
Abstract
Margaritifera margaritifera is a critically endangered species in Europe. Among the causes explaining its decline, metal pollution had never been deeply studied. Thus, an ecotoxicological investigation was developed on this species which comes from the Dronne River (South-West of France). Cadmium (Cd) exposure of mussels at 2 and 5 μg/L for 7 days was conducted to test their vulnerability to this metal, and also the potential endocrine disruption power of Cd. Morphometric analyses, gonad histological observations, metal bioaccumulation, metallothionein (MTs) production, measures of malondialdehyde (MDA), and finally quantitative relative expression analysis of genes involved in various metabolic functions were performed.
The main results showed Cd accumulation increasing in a dose-dependent manner, especially in the gills. The same trend was observed for gene expression relative to oxidative stress. Histological analysis of the gonads highlighted a predominance of hermaphrodite individuals, but after 7 days of exposure to Cd, the percentage of female was largely increased compared with controls, from 17 to 33%. These results demonstrate the endocrine disruption effect of Cd on freshwater pearl mussels.
The pearl mussel Margaritifera margaritifera is sensitive to cadmium since the metallothioneins are poorly induced, gene expression reveals oxidative stress, and gonads tend to be feminized.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Indigenous freshwater mollusks are threatened by extinction in most parts of the world (Vaughn and Taylor 1999; Strayer et al. 2004), and because little is known about their physiology and behavior, conservation efforts are difficult (Rotchell and Ostrander 2003; Geist 2010). Among them, the freshwater pearl mussel Margaritifera margaritifera was in the past the most common mussel type in Europe (Bauer 1988). But populations have declined by more than 90% and only a small number of recruiting (functional) populations remain (Cosgrove and Hastie 2001; Ziuganov 2005; Hastie et al. 2008).
Several factors can be advanced to explain their disappearance during the nineteenth and twentieth centuries: pearl fishing; general deterioration of water quality; reduced host fish density; reduction of the habitats (Gumpinger et al. 2015); filling of sediments (Hauer 2015); pollution by pesticides, phosphates, and nitrogen responsible for eutrophication (Scheder et al. 2015); siltation; and impoundments (Addy et al. 2012). Indeed, this species has particular ecological requirements (see appendix 1 in Varandas et al. 2013). The quality of the water must be excellent and the substrate healthy in particular for the survival of the juvenile mussels which are sensitive to the changes in their surrounding environment (Geist and Kuehn 2005). For adults, the top of the valves (the oldest part of the mussel pearl-bearing) is the most damaged part and can prove to be fragile with the eventual chemical or physical attacks of water (Bensettiti and Gaudillat 2004; Motte 2005). Moreover, this mussel has a very particular life cycle since it has a very long lifespan (150 years on average in Europe) and requires the presence of a fish host. This life cycle is divided into four stages of development: glochidia (larval stage), parasitic stage, juvenile, and adult stage. The sexes are normally separated, but females can become hermaphrodite when isolated and under stress (Bauer 1987). First of all, the male releases sperm (June/July), which are trained by the current and then recovered by the female through its filtration system, after producing eggs. Fertilization produces a cell that evolves into a larva called glochidia. These larvae are incubated for 4 to 6 weeks in a part of the mussel mantle called marsupium. Then, the glochidia are released into the current and, to continue their development, must be fixed in the week on the branchial system of a salmonid with a preference for brown trout (Salmo trutta f. fario) in central European populations (Wächtler et al. 2001). This favorable parasitic part of the cycle constitutes both a phase of larval development and a phase of dissemination of the species with the displacement of the host fish (Treasurer et al. 2006). Once fixed, glochidia parasitize their host for a period of several months. When they leave the gills of the host, the young mussels measure about half a millimeter and have become true bivalves. They then bury themselves in the sediment for a period of 4 to 5 years in order to continue their growth and they progressively rise to position at the sediment-water interface. The fish host can also be affected by water quality and a lack of ecological continuity, which participate in the disappearance of the freshwater pearl mussel. Among pollutants introduced in aquatic systems by anthropogenic activities, metals could weaken populations of M margaritifera and jeopardize its survival.
Currently, this species is only little represented in the rivers of France. Approximately 100,000 individuals are listed. The Department of the Dordogne (South-West of France) shelters two of these populations, on the Bandiat and the Dronne rivers. The population of the Dronne is estimated at 15,000 individuals. Until now, the catchment area of the Dronne was relatively free of pollution. However, some observations made by agents of the AFB (Agence Française de la Biodiversité) or PNRPL (Parc Naturel Régional Périgord-Limousin) question a possible deterioration of the river quality.
In this catchment area, there are several potential sources of impacts (agriculture, release of suspended matter, and artificial ponds which generate eutrophication). Among them, a source of specific pollution was listed in the commune of Saint Saud Lacoussière: an illegal waste dump which had received all kinds of waste for many years, including, for instance, batteries and drugs. This discharge is located on a slope on which streams the resurgence and rainwater. The site was destabilized by work and trace metals or chemicals can be brought out by scrubbing in the Dronne River downstream. However, at this place, the river shelters a population of approximately 1000 to 1500 individuals of freshwater pearl mussels. An exceptional authorization (Agreement N°60/2008) delivered by the French ministry of the environment allowed us to collect several individuals in order to conduct an experiment and to evaluate if this discharge may have an impact on mussels. Recently, a study using some of these collected mussels demonstrated the presence of metals (As, Cd, Co, Cr, Ni, and Zn) accumulated in tissues of M. margaritifera located downstream from this dump and impacting their gene expressions (Bertucci et al. 2017).
In fact, filter-feeders such as bivalves are known to accumulate metals and other pollutants in their tissues (Couillard et al. 1993, 1995; Baudrimont et al. 1999; Giguère et al. 2003; Marie et al. 2006a, 2006b; Paul-Pont et al. 2010; Arini et al. 2014). After accumulation, metals can generate toxic effects, like, in particular, the production of oxidative stress which can poison the animal until death (Cossu et al. 2000; Gonzalez et al. 2006; Farcy et al. 2011; Gillis 2012; Gillis et al. 2014). Among these metals, cadmium (Cd) is considered as a priority substance in Europe (European Water Framework Directive 2008/105/EC) as it is a known toxic metal that accumulates and exerts its toxic action primarily in the kidneys (nephrotoxicity) and liver (hepatotoxic) in aquatic organisms, in conditions of chronic exposure. Cd is well known to provoke cellular or physiological dysfunctioning in varying degrees according to the species (Viarengo 1989; Perceval et al. 2004). It is responsible for a disturbance of the calcium metabolism in the freshwater pearl mussel (Frank and Gerstmann 2007), because of its ability to compete with Ca2+ for membrane carriers of this trace element, due to an atomic radius very close to the latter (0.97 and 0.99 Å). Cd is one of the main factors correlated with gene transcription levels, with effects on translation, apoptosis, immune response, response to stimulus, and transport pathways in M. margaritifera (Bertucci et al. 2017). Cd has also been described as an endocrine disruptor in fish (Pierron et al. 2008; Pierron et al. 2009), and as described in Margaritifera margaritifera, a gene of estrogen receptor was present (Bertucci et al. 2017) which can ask the question of this mode of action in the pearl mussel too. Among the possible lines of defense, the synthesis of small proteins such as the metallothioneins (MTs) can be induced under the influence of metals (Baudrimont et al. 2003; Amiard et al. 2006). These cytosolic proteins are able to sequester the metal ions and thus to neutralize their potential toxic impacts on other cellular targets. They are in the same way able to act directly against the generation of oxidative stress by protecting the cell from the presence of free radicals (Yang et al. 2009). This defense mechanism can be measured at the protein as well as at the gene level in tissues of aquatic organisms (Marie et al. 2006a, 2006b; Paul-Pont et al. 2010). Among the effects of oxidative stress, the oxidative degradation of fatty acids (lipid peroxidation) at the level of the cellular membranes leads to the formation of an end product, malondialdehyde (MDA), which accumulates in the cell, and that we can easily quantify in tissues, as a lipidic marker of peroxidation (Legeay et al. 2005). Metals exert other toxic effects, like the disturbance of the mitochondrial metabolism, the apoptosis (programmed cellular death), or the attacks of the DNA. All these effects can be followed by quantitative real-time PCR of specific gene mRNA implied in all these functions (Gonzalez et al. 2006; Paul-Pont et al. 2010).
The objectives of this work were thus to study the potential impacts of cadmium on several individuals of the freshwater pearl mussel Margaritifera margaritifera collected around the dumping site during the reproductive period. Owing to the fact that Cd can be a potential endocrine disruptor, in parallel, we exposed freshwater pearl mussels to estradiol as a positive control to compare its effect with Cd. The synthesis of detoxification proteins such as MTs was quantified, in parallel with MDA concentrations and relative gene expressions by quantitative PCR. Some histological preparations were also carried out in order to observe the gonads of the freshwater pearl mussels during Cd exposure in controlled conditions.
Materials and methods
Sampling of the freshwater pearl mussels in the Dronne River
Adult Margaritifera margaritifera sampling was carried out during July 2009, upstream and downstream from the illegal waste dump of Saint Saud Lacoussière (44° 33.019; 44° 51.008), following a request for authorization of sampling which was given by the DIREN (French Ministry of the Environment, Agreement N°39/2016) in 2008.
Experimental study of Cd exposure during the reproductive period
Thirty individuals of M. margaritifera were used to conduct the exposure of mussels to Cd under controlled conditions. Pearl mussels were 89 ± 7 mm long (ranging from 70 to 99 mm) and their mean age was estimated at 39 years old according to Bertucci et al. 2017. Five experimental conditions were performed, with six individuals per experimental unit chosen in order to have the same average size in each of them: “upstream control, UC,” corresponding to individuals collected upstream from the wild waste dump; “downstream control, DC,” corresponding to individuals collected downstream from the wild waste dump; Cd exposure in a direct way with 2 or 5 μg/L with individuals coming from upstream of the wild waste dump; and estradiol exposure with 100 μg/L with individuals coming from upstream of the wild waste dump, in order to get a positive control of endocrine disruption, and notably of reproductive hormones (Flynn and Spellman 2009). The contamination exposure lasted 7 days, after an acclimatization period of 14 days. Note that just before the beginning of the experiment, one individual died in the DC experimental unit, corresponding to only 3% of mortality for the entire experiment.
An experimental glass unit (volume = 40 L) consisted of 7 cm of ultrapure sand SILAQ (0.7–1.4 mm) which was rinsed beforehand and deposited on the bottom of the unit previously protected with a food standard plastic bag (Plastiluz). Each unit was filled using the water of the Dronne River taken upstream from the dump and was placed at 19 °C, near to the temperature measured in the river at this period, using a system of thermic regulation. The photoperiod was controlled with 16 h/8 h light/dark and oxygenation was ensured thanks to a system of ventilation by air pumps. Water was permanently filtered in order to avoid the intoxication of the freshwater pearl mussels by their own waste and the mussels were fed for the period of acclimatization using phytoplanctonic green algae Scenedesmus subspicatus. Cd was added in the experimental units every day using a stock solution of CdCl2. The concentration was measured in water samples collected every morning and adjusted daily by the compensated additions method, in order to maintain it constant for all the duration of the experiment. Temperature, pH, and oxygen concentrations were daily measured in the experimental units and remained constant (T °C = 18.76 ± 0.13; pH = 7.2 ± 0.05; 02 = 9.11 ± 0.03 mg/L).
Cd bioaccumulation in tissues of Margaritifera margaritifera
Cd bioaccumulation in three tissues of mussels (gills, visceral mass, and the rest of the body) was analyzed by atomic absorption spectrophotometry (AAS) with graphite furnace (M6 solaar AA spectrometer, Thermoptec). After dissecting, weighing, and drying at 50 °C during 48 h, samples were analyzed. Tissues were first digested for 3 h at 100 °C after the addition of nitric acid 63% (3 mL for 100 mg of dry tissue) in polypropylene tubes. Samples were diluted sixfold with pure water (15 mL) and stored at 4 °C until analysis. For each mineralization procedure, six certified samples (50 mg of Tort-2 (Lobster Hepatopancreas Certified Reference Material for Trace Metals) and 50 mg of Dolt-3 (Dogfish Liver Certified Reference Material for Trace Metals), NRC-CNRC) and three blanks were analyzed in order to test the validity of the method. The values obtained were systematically within the expected range. Results are expressed in μg of metal per gram of dry tissue. The detection limit is 0.1 μg/L for Cd.
Histological analyses
In order to determine precisely the sex of individuals, histological analyses were carried out on all the individuals used for the experiment (six individuals per condition). Samples of gonads not exceeding 5 mm thickness were delicately cut using a scalpel, in a standardized manner. The fixing of tissues was carried out immediately after sampling by immersion of samples in liquid of Bouin (picric acid 0.9% + formol 7.4% + acetic acid 5%) for one minimal period of 48 h at 4 °C. After several rinsings, the samples were dehydrated by successive alcohol baths of increasing degree. Complete dehydration was obtained by immersion of the samples for 1 day in a pure butanol bath renewed every 2 h. The samples were then placed in three successive paraffin baths (56 °C) at a rate of 15 min per bath. At the end of the last paraffin treatment, the samples were left on the straw mattress at ambient temperature. After solidification, blocks of paraffin were cut out in a ribbon of 10 μM thickness using a microtome (Reichert). Before coloring, the paraffin was eliminated by immersion of the blades in two baths of xylene for 5 min. The cuts were then rehydrated. Gonads were colored according to the hematoxylin/eosin method described by Gabe (1968). The determination of the stages of evolution are as follows: stage 0, sexual rest; stage 1, proliferation of the gonads; stage F female sexual maturity; and stage MF, male and female sexual maturity.
Metallothionein quantification
The aliquots of tissues of M. margaritifera stored beforehand at − 80 °C under nitrogen atmosphere were analyzed by the method of saturation by inorganic mercury, coupled with Hg determination by cold vapor AAS (Baudrimont et al. 2003). After defrosting and drying on absorbing paper, the tissue aliquots were weighed then immersed in 500 μL to 1 mL of Tris 25 mM (pH 7.2) buffer. Under nitrogen atmosphere, they were homogenized using an ultra-Turrax mechanical crusher (IKA T10). The homogenates were centrifuged to 20,000g at 4 °C for 1 h; the supernatants were then saturated with mercury (50 mg/L Hg (II); TCA 10%). The mercury not fixed to MTs was recovered by the addition of porcine hemoglobin in excess. After centrifugation of 20 min to 20,000g, the supernatant containing only MTs fixing mercury was recovered, weighed, and 100 μL were analyzed by cold vapor AAS under O2 flow (LECO AMA 254, Altec, Prague, Czech Republic) in order to determine the quantities of mercury contained in the samples. The detection limit of the device is 1 ng mercury. By calculation, the number of nmoles of sites having fixed Hg per gram of fresh weight of the sample was deduced as an estimation of the MT concentration.
Malondialdehyde determination
In the experiments, 200 μL of supernatant was mixed with 650 μL of solution of NMPI (N-methyl-2-phenylindole, according to the LPO-586 method), the whole was homogenized using a vortex, and 150 μL of chlorydric acid (HCl) 12 N was added. The mixture was incubated 1 h at 45 °C then centrifuged at 15,000g at 4 °C for 10 min. The supernatant was recovered and its absorbance (corresponding to the presence of carbocyanine formed by the presence of a molecule of MDA and two molecules of NMPI) was measured by spectrophotometry at 586 nm, thus giving the quantity of MDA present in μmol per liter of supernatant. To be able to compare various samples, the concentration of MDA was reported to the quantity of proteins (μmol per gram of protein; Legeay et al. 2005).
Gene analyses
Total RNA extraction
Total RNAs were extracted from 20 to 40 mg of gills (preserved beforehand in RNA-later and glycerol at − 80 °C) by using the Agilent kit “Absolutely RNA Miniprep Kit” following the manufacturer’s instructions.
Reverse transcription of RNA into cDNA
Reverse transcription was carried out with the “Stratascript first strand synthesis system” kit (Stratagene) using total RNA (3 to 5 μg). A total of 1 μL of oligo-dT (1 μM), 1 μL of hexanucleotides (1 μM), 0.8 μL of dNTP (10 mm), and 2 μL of 10X activity buffer was added to the RNA. This mix spent 5 min at 65 °C in the thermocycler in order to linearize RNAs. Then, 1 μL of reverse transcriptase (1 U) and 0.5 μL of RNAse-block (0.5 U) are added then heated at 42 °C for 1 h with the thermocycler to allow reverse transcription.
Real-time quantitative PCR
The quantitative PCR (qPCR) was carried out using the thermocycler LightCycler (Roche) and LightCyclerRun software 5.32. In this device, the follow-up in real time of the quantity of products of amplification is made possible by the addition of an intercalating agent: SybrGreen in the reactional medium. The reaction takes place in capillaries of glass of 20 μL prepared with (I) 17 μL of a mix of reagent consisted of 12.8-μL water, 3.2 μL of MgCl2 (25 mM), and 1 μL of reactional mixture (Taq with 5 U/mL, dNTP, 10X buffer, and SybrGreen); (II) 2 μL of a mix primers with 3 μM specific from each gene (Table 2); and (III) 1 μL of cDNA resulting from the RT. The protocol used is as follows: activation of the enzyme (95 °C, 10 min), then 50 cycles of PCR (denaturation 95 °C, 5 s; fixing of the primers 60 °C, 5 s; and elongation 72 °C, 20 s). The reaction specificity was determined for each reaction from the dissociation curve of the PCR product. This dissociation curve was obtained by following the SybrGreen fluorescence level during gradual heating of the PCR products from 60 to 95 °C. Relative quantification of each gene expression level was normalized according to the β-actin gene using the 2−ΔCT method as described by Livak and Schmittgen (2001).
Statistical treatment of data
The data processing was carried out using Statistica software (Statistica 12). After checking of the homogeneity of the variances, the factors “conditions” and “tissues” were tested by a two-way factor ANOVA or a one-way ANOVA for histological analyses. In the event of significance for one or the other of the factors taken into account, the mean comparison test (LSD Fisher) was carried out. In the event of nonconformity of the homogeneity of the variances, the non-parametric test of Kruskal–Wallis was used in order to detect the significant differences between the sites or the tissues or conditions and this, for each parameter. The different letters mentioned on the graphs correspond to significant differences with the threshold α = 0.05.
Results and discussion
Cadmium bioaccumulation
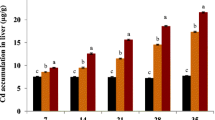
When exposed for 7 days to Cd, freshwater mussels showed significant accumulations in all the studied organs. In gills, Cd concentrations reached values of 12.4 and 17.2 μg/g (dw) under the conditions 2 and 5 μg/L, respectively, compared with 1.1 μg/g for the upstream control individuals (Fig. 1a). These accumulations corresponded to a direct route which involved exposure to the metal transferred from the dissolved phase of the water through the gill epithelia during filtration for respiratory and nutritional purposes. Cd bioaccumulation was also effective in the visceral mass and the rest of the tissues, but less pronounced, attesting a blood transfer of Cd from the gills to the internal organs. Such concentrations were dependent on the exposure dose with the highest values measured for the condition of 5 μg/L compared with 2 μg/L, especially in the gills and in the rest of the body. In contrast, concentrations measured in the conditions “downstream control” and “estradiol” showed no accumulation of this metal. The freshwater pearl mussel M. margaritifera therefore accumulated relatively large quantities of Cd, since in 7 days, it multiplied its concentration in the gills by a factor of about 15. According to Frank and Gerstmann (2007), the freshwater pearl mussel indeed accumulated trace metals including Cd easily, with concentrations in Germany of about 50 μg/g (dw) in some contaminated sites in the visceral mass, compared with measures conducted in Finland, showing concentrations of this metal around 7 μg/g (dw) in these tissues. By comparison, here, we measured values in control individuals acclimated for 14 days in the laboratory at around 2.5 μg/g (dw), which appears to be relatively low and consistent with measurements made in situ in the Dronne River upstream from the dump (Bertucci et al. 2017).

Cadmium bioaccumulation (a) and metallothionein concentrations (b) in tissues (G, gills; VM, visceral mass; R, rest of the tissues) of Margaritifera margaritifera for control individuals upstream (UC) and downstream (DC) the wild waste dump in the Dronne River, the exposure conditions with estradiol (100 μg/L) and Cd (2 and 5 μg/L) (mean ± SE, n = 6). The letters on the graph indicate significant differences between conditions
Metallothionein response
In freshwater pearl mussel M. margaritifera, concentrations of MTs measured in organs exposed to Cd 2 or 5 μg/L showed a weak response to this metal in comparison with the condition “upstream control,” which seemed very surprising given the bioaccumulation results of this metal for the species (Fig. 1b). This means that unlike most species, the freshwater pearl mussel seemed to react very little in terms of detoxification by these proteins in the presence of Cd after a 7-day exposure period. This result could suggest a particular sensitivity of the freshwater pearl mussel to this metal, given the very strong bioaccumulation levels measured in this species in the field (Frank and Gerstmann 2007).
However, a strong induction of these proteins was measured in the visceral mass of M. margaritifera downstream from the dump on the Dronne River. The induction of these proteins was at a factor of 3 compared with individuals from upstream, which was very high. Given the absence of Cd accumulation in these individual downstream relative to those upstream, we were not able to explain this very strong induction which was probably related to the presence of other metals as observed by Bertucci et al. (2017) in situ, or organic contaminants.
Malondialdehyde quantification
MDA is a marker of oxidative degradation of membrane lipids (peroxidation) following the generation of oxidative stress. The accumulation of this molecule demonstrates a severe breach of physiological functioning at the cell level, which can lead to death. In our experiment, under laboratory conditions, we did not observe any accumulation of this marker, whatever the conditions studied (data not shown). This result is positive in the sense that it shows no severe toxic attack of Cd after 7 days of laboratory exposure on membrane lipids.
Gene expression
Five major genes were analyzed (Table 1). Whatever the organ, the expression of the main gene involved in mitochondrial metabolism (cox1) was higher in individuals from downstream compared with those from upstream individuals (Table 2). For the mt gene, the highest overexpression was noted in the kidney with a very significant induction factor (1085), followed by the gills where this gene was found 36 times more expressed than in individuals from upstream, and the visceral mass (15 fold higher). The mt gene expressions are consistent with MT synthesis we observed before, notably in the visceral mass. Some differences are on the contrary observed in the gills where MT proteins are not significantly produced compared with upstream despite a significant gene induction. These results can be explained by the delays between gene expression and MT production, which was already observed in mollusks (Paul-Pont et al. 2010). In the gills, a higher expression of the sodMn gene (factor 6) was also demonstrated. The results in these two groups of individual suggested different levels of basic expression. This suggests that animals from downstream probably adapted to contamination, but the nature of which remains unclear. This contamination impacted the mitochondria and detoxification systems. The fact that the gills and kidney were the two most affected organs suggests contamination by both the direct and trophic routes. Indeed, the gills constitute the first biological barrier crossed by contaminants present in the water. The kidneys, in turn, are the organs involved in the elimination and thus the detoxification of these compounds. The fact that sodMn was overexpressed in the gills of downstream individuals also strengthens this hypothesis. This overexpression reflects a greater generation of ROS in the mitochondria of this organ.
The expression levels observed in conditions of Cd contamination were compared with those obtained from control individuals from upstream. A dose-dependent response was determined in the three organs studied. For example, in the gills, Cd caused overexpression of coxI and mt genes and also a repression of cytoplasmic sod. The same trends were found in the kidney. However, once again, the factors of inductions raised for this organ showed the highest values; 22 compared with 18 for the mt gene and 181 compared with 19.5 for the coxI gene in the kidney and gills, respectively, after exposure to 5 μg/L. In the visceral mass, the same dose-dependent overexpression of mitochondrial and detoxification genes was found. However, an induction of sodMn for a factor of 6 was also visible. These results were consistent with previous exposure to Cd studies in model organisms such as the zebrafish Danio rerio (Gonzalez et al. 2006; Orieux et al. 2011), the amphibian Xenopus laevis (Mouchet et al. 2006); or bivalves Corbicula fluminea, Crassostrea gigas, or even Cerastoderma edule (Baudrimont et al. 2003; Legeay et al. 2005; Marie et al. 2006a, 2006b; Paul Pont et al., 2010). Thus, the production of defense proteins such as metallothioneins allows the sequestering of large quantities of metal ions at intracellular level in order to reduce the toxicity of these elements with respect to other cellular targets. These proteins are inducible by many other factors, such as cytotoxic agents, reproductive factors, the presence of parasites, bacterial infections, and other xenobiotics (Kägi 1991; Baudrimont et al. 1997, 2006), which leads to then being considered more as representative of a comprehensive response to stress (Amiard et al. 2006). Cd, once in the cell, is able to inhibit the transfer of electrons along the mitochondrial respiratory chain and also to induce the production of reactive oxygen species (Wang et al. 2004; Gonzalez et al. 2006). The oxidative stress thus generated is at the origin of DNA breaks and induction of apoptotic mechanisms leading to cell death (Bagchi et al. 2000; Chan and Cheng 2003; Mouchet et al. 2006). The differential expression observed in M. margaritifera can account for these phenomena.
Unlike the Cd, the response to estradiol was fundamentally different depending on the organ. In the gills, a single increase of the 12S gene (factor 4), indicating a greater number of mitochondria and therefore an increasing energy demand, was visible. Instead, repression of this gene and of the sod gene was observed in the kidneys, associated with an overexpression of coxI. This suggests an inhibitory effect of estradiol on this organ. The increase of coxI surely compensates for this decrease in activity and thus maintains a sufficient level of ATP in the cells. These results are consistent with studies of estradiol and its metabolites which showed that these compounds can inhibit mitochondrial respiration (Hagen et al. 2004). In the kidney, estradiol caused in hamsters, through deletion and mutation mechanisms, a decline in the number of copies of mitochondrial DNA leading to a decrease in energy capacity of these organelles and an increase in the sensitivity of the kidney (Bhat 2002). Thus, this type of damage would be involved in the onset and progression of cancers in this organ. In the liver, only coxI and sodMn genes were found induced after exposure to estradiol, suggesting a greater demand for energy and the generation of oxidative stress in the mitochondria. In this context, the production of ROS has recently been observed in the metabolism of estradiol, especially in the liver (Spencer et al. 2012). This production can generate ROS damage to DNA.
Histological analyses
The histological analysis of gonads was performed on individuals from this experiment of exposure to estradiol and Cd to evaluate the potential endocrine disruptive effect of Cd, compared with the positive control condition represented by “estradiol” condition (Table 3).
The results obtained allowed us to demonstrate for control individuals, whether from upstream or downstream, a predominance of hermaphrodite individuals (0% of female individuals). It is largely accepted that freshwater pearl mussels have separate sexes; however, in adverse conditions (poor water quality, for example, or low population density), the female individuals can develop hermaphroditism to maintain their reproduction (Bauer 1987). Concerning the mussels of the Dronne River, it seems that we are in this situation, which is positive in the sense that these individuals are able to reproduce (mature gonad oocytes and sperm; Fig. 2) contrary to what we would have expected following the direct observation of the gonads (data not shown) showing a predominant presence of female individuals. However, these results suggest a reaction or adaptation of populations to their current location in the river, which can be judged in a rather unfavorable manner. We can also add from this point of view that the estradiol and Cd (2 and 5 μg/L) conditions of exposure are less adverse conditions than downstream condition, which is also confirmed by gene expressions (Table 2). Nevertheless, we must consider carefully these first results since only 11 individuals were analyzed (six in the condition “upstream control” and only five in the condition “downstream control”).

Histological observations of the gonads of freshwater pearl mussels Margaritifera margaritifera in function of the different exposure conditions: Upstream control, estradiol, Cd 5 μg/L. H, hermaphrodite individual; F, female; O, oocytes; SPZ, spermatozoids
For the results obtained from exposure to estradiol, the female sex hormone stimulating oocyte development, we saw that after only 7 days of exposure, the percentage of strictly female individuals was largely increased compared with the control conditions (33% against 0%). We have therefore a marked effect of this hormone on the oocyte development of M. margaritifera, as has already been demonstrated by Flynn and Spellman (2009) in the freshwater bivalve Elliptio complanata. Following exposure to Cd, we observed the same phenomenon, with a range effect of this metal, since the percentage of female individuals increased from 17 to 33% for 2 and 5 μg/L, respectively. From this analysis, we can observe that the Cd mimics the effect of estradiol on the development of female sexual characteristics of the freshwater pearl mussel, which attests to an endocrine disruptor effect in this bivalve. This is important given the effects already observed previously in terms of cell and molecular impact on the tissues of the freshwater pearl mussel, and facing significantly higher accumulations of Cd downstream relative to upstream from the waste dump of St Saud Lacoussière on the Dronne River in April 2009. Measured Cd accumulation still remained low compared with the levels found in Germany or Finland, but may become a problem, especially concerning endocrine disruption, because of other metals accumulated in parallel in the field, or other potential contaminants, including organics that might amplify this phenomenon as previously observed in freshwater mussels exposed to municipal effluents (Blaise et al. 2003; Gagné et al. 2004).
Conclusion
This study features a novel approach in the potential toxicological impacts of trace metals, and especially Cd, on the freshwater pearl mussel Margaritifera margaritifera from the Dronne River in France. We measured metallothionein concentrations and malondialdehyde production for the first time in this species and we quantified the relative expression of five genes implied in mitochondrial metabolism, oxidative stress, and detoxification. The results showed, after Cd exposure of the freshwater pearl mussels in the laboratory, significant ability of mussels to accumulate this metal after only 7 days of exposure, but at the same time, the relative slow response of MTs (only at the gene level but not at the protein level) suggesting a certain sensitivity of these mussels to metals. We observed also that the majority of the mussels from the Dronne River seemed to be hermaphrodite, probably in relation with the decrease of the water quality of the river or with the low density of the population. But after exposure to Cd or estradiol, we observed an increase in the amount of female individuals, confirming for this species the endocrine disruptor role of Cd. These results thus bring new knowledge on the sensitivity of this species to metal contamination, which can help to improve reintroduction strategies on the European scale, taking into account the presence of this type of contamination in the field.
References
Addy S, Cooksley SL, Sime I (2012) Impacts of flow regulation on freshwater pearl mussel (Margaritifera margaritifera) habitat in a Scottish montane river. Sci Total Environ 432:318–328
Amiard JC, Amiard-Triquet C, Barka S, Pellerin J, Rainbow PS (2006) Metallothioneins in aquatic invertebrates: their role in metal detoxification and their use as biomarkers. Aquat Toxicol 76:160–202
Arini A, Daffe G, Gonzalez P, Feurtet-Mazel A, Baudrimont M (2014) Detoxification and genetic recovery capacities of Corbicula fluminea after an industrial metal contamination (Cd and Zn): a one-year depuration experiment. Environ Pollut 192:74–82
Bagchi D, Joshi SS, Bagchi M, Balmoori J, Benner EJ, Kuszynski CA, Stohs SJ (2000) Cadmium- and chromium-induced oxidative stress, DNA damage, and apoptotic cell death in cultured human chronic myelogenous leukemic K562 cells, promyelocytic leukemic HL-60 cells, and normal human peripheral blood mononuclear cells. J Biochem Mol Toxicol 14:33–41
Baudrimont M, Andrès S, Durrieu G, Boudou A (2003) The key role of metallothioneins in the bivalve Corbicula fluminea during the depuration phase, after an in situ exposure to Cd and Zn. Aquat Toxicol 63:89–102
Baudrimont M, Andrès S, Métivaud J, Lapaquellerie Y, Ribeyre F, Maillet N, Latouche C, Boudou A (1999) Field transplantation of the freshwater bivalve Corbicula fluminea along a polymetallic contamination gradient (river Lot, France) - Part II: metallothionein response to metal exposure. Environ Toxicol Chem 18:2472–2477
Baudrimont M, de Montaudouin X, Palvadeau A (2006) Impact of digenean parasite infection on metallothionein synthesis by the cockle (Cerastoderma edule): multivariate field monitoring. Mar Pollut Bull 52:494–502
Baudrimont M, Lemaire-Gony S, Métivaud J, Ribeyre F, Boudou A (1997) Seasonal variations of metallothionein concentrations in the Asiatic clam (Corbicula fluminea). Comp Biochem Physiol 118C(3):361–367
Bauer G (1987) Reproductive strategy of the freshwater pearl mussel Margaritifera margaritifera. J Anim Ecol 56:691–704
Bauer G (1988) Threats to the freshwater pearl mussel Margaritifera margaritifera L. in Central Europe. Biol Conserv 45:239–253
Bensettiti F and Gaudillat V 2004. Cahiers d’habitats Natura 2000. Connaissance et gestion des habitats et des espèces d'intérêt communautaire. Tome 7. Espèces animales. La Documentation française, pp 318–321
Bertucci A, Pierron F, Thébault J, Klopp C, Bellec J, Gonzalez P, Baudrimont M (2017) Transcriptomic responses of the endangered freshwater mussel Margaritifera margaritifera to trace metal contamination in the Dronne River, France. Environ Sci Pollut Res 24:27145–27159. https://doi.org/10.1007/s22356-017-0294-6
Bhat HK (2002) Depletion of mitochondrial DNA and enzyme in estrogen-induced hamster kidney tumors: a rodent model of hormonal carcinogenesis. J Biochem Mol Toxicol 16:1–9
Blaise C, Gagné F, Salazar M, Salazar S, Trottier S, Hansen PD (2003) Experimentally-induced feminisation of freshwater mussels after long term exposure to a municipal effluent. Fresenius Environ Bull 12:865–870
Chan PK, Cheng SH (2003) Cadmium-induced ectopic apoptosis in zebrafish embryos. Arch Toxicol 77:69–79
Cosgrove PJ, Hastie LC (2001) Conservation of threatened freshwater pearl mussel populations: river management, mussel translocation and conflict resolution. Biol Conserv 99:183–190
Cossu C, Doyotte A, Babut M, Exinger A, Vasseur P (2000) Antioxidant biomarkers in freshwater bivalves, Unio tumidus, in response to different contamination profiles of aquatic sediments. Ecotoxicol Environ Saf 45:106–121
Couillard Y, Campbell PGC, Tessier A (1993) Response of metallothionein concentrations in a freshwater bivalve Anodonta grandis along an environmental cadmium gradient. Limnol Oceanogr 38:299–313
Couillard Y, Campbell PGC, Pellerin-Massicottee J, Auclair JC (1995) Field transplantation of a freshwater bivalve, Pyganodon grandis, across a metal contamination gradient. II. Metallothionein response to Cd and Zn exposure, evidence for cytotoxicity, and links to effects at higher levels of biological organization. Can J Fish Aquat Sci 52:703–715
Farcy E, Gagné F, Martel L, Fortier M, Trépanier S, Brousseau P, Fournier M (2011) Short-term physiological effects of a xenobiotic mixture on the freshwater mussel Elliptio complanata exposed to municipal effluents. Environ Res 111:1096–1106
Flynn K, Spellman T (2009) Environmental levels of atrazine decrease spatial aggregation in the freshwater mussel, Elliptio complanata. Ecotoxicol Environ Saf 72:1228–1233
Frank H, Gerstmann S (2007) Declining populations of freshwater pearl mussels (Margaritifera margaritifera) are burdened with heavy metals and DDT/DDE. Ambio 36(7):571–574
Gabe M 1968. Techniques histologiques. Edition Masson et Cie
Gagné F, Blaise C, Hellou J (2004) Endocrine disruption and health effects of caged mussels, Elliptio complanata placed downstream from a primary-treated municipal effluent plume for 1 year. Comp Biochem Physiol C Toxicol Pharmacol 138(1):33–44
Geist J (2010) Strategies for the conservation of endangered freshwater pearl mussels (Margaritifera margaritifera; L.): a synthesis of conservation genetics and ecology. Hydrobiologia 644:69–88
Geist J, Kuehn R (2005) Genetic diversity and differentiation of central European freshwater pearl mussel (Margaritifera margaritifera L.) populations: implications for conservation and management. Mol Ecol 14(2):425–439
Giguère A, Couillard Y, Campbell PGC, Perceval O, Hare L, Pinel-Alloul B (2003) Steady-state distribution of metals among metallothionein and other cytosolic ligands and links to cytotoxicity in bivalves living along a polymetallic gradient. Aquat Toxicol 64:185–200
Gillis PL (2012) Impacts of urban runoff and municipal wastewater effluents on the health of the freshwater mussel, Lasmigona costata. Sci Total Environ 431:348–356
Gillis PL, Higgins SK, Jorge MB (2014) Evidence of oxidative stress in wild freshwater mussels (Lasmigona costata) exposed to urban-derived contaminants. Ecotoxicol Environ Saf 102:62–69
Gonzalez P, Baudrimont M, Boudou A, Bourdineaud JP (2006) Comparative effects of direct cadmium contamination on gene expression in gills, liver, skeletal muscles and brain of the zebra fish (Danio rerio). Biometals 19:225–235
Gumpinger C, Hauer C, Scheder C (2015) The current status and future challenges for the preservation and conservation of freshwater pearl mussel habitats. Limnologica 50:1–3
Hagen T, D'Amico G, Quintero M, Palacios-Callender M, Hollis V, Lam F, Moncada S (2004) Inhibition of mitochondrial respiration by the anticancer agent 2-methoxyestradiol. Biochem Biophys Res Commun 322:923–929
Hastie LC, Young MR, Boon PJ, Cosgrove PJ, Henniger B (2008) Sizes, densities and age structures of Scottish Margaritifera margaritifera (L.) populations. Aquat Conserv Mar Freshwat Ecosyst 10:229–247
Hauer C (2015) Review of hydro-morphological management criteria on a river basin scale for preservation and restoration of freshwater pearl mussel habitats. Limnologica 50:40–53
Kägi JHR (1991) Overview of metallothionein. In: Abelson JN, Simon MI (eds) Methods in enzymology. Academic Press, San Diego, CA, pp 613–626
Legeay A, Achard-Joris M, Baudrimont M, Massabuau JC, Bourdineaud JP (2005) Impact of cadmium contamination and oxygen levels on biochemical responses in the Asiatic clam Corbicula fluminea. Aquat Toxicol 74:242–253
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 25(4):402–408
Marie V, Baudrimont M, Boudou A (2006a) Cadmium and zinc bioaccumulation and metallothionein response in two freshwater bivalves (Corbicula fluminea and Dreissena polymorpha) transplanted along a polymetallic gradient. Chemosphere 65:609–617
Marie V, Gonzalez P, Baudrimont M, Bourdineaud JP, Boudou A (2006b) Metallothionein response to cadmium and zinc exposures compared in two freshwater bivalves, Dreissena polymorpha and Corbicula fluminea. Biometals 19:399–407
Mouchet F, Baudrimont M, Gonzalez P, Cuenot Y, Bourdineaud JP, Boudou A, Gauthier L (2006) Genotoxic and stress inductive potential of cadmium in Xenopus laevis larvae. Aquat Toxicol 78:157–166
Motte G (2005) Biologie et statut de la moule perlière en Wallonie. Forêt wallonne No 74:10–16
Orieux N, Cambier S, Gonzalez P, Morin B, Adam C, Garnier-Laplace J, Bourdineaud JP (2011) Genotoxic damages in zebrafish submitted to a polymetallic gradient displayed by the Lot River (France). Ecotoxicol Environ Saf 74:974–983
Paul-Pont I, Gonzalez P, Baudrimont M, Nili H, de Montaudouin X (2010) Short-term metallothionein inductions in the edible cockle Cerastoderma edule after cadmium or mercury exposure: discrepancy between mRNA and protein responses. Aquat Toxicol 97:260–267
Perceval O, Couillard Y, Pinel-Alloul B, Giguère A, Campbell PGC (2004) Metal-induced stress in bivalves living along a gradient of Cd contamination: relating sub-cellular metal distribution to population-level responses. Aquat Toxicol 69:327–345
Pierron F, Baudrimont M, Dufour S, Elie P, Bossy A, Baloche S, Mesmer-Dudons N, Gonzalez P, Bourdineaud J-P, Massabuau J-C (2008) How cadmium could compromise the completion of the European eel’s reproductive migration. Environ Sci Technol 42:4607–4612
Pierron F, Baudrimont M, Dufour S, Elie P, Bossy A, Lucia M, Massabuau JC (2009) Ovarian gene expression and effects of cadmium pre-exposure during artificial sexual maturation of the European eel (Anguilla anguilla). BioMetals 22:985–994
Rotchell JM, Ostrander GK (2003) Molecular markers of endocrine disruption in aquatic organisms. J Toxicol Environ Health 6:453–495
Scheder C, Lerchegger B, Flodl P, Csar D, Gumpinger C, Hauer C (2015) River bed stability versus clogged interstitial: depth-dependent accumulation of substances in freshwater pearl mussel (Margaritifera margaritifera L.) habitats in Austrian streams as a function of hydromorphological parameters. Limnologica 50:29–39
Spencer WA, Vadhanam MV, Jeyabalan J, Gupta RC (2012) Oxidative DNA damage following microsome/Cu(II)-mediated activation of the estrogens, 17β-estradiol, equilenin, and equilin: role of reactive oxygen species. Chem Res Toxicol 25:305–314
Strayer DL, Downing JA, Haag WR, King TL, Layzer JB, Newton TJ (2004) Changing perspectives on pearly mussels, North America's most imperiled animals. Bioscience 54:429–439
Treasurer JW, Hastie LC, Hunter D, Duncan F, Treasurer CM (2006) Effects of Margaritifera margaritifera glochidial infection on performance of tank-reared Atlantic salmon (Salmo salar). Aquaculture journal 256:74–79
Varandas S, Lopes-Lima M, Teixeira A, Hinzmann M, Reis J, Cortes R, Machado J, Sousa R (2013) Ecology of southern European pearl mussels (Margaritifera margaritifera): first record of two new populations on the rivers Terva and Beça (Portugal). Aquat Conserv: Mar Freshw Ecosyst 23:374–389
Vaughn CC, Taylor CM (1999) Impoundments and the decline of freshwater mussels: a case study of an extinction gradient. Conserv Biol 13:912–920
Viarengo A (1989) Heavy metals in marine invertebrates: mechanisms of regulation and toxicity at the cellular level. Aquat Sci 1:295–317
Wächtler K, Dreher-Mansur MC and Richter T 2001. Larval types and early postlarval biology in naiads (Unionoida). In: Bauer G, Wäxhtler K (eds) Ecology and evolution of the freshwater mussels Unionoidea. Ecological studies, 145. Springer Verlag Heidelberg, pp. 93–125
Wang Y, Fang J, Leonard SS, Rao KMK (2004) Cadmium inhibits the electron transfer chain and induces reactive oxygen species. Free Radic Biol Med 36:1434–1443
Yang HY, Wang YM, Peng SQ (2009) Basal expression of metallothionein suppresses butenolide-induced oxidative stress in liver homogenates in vitro. Toxicon 53:246–253
Ziuganov VV (2005) A long-lived parasite extending the host life span: the pearl mussel Margaritifera margaritifera elongates host life by turns out the program of accelerated senescence in salmon Salmo salar. Dokl Biol Sci 403:291–294
Acknowledgments
We want to acknowledge the PNRPL (Parc Naturel Régional Périgord Limousin) for their cooperative assistance in the field and their interest in this study. We also thank Eric Goursolle, Julie Chevalier, Bénédicte Pécassou, and Romain Papin-Vincent for their participation in the realization of the different analyses.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Philippe Garrigues
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Baudrimont, M., Gonzalez, P., Mesmer-Dudons, N. et al. Sensitivity to cadmium of the endangered freshwater pearl mussel Margaritifera margaritifera from the Dronne River (France): experimental exposure. Environ Sci Pollut Res 27, 3715–3725 (2020). https://doi.org/10.1007/s11356-019-05025-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-019-05025-0