
Abstract
Although cell phones have been used worldwide, some adverse and toxic effects were reported for this communication technology apparatus. To analyze in vivo effects of exposure to radiofrequency-electromagnetic field (RF-EMF) on protein expression in rat testicular proteome, 20 Sprague-Dawley rats were exposed to 900 MHz RF-EMF for 0, 1, 2, or 4 h/day for 30 consecutive days. Protein content of rat testes was separated by high-resolution two-dimensional electrophoresis using immobilized pH gradient (pI 4–7, 7 cm) and 12% acrylamide and identified by MALDI-TOF/TOF-MS. Two protein spots were found differentially overexpressed (P < 0.05) in intensity and volume with induction factors 1.7 times greater after RF-EMF exposure. After 4 h of daily exposure for 30 consecutive days, ATP synthase beta subunit (ASBS) and hypoxia up-regulated protein 1 precursor (HYOU1) were found to be significantly up-regulated. These proteins affect signaling pathways in rat testes and spermatogenesis and play a critical role in protein folding and secretion in the endoplasmic reticulum. Our results indicate that exposure to RF-EMF produces increases in testicular proteins in adults that are related to carcinogenic risk and reproductive damage. In light of the widespread practice of men carrying phones in their pockets near their gonads, where exposures can exceed as-tested guidelines, further study of these effects should be a high priority.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The possible health effects of radio-frequency modulated electromagnetic fields (RF-EMF) emitted by cell phones, cordless phones, base stations, and Wi-Fi routers are a matter of growing public concern around the world today. By the end of 2015, many nations will have more cell phones than people, and the use of tablets and other wireless transmitting devices will have grown exponentially within the past few years. Current standards for these devices are based on the assumption that they can have no adverse effect unless they produce a measurable change in temperature. But, a number of research teams have reported that levels of RF-EMF which do not produce temperature changes are associated with important biological impacts. These range from increasing the production of free radicals and heat shock proteins (HSP) to impairing mitochondrial DNA and otherwise affecting human and experimental spermatogenesis (Adams et al. 2014; Agarwal et al. 2009; Azadi Oskouyi et al. 2015; De Iuliis et al. 2009).
In recent years, a number of in vitro and in vivo studies suggested that RF-EMF might impair functioning of male human reproductive organs (Agarwal et al. 2009; Celik et al. 2012; Deepinder et al. 2007; Desai et al. 2009; Forgacs et al. 2004; Iorio et al. 2007). Also, as the testes have the thinnest external membrane of the human body, and among the highest dielectric constant, they may be especially vulnerable to exposures to pulsed digital radiation. However, the mechanisms by which non-thermal levels of non-ionizing radiation can cause damage are unclear. One way to determine the biological impact of RF-EMF in male testis is to analyze changes in gene and protein expressions by using proteomics, transcriptomics, and DNA microarray (Chueca et al. 2014; Fragopoulou et al. 2012; Gerner et al. 2010; Huwiler et al. 2012; Karinen et al. 2008; Le Quément et al. 2012; Leszczynski 2013; Polidori et al. 2012; Sakurai et al. 2011).
Consistent with this literature, we previously reported that RF-EMF exposure induced pathological (Sepehrimanesh et al. 2014a), endocrine (Sepehrimanesh et al. 2014c), and some proteomic (Sepehrimanesh et al. 2014b) effects in animal models. Here, we evaluated changes in protein expression in testes of adult male rats exposed in vivo to RF-EMF for various periods of time.
Methods
Chemicals and reagents
Urea, 3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate (CHAPS), iodoacetamide, Tris, acrylamide, methylene bis-acrylamide, sodium dodecyl sulfate (SDS), ammonium persulfate (APS), tetramethylethylenediamine (TEMED), glycerol, dithiothreitol (DTT), 2-mercaptoethanol (2-ME), and bromophenol blue were purchased from Sigma-Aldrich (Munich, Germany). Seven-centimeter-long IPG strips were purchased from Bio-Rad (Hercules, CA, USA). Sodium monophosphate, hydrochloric acid (HCl), and sodium hydroxide were purchased from Merck (Darmstadt, Germany). All other chemical reagents used were of analytical grades and supplied by commercial sources.
Animals and housing
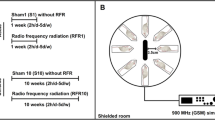
Twenty male Sprague-Dawley rats, weighing 180 ± 10 g and aged 12 weeks, obtained from the Laboratory Animal Center of Shiraz University of Medical Sciences, Shiraz, Iran, and were used in this study. All procedures were approved by local ethical committee and were performed based on animal welfare issues recommended by National Institute of Health (NIH). Animals were housed in standard transparent polycarbonate cage of 42 cm × 26.5 cm × 15 cm, without any metallic fittings under a 12-h light cycle (lights on at 7:00 AM) with an ambient temperature of 22 ± 2 °C, and 50% relative humidity and were removed from their cages every 3 days for cleaning the cages and renewing their food and water supply. After a 3-day acclimation period, rats were allocated randomly into four equal independent groups. These included one sham-exposed (without RF-EMF), and three exposure groups, based on the time of exposure to RF-EMF (1, 2, and 4 h) as short, moderate, and long-time exposure and simulation of total answering/calling time of cell phone use in human.
EMF system and exposure
Exposure at frequency of 900 MHz with the average power density of 86 mW/cm2 (22.8–146.8 mW/cm2) and an average whole body specific absorption rate (SAR) of 0.19–1.22 W/kg was experienced by freely moving animals for 30 consecutive days. The animals were awake during the exposure which was produced through an RF-EMF generating system that mimicked cell phone signals. These SAR values are only rough estimates and cannot be stressed exactly. The cage diameters were 26.5 cm × 42 cm. The RF-EMF generator antenna was placed near the cage and rats could move freely in a cage. Therefore, the shortest and longest distances from the antenna were 20 and 51 cm, respectively. The sham-exposed group was kept in a separate room with similar temperature, light, and ventilation conditions except RF-EMF signal generator. The RF-EMF exposure was started at 8 AM on each day. The use of any RF-EMF-emitting devices, such as an extracellular phone, centrifuge, and computers, was not permitted in the rooms housing experimental animals. In addition, any metal objects include cage roof and metal water container was replaced by plastic ones. Therefore, metal could not interfere with RF-EMF exposure. As an additional environmental control, the temperature was monitored in each cage during the exposure using a conventional white-backed mercury thermometer unperturbed by RF-EMF. Temperature readings were taken at 5-min intervals. The temperature of air of each experimental cage was maintained at 23 ± 1 °C using an air-cooling system.
Sampling and protein extraction
At the end of the experimental period, all rats in four groups were euthanized under deep anesthesia with ether as inhalation route, and the testes were removed from the body by non-traumatic surgery. In this method, whole testes without any damage or extra bleeding were removed. Briefly, abdominal cavity was dissected at the umbilical base, followed by a midline incision to the diaphragm, and then, both testes were removed, frozen, and stored in liquid nitrogen, until use. As soon as possible, 0.3 g of testicular tissue was placed in a crucible on ice, thoroughly minced with surgical scissors, and was then ground manually under liquid nitrogen. After that, 6 vol of a solution of 0.05 M Tris-HCl, 10% sucrose, pH 6.8, was added, and the homogenates were centrifuged at 14,000 rpm for 15 min at 4 °C to remove tissue, and cellular debris and supernatants were used as the source of protein (Sepehrimanesh et al. 2014b).
Acetone precipitation and Laemmli-SDS-polyacrylamide gel electrophoresis analysis
Acetone precipitation for removal of high abundant proteins and concentrating of sample was performed according to the methods which was described previously (Fattahi et al. 2014). SDS-PAGE was performed on a Mini Protein cell (Bio-Rad) using a 12% T gel with 0.1% SDS according to the instructions of the manufacturer. Prior to analysis, samples were boiled for 5 min in SDS sample buffer containing 0.1 M DTT, then cooled down, and applied directly to the gel, using established methods of preparation and analysis.
Two-dimensional electrophoresis
Two-dimensional proteomic map of whole testicular tissue was produced according to our previously reported procedure that relies on accepted protocols (Sepehrimanesh et al. 2014b). All preparations were made at the same time, by using one expert laboratory technician and same laboratory apparatus. An approximately high number of replicates are necessary to lessen technical and biological variability when using two-dimensional gels. Thus, three independent protein preparations were prepared from each testes of each rat in different groups. Then, each prepared sample was analyzed by 2-DE. Briefly, after dissolving of protein samples in 165 μl of gel rehydration solution at 20 °C, linear pH range 4–7 IPG strips (7 cm length) were rehydrated under passive conditions at 20 °C for 17 h in the protein IEF cell (Bio-Rad, USA). After IEF, the SDS-PAGE followed by silver nitrate staining (SNS) was performed. More than 85% of visible protein spots were in the pH range of 4–7. The IEF was carried out in six steps: (I) 0–50 V for 15 min, linear; (II) 50–100 V for 15 min, gradient; (III) 100–150 V for 15 min, gradient; (IV) 150–250 V for 15 min, gradient; (V) a linear voltage ramping step for 2 h at 4000 V; and (VI) the maximum voltage of the ramp step was maintained at up to 24,000 Vh. After IEF, the strips with focused proteins were stored overnight at −70 °C, and then equilibration with two buffers, DTT and idoacetamide. SDS-PAGE of proteins in IEF strips was done at 30 V for 15 min followed by approximately 60 min at 140 V using 12% acrylamide gels at 4 °C. Resolved proteins were visualized with modified SNS. The SNS modification omitted glutaraldehyde fixation to permit processing of extracted protein spots by mass spectroscopy.
Image analysis, spot extraction, and MALDI-TOF/TOF
Stained gels were scanned on Image Scanner (Amersham Bio-sciences). Images of scanned gels were analyzed using Melanie software, version 6.0.2.0 (Geneva Bioinformatics (GenBio), Switzerland). The proteins of interest were manually excised from the two-dimensional gels and allowed to dry at 4 °C in a cool and clean air shed. Peptides were analyzed using a MALDI-TOF/TOF-MS in reflection mode with delayed extraction. For protein identification, the mass spectra were saved in the Mascot generic file (.mgf) format, submitted to the Mascot database search engine, and the Swiss-Prot (us.expasy.org) and NCBInr (www.ncbi.nlm.nih.gov) databases were searched as reported previously (Kazemipour et al. 2016). The search parameters for Mascot-searching included NCBInr database; Metazoa (Animals) taxonomy; trypsin as enzyme; carbamidomethyl (C) as fixed modifications and oxidation (M) as variable modifications; monoisotopic mass values; unrestricted protein mass; ±100 ppm peptide mass tolerance; ±0.5 Da fragment mass tolerance; maximum missed cleavages of 1 and instrument type of MALDI-TOF-TOF (Kazemipour et al. 2015).
Statistical analysis
Values are expressed as the mean ± SD. Differences between the RF-EMF exposure and sham-exposed groups were determined by analysis of variance (ANOVA), followed by Tukey method for all pair wise multiple comparisons. P value <0.05 was considered statistically significant.
Results
After comparing the 2-DE map of testicular proteome of all groups, significant differences in expression of seven protein spots were detected in both intensity (Fig. 1) and volume (Fig. 2) percentages. Among them, our analysis has revealed four protein spots which were differentially expressed (P < 0.05) in aspects of both intensity and volume with induction factors above 1.7 that indicated the RF-EMF response (Fig. 3). Expression of the two of the proteins (spot nos. 1 and 7) was found to be up-regulated in all three exposed groups and two of the proteins (spot nos. 3 and 5) up-regulated only in LTE rats by the cell phone radiation exposure (Fig. 4). Further analysis of these protein spots by MALDI/TOF-TOF demonstrated that two of them did not match with any protein in all searched databases (spot nos. 1 and 7). However, two other proteins (spot nos. 3 and 5) were identified as ATP synthase beta subunit (ASBS) and hypoxia up-regulated protein 1 precursor (HYOU1), respectively. Properties of all four proteins are presented in Table 1.

Comparison of volume of seven differential expressed protein spots on the two-dimensional electrophoresis (2-DE) gels. The data are reproduced from three individual 2-DE gels in each experimental group. Statistically significant differences in volume of each spots between groups are identified by superscript letters. SE sham exposed, STE short time exposure, MTE moderate time exposure, LTE long time exposure

Comparison of intensity of seven differential expressed protein spots on the two-dimensional electrophoresis (2-DE) gels. The data are reproduced from three individual 2-DE gels in each experimental group. Statistically significant differences in spot intensity between groups are identified by superscript letters. SE sham exposed, STE short time exposure, MTE moderate time exposure, LTE long time exposure

Partial comparison of patterns of four differential expressed protein spots on the two-dimensional electrophoresis (2-DE) gels between different experimental groups. Statistically significant differences in volume of each spots between groups are identified by superscript letters. SE sham exposed, STE short time exposure, MTE moderate time exposure, LTE long time exposure

Histograms show the relative intensity (white square) and volume (black square) of four differential expressed protein spots on the two-dimensional electrophoresis (2-DE) gels and compared between experimental groups. The data are represented as mean ± SD from three individual 2-DE gels in each experimental group. Statistically significant differences in intensity and volume of each spots between each group are identified by uppercase and lowercase superscript letters, respectively. SE sham exposed, STE short time exposure, MTE moderate time exposure, LTE long time exposure
Discussion
This study compared the down- and up-regulated proteins in whole proteome of rat testicular tissue in response to RF-EMF exposure in vivo. The average protein spot numbers in the sham-exposed, short, moderate, and long-time exposure rats were 439, 382, 393, and 365, respectively. Among them, we found four spots that up- or down-expressed in response to RF-EMF. These include two acidic (approximate pI < 6) and two neutral (approximate pI 6 to 7) proteins with Mw ranging from 22 to 112 KDa. Two of them were not matched with any other proteins in Mascot and NCBI protein databases. However, another two proteins have regulatory roles and include ASBS and HYOU1. These two proteins, ASBS and HYOU1, have pivotal roles in energy providing across electron transport complexes of the respiratory chain and cytoprotective cellular mechanisms triggered by oxygen deprivation.
Several epidemiologic and in vivo and in vitro experiments have evaluated and highlighted the role of cell phone exposure on sperm motility and morphology (Agarwal et al. 2008; Agarwal et al. 2009; Fejes et al. 2005) reproductive endocrine (Black and Heynick 2003; Sepehrimanesh et al. 2014c) and reproductive histopathology (Celik et al. 2012; Forgacs et al. 2004) and found evidence for a substantial reduction in male fertilization potential. However, the mechanism of these effects is not known.
In previous research, we found that the 4-h exposure to 900 MHz RF-EMF affected the expression of proteins involved in regulation of oxidative stress, HSP, protein folding, cytoskeleton, and pentose phosphate pathway (Sepehrimanesh et al. 2014b). HSP also known as stress proteins are highly conserved and present in all organisms. These include several families which are named according to their molecular weight. HYOU1 is a protein that rat is encoded by the Hyou1 gene and contains 998 amino acids and belongs to HSP70 family (Meunier et al. 2002). HYOU1 has a pivotal role in cytoprotective cellular mechanisms triggered by oxygen deprivation that are critical to the testicular endoplasmic reticulum. Also, HYOU1 may play a role as a molecular chaperone and participate in protein folding (Meunier et al. 2002). We and others have reported that free radical induction due to RF-EMF exposure can cause biological effects such as oxidative stress (Sepehrimanesh et al. 2014b). HSPs are able to inhibit the release of free radicals and protect cells against destructive effects (Gordon et al. 1997).
Another protein which induced in this study in response to RF-EMF exposure was ASBS. This is a mitochondrial membrane protein that produces ATP from ADP in the presence of a proton gradient across the membrane which is generated by electron transport complexes of the respiratory chain (Boulet et al. 1989; Meyer et al. 2007). ATP synthase is a co-chaperone of HSP 90 which is known as ATPase and hydrolyzes ATP to stabilize substrates (Roe et al. 1999). Mitochondrial DNA is sensitive to damage by mutagens and predisposes mitochondria to injury on exposure of cells to genotoxic agents or oxidative stress. Damage to the mitochondrial genome can cause mutations or loss of mitochondrial gene products, or damage to some nuclear genes encoding mitochondrial membrane proteins (Bandy and Davison 1990). There are no studies about the relationship between alteration of ASBS expression and RF-EMF exposure in human and animals. Hojlund and collaborators reported that ATP synthase β-subunit was down-regulated in type 2 diabetes mellitus due to phosphorylation (Højlund et al. 2003). On the other hand, dephosphorylation of some proteins and enzymes, especially regulatory protein, in response to oxidative stress were reported (Cicchillitti et al. 2003; Davis et al. 1997; Perry et al. 1999; Zambrano et al. 2004). It seems that 900-MHz RF-EMF exposure of rat testes at 4 h for 30 consecutive days has some capacity to induce changes in phosphorylation/dephosphorylation status of ASBS and therefore induce up-expression.
Another possible mechanism that can account for these impacts may involve weakening of cell membranes. This can alter the response of the cells to environment and exert changes in protein expression as off/on and up/down regulation. For instance, Sirav and Sihan in 2009 reported that exposure to 900 and 1800 MHz could increase the permeability of blood-brain barrier (Sirav and Seyhan 2009). However, such breakdown pathways and mechanisms must be evaluated in future in vitro and in vivo studies.
Finally, it must be presented that our study has two limitations. First, the absence of control group in addition to sham-exposed group which does not allow getting control-sham comparison to estimate natural variability of the results. Second, the low number of animals per each group. Despite of these limitations, our study provides the first report showing the induction of two regulatory proteins, ATP synthase beta subunit and precursor in response to 4-h exposure to 900 MHz RF-EMF. Both the up-regulated proteins found here are co-chaperones or HSPs, a family of molecules that arise in response to abnormal conformations. HSPs prevent non-specific aggregation and support repair and maintenance of functional structure (Frydman 2001). This work is a preliminary report, and further, complementary studies should be pursued using other methods such as Western blot, immunohistochemistry, and/or real-time PCR. Also, complete sequencing of the two unnamed proteins we have noted and other complementary analysis for identifying these proteins is in order.
References
Adams JA, Galloway TS, Mondal D, Esteves SC, Mathews F (2014) Effect of mobile telephones on sperm quality: a systematic review and meta-analysis. Environ Int 70:106–112
Agarwal A, Deepinder F, Sharma RK, Ranga G, Li J (2008) Effect of cell phone usage on semen analysis in men attending infertility clinic: an observational study Fertility and Sterility 89:124–128
Agarwal A, Desai NR, Makker K, Varghese A, Mouradi R, Sabanegh E, Sharma R (2009) Effects of radiofrequency electromagnetic waves (RF-EMW) from cellular phones on human ejaculated semen: an in vitro pilot study. Fertil Steril 92:1318–1325
Azadi Oskouyi E, Rajaei F, Safari Variani A, Sarokhani MR, Javadi A (2015) Effects of microwaves (950 MHZ mobile phone) on morphometric and apoptotic changes of rabbit epididymis. Andrologia 47:700–705. doi:10.1111/and.12321
Bandy B, Davison AJ (1990) Mitochondrial mutations may increase oxidative stress: implications for carcinogenesis and aging? Free Radic Biol Med 8:523–539. doi:10.1016/0891-5849(90)90152-9
Black DR, Heynick LN (2003) Radiofrequency (RF) effects on blood cells, cardiac, endocrine, and immunological functions. Bioelectromagnetics 24:S187–S195
Boulet D, Poirier J, Cote C (1989) Studies on the biogenesis of the mammalian ATP synthase complex: isolation and characterization of a full-length cDNA encoding the rat F1-beta-subunit. Biochem Biophys Res Commun 159:1184–1190
Celik S, Aridogan IA, Izol V, Erdogan S, Polat S, Doran S (2012) An evaluation of the effects of long-term cell phone use on the testes via light and electron microscope analysis. Urology 79:346–350
Chueca B, Pagan R, Garcia-Gonzalo D (2014) Transcriptomic analysis of Escherichia coli MG1655 cells exposed to pulsed electric fields. Innovative Food Sci Emerg Technol. doi:10.1016/j.ifset.2014.09.003
Cicchillitti L, Fasanaro P, Biglioli P, Capogrossi MC, Martelli F (2003) Oxidative stress induces protein phosphatase 2A-dependent dephosphorylation of the pocket proteins pRb, p107, and p130. J Biol Chem 278:19509–19517
Davis DR, Anderton BH, Brion JP, Reynolds CH, Hanger DP (1997) Oxidative stress induces dephosphorylation of τ in rat brain primary neuronal cultures. J Neurochem 68:1590–1597
De Iuliis GN, Newey RJ, King BV, Aitken RJ (2009) Mobile phone radiation induces reactive oxygen species production and DNA damage in human spermatozoa in vitro. PLoS One 4:e6446. doi:10.1371/journal.pone.0006446
Deepinder F, Makker K, Agarwal A (2007) Cell phones and male infertility: dissecting the relationship. Reprod BioMed Online 15:266–270
Desai NR, Kesari KK, Agarwal A (2009) Pathophysiology of cell phone radiation: oxidative stress and carcinogenesis with focus on male reproductive system. Reprod Biol Endocrinol 7:1–9
Fattahi S, Kazemipour N, Hashemi M, Sepehrimanesh M (2014) Alpha-1 antitrypsin, retinol binding protein and keratin 10 alterations in patients with psoriasis vulgaris, a proteomic approach. Iranian Journal of Basic Medical Sciences 17:651–655
Fejes I, Závaczki Z, Szöllosi J, Koloszár S, Daru J, Kovacs L, Pal A (2005) Is there a relationship between cell phone use and semen quality? Systems Biology in Reproductive Medicine 51:385–393
Forgacs Z et al (2004) Effects of whole-body 50-Hz magnetic field exposure on mouse Leydig cells. Sci World J 4:83–90
Fragopoulou AF et al (2012) Brain proteome response following whole body exposure of mice to mobile phone or wireless DECT base radiation. Electromagnetic Biology and Medicine 31:250–274
Frydman J (2001) Folding of newly translated proteins in vivo: the role of molecular chaperones. Annu Rev Biochem 70:603–647
Gerner C, Haudek V, Schandl U, Bayer E, Gundacker N, Hutter HP, Mosgoeller W (2010) Increased protein synthesis by cells exposed to a 1,800-MHz radio-frequency mobile phone electromagnetic field, detected by proteome profiling. Int Arch Occup Environ Health 83:691–702
Gordon SA, Hoffman RA, Simmons RL, Ford HR (1997) Induction of heat shock protein 70 protects thymocytes against radiation-induced apoptosis. Arch Surg 132:1277–1282
Højlund K et al (2003) Proteome analysis reveals phosphorylation of ATP synthase β-subunit in human skeletal muscle and proteins with potential roles in type 2 diabetes. J Biol Chem 278:10436–10442
Huwiler SG et al (2012) Genome-wide transcription analysis of Escherichia coli in response to extremely low-frequency magnetic fields. Bioelectromagnetics 33:488–496
Iorio R et al (2007) A preliminary study of oscillating electromagnetic field effects on human spermatozoon motility. Bioelectromagnetics 28:72–75
Karinen A, Heinävaara S, Nylund R, Leszczynski D (2008) Mobile phone radiation might alter protein expression in human skin. BMC Genomics 9:77
Kazemipour N, Qazizadeh H, Sepehrimanesh M, Salimi S (2015) Biomarkers identified from serum proteomic analysis for the differential diagnosis of systemic lupus erythematosus. Lupus 24(6):582–587
Kazemipour N, SalehiInchebron M, Valizadeh J, Sepehrimanesh M (2016) Clotting characteristics of milk by Withania coagulans: proteomic and biochemical study international. Journal of Food Properties. doi:10.1080/10942912.2016.1207664
Le Quément C et al (2012) Whole-genome expression analysis in primary human keratinocyte cell cultures exposed to 60 GHz radiation. Bioelectromagnetics 33:147–158
Leszczynski D (2013) Effects of radiofrequency-modulated electromagnetic fields on proteome. In: Radiation Proteomics. Springer, Netherlands, pp 101–106
Meunier L, Usherwood YK, Chung KT, Hendershot LM (2002) A subset of chaperones and folding enzymes form multiprotein complexes in endoplasmic reticulum to bind nascent proteins. Mol Biol Cell 13:4456–4469
Meyer B, Wittig I, Trifilieff E, Karas M, Schagger H (2007) Identification of two proteins associated with mammalian ATP synthase. Mol Cell Proteomics 6:1690–1699
Perry G et al (1999) Activation of neuronal extracellular receptor kinase (ERK) in Alzheimer disease links oxidative stress to abnormal phosphorylation. Neuroreport 10:2411–2415
Polidori E et al (2012) Gene expression profile in cultured human umbilical vein endothelial cells exposed to a 300 mT static magnetic field. Bioelectromagnetics 33:65–74
Roe SM, Prodromou C, O’Brien R, Ladbury JE, Piper PW, Pearl LH (1999) Structural basis for inhibition of the Hsp90 molecular chaperone by the antitumor antibiotics radicicol and geldanamycin. J Med Chem 42:260–266
Sakurai T, Narita E, Suzuki Y, Taki M, Miyakoshi J (2011) Microarray analysis of human-derived glial cells exposed to 2.45 GHz microwave. In: Microwave Workshop Series on Innovative Wireless Power Transmission: Technologies, Systems, and Applications (IMWS), 2011 I.E. MTT-S International. IEEE, pp 105–108
Sepehrimanesh M, Azarpira N, Saeb M, Nazifi S, Kazemipour N, Koohi O (2014a) Pathological changes associated with experimental 900-MHz electromagnetic wave exposure in rats. Comp Clin Pathol 23:1629–1631
Sepehrimanesh M, Kazemipour N, Saeb M, Nazifi S (2014b) Analysis of rat testicular proteome following 30-day exposure to 900 MHz electromagnetic field radiation. Electrophoresis 35:3331–3338
Sepehrimanesh M, Saeb M, Nazifi S, Kazemipour N, Jelodar G, Saeb S (2014c) Impact of 900 MHz electromagnetic field exposure on main male reproductive hormone levels: a Rattus norvegicus model. Int J Biometeorol 58:1657–1663
Sirav B, Seyhan N (2009) Blood-brain barrier disruption by continuous-wave radio frequency radiation. Electromagnetic Biology and Medicine 28:215–222
Zambrano CA, Egaña JT, Núñez MT, Maccioni RB, González-Billault C (2004) Oxidative stress promotes τ dephosphorylation in neuronal cells: the roles of cdk5 and PP1. Free Radic Biol Med 36:1393–1402
Acknowledgements
This work was supported by Research Council of Shiraz University, Shiraz, Iran (Grant No. 71-GR-VT-5). Animals were kindly provided by Dr. Mahjoob Vahedi at the Laboratory Animal Center of University of Medical Science, Shiraz, Iran. Proteomic analysis was kindly performed with the cooperation of Biotechnology Research Center of Shiraz University, Shiraz, Iran. Mr. Omid Koohi-Hosseinabadi provided expert technical assistance with animal handling and sampling.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Responsible editor: Philippe Garrigues
Rights and permissions
About this article

Cite this article
Sepehrimanesh, M., Kazemipour, N., Saeb, M. et al. Proteomic analysis of continuous 900-MHz radiofrequency electromagnetic field exposure in testicular tissue: a rat model of human cell phone exposure. Environ Sci Pollut Res 24, 13666–13673 (2017). https://doi.org/10.1007/s11356-017-8882-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-017-8882-z