
Abstract
The establishment of nature reserves is a key approach for biodiversity conservation worldwide. However, the effectiveness of nature reserves established by protecting the habitat needs of surrogate species is questioned. In this study, the Baishuijiang National Nature Reserve (Baishuijiang NNR), located in the Minshan Mountains, China, which is established mainly for the conservation of giant panda (a surrogate for the conservation of other endangered species) was selected. We quantitatively evaluated the conservation effectiveness of the reserve for giant panda and co-occurring species (here, seven protected species) using a maximum entropy model (Maxent), and analyzed spatial congruence between giant panda and other seven species. Results shown that the habitat of giant panda generally included the habitat of other seven protected species, suggesting that conservation of giant panda habitat also allows the conservation for the habitat of almost co-occurring species. Hence, the natural reserve established for giant panda as a surrogate species has a relatively high effectiveness. A high proportion of the suitable habitat for six species is inside the core zone, but a high proportion of the suitable habitat for two species is located in the experimental and buffer zones. Thus, the two species are affected by human activities. To improve the conservation effectiveness of the nature reserve, the management zones need to be amended. The result of the study will be beneficial for future conservation and management of the reserve. This study provides an effective method for evaluating the conservation effectiveness of nature reserves in other area of the worldwide.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
To protect natural resources and biodiversity, nature reserves have been established worldwide (Suski and Cooke 2007; Laurance et al. 2012; Yu and Czarnezki 2013; Zheng et al. 2013; Stewart et al. 2017). However, there are too many species in the world that need to be protected directly (Williams et al. 2006). Thus, we have to rely on surrogates that allow conservationists to identify nature reserves based on the land requirement of a small number of species (Favreau et al. 2006). Usually, the location and areal extent of nature reserves have been defined based on empirical information as well as experts’ opinion on the distribution of surrogate species (Lambeck 1997; Caro and O’Doherty 1999). Ideally, once a natural reserve is built, both surrogate species and sympatric species can be protected with high effectiveness. However, the habitat needs may be different among surrogate and sympatric species. Therefore, the establishment of a nature reserve by using surrogate species has been questioned (Simberloff 1998; Andelman and Fagan 2000; Favreau et al. 2006; Grantham et al. 2010; Lindenmayer and Likens 2011; Westgate et al. 2014). Some researches considered the surrogate species measurement is disappointed (Banks et al. 2010; Wesner and Belk 2012), while others suggest it works well (Poiani et al. 2001; Brooker 2002; Lambeck 2002; Xu et al. 2014).
Assessing the effectiveness of a nature reserve is not only one of the important means to improve the sustainable management level of the nature reserve, but also a hotspot in recent research of biodiversity conservation (Kang et al. 2013; Xu et al. 2014). There are many methods to assess the effectiveness, including correlation analysis (Wei et al. 1999; Shen et al. 2009), discriminant function analysis (Kang et al. 2013), and model analysis (Xu et al. 2014). These methods can be classified into two groups: statistics methods and habitat models. Statistics methods offer some insights into the habitat preference of surrogate species and have been widely used (Williams et al. 2006), but they are only based on the point scale (Grantham et al. 2010). In contrast, habitat models can provide spatial information, but has not been widely used for the effectiveness assessment.
Many habitat models have been developed, such as the Genetic Algorithm for Rule-set Prediction (GARP) (Stockwell et al. 2006), BIOCLIM (Beaumont et al. 2005), DOMAIN (Carpenter et al. 1993), Ecological Niche Factor Analysis (ENFA) (Hirzel et al. 2002), and Maximum Entropy (Maxent) (Phillips 2006). Once a fitted model is selected, the potential distribution of suitable habitat for endangered species can be accurately defined. Among the habitat models, the Maxent uses presence-only data to predict species potential distribution and has robust prediction performance (Elith et al. 2006, 2011; Phillips 2006; Tognelli et al. 2009).
Giant panda (Ailuropoda melanoleuca) is a protected species listed in category I of the Red List of China (Wei et al. 2000). It is a flagship species for nature conservancy and one of the rarest and endangered species in the world. Giant panda is distributed in the Minshan, Qionglai, Xiangling, Liangshan, and Qinling Mountains, China (Wei et al. 2000; Hu et al. 2011). To protect the endangered species, the Chinese government has established 67 nature reserves since 1963 taking giant panda as surrogates (Tang et al. 2015; Wei et al. 2015). The largest one is the Baishuijiang NNR. However, its effectiveness for conserving the giant panda and other species has been seldom evaluated.
In this study, Baishuijiang NNR was selected. Our objectives were to (1) predict the habitat suitability of giant panda and other sympatric species, (2) evaluate the effectiveness of the present nature reserve established by giant panda as a surrogate species, and (3) design a scientific and reasonable core zone based on the surrogate and sympatric species.
Materials and methods
Study area
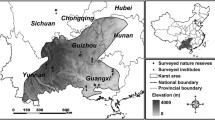
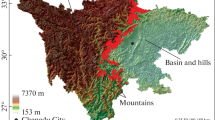
The Baishuijiang NNR located in the southern part of Gansu Province (E 104° 16′–105° 25′, N 32° 36′–33° 00′), northwest China (Fig. 1). The reserve was established in 1978 mainly to protect giant panda population and its habitat, covering an area of 1857.5 km2. The reserve is classified into three management zones, that is, core zone, buffer zone, and experimental zone. Areas of the core zone, buffer zone, and experimental zone are 917.8 km2, 278.5 km2, and 664.8 km2, respectively. The percentage of the core zone area is 49.4% to the reserve.

Location and management zones of the Baishuijiang NNR
The study area is dominated by mountains with elevation increase from southeast to northwest, ranging from 585 to 4041 m a.s.l. The climate is characterized by a transition climate between subtropical and warm-temperate types. The mean annual temperature is 14.8 °C with the monthly maximum 24.6 °C (July) and the monthly minimum 3.7 °C (January). Precipitation ranges from 840 to 950 mm in the eastern part and from 460 to 1200 mm in the western part (Zheng et al. 2014). The study area has diverse ecosystems. With increasing elevation, the sequence of vegetation type is mountain subtropical evergreen broad-leaved forests, mountain evergreen broad-leaved and deciduous broad-leaved mixed forests, mountain deciduous broad-leaved forests, warm-temperate needle-leaved and deciduous broad-leaved mixed forests, mountain cold-temperate needle-leaved forests, subalpine frigid shrubs and meadows. There are 52 species of rare animals under state protection in the reserve, including 10 species were class I protected species and 42 belonged to class II. Among those species, the population and distribution of giant panda population are relatively clear. According to the 4th national survey report of giant pandas (State Forestry Administration 2015), there exists 110 giant pandas in the reserve. Pandas mainly feed on bamboos. There are 9 species of bamboo in the reserve, of which, six species are staple bamboo of pandas (i.e., Fargesia nitida (Mitford) Keng f., Fargesia denudata Yi, F.rufa Yi, Fargesia dracocephala Yi, Fargesia scabrida Yi, and Fargesia obliqua Yi).
Species selection
We selected giant panda as a surrogate species and other seven endangered species as target species by three criteria: (1) being a protected species in the China protection class list or were the national protected terrestrial wild animals with important ecological, economic, and scientific values; (2) having different habitat requirement; and (3) having data accessibility. Eight protected species are chosen according to Table 1.
Habitat modeling
Model selection
Maxent is based on the machine-learning method to predict the distribution of species from incomplete information (Phillips 2006). A set of environmental variables and species occurrence data are needed to drive this model. The output of Maxent model is a habitat suitability index (HSI), with its value ranges from 0 (unsuitable) to 1 (perfectly suitable) (Liu et al. 2005). Maxent has been widely used in the world due to its five advantages: (1) needing only species occurrence data, which makes data to be easily collected; (2) using both continuous and categorical environmental data at the same time; (3) generating a continuous probabilistic output, that is apt to classify species suitability; (4) reducing laborious jobs in data collection with a small sample size to meet its demand (Pearson et al. 2007; Wisz et al. 2010); (5) facilitating model interpretation (Scheldeman and Zonneveld 2010). Maxent has been found to be consistently robust in its prediction performance (Tognelli et al. 2009). There are many cases of its application in predicting species distribution (e.g., Fuller et al. 2012; Reiss et al. 2011). Detailed information about the Maxent model was given by Phillips (2006), as well as Phillips and Dudík (2008).
Species occurrence data
The spatial locations of the occurrence for 8 protected species were obtained from the field surveys conducted during 2015–2016. These field surveys were carried out a whole year round (in four seasons) so that the differences in habitat preferences were observed throughout the year. There are 550 monitoring routes were arranged along elevation gradient over the study area. During these field surveys, the geographic locations of the occurrence of protected species (e.g., feces, tracks) were recorded by global positioning system receivers (GPS). A total of 833 points were obtained, including the least points (15 points) for the blood pheasant and the most points (252 points) for the panda (Table 1).
Environmental variables
Environmental variables including topography, land cover, and human disturbance were chosen based on known species habitat associations (Liu et al. 1999; Loucks et al. 2003; Xu et al. 2006; Qi et al. 2011; Zhang 2011; Hong et al. 2015; Wei et al. 2015; Hong et al. 2016, Wei et al. 2018a, b, c) (Table 2). The topographic variables (i.e., elevation, slope, and aspect) were derived from a digital elevation model (DEM) collected from the Computer Network Information Center, Chinese Academy of Sciences (http://datamirror.csdb.cn). The human disturbance variables, including hydropower stations, roads, and residential areas, were collected from the Baishuijiang NNR administration. Vegetation type was classified by the Landsat ETM data (path-row: 129–037) with 30 m spatial resolution in July 2015. The data was obtained from the Computer Network Information Center (http://datamirror.csdb.cn). A supervised classification was carried out using a maximum likelihood classification algorithm. Seven classes were obtained: deciduous forest, deciduous broad-leaf and needle-leaf mixed forests, conifer forest, shrub, meadow, farmland, and bare land. The accuracy of the classification was 92%. All category data were made buffer analysis. According to the buffer area, the distance between a cell containing evidence of 8 species habitat requirement and the closest cell containing a given feature were calculated. All analyses were performed in ENVI5.1 software and GIS 10.3. All variables running the Maxent model had spatially explicit characteristics and were transformed in UTM 48N reference system using WGS 1984 datum, and re-sampled to a common grid of 30 × 30 m.
Model validation
We used the area under the receiver operating characteristic curve (AUC) (Hanley and McNeil 1982) to evaluate the prediction accuracy. The AUC has been extensively applied as a measure of model’s overall performance (Vanagas 2004). The AUC validation procedure was implemented in the software Maxent 3.3.3 using 10,000 randomly selected background pixels that were used as pseudo-absence. We divided the occurrence data of 8 species into training and validation through a randomization procedure in which 75% of the data were used for training and the remaining 25% for validation. The AUC has the values ranging from 0 to 1. The larger AUC indicates a preferred model. Generally, based on the AUC values, model performance is categorized as failing (0.5–0.6), poor (0.6–0.7), fair (0.7–0.8), good (0.8–0.9), or excellent (0.9–1) (Swets 1988).
Analyzing the relationship between giant panda and other species habitats
Once giant panda habitat is protected, does it mean that the habitats of other 7 species are protected? To answer this question, we conducted the study with following two procedures. The first, the habitat prediction values were extracted corresponding with occurrence of species by ArcGIS 10.3. Then we calculated Spearman rank-ordered correlation coefficients between giant panda’s habitat prediction values and the other 7 species’ habitat prediction values by SPSS software. The second, we identified the suitable habitat of eight species by the minimum threshold value method (by extracting predicted values of species distribution using occurrence of species, selecting the minimum value among these values). The minimum threshold value is the most conservative threshold and can identify the minimum predicted area possible while still maintaining a zero omission rate for both training and test data (Liu et al. 2005). Ecologically, the MTP can be interpreted to contain those cells that are predicted to be at least as suitable as those where the species is identified as present. The suitable area of giant panda was overlapped by the suitable area of each targeted species. The ratio, the overlapped area to the habitat area for the targeted species, was calculated.
Evaluating the effectiveness of the nature reserve
We overlapped the suitable area of eight species with management zones of the Baishuijiang NNR. The suitable area of each species in the core zone was calculated. At the same time, we overlapped the potential distribution of 8 species and obtained the potential abundance in the study area. According to the potential abundance, we can provide suggestions to define new management zones.
Results
Potential distribution of eight species
The AUC values were all above 0.85 for eight species, suggesting a high prediction accuracy of the Maxent model. The potential distribution of eight species is shown in Fig. 2. Some species, such as giant panda, golden snob-nosed monkey, Chinese goral, and wild boar have large distribution areas, while other species such as takin, blood pheasant, red tragopan, and mainland serow exhibit comparatively smaller habitat areas. The most suitable habitat for giant panda, blood pheasant, mainland serow, and red tragopan locate in the western parts of the study area and the suitable habitat mainly distribute in southern and northeastern parts for golden snob-nosed monkey, takin, and Chinese goral. The suitable habitat for wild boar covers almost the whole study area.

The potential distribution of 8 species derived from the Maxent model. a Giant panda, AUC = 0.92. b Golden snob-nosed monkey, AUC = 0.88. c Takin, AUC = 0.93. d Blood pheasant, AUC = 0.96. e Chinese goral, AUC = 0.89. f Mainland serow, AUC = 0.90. g Red tragopan, AUC = 0.97. h Wild boar, AUC = 0.93
The potential distribution of eight species was overlapped, and the sum of the generated models for eight species was presented in the potential abundance map (Fig. 3). The potential abundance is divided into nine classes (Table 3). The area with potential abundance eight accounted for 1.4% of the reserve area, while the area with potential abundance zero accounted for 22.7% of the reserve area. Some areas with higher potential abundance were outside the core zone (e.g., yellow circles in Fig. 3).

Current core zone of the nature reserve and the distribution of potential abundance (yellow circle indicating areas of the suitable habitat outside the core zone)
Relationship between giant panda habitat and other species habitat
Spearman rank-correlation coefficients between the habitat prediction values of giant panda and that of the each other species ranged from 0.01 to 0.78 (p < 0.01) (Table 4). The coefficient was 0.78 for mainland serow and 0.54 for red tragopan, indicating high spatial congruence between the habitat of these two species and that of giant panda. In contrast, coefficients for golden snob-nosed monkey, takin, blood pheasant, and Chinese goral were small (below 0.50), implying high spatial difference between the habitat of these six species and that of giant panda.
The ratio of the overlapped area between habitat of panda and that of each other species to the total habitat area of the targeted species was calculated (Table 4). Six species had higher ratio (above 50%), with the highest ratio for blood pheasant (97.8%) and the lowest ratio for wild boar (57.3%). From Table 4, we can derive two combinations of Spearman rank-correlation coefficient and the ratio: firstly, high coefficient and high ratio (e.g., mainland serow); secondly, low coefficient and high ratio (e.g., golden snob-nosed monkey, blood pheasant). The first combination indicates the overlapped area where both species preferred. The second combination expresses the overlapped area where was suitable to both species, but only one preferred to live.
Effectiveness of the nature reserve
The suitable habitat areas of giant panda occupied 40.7% of the study area (Fig. 4a), and the proportions for golden snob-nosed monkey, takin, Chinese goral, and wild boar were a little higher than those of giant panda. The proportions of other three species were less than 40%.

Ratios of suitable habitat to total nature reserve (a); the ratios of suitable habitat in the core zone to total suitable area (b). Dotted line represents ratio of 50%. A, giant panda; B, golden snob-nosed monkey; C, takin; D, blood pheasant; E, Chinese goral; F, mainland serow; G, Red tragopan; H, wild boar
Figure 4b shows the proportions of suitable habitat areas of eight species inside the core zone to total suitable habitat area. There were high proportions for eight species, indicating that the suitable habitats of selected species were located in the core zone. This result suggests that the core zone having higher conservation effectiveness. However, red tragopan and wild boar exhibited relatively low proportions (about 60%), indicating approximately 40% suitable habitats of the two species were located outside the core zone (buffer and experimental zones), implying these two species being affected by human activities.
Discussion and conclusions
There are enormous amount of species needed to be protected, but it is difficult to build nature reserves for all protected species. Thus, it is necessary to select a surrogate species to establish natural reserves. It is commonly believed that conservation of a surrogate species will automatically be beneficial for co-occurring species (Kang et al. 2013). Hence, the use of surrogate species as a conservation management strategy has been well adopted in China. This stems from the assumption that many endangered species have similar distributions and experience the same habitat (Wesner and Belk 2012). However, whether the reserve built by a surrogate species can protect other endangered ones is a challenge (Wesner and Belk 2012). But for the moment, using surrogate species is the best way to build a nature reserve due to the realities of limited funding and information for every species (Simberloff 1998; Caro and O’Doherty 1999).
The Baishuijiang NNR was built using giant panda as a surrogate species. Our results show that the current reserve has high conservation effectiveness. On the one hand, the suitable habitat of giant panda has high spatial congruence with that of seven targeted species. On the other hand, the core zone has covered major areas of suitable habitats of the eight species. However, the extent of the present core zone still excludes the partial suitable area of selected species, especially, for red tragopan and wild boar (Fig. 3b). This suggests that the effectiveness of the nature reserve for protecting the overall biodiversity in this region needs to improve. If the partial suitable areas are excluded the core protection area, these species may be subjected to higher pressure deriving from human activities. Therefore, the boundaries of present core zone need to be modified so that the excluded suitable habitat could be involved. For instance, some areas (yellow circle in Fig. 3) should be included in the modified core zone, since these areas contain a large area of the suitable habitat of protected species. The modified core zone should be defined by the potential abundance (Fig. 3). According to the potential abundance, the boundary of the core zone can be extended (Table 3). If the potential abundance is three, the ratio of the new core zone to the total reserve area is 53.5%. The area percentage of the new core zone is 4.1% higher than the present core zone. If the potential abundance is one, the ratio of the new core zone to the total reserve area is 77.3%, and that percentage is 27.9% higher than the present core zone.
Studies have widely reported different results on the effectiveness of nature reserves established by surrogates (Grantham et al. 2010; Larsen et al. 2009). The previous study results varied depend on study area location, spatial scale, spatial resolution, type of surrogates, and analytical methods (Grantham et al. 2010). Kang (2013) revealed that the takin had not be well protected under the nature reserve focused solely on panda protection, as only 25.4% of the takin’s habitat would be protected. Xu et al. (2014) reported that the co-occurring species will be protected in the nature reserve network of Qinling Mountans built by giant panda as a surrogate, which is consistent with our results. In addition, the biodiversity of giant panda nature reserve reserves is among the highest in the temperate world (Mackinnon 2008). Wei et al. (2018) found that protecting the panda as an umbrella species and the habitat that supports it yields roughly 10–27 times the cost of maintaining the current reserves (Wei et al. 2018).
Our study found that although the major habitats of seven selected species are contained by giant panda habitats, the correlation between habitats of seven selected species and that of giant panda is low. This may be due to microhabitat separation. It has been generally accepted that microhabitat separation is the most basic form of niche partitioning in sympatric mammals as it contributes to multiple-species coexistence (Wei et al. 2017). Previous studies found that sympatric species often avoid excessive interspecific competition by partitioning resource utilization along three main niche dimensions, i.e., in space (Campbell et al. 2007), food (Siemers and Swift 2006), and activity time (Jacobs and Barclay 2009; Jiang et al. 2013). If sympatric animals want to coexist harmoniously in similar overlapping distribution areas, they need to show some niche differences in at least one spatial dimension to decrease excessive interspecific competition, e.g., in dietary differentiation, feeding sites, microhabitat selection(Wei 2018). The existence of these differences may have led to mutual adaptation of sympatric species and reduce competition to promote their long-term coexistence (Berger 1997; Zhang et al. 2009).
Evaluating the effectiveness of nature reserves was mainly based on expert opinion, correlation analysis, and discriminant function analysis (Williams et al. 2006; Shen et al. 2009). In our study, we used the habitat model that is tested to be useful. It provides a meaningful criterion for establishing boundaries not only for nature reserves but also for management zones. So the nature reserve can be scientifically and reasonably established to protect wildlife species. In addition, the approach can be carried out at different scales, as long as appropriate species occurrence and spatially explicit environmental variables are available.
Nevertheless, uncertainties still remain as to the generalizability of our approach. Firstly, only the suitable habitat of eight species was evaluated, accounting for 13.4% of the total number of protected wildlife species in the study area. The eight species cannot represent all taxa. Thus, the future studies need to incorporate as many endangered species as possible. Secondly, the model prediction accuracy varied across the model type (Guisan and Zimmermann 2000) and the availability of environmental variables. A few environmental variables were used in this study, and more environmental variables should be selected to improve the prediction accuracy. Finally, our study conducted in the Baishuijiang NNR, while other regions are underrepresented. The further research will be focused on the entire geographic range of giant panda to obtain a comprehensive evaluation on the effectiveness of the entire nature reserve network built by giant panda as a surrogate species. It will provide new insights for the planning and construction of the National Park for Giant Pandas.
References
Andelman SJ, Fagan WF (2000) Umbrellas and flagships: efficient conservation surrogates or expensive mistakes? Proc Natl Acad Sci U S A 97:5954–5959. https://doi.org/10.1073/pnas.100126797
Banks JE, Ackleh A, Stark JD (2010) The use of surrogate species in risk assessment: using life history data to safeguard against false negatives. Risk Anal 30:175–182. https://doi.org/10.1111/j.1539-6924.2009.01349.x
Beaumont LJ, Hughes L, Poulsen M (2005) Predicting species distributions: use of climatic parameters in BIOCLIM and its impact on predictions of species’ current and future distributions. Ecol Model 186:251–270. https://doi.org/10.1016/j.ecolmodel.2005.01.030
Brooker L (2002) The application of focal species knowledge to landscape design in agricultural lands using the ecological neighbourhood as a template. Landscape Urban Plan 60:185–210
Campbell P, Schneider CJ, Zubaid A, Kunz ATH (2007) Morphological and ecological correlates of coexistence in Malaysian fruit bats (chiroptera: pteropodidae). J Mammal 88(1):105–118. https://doi.org/10.2307/4126856
Caro TM, O’Doherty G (1999) On the use of surrogate species in conservation biology. Conserv Biol 13:805–814. https://doi.org/10.1046/j.1523-1739.1999.98338.x
Carpenter G, Gillison AN, Winter J (1993) Domain: a flexible modeling procedure for mapping potential distributions of plants and animals. Biodivers Conserv 2:667–680. https://doi.org/10.1007/BF00051966
Elith J, Graham CH, Anderson RP, Dudik M, Ferrier S, Guisan A, Hijmans RJ, Huettmann F, Leathwick JR, Lehmann A (2006) Novel methods improve prediction of species’ distributions from occurrence data. Ecography 29:129–151. https://doi.org/. https://doi.org/10.1111/j.2006.0906-7590.04596.x
Elith J, Phillips, SJ, Hastie T, Dudík M, Chee YE, Yates CJ (2011) A statistical explanation of MaxEnt for ecologists. Divers Distrib 17:43–57. https://doi.org/10.1111/j.1472-4642.2010.00725.x
Favreau JM, Drew CA, Hess GR, Rubino MJ, Koch FH, Eschelbach KA (2006) Recommendations for assessing the effectiveness of surrogate species approaches. Biodivers Conserv 15:3949–3969. https://doi.org/10.1007/s10531-005-2631-1
Fuller DO, Ahumada ML, Quiñones ML, Herrera S, Beier JC (2012) Near-present and future distribution of Anopheles albimanus in Mesoamerica and the Caribbean Basin modeled with climate and topographic data. Int J Health Geogr 11:13
Grantham HS, Pressey RL, Wells JA, Beattie AJ (2010) Effectiveness of biodiversity surrogates for conservation planning: different measures of effectiveness generate a kaleidoscope of variation. PLoS One 5:e11430. https://doi.org/10.1371/journal.pone.0011430
Guisan A, Zimmermann NE (2000) Predictive habitat distribution models in ecology. Ecol Model 135:147–186. https://doi.org/10.1016/S0304-3800(00)00354-9
Hanley JA, McNeil BJ (1982) The meaning and use of the area under a receiver operating characteristic (ROC) curve. Radiology 143:29–36 10.1.1.139.4820
Hirzel AH, Hausser J, Chessel D, Perrin N (2002) Ecological-niche factor analysis: how to compute habitat-suitability maps without absence data? Ecology 83:2027–2036. https://doi.org/10.2307/3071784
Hong M, Yuan S, Yang Z, Yang X, Gu X, Huang F, Zhang Z (2015) Comparison of microhabitat selection and trace abundance of giant pandas between primary and secondary forests in Liziping nature reserve, China: effects of selective logging. Mamm Biol 80(5):373–379. https://doi.org/10.1016/j.mambio.2015.05.003
Hong M, Wei W, Yang Z, Yuan S, Yang X, Gu X, Zhang Z (2016) Effects of timber harvesting on arundinaria spanostachya bamboo and feeding-site selection by giant pandas in Liziping nature reserve, China. For Ecol Manag 373:74–80. https://doi.org/10.1016/j.mambio.2015.05.003
Hu JC, Zhang ZJ, Wei FW (2011) History, current situation and prospects on nature reserve for giant pandas (Ailuropioda melanoleuce) in China. Acta Theriol Sin 31:10–14 (in Chinese). https://doi.org/10.16829/j.slxb.2011.01.004
Jacobs DS, Barclay RMR (2009) Niche differentiation in two sympatric sibling bat species Scotophilus dinganii and Scotophilus mhlanganii. J Mammal 90:879–887. https://doi.org/10.1644/08-MAMM-A-235.1
Jiang TL, Lu GJ, Sun KP, Luo JH, Feng J (2013) Coexistence of Rhinolophus affinis and Rhinolophus pearsoni revisited. Acta Theriol 58:47–53. https://doi.org/10.1007/s13364-012-0093-x
Kang D, Yang H, Li J, Chen Y (2013) Can conservation of single surrogate species protect co-occurring species? Environ Sci Pollut Res 20(9):6290–6296. https://doi.org/10.1007/s11356-013-1675-0
Lambeck RJ (1997) Focal species: a multi-species umbrella for nature conservation. Conserv Biol 11:849–856. https://doi.org/10.1046/j.1523-1739.1997.96319.x
Lambeck RJ (2002) Focal species and restoration ecology: response to Lindenmayer et al. Conserv Biol 16:549–551
Larsen FW, Bladt J, Rahbek C (2009) Indicator taxa revisited: useful forconservation planning? Divers Distrib 15:70–79. https://doi.org/10.1111/j.1472-4642.2008.00507.x
Laurance WF, Useche DC, Rendeiro J et al (2012) Averting biodiversity collapse in tropical forest protected areas. Nature 489:290–294. https://doi.org/10.1038/nature11318
Lindenmayer DB, Likens GE (2011) Direct measurement versus surrogate Indicator species for evaluating environmental change and biodiversity loss. Ecosystems 14:47–59. https://doi.org/10.1007/s10021-010-9394-6
Liu J, Ouyang Z, Taylor WW, Groop R, Tan Y, Zhang H (1999) A framework for evaluating the effects of human factors on wildlife habitat: the case of giant pandas. Conserv Biol 13(6):1360–1370. https://doi.org/10.1046/j.1523-1739.1999.98418.x
Liu C, Berry PM, Dawson TP, Pearson RG (2005) Selecting thresholds of occurrence in the prediction of species distributions. Ecography 28:385–393. https://doi.org/10.1111/j.0906-7590.2005.03957.x
Loucks CJ, Lü Z, Dinerstein E, Dajun W, Hao DW (2003) The giant pandas of the qinling mountains, China: a case study in designing conservation landscapes for elevational migrants. Conserv Biol 17(2):558–565. https://doi.org/10.2307/3095373
Mackinnon J (2008) Species richness and adaptive capacity in animal communities: lessons from China. Integr Zool 3:95–100. https://doi.org/10.1111/j.1749-4877.2008.00081.x
Pearson RG, Raxworthy CJ, Nakamura M, Peterson AT (2007) Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. J Biogeogr 34:102–117. https://doi.org/10.1111/j.1365-2699.2006.01594.x
Phillips SJ (2006) Maximum entropy modeling of species geographic distributions. Ecol Model 190:231–259. https://doi.org/10.1016/j.ecolmodel.2005.03.026
Phillips SJ, Dudík M (2008) Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation. Ecography 31:161–175. https://doi.org/10.1111/j.0906-7590.2008.5203.x
Poiani KA, Merrill MD, Chapman KA (2001) Identifying conservation-priority areas in a fragmented Minnesota landscape based on the umbrella species concept and selection of large patches of natural vegetation. Conserv Biol 15:513–522. https://doi.org/10.1046/j.1523-1739.2001.015002513.x
Qi D, Zhang S, Zhang Z, Hu Y, Yang X, Wang H, Wei F (2011) Different habitat preferences of male and female giant pandas. J Zool 285(3):205–214. https://doi.org/10.1111/j.1469-7998.2011.00831.x
Reiss H, Cunze S, König K, Neumann H, Kröncke I (2011) Species distribution modelling of marine benthos: a North Sea case study. Mar Ecol Prog Ser 442:71–86. https://doi.org/10.3354/meps09391
Scheldeman X, Zonneveld MV (2010) Training manual on spatial analysis of plant diversity and distribution. Bioversity International, Rome
Shen D, Hexun Z, Wang Y et al (2009) Winter habitats of dwarf blue sheep and goal in Batang County, Sichuan. Acta Theriol Sin 29:2320–2330 (in Chinese)
Siemers BM, Swift SM (2006) Differences in sensory ecology contribute to resource partitioning in the bats myotis bechsteinii and myotis nattereri (chiroptera: vespertilionidae). Behav Ecol Sociobiol 59(3):373–380. https://doi.org/10.1007/s00265-005-0060-5
Simberloff D (1998) Flagships, umbrellas, and keystones: is single-species management passé in the landscape era? Biol Conserv 83:247–257. https://doi.org/10.1016/s0006-3207(97)00081-5
State Forestry Administration (2015) Results of the fourth national survey on the giant panda. [EB/OL]. http://www.forestry.gov.cn/main/72/content-742880.html. Accessed 28 Feb 2015 (in Chinese)
Stewart DR, Underwood ZE, Rahel FJ, Walters AW (2017) The effectiveness of surrogate taxa to conserve freshwater biodiversity. Conserv Biol 32:183–194. https://doi.org/10.1111/cobi.12967
Stockwell DRB, Beach JH, Stewart A, Vorontsov G, Vieglais D, Pereira RS (2006) The use of the GARP genetic algorithm and internet grid computing in the Lifemapper world atlas of species biodiversity. Ecol Model 195:139–145. https://doi.org/10.1016/j.ecolmodel.2005.11.016
Suski CD, Cooke SJ (2007) Conservation of aquatic resources through the use of freshwater protected areas: opportunities and challenges. Biodivers Conserv 16(7):2015–2029. https://doi.org/10.1007/s10531-006-9060-7
Swets JA (1988) Measuring the accuracy of diagnostic systems. Science 240:1285–1293. https://doi.org/10.1126/science.3287615
Tang XP, Jia JS, Wang ZC, Zhang DH, Yu BC, Yue JB, Gong MH, Liu Y (2015) Scheme design and main result analysis of the fourth national survey on giant pandas. For Resour Manag 2015(01):11–16 (in Chinese). https://doi.org/10.13466/j.cnki.Lyzygl.2015.01.002
Tognelli MF, Roigjuñent SA, Marvaldi AE, Marvaldi AE, Flores GE, Lobo JM (2009) An evaluation of methods for modelling distribution of Patagonian insects. Rev Chil Hist Nat 82:347–360. https://doi.org/10.4067/s0716-078x2009000300003
Vanagas G (2004) Receiver operating characteristic curves and comparison of cardiac surgery risk stratification systems. Interact Cardiovasc Thorac Surg 3(2):319–322. https://doi.org/10.1016/j.icvts.2004.01.008
Wei FW, Feng ZJ, Wang ZW (1999) Habitat selection by giant pandas and red pandas in Xiangling Mountains. Acta Zool Sin 45:57–63 (in Chinese)
Wei F, Feng Z, Wang Z, Hu J (2000) Habitat use and separation between the giant panda and the red panda. J Mammal 81:448–455. https://doi.org/10.1093/jmammal/81.2.448
Wei F, Swaisgood R, Hu Y, Nie Y, Yan L, Zhang Z, Qi D, Zhu L (2015) Progress in the ecology and conservation of giant pandas. Conserv Biol 29:1497–1507. https://doi.org/10.1111/cobi.12582
Wei F, Costanza R, Dai Q, Stoeckl N, Gu X, Farber S, Zhang W (2018a) The value of ecosystem services from giant panda reserves. Curr Biol 28(13):2174–2180.e7. https://doi.org/10.1016/j.cub.2018.05.046
Wei W, Swaisgood RR, Dai Q, Yang ZS, Yuan SB, Owen MA, Pilfold NW, Yang XY, Gu XD, Zhou H, Han H, Zhang JD, Hong MS, Zhang ZJ (2018b) Giant panda distributional and habitat-use shifts in a changing landscape. Conserv Lett 11(11):e12575. https://doi.org/10.1111/conl.12575
Wei W, Han H, Zhou H, Hong M, Cao S, Zhang Z (2018c) Microhabitat use and separation between giant panda (Ailuropoda melanoleuca), takin (Budorcas taxicolor), and goral (Naemorhedus griseus) in Tangjiahe Nature Reserve, China. Folia Zool 67(3–4):198–206. https://doi.org/10.25225/fozo.v67.i3-4.a10.2018
Wesner JS, Belk MC (2012) Habitat relationships among biodiversity indicators and co-occurring species in a freshwater fish community. Anim Conserv 15:445–456. https://doi.org/10.1111/j.1469-1795.2012.00538.x
Westgate MJ, Barton PS, Lane PW, Lindenmayer DB (2014) Global meta-analysis reveals low consistency of biodiversity congruence relationships. Nat Commun 5(3899). https://doi.org/10.1038/ncomms4899
Williams P, Dan F, Manne L, Sechrest W, Preston C (2006) Complementarity analysis: mapping the performance of surrogates for biodiversity. Biol Conserv 128:253–264. https://doi.org/10.1016/j.biocon.2005.09.047
Wisz MS, Hijmans RJ, Li J, Peterson AT, Graham CH, Guisan A (2010) Effects of sample size on the performance of species distribution models. Divers Distrib 14:763–773. https://doi.org/10.1111/j.1472-4642.2008.00482.x
Xu W, Ouyang Z, Vina A, Zheng H, Liu J, Xiao Y (2006) Designing a conservation plan for protecting the habitat for giant pandas in the Qionglai mountain range, China. Divers Distrib 12(5):610–619. https://doi.org/10.1111/j.1366-9516.2006.00236.x
Xu W, Viña A, Qi Z, Ouyang Z, Liu J, Liu W, Wan H (2014) Evaluating conservation effectiveness of nature reserves established for surrogate species: case of a giant panda nature reserve in Qinling Mountains, China. Chinese Geore Sci 24:60–70. https://doi.org/10.1007/s11769-014-0656-7
Yu W, Czarnezki JJ (2013) Challenges to China’s natural resources conservation & biodiversity legislation. Soc Sci Electron Publ 43. https://doi.org/10.2139/ssrn.2133580
Zhang ZJ (2011) Old-growth forest is what giant pandas really need. Biol Lett 7:403–406. https://doi.org/10.1098/rsbl.2010.1081
Zheng ZR, Lv SH, Gao JJ, Feng CY (2013) Assessment of nature reserve management effectiveness-a case study of huihe nature reserve. Adv Mater Res 726-731:1093–1100. https://doi.org/10.4028/www.scientific.net/amr.726-731.1093
Zheng Z, Gong DJ, Zhang Q, Zhao HB (2014) Vertical patterns of plant species diversity in the Baishuijiang Nature Reserve: explanation of area, climate and boundary constraint. Chin J Appl Ecol 25:3390–3398 (in Chinese). https://doi.org/10.13292/j.1000-4890.2014.0039
Funding
The research was financially supported by the National Natural Science Foundation of China (91025015, 51178209) and Project of Arid Meteorological Science Research Foundation of China Meteorological Administration (IAM201608).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Philippe Garrigues
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Rong, Z., Liu, X., Zhao, C. et al. Evaluating giant panda as a surrogate species for conservation co-occurring species in the Baishuijiang National Nature Reserve. Environ Sci Pollut Res 26, 8939–8948 (2019). https://doi.org/10.1007/s11356-019-04420-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-019-04420-x