
Abstract
Fipronil (FIP) is a highly effective, broad-use insecticide that belongs to the phenylpyrazole chemical group. It is extensively used in the agriculture and veterinary medicine for controlling a wide variety of pests. Though FIP showed lower toxicity in vertebrates than in insects, it was recognized to have a variety of toxic effects in mammals. The present study was undertaken to evaluate FIP-induced alterations in the blood biochemical markers and oxidative stress parameters in male albino mice via oral sub-acute toxicity exposure. The possible ameliorative effect of the pretreatment with selenium plus α-tocopherol (vitamin E) against the harmful effects of FIP was also investigated. Mice in FIP-test groups were exposed to different sublethal doses, i.e., 1.43, 2.87, and 4.78 mg active ingredient (AI)/kg body weight (b.w.), equal to 1/100, 1/50, and 1/30 LD50 of FIP, respectively, for 28 days. Mice in the amelioration groups were orally administered with selenium + vitamin E (0.3 mg + 22.5 mg/kg b.w., respectively) 14 days prior to exposure to the higher dose (4.78 mg/kg) of FIP for another 14 days. Fipronil exposure at medium and high doses showed lowered values of red blood cell count (RBC), hematocrit (HCT), hemoglobin (HGB), white blood cell (WBC), and platelet (PLT) counts after 28-day exposure, compared to the control. All three doses caused significant increases in levels of liver-function biomarkers, i.e., aspartate amino transaminase (AST), alanine amino transaminase (ALT), alkaline phosphatase (ALP), cholesterol, and bilirubin levels compared to the control. Levels of biomarkers related to kidney functions, i.e., urea, uric acid, and creatinine, increased significantly than these of the control. Likewise, the oxidative stress indices, i.e., hydrogen peroxide (H2O2) and malondialdehyde (MDA), significantly increased at the higher and medium doses, while antioxidant enzymes, catalase (CAT) and superoxide dismutase (SOD), decreased significantly. On the other hand, prior administration of selenium + vitamin E in the FIP-exposed mice led to restore values of most hematological parameters nearly to these of the control. Also, the levels of AST, total protein, and creatinine seemed to be restored to the control values. Interestingly, pretreatment with selenium + vitamin E restored the levels of antioxidant enzymes, CAT and SOD, to the control values, whereas, oxidative stress indices, H2O2 and MDA, remained significantly high. It is our thought that the sublethal dose less than 1.43 mg/kg b.w. of commercial formulation of FIP (COACH® 200 SC) could be considered as no-observed-adverse-effect-level(NOAEL) under our present experimental conditions at short-term toxicity study. On the other hand, the higher sublethal doses, 4.78 and 2.87 mg/kg b.w., induced significant adverse effects in biomarkers and may be deleterious to human health following long-term exposure.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Fipronil (FIP), (5-amino-1-[2, 6-dichloro- 4(trifluoromethyl) phenyl]-4-[(trifluoromethyl) sulfinyl]-1H-pyrazole-3-carbonitrile)), is broad-spectrum insecticide that belongs to the second-generation phenylpyrazole chemical family. Recently, it has been widely used to control an extensive species of agricultural crop pests and also public hygiene, and veterinary pests (Tingle et al. 2003). It is well known that FIP acts on the nervous system through a non-competitive binding affinity to the gamma-aminobutyric(GABA) receptor, and therefore blocking the normal passage of chloride ions and the transmission of normal neural impulse causing paralysis and finally death (Kidd and James 1991). The lower binding affinity for mammalian receptors enhances selectivity against insects and increases its boundary of safety (Hainzl et al. 1998). In spite of its obviously positive toxicological profile for mammals, FIP could have undesirable effects independent on non-neuronal sites like kidney, liver, and thyroid and influence reproductive function (Khan et al. 2015; de Oliveira et al. 2012; Leghait et al. 2010; Ohi et al. 2004) in non-target organisms. Liver, kidney, and thyroid seemed to the most affected organs showing toxic effects on chronic exposure of fipronil in rats (Tingle et al. 2003; Badgujar et al. 2015). Short-term exposure to fipronil induced oxidative stress in the kidney, brain, and liver of mice (Badgujar et al. 2015, 2016). Several researchers have expressed concern of probable undesirable human public health effects due to extensive use of FIP in commercial and for home applications (Tingle et al. 2003; Jennings et al. 2002a, b).
Chronic toxicity study using FIP-technical grade revealed a number of toxicological effects at relatively low doses (Tingle et al. 2016). An important environmental concern of FIP exposure has been reported by Tingle et al. (2003) who stated that cyclic, systemic applications of FIP have bioaccumulative effects which in turn affect all animals along the trophic chain. Hematological indicators are considered significant biomarker for suggesting altered internal and/or external environment of animals, and variations in their indicators within an individual may cause insufficient responses to chemical stressors; however, these variations are nonspecific to a wide scope of substances (Prashanth and David 2006). Hence, exposure to pesticides is supposed to increase or decrease in hematological levels frequently influencing the survival of exposed organisms (Kori-Siakpere and Oboh 2011). The biological markers including both hematological and biochemical parameters are exercised as good indicators of health conditions and outcome from the pesticide-induced toxicity within organisms (Thrall 2004; Pimpao et al. 2007).
Serum aminotransferase levels, alanine aminotransferase (ALT), aspartate aminotransferase (AST), and alkaline phosphatase (ALP), are considered the most useful measures of liver cell injury. These components and the end products of the metabolic pathways are highly sensitive for deciding the incurred unusual and have been considered as biochemical markers of liver dysfunction (Schomaker et al. 2013). Increase in the levels of serum AST, ALT, and ALP reveals the loss of biochemical and basic consistency of the liver and is the most favored indicative tool to estimate hepatocellular damage (Giannini et al. 2005). On the other hand, the toxic effects of pesticides might be correlated with increased production of reactive oxygen species (ROS) which sequentially induce pathological damage to the liver, kidneys and brain tissues (Verma et al. 2007; El-Gendy et al. 2010). It is well known that oxidative stress happens once the production of ROS overcomes antioxidant capacity in the target cells or biological system, in which membrane lipid peroxidation is one of the most serious types of oxidative stress (Scandalios 2005). The antioxidant defense mechanism in animals is comprised of enzymes including catalase (CAT), superoxide dismutase (SOD), glutathione peroxidase (GPx), and non-enzymatic antioxidants, i.e., non-protein thiols, particularly glutathione (GSH). Therefore, the activities of enzymatic antioxidants, as well as the incidence of oxidative damage, have been suggested to be good indicators of oxidative stress induced by pollutants including pesticides (Li et al. 2003; Ahmad et al. 2000). The enzymatic- and non-enzymatic antioxidants were found to be markedly affected by pesticide exposure (Banerjee et al. 2001). In vitro study revealed that fipronil induced oxidative stress in the neuronal cell lines (Ki et al. 2012). However, a few studies have been done to investigate the effects of fipronil on oxidative stress in an animal model (in vivo). Recently, fipronil was found to cause significant increases in the activities of serum aminotransferases, AST and ALT, with connected increase in the absolute and relative weight of liver (Badgujar et al. 2015). Moreover, high dose of FIP caused significant increase in the lipid peroxidation (LPO) and decrease in the activity of antioxidant enzymes, SOD, CAT, and GST as well as the content of non-antioxidant enzymes, glutathione, and total thiol (Badgujar et al. 2015). Similarly, Mossa et al. (2015) reported that FIP induced LPO, oxidative stress, and liver and kidney injury in male albino rats. Many researchers have reported the protective/ameliorative effects of a large number of antioxidants, such as selenium, vitamins E and C, and plant derivatives against pesticide-induced toxicity (Singh et al. 2008; El-Shenawy et al. 2010; Mossa et al. 2011; El-Desoky et al. 2012; Badgujar et al. 2016, 2017). Vitamins E and C were able to scavenge free radicals produced by normal metabolic processes in organisms as well as by environmental pollutants, particularly pesticides, and stop their attack on polyunsaturated fats (Laurence et al. 1997). Vitamin E has been reported to be the most effective lipid-soluble antioxidant found in the biological system that scavenges ROS, thus preventing LPO and the instigation of oxidative tissue damage (Verma et al. 2007). Pretreatment of rats with vitamin E provided significant protection against the elevation of malondialdehyde (MDA) concentrations, a biomarker of lipid peroxidation, in cerebral and hepatic tissues, induced by cypermethrin (Giray et al. 2001). Selenium (Se) is an essential dietary trace element, which plays a vital role as an antioxidant and as an integral part of many proteins with catalytic and structural functions. Its nutritional deficiency leads to muscular dystrophy, endemic fatal cardiomyopathy (Keshan disease), and chronic degenerative diseases in humans that could be prevented by Se supplementation when used alone or in combination (Rayman 2008). The most important metabolic roles of Se in mammalian cells are due to its function in the active site of many antioxidant enzymes, for example thioredoxin reductase and glutathione peroxidase (GPx)(Flora et al. 2002). Selenium, in the form of GSH-Px, plays an important role in the protection of tissue from oxidative damage. Aside from being an integral part of GSH-Px, Se can antagonize the toxic effects of some chemical substances (Combs and Combs 1986; Raneva et al. 2002). On the other hand, increases of dietary and tissue concentration of selenium increase plasma and hepatic GSH-Px activities, followed by ultimately hepatic lipid peroxidation (Stewart et al. 1999; Zia and Islam 2000; Hoffman 2002). The use of selenium prior to cypermethrin was observed to increase both MDA concentrations and antioxidant systems such as GSH-Px and CAT activities (Ateşşahin et al. 2012). On the other hand, vit E plus selenium could cause complex alterations in the antioxidant system and adversely could alleviate the oxidative stress induced by cypermethrin (Ateşşahin et al. 2012). Administration of vitamin C and selenium exhibited protective effect by inhibiting methomyl-induced toxicity in liver and kidney (Djeffal et al. 2012). Based on the above information, we hypothesize that vitamin E and/or selenium can reduce FIP-induced toxicity.
Therefore, the present study was carried out to investigate the effect of commercial formulation of fipronil “COACH®” on several biomarkers associated with liver and kidney functions, and these of oxidative stress of mice after 28 days repeated dose exposure. Another goal of the study was to evaluate the possible protective role of pre-treatment of selenium (plus vitamin E) against fipronil-induced toxicity. Additionally, the no-observed-adverse-effects level (NOAEL) of FIP formulation was also estimated.
Materials and methods
Chemicals
Commercial product of fipronil (COACH®, 200 SC) used in this study was purchased from the local market of pesticides, Shoura Chem. Co., Egypt. The chemicals used in biochemical analyses were of commercially available biodiagnostic grade kits and purchased locally from Diagnostic & Research Reagents Co., Dokki, Giza, Egypt. Selenium was obtained from a local veterinary pharmaceutical company (Movartis-Agro, Egypt), which marketed commercially sodium selenite (Na2SeO3) mixed with vitamin E (α-tocopherol acetate) (100 + 7500 mg/L, respectively).
Animal’s husbandry
The study was conducted on adult albino mice, 8–10 weeks of age, weighing 25–30 g obtained from the Egyptian Company for Veterinary Drugs and Vaccines (VACSERA), Helwan Farm, Egypt. The animals were housed in groups of four to five animals in each polycarbonate cage (12″ × 10″ × 8″) containing a layer of sawdust that was changed every 2 days to maintain hygienic conditions under controlled conditions of temperature (26 ± 2 °C), humidity (40–70%), and photoperiod (Light: Dark12:12 h), with free access to water. The animals were fed daily on a commercial pelleted diet purchased from a local company (Ayad Company, Sadat City, Egypt). The diet provides essential nutrients, with approximately 17.51% proteins, 5% fats, 62% carbohydrates, 3.3% calcium, and 0.48% phosphorus, to meet the requirements of breeding animals. The mice were acclimatized for 2 weeks in our lab, at Department of Pesticides, before using them for experimentation. The handling procedures of mice were conducted in strict conformation with the guidelines and welfare regarding animal protection approved by the local institute ethical committee of our institution.
Experimental design
Oral acute toxicity test—24-h LD50
The acute toxic method as described in the Organization for Economic Cooperation and Development (OECD) Guideline (OECD 1998) was used to estimate LD50 (the median lethal dose of each pesticide). Healthy young adult males weighing 30 ± 2 g were fasted for 8 h prior to dosing. For preliminary toxicity tests, various diluted doses of each pesticide chosen with respect to previously reported oral LD50 were screened for determination of maximum tolerated dose (MTD) in pilot dose range. Oral gavage needle was used for administration of pesticide dose in mice. The pilot study so called up-and-down procedure was followed as described by Bruce (1985). Based on the results of pilot study, we performed confirmatory experiments according to OECD technique (1998). The animals were organized into five groups of five individuals each. The control group received only distilled water, while the test groups received the test doses diluted or suspended in distilled water via single oral administration technique using gavage needle. The selected doses of fipronil used for toxicity test ranged from 33.3 to 333.0 mg AI/kg b.w. Then, mortality from 0 to 100% was calculated after 24 h. A dose-response relationship was calculated to estimate values of LD50, 95% confidence limits (CLs), and slope (b), using Probit and Logit Analysis software program (Polo Plus, ver. 2.0, 2008) based on Finney analysis (Finney 1971).
Clinical symptoms
The different toxicity symptoms induced in response to exposure to each dose such as salivation, lacrimation, diarrhea, tremor, convulsion, fatigue and death were observed in each group for early 3 h and then once daily throughout 14 days following the exposure.
Repeated dose 28-day sub-acute toxicity study
Subacute-oral toxicity tests were carried out based on the procedure described by OECD Guidelines for the Testing of Chemicals (OECD 2008). This test provides information on the possible health hazards or toxicities which may arise from repeated exposure, over a relatively short period of time (28 days). Subacute toxicity tests are also undertaken as preliminary testing to longer-term toxicity studies. In this experiment, adult male mice, weighing 30 ± 2 g, were organized into four groups of at least 10 individuals in each polycarbonate cage, under controlled conditions of temperature and photoperiod with free access to food and water as described above. Adult male mice were divided into four groups. One group served as control and was given distilled water through oral gavage. The other three groups were given the selected sublethal doses, i.e., 1.44, 2.87, and 4.78 mg AI/kg b.w./day (corresponding to 1/100, 1/50, and 1/30 LD50 of FIP, respectively) suspended in water for 28 consecutive days. The test doses were calculated from the percentage of active ingredients of the commercial formulation. Two replicates were used for each group. Body weight and clinical signs of toxicity were recorded throughout the period of experiment.
Protective effect of selenium plus α-tocopherol against FIP-induced toxicity
In this experiment, the possible protective effects of pre-treatment with selenium + α-tocopherol (vitamin E) against toxicity of FIP were evaluated. Adult male mice were segregated into four groups, each group having 18–20 animals, in two replicates. Group 1 served as negative control where mice received only distilled water; in group 2, the mice received FIP alone (at 4.78 mg/kg b.w./day) for 28 days; in group 3, the animals received only Se + Vit E (at 0.3 mg + 22.5 mg/kg b.w,, respectively) for 28 days; and in group 4, the mice received Se + Vit E (at 0.3 mg + 22.5 mg/kg b.w., respectively) for 14 days prior to administration with FIP (at 4.78 mg/kg b.w./day) for another 14 days.
Hematological evaluation
At the end of the experiment (on the 28th day), all survived animals of each group were sacrificed, and blood was taken directly from the heart using heparinized hypodermic syringe and collected in a test tube rinsed with heparin saline solution as anticoagulant, in the proportion of 50 μl for each 1.0 mL of blood and used for the complete blood count. Whole blood was used to count leucocytes (white blood cells), red cells (RBCs), and platelets (PLTs), and also determine hemoglobin (HGB) and hematocrit (HCT) levels. The analyses were performed using an auto hematology analyzer (Swelab Alfa hematology analyzers, Boule Diagnostics AB, Domnarvsgatan 4, Sweden).
Biochemical analysis
Blood serum was used for clinical biochemical determinations. To collect the serum, blood samples were left to clot in clean dry tubes and centrifuged at 1500×g for 10 min at 4 °C to obtain the sera. The clear serum samples were carefully aspirated off with a fine-bore pipette to avoid extracting red cells and stored frozen at − 20°C until using for biochemical analysis. All biochemical analyses were conducted according to the manufacturers’ instructions of the corresponding specific biodiagnostic kits (Diagnostic & Research Reagents Co., Dokki, Giza, Egypt).
Liver and kidney function biomarkers
The activities of serum aminotransferases, i.e., aspartate aminotransferase (AST) and alanine aminotransferase (ALT), were determined colorimetrically (at 450 nm) according to the methods of Reitman and Frankel (1957). For AST, the transfer of an amino group from aspartate to α-ketoglutarate results in the generation of glutamate, resulting in the production of a colorimetric product proportional to the AST enzymatic activity present. For ALT assay, it catalyzes the reversible transfer of an amino group from alanine to α-ketoglutarate, generating pyruvate and glutamate. Diagnostically, the activities of these enzymes are almost measured in units/liter (U/L). The activity of alkaline phosphates (ALPs) was determined based on the kinetic assay described by Wenger et al. (1984). ALP catalyzes the hydrolysis of phenyl phosphate at pH 10.4, liberating phenol and phosphate. The rate of p-nitrophenol formation, measured photometrically (at 405 nm), is proportional to the catalytic concentration of alkaline phosphatase present in the sample, which is expressed as units per liter of sample (U/L). Serum cholesterol was determined according to the method of Roschlau et al. (1974). Cholesterol assay is based on cholesterol esterase hydrolysis of cholesterol esters to form free cholesterol and cholesterol dehydrogenase catalyzed conversion of cholesterol to cholest-4-ene3-one, in which NAD is reduced to NADH. The optical density of the formed NADH at 340 nm is directly proportionate to the cholesterol concentration in the sample. Total serum cholesterol is expressed as mg/dL. The total protein content was determined by the method of Bradford and Marion (1976) which is based on employing the dye, Brilliant Blue G. When dissolved in an acid-alcoholic medium, the Brilliant Blue G dye reacts almost immediately with protein to form a blue-colored protein dye complex. The protein-dye complex causes a shift in the absorption maximum of the dye from 465 to 595 nm. The amount of absorption at 595 nm is proportional to the protein present. Protein concentration was expressed as mg/ml. The level of serum bilirubin was determined by following the Malloy and Evelyn method (1937). Bilirubin is converted to colored diazotized sulfanilic acid and measured photometrically (at 510–550 nm). The total bilirubin concentration was expressed as mg/dL.
Creatinine test is most widely used to assess kidney function. The Henry method (Henry 1974) was followed to determine the level of serum creatinine. The method was based on the reaction of creatinine with alkaline picrate. The assay utilizes a kinetic absorbance method to overcome interference by colored compounds in serum. In this method, creatinine reacts with picric acid in alkaline conditions to form a color complex which absorbs at 492 nm. Serum urea concentration (mg/dL) was determined according to the method of Patton and Crouch (1977). The reaction based on hydrolysis of urea in the presence of water and urease to produce ammonia and carbon dioxide. The free ammonia in the presence of indicator forms a colored complex, measured at 557 nm, which is directly proportional to the urea concentration in the serum. Uric acid concentration in serum was estimated by the method of Fossati et al. (1980). This assay is based upon the methods of modified trinder peroxidase assay using 3,5-dichloro-2-hydroxy-benzene-sulforic acid (DCHB). The assay was based on the oxidation of uric acid to allantoin by uricase production of hydrogen peroxide which reacts with 4-amino-antipyrine and DCHB in the presence of peroxidase to yield a quinone imine dye. The subchange in absorbance at 546 nm is proportional to uric acid concentration in the sample.
Oxidative stress and enzymatic antioxidant biomarkers
Determination of oxidative stress biomarkers, i.e., lipid peroxidation and hydrogen peroxide, and enzymatic-antioxidants, i.e., catalase (CAT) and superoxide dismutase (SOD), were performed according to the details given in BioDiagnostic kit’s instructions (Diagnostic & Research Reagents Co., Dokki, Giza, Egypt). Oxidative damage was estimated by measuring the degree of lipid peroxidation. Malondialdehyde (MDA) formation of thiobarbituric acid-reactive substances (TBARSs) which was expressed in forms of malondialdehyde content according to the method of Yagi (1998). Though this assay is sensitive and widely used, it is not specific and TBA reacts with a number of components present in biological samples. In this technique, lipid peroxidation is determined by the reaction of MDA with thiobarbituric acid (TBA) to form a colorimetric (532 nm) product, proportional to the MDA present. MDA concentration was expressed as nmol/ml. To estimate concentration of serum hydrogen peroxide, the method of Nourooz-Zadeh et al. (1994) was followed. In brief, this assay utilizes the chromogenic Fe3+-xylenol orange reaction, in which a purple complex is formed when Fe2+ is oxidized to Fe3+ by peroxides present in the sample, generating a colorimetric (585 nm) result, proportional to the level of peroxide present. The concentration of serum hydrogen peroxide was expressed as ng/ml.
Catalase (CAT) was assayed by the composition of hydrogen peroxide according to the method of Aebi (1984). Catalase reacts with a known quantity of H2O2; the reaction is stopped after exactly 1 min with catalase inhibitor. In the presence of peroxidase (HRP), the remaining H2O2 reacts with 3,5-dichloro-2-hydroxybenzene sulfonic acid (DHBS) and 4-aminophenazone(AAP) to form a chromophore with a color intensity proportional to the amount of catalase in the original sample. The change in absorbance of reaction solution was determined at 510 nm. The catalase activity was calculated and expressed as U/mL. The superoxide dismutase (SOD) activity was estimated by the method of Marklund and Marklund (1974). In the assay, superoxide (O2-) is provided by xanthine oxidase (XO) catalyzed reaction. O2– reacts with a WST-1 dye to form a colored product. The color intensity (OD440nm) is used to determine the SOD activity in a sample. The SOD activity was expressed as U/mL.
All the above biochemical analyses were conducted using UV-Spectrophotometer (model: B01-CT-2200 Spectrophotometer, E-Chrom Tech Co., Taiwan).
Statistical analysis
Biochemical analyses were presented as the mean ± standard deviation of means (SD). Comparisons were made between control and treated groups using one-way analysis of variance (ANOVA) method. The least significant difference (LSD) test was used for mean separation at p < 0.05. All statistical analyses were made using the software SPSS (version 22.0).
Results
Acute toxicity and clinical symptoms
The toxicity data of fipronil formulation (FIP) showed that the LD50 value obtained after 24 h was 143.50 mg AI/ kg b.w. with 95% confidence limits (84.9–203.9). The regression equation was: Y = 2.790 + b×, and the slope (b) was 3.48 ± 1.213. The single dose-acute toxicity assays of FIP showed that the main clinical symptoms following exposure were generally observed at within 24 h of treatment, and the major toxicity signs included hyperactivity, tremors and convulsions 7of various types, effects on activity or gait, hunched posture, wetness in various body areas, and seizures. It was also obvious that the toxicity signs did not reach their peak until 7 h after dosing.
Sub-acute toxicity study
Effects of sublethal doses of Fipronil
After repeated dose 28-day exposure to fipronil (FIP), the results of the blood profile (Table 1) showed that erythrocyte indices including mean cell volume (MCV) and mean cell hemoglobin (MCH) were significantly decreased in the FIP groups exposed to 1/20 and 1/50 LD50, as compared to these in the control. The data also revealed that levels of hemoglobin (HGB), red blood cells (RBCs), white blood cells (WBCs), and hematocrit (HCT) levels were significantly decreased. The lymphocyte (LYM) and platelet (PLT) levels were also significantly decreased. It was obvious that the decreased levels were in dose-dependent manner.
The data presented in Table 2 showed clearly that FIP induced hepatoxicity and nephrotoxicity as demonstrated by the elevation of serum liver and kidney biomarkers. A high significant increase was found in the activities liver enzymes and aminotransferases, i.e., aspartate aminotransferase (AST) and alanine aminotransferase (ALT), in the group exposed to the higher dose (1/30 LD50) compared to those of the control. It was obvious that increasing rate was in a dose-dependent manner. Likewise, alkaline phosphatase (ALP) activity was also significantly increased in all FIP-groups, in a dose-dependent manner. The data also showed high significant increase in the total contents of bilirubin, protein, and cholesterol as compared to these in the negative control.
For the overall kidney function markers, the results in Table 2 showed significant increases in creatinine levels in mice that received 1/30 and 1/50 LD50 of FIP compared to that of the control. The increased levels were in a dose-dependent manner. The similar trend was observed for urea levels, where all sublethal doses, 1/30, 1/50, and 1/100 LD50, significantly (P< 0.05) increased urea levels. The data also revealed that FIP induced high significant increase in the levels of uric acid, in a dose-dependent manner.
The data of oxidative stress indices revealed that high significant increase (P< 0.05) in hydrogen peroxide (H2O2) levels in mice exposed to FIP at doses of 1/30 and 1/50 LD50, compared to that of the control (Table 3). Similar trend was also reported for malondialdehyde (MDA), but increasing levels seemed to be much lower than these for hydrogen peroxide (Table 3). The increased percentages for either H2O2 or MDA were in dose-dependent manner. On the other hand, the levels of antioxidant enzymes, catalase (CAT) and superoxide dismutase (SOD), are presented in Table 3. The results showed that FIP-repeated exposure, at the higher dose, 4.78 mg/kg b.w. (1/30 LD50), induced significant decreases in both CAT and SOD, in comparison with these of the control. Slight insignificant decrease was also observed in CAT and SOD by the other sublethal doses, 1/50 and 1/100 LD50.
Ameliorative effect of pre-treatment with selenium (plus vitamin E)
The data presented in Tables 4, 5, and 6 show the protective effects of pre-treatment administration of male mice with selenium (plus vitamin E) against the adverse effects induced by high dose of fipronil. The results in Table 4 demonstrated that 28-day repeated exposure to FIP alone, at 1/30 LD50, induced high significant decreases (푃 < 0.05) in the levels of erythrocyte count, RBC, HGB, MCV, MCH, and HCT than these of the negative control. Interestingly, pre-treatment of mice via oral administration with selenium (plus vitamin E), for 2 weeks, showed beneficial effects by increasing the levels of hematological parameters: RBC, HGB, MCV, MCHC, and HCT. Hemoglobin (HGB) and red blood cell size (MCV) were mostly recovered to the normal values in case pre-treatment with selenium plus vitamin E. Similar trend was also observed for the levels of both white blood cells (WBCs) and lymphocytes (LYMs). On the contrary, the parameter of hemoglobin concentration per red blood cell (MCHC) was significantly increased in mice received FIP alone as compared to the control group. Remarkably, pre-treatment of animals with selenium (plus vitamin E) before exposure to FIP recovered the concentrations of MCHC near to the control levels. However, pre-treatment with selenium (plus vitamin E) did not recover platelet concentration (PLT) to be like that of the control.
The effects of FIP intoxication and pre-treatment with selenium (plus vitamin E) on biochemical parameters associated with liver and renal functions are shown in Table 5. It is obvious that FIP induced hepatoxicity and nephrotoxicity as demonstrated by the elevation of serum liver and kidney biomarkers. The AST, ALT, ALP, bilirubin, protein, cholesterol, urea, creatinine, and uric acid levels were substantially increased (P ≤ 0.05) in response to FIP administered orally alone to mice, compared to those of the control group. However, these parameters were significantly reduced (P ≤ 0.05) when mice were pre-treated with selenium (plus vitamin E) suggesting amelioration of FIP-induced toxicity.
For oxidative stress indices, the results in Table 6 revealed that repeated-dose exposure of FIP at high dose (1/30 LD50) induced high significant increase in the level of hydrogen peroxide (H2O2) compared to that of the control. However, pre-treatment of mice with selenium + vitamin E for 2 weeks showed beneficial effect by reducing the increasing level of H2O2 from 0.646 (in case of FIP alone) to 0.381 ng/mL, in comparison with that of the control (0.199 ng/mL). The data also showed similar trend for the other oxidative stress marker, i.e., malondialdehyde (MDA), where significant increase in its level was observed in animals received FIP alone compared to that of the control. However, slight improvement in the levels of MDA was observed in the group pre-treated with selenium (plus vitamin E) prior exposure to FIP, by reducing the increasing level of MDA from 9.15 (in case of FIP alone) to 7.04 nmol/mL, in comparison with that of the control (5.19 nmol/mL).
On the other hand, the results in Table 6 show the protective effects of pre-treatment administration of mice with selenium (plus vitamin E) on ameliorating the negative effects of FIP on two important antioxidant enzymes, CAT and SOD. It was clear that repeated exposure of mice to the higher sub-lethal dose of FIP significantly decreased the levels of both CAT and SOD, in comparison with these in the negative control. Interestingly, the pre-treatment of mice with selenium (plus vitamin E) showed better effects by restoring the levels of both CAT and SOD to be nearly like these of the control group.
Discussion
The acute toxicity of commercial product of fipronil (FIP) “COACH® 200 SC” used in the present study showed that the 24-h LD50 was 143.50 mg AI/kg body weight with 95% confidence limits (84.9–203.9). The estimated LD50 value of FIP in the current study increased than that published by other researchers, where FIP was reported as moderately toxic with LD50 equal 91.0 mg/kg b.w. in mice (Mondot and Dange 1995; Tomlin 2006) and with 97.0 mg/kg b.w. in rats (Gardner 1988; Tingle et al. 2003). The low toxicity of FIP in the present study compared to that published in the literature can be explained as the toxicity of pesticides is probably affected by the type of their formulations (Pereira et al. 2009). It is known that US EPA pesticide registration is based on toxicity data of the active ingredient (technical grade), not formulations as they are sold and applied. The acute oral toxicity of fipronil formulation, REGENT® 200SC, in aqueous vehicle was low in rats (LD50 = 1099.0 mg/kg b.w.) and moderate in mice (LD50 = 324.0 mg/kg b.w.) (Dreher 1990a,b).
Clinical symptoms of FIP observed, in the current study, were mostly distinguished within 24 h of animal exposure, and this is reliable with the known mode of action for FIP which acts as a GABA antagonist. The observed clinical symptoms involved hyperactivity, tremors and convulsions, effects on animal activity or gait, hunched posture, wetness in various body areas, and seizures (Gardner 1988; Cracknell 1991; Nachreiner 1995). The toxicity signs did not reach their ultimate until 7 h after oral administration of FIP (purity, 97.9%) to Crl:CD BR rats at different doses (2.5, 7.5, or 25 mg/kg b.w.) (Hughes 1997). The changes in motor activity involved decreased large movements were observed during the first 10 min of the 7-h evaluation (Hughes 1997). The no-observed-adverse-effects level (NOAEL) of FIP, in acute neurotoxicity study, on rats was reported 2.5 mg/kg b.w., where the lowest dose that produced adverse effects (LOAEL) was 7.5 mg/kg(Hughes 1997; Gill et al.1993). Researches also reported that decreased hind-foot splay was observed in males at 7.5 and 25 mg/kg b.w. 7 h after single dosing, where females showed decreased grooming and decreased body-weight gain, food consumption, and food efficiency. All treatment-linked effects were resolved by 14 days of exposure to FIP via acute-toxicity assay, except decreased grooming among female rats (Jennings et al. 2002a, b). In the present study, three sublethal doses were selected to use in the subacute toxicity assay including the published acute neurotoxicity NOAEL dose as medium dose, 2.87 mg/kg b.w. The NOAEL reported previously was 2.5 mg/kg b.w., based on decreased body-weight gain, food consumption, decreased hindlimb splay in males, and decreased grooming in females (Hughes 1997; Jackson et al. 2009). The LOAEL of FIP in mice was reported 2.4 mg/kg b.w. in a short-term(6-week)repeated-dose study (Holmes 1990).
Blood can act as a good biomarker for some pathological and physiological alterations of animal health (Jorum et al. 2016). The biomarker was defined as a biological response to a chemical or a group of chemical agents (Walker et al. 1996) but not the presence of the agent or its metabolites within the body (internal dose). It is also defined by Silbergeld and Davis (1994) as a physiological signal that reflect exposure, early cellular response, or inherent or acquired susceptibilities, which provide a new strategy for resolving some toxicological problems. Hence, changes in hematological parameters are considered to be one of the most important bioindicators of physiological stress (Akinrotimi et al. 2009). Hematological indices, like hemoglobin, hematocrit, and counts of erythrocytes and white blood cells, can be used as biomarkers of toxicity. Such parameters have a wide potential application in the environmental and occupational monitoring (Sancho et al. 2000; Barcellos et al. 2003). In this study, it was clear that the 28-day repeated dose exposure to FIP resulted in significant decrease in erythrocyte count, RBC, in the groups exposed to 1/30 and 1/50 LD50. The reduction in RBCs subsequently led to a significant decrease in hemoglobin (HGB) concentrations, MCH (hemoglobin amount per red blood cell), MCV (average red blood cell size), and hematocrit (HCT) values in these groups compared to these in control. Red blood cell parameters reflect the size (MCV) and level of hemoglobin concentrations (MCH and MCHC) of the red blood cells and help in the diagnosis of the reason of anemia. Also, leukocyte counts (WBCs), LYM (lymphocyte), and PLT (platelet) levels were found to be significantly decreased at 1/30 LD50 and 1/50 LD50 doses, compared to these of the control group. In the cellular immune response, lymphocytes are considered the most important cells of the immune response since they contain in their membrane receptors which are able to distinguish certain antigenic molecules and correspond to the higher percentage of the white blood cells (Ruiz et al. 2007). Our present results are in agreement with these reported by Kartheek and David (2017) who demonstrated that administration of Wistar rats to sublethal doses of fipronil (6.46, 12.12 and 32.33 mg/kg b.w.) for 90 days caused significant decreases in the levels of RBC, WBC, HGB, HCT, MCH, and MCHC as compared to the control. Similar findings were reported by Goel et al. (2006) and Ambali et al. (2010) who stated that exposure of rats to chlorpyrifos under long-term durations have been known to cause anomalies including anemia in non-target organisms (Bigoniyaa et al. 2015).
In the present study, repeated sub-lethal doses (1/30 LD50, 1/50 LD 50, and 1/100 LD50) of FIP for 28 days showed significant changes in serum biochemical parameters associated with disturbance in liver and kidney functions. The liver has a number of critical functions in the body, such as glycogen storage, plasma protein synthesis, and detoxification of xenobiotics. It also produces bile, which is important for digestion. Therefore, to evaluate hepatic damage, serum aminotransferase enzymes, AST and ALT, were monitored. The present study demonstrated that activities of serum ALT and AST, as well as cholesterol levels in male mice exposed daily for 28 days to sublethal doses of FIP at 1/30 and 1/50 LD50 showed significant increases when compared to those in the control group. The alterations that happened in activities of liver marker enzymes are often utilized as a critical indicator of cellular damage among hepatocytes with a further indication of changed plasma membrane permeability (Dhouib et al. 2015; Kaneko et al. 1997). Similar findings have been reported by several researchers wherein the increase in liver indicator enzymes has been attributed to the histopathological incidence. Our present results are in agreement with Abdel-Daima and Abdeen (2018) who reported similar conditions of elevated AST, ALT, and ALP activities in the serum of Wistar albino rats following oral administration with FIP at 20 mg/kg b.w. Similar findings were also reported by Mossa et al. (2015) who found that FIP administered to albino male rats at concentrations 0.1, 1, and 10 mg/L in drinking water for 45 days induced highly significant increases in the activities of serum AST, ALT, ALP, and lactate dehydrogenase (LDH). Likewise, Kartheek and David (2018) reported that FIP caused a significant elevation in the levels of AST, ALT, and ALP in the serum of Wistar rats. The elevation of AST activity, a cytosolic enzyme of the hepatocytes, reflects the increase of plasma membrane permeability as a result of the damage of hepatocytes (Plaa and Hewitt 1982), and this is a biomarker of liver injury (Klaassen and Eaton 1991). The type of liver damage was found to be associated with the relative rise of ALT and alkaline phosphatase (ALP)(Hussaini and Farrington 2007).
Kidney function assessment was generally made using crucial parameters, i.e., serum creatinine concentration, blood urea nitrogen level, and urine analysis (Lopez and Madero 2015). It is known that kidney needs high energy levels for active transport of several molecules via the nephrons, mostly in the cortical segments, where mitochondria are plentiful (Small et al. 2012). It is known that creatinine is a breakdown product of creatine phosphate in muscle and is regularly produced at a fairly constant level by the animal body depending on muscle mass (Yuegang et al. 2008). The diagnosis of renal failure is usually assumed when serum creatinine is greater than the upper limit of the “normal” level. Urea is the main nitrogenous end-product of protein and amino acid catabolism, produced by the liver distributed through intracellular and extracellular fluid. Urea is then filtered out of blood in kidney by glomeruli and is somewhat being reabsorbed with water (Miller et al. 2005). The most commonly determined clinical markers for estimating renal function depends upon concentration of urea in the serum. Therefore, blood urea and serum creatinine have typically been considered as good biomarkers to diagnose kidney damage (Edelstein 2008). In the present study, significant increases in the levels of serum creatinine of male mice received FIP at high (4.78 mg/kg b.w.) and medium (2.87 mg/kg b.w.) doses through 28 days repeated-dose exposure as compared to that of the control. Moreover, our study revealed that FIP administered to mice, at all sublethal doses, significantly increased urea and uric acid levels, in a dose-dependent manner. Kidney requires high energy levels for active transport of various molecules through the nephrons, particularly in the cortical segments, where mitochondria are abundant (Small et al. 2012). It was suggested that FIP-induced histological alterations in kidney tissue with changes in kidney function markers, including creatinine and urea, were mediated by oxidative damage (Abdel-Daima and Abdeen 2018). These findings are in consistent with those reported recently by Abdel-Daima and Abdeen (2018) who found that FIP administered to Wistar rats (20 mg/kg b.w.) induced an elevation of serum kidney biomarkers, urea and creatinine levels, suggesting that such alterations were mediated by oxidative damage. Similar trend was also reported by Mossa et al. (2015) who found that the levels of uric acid and creatinine were significantly increased in albino rats exposed to FIP at 1 and 10 mg/L in drinking water for 45 days. The significant increase in urea and creatinine levels observed suggests that the pesticide is possibly nephrotoxic. The elevation in serum uric acid concentration might be caused by degradation of purines and pyrimidines or because of overproduction and inability of excretion (Mossa and Abbassy 2012; El-Sebae et al. 1981), where the increase of urea level might be attributed to reduction in glomerular filtration in the kidney that reflects dysfunction of the kidney tubules (Walmsley and White 1994).
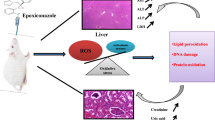
It is known that exposure to pesticides has been associated with several harmful effects including excessive production of reactive oxygen species (ROS) which led to defective organ dysfunction (Murphy 2009). Production of oxygen free radicals which may occur as a result of exposure to pesticides leads to tissue injury by triggering several oxidative mechanisms and lipid peroxidation (Kanbur et al. 2008). Since it is a complicated process to estimate free radicals directly in vivo, it is essential to act upon the quantification of cellular components which can react with these free radicals, like DNA (Seto et al. 1981), proteins (Halliwell and Chirico 1993), and mostly lipids (Esterbauer and Cheeseman 1990). In physiological conditions, oxy-radicals are part of the normal regulatory sequence of the organism and the cellular redox state is thoroughly controlled by antioxidants. When the levels of free radicals increase while both the enzymatic systems and low molecular antioxidants are insufficient to protect the organism, these radicals attack membranes and cells. This process is mostly energetic in the vascular system, where they can damage the endothelium by peroxidation and initiate the atherosclerotic lesion (Shaw et al. 2005). Estimating the level of lipid peroxidation (LPO) has a high significance in pathologies and in toxicology related to oxidative stress. One of the most recognized secondary products of lipid peroxidation is malondialdehyde (MDA) which can be used as an important indicator of cell membrane damage (Esterbauer et al. 1991). It is well known that MDA is the most important end-product of lipid peroxidation (LPO) and is often thought to proliferate and magnify oxidative injury (Pizzimenti et al. 2013). Though MDA assay is sensitive and widely used, the most common methods to MDA detection, thiobarbituric acid reactive substances (TBA-RS) reaction by spectrophotometer detection, are insufficiently sensitive and specific because TBA can react with numerous cellular constituents other than those derived from lipid peroxidation and less specifically with MDA (Grotto et al. 2009).
Estimation of the LPO in tissues is being a valuable marker to determine the extent of the oxidative stress, and its determination helps in validating the intensity of injury to biological membranes (Kartheek and David 2016). But it is necessary to consider that these biological samples may only be used in experimental studies. Hence, plasma sample is still the most used in such assays (Grotto et al. 2009). In the present study, the effect of different doses of FIP on LPO was measured in terms of MDA. It was obvious that administration of FIP at high (4.78 mg/kg b.w.) and medium (2.87 mg/kg b.w.) doses significantly increased (P ≤ 0.05) the MDA levels as compared to that of the control. Similar trend was also reported of hydrogen peroxide (H2O2), where the increased levels seemed to be in dose-dependent manner. In agreement with previous reports, fipronil provoked strong oxidative stress, increasing MDA levels more than threefold at 30 μM and sixfold at 50 μM (Lassiter et al. 2009). Insignificant increases in both MDA and H2O2 levels were observed at the lower dose, 1.44 mg/kg b.w. These findings are quite consistent with those reported by Badgujar et al. (2016) who demonstrated that FIP treatments in male mice, at different doses of 2.5, 5.0, and 10.0 mg/kg b.w./day, for 4 weeks, caused significant increase in the level of MDA in the hepatic tissue of mice, and this increase was dose-dependent in nature. Similar findings were also reported recently by Kartheek and David (2018) who found that Wistar albino rats exposed to high dose of FIP (32.33 mg/kg b.w.) indicated the highest MDA level as compared to control. A probable explanation for this may possibly be that a much stronger oxidant than superoxide anion-radical could initiate the chain oxidation of polyunsaturated phospholipids, consequently resulting to damage of membrane function (Schneider et al. 2008).
It is well known that both enzymatic- and non-enzymatic antioxidants are considered as the first line of defense against free radical-induced toxicity. A redox equilibrium between prooxidants and antioxidants is crucial for the normal cellular functioning. Therefore, the animals have antioxidant defense mechanism to counterbalance reactive oxygen species, composed of enzymes involving catalase (CAT), superoxide dismutase (SOD), glutathione peroxidase (GPx), and glutathione reductase (GR), as well as non-enzymatic antioxidants including non-protein thiols, especially glutathione (GSH). The oxidative injury may occur when the defenses of the organism are inadequate to counteract the ROS, one of the most important types of which is membrane lipid peroxidation (Scandalios 2005). In the present study, oral administration of male mice to FIP at high dose, 4.78 mg/kg b.w., induced significant decrease in both CAT and SOD, where insignificant decreases were observed at the other sublethal doses, as compared to these of the control. These results are in complete agreement with the findings of Badgujar et al. (2016) who have reported that FIP- treatments in male mice, at doses of 5.0 and 10.0 mg/kg b.w./day, for 4 weeks, caused significant decreases in the activities of antioxidant enzymes; CAT, SOD, and glutathione-S-transferase(GST) as well as the content of non-antioxidant enzymes; glutathione and total thiol. Similar decrease in the activities of both SOD and CAT enzymes was also observed in the kidney and brain tissues of mice following FIP exposure (Badgujar et al. 2015). Likewise, our results are in accordance with those reported by Swelam et al. (2017) and Mossa et al. (2015) who found that the activities of antioxidant-enzymes, SOD, CAT, GPx, and GST were significantly decreased, while LPO was significantly increased in male albino mice and albino rats administered with FIP at 1.0 and 10.0 mg/L in drinking water for 45 days, compared to the control. Recently, similar findings have been reported by Kartheek and David (2018) who reported significant variation in antioxidant enzymes, CAT and SOD in the blood serum of Wistar rats exposed to high (32.33 mg/kg b.w.) and medium (12.12 mg/kg b.w.) doses of FIP. A likely explanation for the increase of LPO by FIP may be owing to the production of free radicals as a result of metabolism of FIP that could have chemically modified essential biomolecules specifically lipids and proteins causing the condition of LPO (Young and Woodside 2001).
On the other hand, the possible ameliorative role of selenium plus vitamin E on FIP-induced toxicity was evaluated in mice pre-treated with selenium (plus Vit E) 14 days prior to exposure to the higher sublethal dose of FIP, 4.78 mg/kg b.w. This dose was the most effective dose on treated mice causing significant alterations in the biochemical parameters related to liver and kidney functions, and these associated with oxidative stress. To protect against FIP-induced oxidative stress, a variety of compounds have been evaluated for their anti-oxidative effects such as vitamin C, vitamin E, and selenium. However, little literature has been found on the protective role of selenium and vitamin E in ameliorating the adverse effects of FIP in mice or rats.
It is well known that the free radicals produced by normal metabolic processes and/or by environmental pollutants have been scavenged by antioxidant vitamins E and C and stop their attack on polyunsaturated fats (Laurence et al. 1997). Interestingly, our present data demonstrated that pre-treatment of mice via oral administration with selenium (plus vitamin E) 2 weeks prior to exposure to high dose of FIP restored the levels of hematological parameters, especially HGB, MCV, MCHC, WBC, and PLT to the control values. The levels of biomarkers associated with liver functions, especially AST, albumin, and total protein, were also obviously restored to those of the untreated control, whereas the other parameters, ALT, ALP, total bilirubin, and cholesterol, still significantly increased than those of the control. Vitamins E and C were found to protect cellular injury to hepatocytes from the free radicals caused by FIP treatment and thus improved histological lesions. Similar findings were also reported by several researchers (Sulak et al. 2005; Sankar et al. 2013; Jebur et al. 2014). In addition, our results showed that pre-treatment of mice with selenium (plus vitamin E) before exposure to high dose of FIP exhibited ameliorative effect by restoring levels of biomarkers associated with kidney injury, i.e., creatinine and urea, nearly to those of the control group
The present study showed also that pre-treatment of mice with selenium plus vitamin E resulted in rather positive effect on amelioration of oxidative stress by reducing lipid peroxidation and hydrogen peroxide levels induced as a result of administration of mice to high dose of FIP. In agreement with our results, Slotkin and Seidler (2010) revealed that addition of vitamin E reduced the MDA concentration to normal levels. Likewise, Badgujar et al. (2016) demonstrated that FIP evoked lipid peroxidation both in vitro and in vivo and vitamin E and vitamin C have protective effect on FIP-induced lipid peroxidation. The role of vitamins E and C in counteracting oxidative stress is well known. Mostly, the ameliorative or protective role of vitamin E is attributed to its ring structure possessing hydrogen giving hydroxyl group; thus, it acts as chain terminating antioxidant (Badgujar et al. 2016). Vitamin C immediately and rapidly scavenges free radicals inhibiting their formation (Badgujar et al. 2016). Vitamin C protects the DNA of cells from harm caused by free radicals and may involve in combating the adverse effects of various toxins, including pesticides and heavy metals (El-Gendy et al. 2010). In the present study, the ameliorative effects of selenium plus vitamin E on FIP-induced toxicity are in agreement with these findings demonstrated by Badgujar et al. (2016) who reported that prior treatment with vitamins E or C against high dose of FIP led to an increase in the activities of SOD, catalase, and GST as well as non-enzymatic antioxidants content of GSH and total thiol in the liver tissue of mice. The protective effect of vitamins seen in this study is also in consistent with researchers who revealed the protective effects of vitamins E and/or C against pesticides such as imidacloprid, methidathion, and chlorpyrifos (Sulak et al. 2005; Verma et al. 2007; El-Gendy et al. 2010, respectively).
Conclusions
It is concluded that short-term repeated exposure to commercial formulation of fipronil (COACH®, 200 SC), even at low doses, can induce serious hepatic and renal dysfunction as well as hematological damage, caused presumably by increased oxidative stress. Based on our present results, it is suggested that the dose of FIP lower than 1.87 mg/kg b.w. administered orally to male albino mice for 28 days could be considered as no-observed-adverse-effect level (NOAEL). Further, more sub-chronic and chronic studies are required to establish the NOAEL at long-term repeated exposure. Moreover, findings of this study suggest that prior administration of antioxidant vitamin E plus selenium combated adverse effects including oxidative stress induced by fipronil. The extensive global use of different FIP formulations underlines the importance of such findings.
References
Abdel-Daima MM, Abdeen A (2018) Protective effects of rosuvastatin and vitamin E against fipronil-mediated oxidative damage and apoptosis in rat liver and kidney. Food Chem Toxicol 114:69–77
Aebi H (1984) Catalase in vitro. Methods Enzymol 105:121–126
Ahmad I, Hamid T, Fatima M, Chand HS, Jain SK, Athar M, Raisudin S (2000) Induction of hepatic antioxidants in freshwater catfish (Channa punctatus Bloch) is a biomarker of paper mill effluent exposure. Biochim Biophys Acta 1523:37–48
Akinrotimi OA, Abu OMG, Ansa EJ, Edun OM, George OS (2009) Haematological responses of Tilapia guineensis to acute stress. J Nat Appl Sci 5:338–343
Ambali SF, Abubakar AT, Shittu M, Yaqub LS, Anafi SB, Abdullahi A (2010) Chlorpyrifos induced alterations of hematological parameters in Wistar rats: Ameliorative effect of zinc. Res J Environ Toxicol 4:55–66
Ateşşahin A, Yilmaz S, Karahan I, Pirinçci I, Taşdemir B (2012) The effects of vitamin E and selenium on cypermethrin-induced oxidative stress in rats. Turk J Vet Anim Sci 29:385–391
Badgujar PC, Pawar NN, Chandratre GA, Telang AG, Sharma AK (2015) Fipronil induced oxidative stress in kidney and brain of mice: protective effect of vitamin E and vitamin C. Pestic Biochem Physiol 118:10–18
Badgujar PC, Chandratre GA, Pawar NN, Telang AG, Kurade NP (2016) Fipronil induced oxidative stress involves alterations in SOD1 and catalase gene expression in male mice liver: protection by vitamins E and C. Environ Toxicol 31:1147–1158
Badgujar PC, Selkar NA, Chandratre GA, Pawar NN, Dighe VD, Bhagat ST, Telang AG, Vanage GR (2017)Fipronil-induced genotoxicity and DNA damage in vivo: Protective effect of vitamin E. Hum Exp Toxicol 36(5):508–519
Banerjee BD, Seth V, Ahmed RS (2001) Pesticides induced oxidative stress: perspectives and trends. Rev Environ Health 16:1–40. https://doi.org/10.1515/reveh.2001.16.1.1
Barcellos LJG, Kreutz LC, Rodrigues LB, Fioreze I, Quevedo RM, Cericato L, Conrad J, Soso AB, Fagundes M, Lacerda LA, Terra S (2003) Haematological and biochemical characteristics of male jundiá (Rhamdia quelen Quoy & Gaima RDT, Pimelodidae): changes after acute stress. Aquac Res 34:1465–1469
Bigoniyaa P, Sahua T, Tiwari V (2015) Hematological and biochemical effects of sub-chronic artesunate exposure in rats. Toxicol Rep 2:280–288
Bradford MM, Marion M (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72(1-2):248–254
Bruce RD (1985) An up-and-down procedure for acute toxicity testing. Fundam Appl Toxicol 6:151–157
Combs GF, Combs SB (1986) The Role of Selenium in Nutrition. Florida: Academic Press.
Cracknell S (1991) M&B 46030: Acute inhalation toxicity study in the rat. Unpublished report No. 90/RHA358/0791 from Life Science Research Ltd. Submitted to WHO by Rhone-Poulenc, Inc., Research Triangle Park, NC, USA
De Oliveira PR, Bechara GH, Denardi SE, Oliveira RJ, Mathias MI (2012) Cytotoxicity of fipronil on mice liver cells. Microsc Res Tech 75:28–35
Dhouib IEB, Annabi A, Montassar ML, Gharbi N, El-Fazaa S (2015)Anti-inflammatory effects of N-acetylcysteine against carbosulfan-induced hepatic impairment in male rats, Recent. Adv Biol Med 1:29–40
Djeffal A, Messarah M, Boumendjel A, Kadeche L, El Feki A (2012) Protective effects of vitamin C and selenium supplementation on methomyl-induced tissue oxidative stress in adult rats. Toxicol Ind Health 31:31–43
Dreher DM (1990a) Formulation EXP60145A: Acute oral toxicity test in the rat. Project Number 282/1. Report number: SLL 3797-282/2. Lab: Safepharm Laboratories Ltd, Derby, UK. Unpublished. 18.1.1990 [RP; sub:10528, Vol. 8]
Dreher DM (1990b) Formulation EXP60145A: Acute oral toxicity test in the mouse. Project Number 282/2. Report number: SLL 3796-282/1. Lab: Safepharm Laboratories Ltd, Derby, UK. Unpublished. 18.1. [RP; sub:10528, Vol. 8]
Edelstein CL (2008) Biomarkers of Acute Kidney Injury. Adv Chronic Kidney Dis 15:222–234
El-Desoky G, Abdelreheem M, AL-Othman A, AL-Othman Z, Mahmoud M, Yusuf K (2012) Potential hepatoprotective effects of vitamin E and selenium on hepatotoxicity induced by malathion in rats. Afr J Pharm Pharmacol 6:806–813
El-Gendy AK, Aly NM, Mahmoud FH, Kenawy A, El-Sebae AK (2010) The role of Vit C as antioxidant in protection of oxidative stress induced by Imidacloprid. Food Chem Toxicol 48:215–221
El-Sebae AH, Enan EE, Soliman SA, El-Fiki S, Khamees E (1981) Biochemical effects of some organophosphorus insecticide on new targets in white rats. J Environ Sci Health Part B 16(4):475–491
El-Shenawy NS, El-Salmy F, Al-Eisa RA, El-Ahmary B (2010) Amelioratory effect of vitamin E on organophosphorus insecticide diazinoninduced oxidative stress in mice liver. Pestic Biochem Physiol 96:101–107
Esterbauer H, Cheeseman KH (1990) Determination of aldehydic lipid peroxidation products: Malonaldehyde and 4 Hydroxynonenal. Methods Enzymol 186:407–421
Esterbauer H, Schaur RJ, Zollner H (1991) Chemistry and Biochemistry of 4-Hydroxynonenal, Malonaldehyde, and Related Aldehydes. Free Radic Biol Med 11:81–128
Finney DJ (1971) Probit Analysis, 3rd edn. Cambridge University Press, Cambridge 333 p
Flora S, Kannan G, Pant B, Jaiswal D, (2002) Combined administration of oxalic acid, succimer and its analogue for the reversal of gallium arsenide-induced oxidative stress in rats. Arch Toxicol 76:269–276
Fossati P, Prencipe L, Berti G (1980) Use of 3,5-dichloro-2-hydroxy-benzenesulfonic acid/4-aminophenazone chromogenic system in direct enzymic assay of uric acid in serum and urine. Clin Chem 26:227–231
Gardner JR (1988) Acute oral toxicity to rats of M&B 46,030. Unpublished report No. 881300D/M&B 290/AC from Huntingdon Research Center Ltd. Submitted to WHO by Rhone-Poulenc, Inc., Research Triangle Park, NC, USA. Gastroenterol Res Pract 2015:738548.
Giannini EG, Testa R, Savarino V (2005) Liver enzyme alteration: a guide for clinicians. CMAJ: Can Med Assoc J 172:367–379
Gill MW, Wagner CL, Driscoll CD (1993) M&B 46030: Single exposure per oral (gavage) neurotoxicity study in Sprague Dawley rats. Unpublished report No. 91N0099 from Union Carbide Bushy Run Research Center. Submitted to WHO by Rhone-Poulenc, Inc., Research Triangle Park, NC, USA
Giray B, Gürbay A, Hincal F (2001)Cypermethrin-induced oxidative stress in rat brain and liver is prevented by Vitamin E or allopurinol. Toxicology Letters 118(3):139–146
Goel A, Danni V, Dhawan DK (2006) Role of zinc in mitigating the toxic effects of chlorpyrifos on hematological alterations and electron microscopic observations in rat blood. Bio Metals 19:483–492
Grotto D, Maria LS, Valentini J, Paniz C, Schmitt G, Garcia SC, Pomblum VJ, Rocha JB, Farina M (2009) Importance of the lipid peroxidation biomarkers and methodological aspects for malondialdehyde quantification. Quim Nova 32(1):169–174
Hainzl D, Cole LM, Casida JE (1998) Mechanisms for selective toxicity of fipronil insecticide and its sulfone metabolite and desulfinyl photoproduct. Chem Res Toxicol 11:1529–1535
Halliwell B, Chirico S (1993) Lipid Peroxidation Its Mechanism, Measurement, and Significance. Am J Clin Nutr 57:715–725
Henry RJ (1974) Clinical Chemistry, Principles and Techniques, 2nd edn. Harper and Row, Hagerstown 525 p
Hoffman DJ (2002) Role of selenium toxicity and oxidative stress in aquatic birds. Aquat Toxicol 57:11–26
Holmes P (1990) M&B 46030: Preliminary toxicity study by dietary administration to CD-1 mice for six weeks. Unpublished report no. 90/RHA299/0325 from Life Science Research, Ltd. Previously submitted to WHO by Rhone-Poulenc, Inc., RTP, North Carolina, USA
Hughes EW (1997) Fipronil: Neurotoxicity to rats by acute oral administration (including a time to peak effect study) Unpublished report No. RNP 536/973345 from Huntingdon Life Sciences. Submitted to WHO by Rhone-Poulenc, Inc., RTP, North Carolina, USA
Hussaini SH, Farrington EA (2007) Idiosyncratic drug-induced liver injury: an overview. Expert Opin Drug Saf 6:673–684
Jackson D, Cornell CB, Luukinen B, Buhl K, Stone D (2009) Fipronil Technical Fact Sheet; National Pesticide Information Center, Oregon State University Extension Services. (2009, January) Retrieved from http://npic.orst.edu/factsheets/fiptech.html.
Jebur AB, Nasr HM, El-Demerdash FM (2014) Selenium modulates b-cyfluthrin-induced liver oxidative toxicity in rats. Environ Toxicol 29:1323–1329
Jennings KA, Keller RJ, Doss RB (2002a) Human Exposure to Fipronil from Dogs Treated with Frontline. Controv Toxicol 44(5):301–303
Jennings KA, Canerdy TD, Keller RJ et al (2002b) Human exposure to fipronil from dogs treated with frontline. Vet Hum Toxicol 44(5):301–303
Jorum OH, Piero NM, Machocho AK (2016) Haematological effects of dichloromethane-methanolic leaf extracts of Carissa edulis(Forssk) Vahl in Normal Rat Models. J Hematol Thromboembolic Dis 5:2
Kanbur M, Atalay O, Ica A, Eraslan G, Cam Y (2008) The curative and antioxidative efficiency of doramectin and doramectin+vitamin AD3E treatment on Psoroptes cuniculi infestation in rabbits. Res Vet Sci 85:291–293
Kaneko JJ, Harvey JW, Bruss ML (1997) "Clinical Biochemistry of Domestic Animals".5th Ed., Academic Press, San Diego, California of Alcohol Abuse. QJM89: 137-144
Kartheek RM, David M (2016) Fipronil induced modulations in biochemical and histopathological aspects of maleWistar albino rats: a subchronic study. J Environ Biosci 5:26–32
Kartheek RM, David M (2017) Modulations in haematological aspects of Wistar rats exposed to sublethal doses of fipronil under subchronic duration. J Pharm Chem Biol Sci 5(3):187–194
Kartheek RM, David M (2018) Assessment of fipronil toxicity on wistar rats: A hepatotoxic perspective. Toxicol Rep 5:448–456
Khan S, Jan MH, Kumar D, Telang AG (2015) Firpronil induced spermotoxicity is associated with oxidative stress, DNA damage and apoptosis in male rats. Pestic Biochem Physiol 124:8–14
Ki YW, Lee JE, Park JH, Shin IC, Koh HC (2012) Reactive oxygen species and mitogen-activated protein kinase induce apoptotic death of SH-SY5Y cells in response to fipronil. Toxicol Lett 211(1):18–28
Kidd H, James D (eds) (1991) The Agrochemicals Handbook, 3rd edn. Royal Society of Chemistry Information Services, Cambridge
Klaassen CD, Eaton DL (1991) Principles of toxicology. In: Amdur MO, Doull J, Klaassen CD (eds) Toxicology, 4th edn. McGraw Hill, New York, p 12
Kori-Siakpere O, Oboh EC (2011) Haematological effects of sublethal concentrations of tobacco leaf dust on the African catfish: Clarias gariepinus (Burchell 1822). Arch Appl Sci Res 3:93–502
Lassiter TL, MacKillop EA, Ryde IT, Seidler FJ, Slotkin TA (2009) Is fipronil safer than chlorpyrifos? Comparative developmental neurotoxicity modeled in PC12 cells. Brain Res Bull 78:313–322
Laurence DR, Bennett PN, Brown MJ (1997) Clinical pharmacology, 8th edn. Churchill Living Stone, New York, pp 669–675
Leghait J, Gayrard V, Toutain P-L, Picard-Hagen N, Viguié C (2010) Is the mechanism of fipronil-induced thyroid disruption specific to the rat: re-evaluation of fipronil thyroid toxicity in sheep? Toxicol Lett 194:351–357
Li W, Yin D, Zhou Y, Hu S, Wang L (2003) 3,4-Dichloroaniline-induced oxidative stress in liver of crucian carp (Carassius auratus). Ecotoxicol Environ Saf 56:251–255
Lopez GS, Madero M (2015) Biomarkers in chronic kidney disease, from kidney function to kidney damage. World J Nephrol 4(1):57–73
Malloy HT, Evelyn KA (1937) The determination of bilirubin with the photoelectric colorimeter. J Biol Chem 119:481
Marklund S, Marklund G (1974) Involvement of the superoxide anion radical in the autoxidation of pyrogallol and a convenient assay for superoxide dismutase. Eur J Biochem 47:469–474
Miller W, Myers G, Ashwood E et al (2005) Creatinine measurement: state of the art in accuracy and interlaboratory harmonization. Arch Pathol Lab Med 129(3):297–304
Mondot S, Dange M (1995) Unpublished report No. R&D/CRSA/TO-PHA3 from Rhone-Poulenc Agrochimie Toxicology (Research Triangle Park, NC, USA, Submitted to WHO by Rhone-Poulenc, Inc)
Mossa AH, Abbassy MA (2012) Adverse haematological and biochemical effects of certain formulated insecticides in male rats. Environ Toxicol 6(4):160–168
Mossa AH, Refaie AA, Ramadan A (2011) Effect of exposure to mixture of four organophosphate insecticides at no observed adverse effect level (NOAEL) dose on rat liver: the protective role of vitamin C. Res J Environ Toxicol 5:323–335
Mossa A, Swelam ES, Mohafrash SMM (2015)Sub-chronic exposure to fipronil induced oxidative stress, biochemical and histopathological changes in the liver and kidney of male albino rats. Toxicol Rep 2:775–784
Murphy PM (2009) How mitochondria produce reactive oxygen species. J Biochem 417(1):1–13
Nachreiner DJ (1995) Fipronil: Acute nose-only dust inhalation study in rats. Unpublished report No. 94N1501 from Union Carbide Bushy Run Research Center. Submitted to WHO by Rhone-Poulenc, Inc., Research Triangle Park, NC, USA
Nourooz-Zadeh J, Tajaddini-Sarmadi J, Wolff SP (1994) Measurement of plasma hydroperoxide concentrations by the ferrous oxidation-xylenol orange assay in conjunction with triphenylphosphine. Anal Biochem 220:403–409
OECD (Organization for Economic Cooperation and Development) (1998) Repeated dose 90-day oral toxicity study in rodents. Guideline 408 (Updated September 21)
OECD (Organization for Economic Cooperation and Development) (2008) Guidelines for the Testing of Chemicals. No 39 Draft Guidance Document on Acute Inhalation Toxicity Testing Version 9, March 2008
Ohi M, Dalsenter PR, Andrade AJ, Nascimento AJ (2004) Reproductive adverse effects of fipronil in Wistar rats. Toxicol Lett 146:121–127
Patton CJ, Crouch SR (1977) Spectrophotometric and kinetics investigation of the Berthelot reaction for the determination of ammonia. Anal Chem 49:464–469
Pereira JL, Antunes SC, Castro BB, Marques CR, Goncalves AM, Goncalves F et al (2009) Toxicity evaluation of three pesticides on non-target aquatic and soil organisms: commercial formulation versus active ingredient. Ecotoxicology 18:455–463
Pimpao CT, Zampronio AR, De Assis HC (2007) Effects of deltamethrin on hematological parameters and enzymatic activity in Ancistrus multispinis (Pisces, Teleostei). Pestic Biochem Physiol 88:122–127
Pizzimenti S, Ciamporcero E, Daga M, Pettazzoni P, Arcaro A, Cetrangolo G, Minelli R, Dianzani C, Lepore A, Gentile F, Barrer G (2013) Interaction of aldehydes derived from lipid peroxidation and membrane proteins. Front Physiol l4:242
Plaa GL, Hewitt WR (1982) Detection and evaluation of chemically induced liver injury. In: Hayes W (ed) Principles and Methods of Toxicology. Raven Press, New York, p 407
Prashanth MS, David M (2006) Changes in nitrogen metabolism of the freshwater fish Cirrhinus mrigala following exposure to cypermethrin. J Basic Clin Physiol Pharmacol 17:63–70
Raneva V, Shimasaki H, Furukawa Y, Ueta N, Yanishlieva N, Aaseng JE, Partali V, Sliwka HR, Yoshida Y, Niki E, (2002) Action of 1-(11-selenadodecyl)-glycerol and 1-(11-selenadodecyl)-3-trolox-glycerol against lipid peroxidation. Lipids 37:633–640
Rayman MP (2008)Food-chain selenium and human health: emphasis on intake. Br J Nutr 100:254–268
Reitman S, Frankel S (1957) A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am J Clin Pathol 28(1):56–63
Roschlau P, Bernt E, Gruber W (1974) Enzymatic determination of total cholesterol in serum. Z Klin Chem Klin Biochem (J Clin Chem Clin Biochem)12:403–408.
Ruiz FS, Andersen ML, Zager A, Martins RCS, Tufik S (2007) Sleep deprivation reduces the lymphocyte count in a non-obese mouse model of type 1 diabetes mellitus. Braz J Med Biol Res 40:633–637
Sancho E, Cerón JJ, Ferrando MD (2000) Cholinesterase activity and hematological parameters as biomarkers of sublethalmolinate exposure in Anguilla anguilla. Ecotoxicol Environ Saf 46:81–86
Sankar P, Telang AG, Kalaivanan R, Karunakaran V, Manikam K, Sarkar SN (2013) Effects of nanoparticle—encapsulatedcur-cumin on arsenic—induced liver toxicity in rats. Environ Toxicol 6:628–637
Scandalios JG (2005) Oxidative stress: molecular perception and transduction of signals triggering antioxidant gene defenses. Braz J Med Biol Res 38:995–1014
Schneider BP, Wang M, Radovich M, Sledge GW, Badve S, Thor A, Flockhart DA, Hancock B, Davidson N, Gralow J, Dickler EA, Cobleigh M, Shenkier T, Edgerton S, Miller KD (2008) ECOG 2100. association of vascular endothelial growth factor and vascular endothelial growth factor receptor-2 genetic polymorphisms with outcome in a trial of paclitaxel compared with paclitaxel plus bevacizumab in advanced Breast cancer: ECOG 2100. J Clin Oncol 26:4672–4678
Schomaker S, Warner R, Bock J, Johnson K, Potter D, Van Winkle J, Aubrecht J (2013) Assessment of emerging biomarkers of liver injury in human subjects. Toxicol Sci 132:276–283
Seto H, Akiyama K, Okuda T, Hashimoto T, Takesue T, Ikemura T (1981) Structure of a new modified nucleoside formed by guanosine-malondialdehyde reaction. Chem Lett 52:707
Shaw CA, Taylor EL, Megson IL, Rossi AG (2005) Nitric oxide and the resolution of inflammation: implications for atherosclerosis. Mem Inst Oswaldo Cruz 100:67–71
Silbergeld EK, Davis DL (1994) Role of biomarkers in identifying and understanding environmentally induced disease. Clin Chem 40(7):1363–1367
Singh M, Kaur P, Sandhir R, Kiran R (2008) Protective effects of vitamin E against atrazine-induced genotoxicity in rats. Mutation Research/Genetic Toxicology and Environmental Mutagenesis 654(2):145–149
Slotkin TA, Seidler FJ (2010) Oxidative stress from diverse developmental neurotoxicants: antioxidants protect against lipid peroxidation without preventing cell loss. Neurotoxicol Teratol 32:124–131
Small DM, Coombes JS, Bennett N, Johnson DW, Gobe GC (2012) Oxidative stress, anti-oxidant therapies and chronic kidney disease. Nephrology 17:311–321
Stewart MS, Spallholz JE, Neldner KH, Barbara, Pence BC, (1999) Selenium compounds have disparate abilities to impose oxidative stress and induce apoptosis. Free Radic Biol Med 26:42–48
Sulak O, Altuntas I, Karahan N, Yildirim B, Akturk O, Yilmaz HR, Delibas N (2005) Nephrotoxicity in rats induced by organophosphate insecticide methidathion and ameliorating effects of vitamins E and C. Pestic Biochem Physiol 83:21–28
Swelam ES, Abdallah IS, Mossa AH (2017) Ameliorating Effect of Zinc Against Oxidative Stress and Lipid Peroxidation Induced by Fipronil in Male Rats. J Pharmacol Toxicol 12(1):24–32
Thrall MA (2004) Haematology of amphibians. Veterinary haematology and clinical chemistry: Text and clinical case presentations. Lippincott Williams and Wilkins, Philadelphia
Tingle CC, Rother CF, Lauer DS, King WJ (2003) Fipronil: environmental fate, ecotoxicology, and human health concerns. Rev Environ Contam Toxicol 176:1–66
Tingle CC, Rother JA, Dewhurst CF, et al. (2016) Health and environmental effects of fipronil, http://www.pan-uk.Org/Publications/Briefing/fipronil.pdf. Accessed 16 Jan 2016.
Tomlin DS (2006) The Pesticide Manual, A World Compendium, 14th edn. British Crop Protection Council, Surry, pp 598–599
Verma RS, Mehta A, Srivastava N (2007) In vivo chlorpyrifos induced oxidative stress: Attenuation by antioxidant vitamins. Pestic Biochem Physiol 88:191–196
Walker CH, Hopkin SP, Sibly RM, Peakall DB (1996) Biomarkers. In: Walker CH, Hopkin SP, Sibly RM, Peakall DB (eds) Principles of Ecotoxicology. Taylor and Francis, London, pp 175–194
Walmsley RN, White GH (1994) A Guide to Diagnostic Clinical Chemistry, 3rd edn. Blackwell Scientific Publication, London
Wenger C, et al (1984) Alkaline phosphatase. In: Kaplan A et al (eds) Clin Chem. The C.V. Mosby Co. Si Louis. Tronto. Princeton, pp. 1094–1098
Yagi K (1998) Simple assay for the level of total lipid peroxides in serum or plasma. Methods Mol Biol 108:101–106
Young S, Woodside JV (2001) Antioxidants in health and disease. J ClinPathol 54:176–186
Yuegang Z, Chengjun W et al (2008) Simultaneous Determination of Creatinine and Uric Acid in Human Urine by High Performance Liquid Chromatography. Anal Sci 24:1589–1592
Zia S, Islam F (2000) Selenium Altered the Levels of Lipids, Lipid Peroxidation, and Sulfhydryl Groups in Straitum and Thalamus of Rat. Biol Trace Elem Res 77(3):251–260
Acknowledgments
The authors are thankful to the Dean of Faculty of Agriculture, and the Chair of Department of Pesticides, Menoufia University, for providing necessary facilities and funds.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Philippe Garrigues
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Abouelghar, G.E., El-Bermawy, Z.A. & Salman, H.M.S. Oxidative stress, hematological and biochemical alterations induced by sub-acute exposure to fipronil (COACH®) in albino mice and ameliorative effect of selenium plus vitamin E. Environ Sci Pollut Res 27, 7886–7900 (2020). https://doi.org/10.1007/s11356-019-06579-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-019-06579-9