors. Frontiers in pharmacology 5:181–181
Reitman S, Frankel S (1957) A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. American journal of clinical pathology 28:56–63
Sahu BD, Kuncha M, Sindhura GJ, Sistla R (2013) Hesperidin attenuates cisplatin-induced acute renal injury by decreasing oxidative stress, inflammation and DNA damage. Phytomedicine 20:453–460
Sakata K, Hirose Y, Qiao Z, Tanaka T, Mori H (2003) Inhibition of inducible isoforms of cyclooxygenase and nitric oxide synthase by flavonoid hesperidin in mouse macrophage cell line. Cancer letters 199:139–145
Selvaraj P, Pugalendi KV (2010) Hesperidin, a flavanone glycoside, on lipid peroxidation and antioxidant status in experimental myocardial ischemic rats. Redox Report 15:217–223
Shaban NZ, Ahmed Zahran AM, El-Rashidy FH, Abdo Kodous AS (2017) Protective role of hesperidin against γ-radiation-induced oxidative stress and apoptosis in rat testis. Journal of Biological Research-Thessaloniki 24:5–5
Stadler RH, Blank I, Varga N, Robert F, Hau J, Guy PA, Robert M-C, Riediker S (2002) Acrylamide from Maillard reaction products. Nature 419:449–450
Szymański M, Młynarek D, Szymański A, Matławska I (2016) Simultaneous determination of diosmin and hesperidin in pharmaceuticals by RPLC using ionic liquids as mobile phase modifiers. Iranian journal of pharmaceutical research : IJPR 15:141–148
Tahir M, Rehman MU, Lateef A, Khan R, Khan AQ, Qamar W, Ali F, O'Hamiza O, Sultana S (2013) Diosmin protects against ethanol-induced hepatic injury via alleviation of inflammation and regulation of TNF-α and NF-κB activation. Alcohol 47:131–139
Tietz NW, Burtis CA, Duncan P, Ervin K, Petitclerc CJ, Rinker AD, Shuey D, Zygowicz ER (1983) A reference method for measurement of alkaline phosphatase activity in human serum. Clinical chemistry 29:751–761
Turk E, Kandemir FM, Yildirim S, Caglayan C, Kucukler S, Kuzu M (2019) Protective effect of hesperidin on sodium arsenite-induced nephrotoxicity and hepatotoxicity in rats. Biol Trace Elem Res 189:95–108
Uthra C, Shrivastava S, Jaswal A, Sinha N, Reshi MS, Shukla S (2017) Therapeutic potential of quercetin against acrylamide induced toxicity in rats. Biomedicine and Pharmacotherapy 86:705–714
Vasco CF, Watanabe M, Fonseca CD, MdFF V (2018) Sepsis-induced acute kidney injury: kidney protection effects by antioxidants. Revista Brasileira de Enfermagem 71:1921–1927
Venkatasubbaiah K, Venkataswamy M, Sandhya D, Suresh K, Rao KJ (2014) Acrylamide induced oxidative stress in rat and chick embryonic liver tissues. Indo American Journal of Pharm Research:4
Wei D, Ci X, Chu X, Wei M, Hua S, Deng X (2012) Hesperidin suppresses ovalbumin-induced airway inflammation in a mouse allergic asthma model. Inflammation 35:114–121
Wojnar W, Kaczmarczyk-Sedlak I, Zych M (2017) Diosmin ameliorates the effects of oxidative stress in lenses of streptozotocin-induced type 1 diabetic rats. Pharmacological Reports 69:995–1000
Yousef MI, El-Demerdash FM (2006) Acrylamide-induced oxidative stress and biochemical perturbations in rats. Toxicology 219:133–141
Yumnam S, Hong GE, Raha S, Saralamma VVG, Lee HJ, Lee W-S, Kim E-H, Kim GS (2016) Mitochondrial dysfunction and Ca 2+ overload contributes to hesperidin induced paraptosis in hepatoblastoma cells, HepG2. Journal of Cellular Physiology 231:1261–1268
Zhang L, Wang E, Chen F, Yan H, Yuan Y (2013) Potential protective effects of oral administration of allicin on acrylamide-induced toxicity in male mice. Food & Function 4:1229–1229
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Acknowledgments
This project was funded by Research supporting project number (RSP-2019/95) king Saud University,Riyadh,Saudi Arabia.
Additional information
Responsible editor: Philippe Garrigues
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Elhelaly, A.E., AlBasher, G., Alfarraj, S. et al. Protective effects of hesperidin and diosmin against acrylamide-induced liver, kidney, and brain oxidative damage in rats. Environ Sci Pollut Res 26, 35151–35162 (2019). https://doi.org/10.1007/s11356-019-06660-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-019-06660-3