
Abstract
The aim of this study was to investigate the overall root/shoot allocation of metal contaminants, the amount of metal removal by absorption and adsorption within or on the external root surfaces, the dose-response of water hyacinth metal uptake, and phytotoxicity. This was examined in a single-metal tub trial, using arsenic (As), gold (Au), copper (Cu), iron (Fe), mercury (Hg), manganese (Mn), uranium (U), and zinc (Zn). Iron and Mn were also used in low-, medium-, and high-concentration treatments to test their dose effect on water hyacinth’s metal uptake. Water hyacinth was generally tolerant to metallotoxicity, except for Cu and Hg. Over 80 % of the total amount of metals removed was accumulated in the roots, of which 30–52 % was adsorbed onto the root surfaces. Furthermore, 73–98 % of the total metal assimilation by water hyacinth was located in the roots. The bioconcentration factor (BCF) of Cu, Hg, Au, and Zn exceeded the recommended index of 1000, which is used in selection of phytoremediating plants, but those of U, As, and Mn did not. Nevertheless, the BCF for Mn increased with the increase of Mn concentration in water. This suggests that the use of BCF index alone, without the consideration of plant biomass and metal concentration in water, is inadequate to determine the potential of plants for phytoremediation accurately. Thus, this study confirms that water hyacinth holds potential for a broad spectrum of phytoremediation roles. However, knowing whether these metals are adsorbed on or assimilated within the plant tissues as well as knowing their allocation between roots and shoots will inform decisions how to re-treat biomass for metal recovery, or the mode of biomass reduction for safe disposal after phytoremediation.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Phytoremediation
The innovative technology of phytoremediation dates back more than three centuries, although its popularity only surfaced from the mid-1970s onwards (Henry 2000). Phytoremediation is broadly defined as the direct use of green plants to reduce environmental contaminants and render them safer (Gerhardt 2009; Paz-Alberto and Sigua 2013). Many aquatic plants have the ability to remove heavy metals and metalloids from water, and of special interest are those with a high tolerance to metals in terms of plant growth rate and biomass production (Sasmaz and Obek 2009). Such growth characteristics are often associated with alien invasive aquatic weeds, and thus, some have been utilized to improve water quality (Malik 2007).
Several aquatic weeds have shown significant ability to remediate heavy metal contaminants in water. For instance, duck weed, Lemna gibba L., is used in constructed wetlands for wastewater treatment and efficiently accumulates large amounts of heavy metal pollutants (Vaillant et al. 2004). The small water fern, Azolla caroliniana, removed about 93 % of 0.1 mg/L mercury (Hg) from polluted water in just 12 days (Bennicelli et al. 2004), while 99.8 % of 0.5 mg/L Hg was removed after 3 weeks by parrot’s feather (Myriophyllum aquaticum), creeping primrose (Ludwigia palustris), and water mint (Mentha aquatica) (Kamal et al. 2004).
Eichhornia crassipes (Mart.) Solms is an aquatic plant originating from South America, which also grows rapidly in tropical and subtropical regions of the world (Center and Spencer 1981). It is a notorious aquatic weed that can survive in a wide range of environmental conditions and possesses an extremely aggressive and invasive nature in areas where it has been introduced (Newete and Byrne 2016). Under ideal conditions, E. crassipes has the capacity to double its biomass within 7–10 days (Malik 2007). A single plant of E. crassipes with six to seven leaves produces a single new leaf per week on average (Center and Spencer 1981; Byrne et al. 2010). Despite its weedy status, this species is one of the most researched aquatic macrophytes for phytoremediation of both eutrophic waters and heavy metal-polluted waters and has been used as such in a number of water resources and wetlands (Liao and Chang 2004; Malik 2007; Newete and Byrne 2016; Mishra et al. 2008). This is largely attributed to its exceptionally high growth rate and large biomass both below and above water. Eichhornia crassipes in a constructed wetland in Taiwan removed large amounts of lead, copper, and zinc (Liao and Chang 2004). Roldán (2002) reported metal removal of over 90 % from aluminum factory effluents by water hyacinth.
Eichhornia crassipes accumulates higher concentrations of heavy metals in its roots than in its shoots (Lu et al. 2004; Liao and Chang 2004; Rahman and Hasegawa 2011). For instance, Kay et al. (1984) found that the concentrations of Pb, Cu, and Cd in E. crassipes decreased in a descending order of roots > stems > leaves after being exposed to varying initial concentrations of 1, 2.5, or 5 mg/L for 6 weeks. Lu et al. (2004) also showed that the highest concentration of cadmium (2044 mg/kg) and zinc (9652.1 mg/kg) was sequestered by the roots of water hyacinth as compared to the shoot system (113.2 and 1926.7 mg/kg, respectively), from Cd and Zn concentrations of 4 mg/L Cd and 40 mg/L Zn in water, respectively. Nevertheless, there is little information on root vs shoot metal accumulation or adsorption vs absorption of metals in water hyacinth plant tissues, which are investigated in this study.
The effects of heavy metals on plant growth
Symptoms of heavy metal phytotoxicity in most aquatic plants are more conspicuous on the leaves. This is because excess heavy metals disrupt photosynthetic and metabolic processes through the inhibition of electron transport at the redox sites in photosystems I and II (Fernandes and Henriques 1991). This generates reactive oxyradicals, leading to “oxidative stress,” that react and decompose membrane lipid peroxides (Fernandes and Henriques 1991; Smolders and Roelofs 1996). Heavy metals also interfere with the function of several metalloenzymes, such as those involved in the dark reaction of photosynthesis (Stiborová et al. 1986). Severe metal-induced stress in plants reduces the rate of oxygen consumption and CO2 and ATP production due to the inhibition of catabolic pathways in the metabolic processes (Lösch and Köhl 1999). Mild metal stress, however, often increases the rate of oxygen consumption. For instance, Prasad et al. (2001) showed that while the exposure of Lemna trisulca (Araceae) to a concentration of 10 mM cadmium (Cd) significantly decreased the net photosynthesis and oxygen consumption, exposure to concentrations of Cd ranging from 1 to 5 mM resulted in an increase in the rate of respiration. This is however, associated to the increased energy demand required for ionic transportation at the plasma membranes or the production of specific metal-binding polypeptides and subsequent removal of such metal-bound chelates from the sensitive organelles in the cytoplasm (Lösch and Köhl 1999). Such strategy of metal tolerance, however, can cost plants up to 20–50 % reduction in productivity compared to a non-mettalliferous plant (Lösch and Köhl 1999).
Among several symptoms of metal phytotoxicity, leaf chlorosis and necrosis, stunted growth, and waterlogging of tissues are very common (Kay et al. 1984; Yruela 2005; Xiong et al. 2006; Burkhead et al. 2009). These, however, depend on the ion species and concentration of the metal concerned. The natural concentration of Cu in fresh water does not usually exceed 0.002 ppm and ranges between 0.05 and 0.2 mg/L in natural waters contaminated with acid mine drainage (Fernandes and Henriques 1991). While the normal Cu concentration range is 3–20 mg/kg dry weight (dw), for most crop and non-metallophyte plant species (Nriagu 1979; Clarkson and Hanson 1980), concentrations exceeding this range in most aquatic plants are toxic. Similarly, Chaney (1989) indicated that the normal range of inorganic arsenic in plants is 0.01–1 mg/kg dw, while the phytotoxic concentration ranges between 3 and 20 mg/kg dw.
The tolerance mechanism in water hyacinth, as in many aquatic macrophytes, largely depends on localizing heavy metals removed from water in their roots and limiting their transportation to the aerial system particularly from the most susceptible plant parts, the photosynthetic apparatus in the shoots (Newete and Byrne 2016). Heavy metals are either adsorbed on the surface of water hyacinth roots or compartmentalized in cell vacuoles and the intercellular wall once taken in and kept out of reach from the interior cells of the bundle sheath to prevent their transportation to the shoots where the damage could be detrimental (Hossain et al. 2012). Metal leakage through the roots of water hyacinth into the growth medium and leaf detachment are also among some of the tolerance mechanisms used by water hyacinth to reduce metal toxicity.
A plant species’ efficiency for phytoremediation is determined by the index of their bioconcentration factor (BCF) and is computed as the final metal concentration in the plant tissues divided by the initial metal concentrations in water (Zhu et al. 1999). According to Zhu et al. (1999), aquatic plants capable of accumulating 5000 mg of metals per kilogram of dry plant biomass or those with BCF >1000 are considered as good accumulators of heavy metals, and this BCF value widely has been used as a criterion in several studies to determine plants that are potentially good candidates for phytoremediation.
Although the metal uptake and removal capacity of many aquatic plants have been well documented, most studies are limited to few metal contaminants such as Cd, Pb, Hg, and Zn in isolation and conducted in beakers with a single plant or clonal material. This limits their usefulness for determining phytoremediation potential. This study used a large container approach with single metal solutions and used some of these above metals to benchmark others such as uranium and gold for the first time to compare their relative uptake by E. crassipes. Agitation of the growth medium similar to water movement in slow flowing rivers in the field was used to determine metal removal by E. crassipes in conditions similar to those in the natural system. These methods were used to investigate the root/shoot metal allocation, the ratio of metal assimilated within to that adsorbed on the external root surfaces, the dose effect on the metal uptake by water hyacinth, and its tolerance to metal toxicity.
Materials and methods
Laboratory experiments were conducted in 65-l plastic tubs, as single-metal trials. The new plastic tubs were first conditioned with sulfuric acid (pH 1.5) for 7 days then thoroughly washed with tap water, rinsed, and dried, before the Eichhornia crassipes were moved into the tubs for the trials. The acidic water was neutralized with sodium hydroxide (NaOH) after the tub conditioning and disposed of. The water hyacinth used in the tubs was transplanted from a pond at the University of the Witwatersrand, Johannesburg, South Africa, and was originally obtained from Delta Park, Johannesburg, 2 years prior to the experiment. Water hyacinth plants were grown with a single metal or metalloid treatment in each tub. The tubs were housed under a clear, non-UV screening, greenhouse plastic tent (UVA CLEAR 200MIC; Vegtech 2000, Cape Town, South Africa). The trial was run for 3 weeks between late spring and early summer of 2011, during the active growing season of water hyacinth. The plants were at the “bulbous” phenostage (short, green, and healthy plants) and were left to grow for a week in the water amended with the nutrient solution to acclimatize before the addition of the metal treatments.
Experimental setup
Three replicates of a total of 39 tubs in 13 treatments were arranged randomly in four rows. Tubs were filled with 45 l of tap water, and one-fourth strength of Hoagland solution, based on literature reviews (Zhu et al. 1999; Weiss et al. 2006; Rajan et al. 2008; Hussain et al. 2010), was added to each tub using a plastic syringe and stirred thoroughly with a plastic rod. Each tub was equipped with a submersible fish tank pump (flow rate 400 l/h model PH400; power head pump) to agitate all treatments.
Ten short, green, healthy non-clonal individuals of E. crassipes of the same phenostage with average fresh biomass weight of 1.2 kg were washed and rinsed three times by dipping plants in a sequence of three glass containers filled with tap water and then were added to each tub. All metal/metalloid treatments were added to each tub in the same way as the Hoagland solution, except that the plants were first raised out of the water before adding the treatments to facilitate the stirring process (and to limit immediate adsorption). All metal compounds used were of chemically pure (CP) analytical grade from the Associated Chemical Enterprises (ACE, Southdale, South Africa). The metals added were to a final concentration of 1 mg/L Au, 2 mg/L Cu, 0.5 mg/L Fe, 1 mg/L Hg, 0.5 mg/L Mn, 1 mg/L U, and 4 mg/L Zn as well as the metalloid 1 mg/L As. A nested dose-response experiment was performed for Fe and Mn only, at concentrations corresponding to low (Mn-L and Fe-L), medium (Mn-M and Fe-M), and high (Mn-H and Fe-M) of 0.5, 2.0, and 4.0 mg/L, respectively. Plants in the control treatment were only grown with the Hoagland solution without the addition of any metal or metalloid. Water loss from each tub due to evapo-transpiration was compensated for by topping up each tub every 4 to 6 days with tap water. The experiment was run for a total of 3 weeks, starting from the day when first the metal treatments were added into the tubs.
Sample preparation for water analysis
Water samples were taken at the start of the experiment immediately after adding the metal treatments and again after 3 weeks (at the end of the trial). All water samples were collected in 250-ml plastic jars and were preserved with 1 % acetic acid and stored in a refrigerator in the lab at a temperature of 4 °C. Before the analysis, all samples were filtered with filter paper (100 % cotton fiber, 0.19 mm thickness, and with filtration speed of 29 s/100 ml) (Macherey-Nagel (MN), GmbH & Co. KG, Düren, Gremany).
Sample preparation for plant tissue analysis
Plant samples were collected at the end of the experiment (3 weeks after the addition of the metal treatments) from each tub (three tubs per treatment). The outer leaves (the petiole and lamina) from each plant sample were then removed with the exception of the last three leaves at the center of the plant. This is because nutrient and other metal elements are concentrated in the most actively growing young leaves at the center of the plant. This portion of the plant was separated into roots and shoots, and then each of these was bisected with a plastic knife into two halves (resulting in two root samples and two shoot samples) to determine the root/shoot metal assimilation. One half of each root and shoot was washed three times in Milli-Q deionized water, while the other half of each sample was first washed in deionized Milli-Q water followed by two washes in Milli-Q water acidified with acetic acid (CH3OOH) to pH 3.5 and finally rinsed in deionized water, for the analysis of metals removed only by root or shoot absorption. The four samples prepared from each three plants per tub were sealed in individual plastic bags, labeled, and stored at −20 °C until transferred to a freeze drier. After 2 weeks in the freeze drier, each sample was ground and placed in a 40-mL plastic jar, sealed, and sent for analysis to the chemistry department, at the University of the Witwatersrand. The inductively coupled plasma-optical emission spectroscopy (ICP-OES) was used for the analysis of all the heavy metals, except for Hg which was analyzed using flow injection atomic spectrometry (FIAS).
Bioconcentration factor
The BCF in this study was calculated as the final metal concentration in plant tissues divided by the initial metal concentration in the medium (water).
Root and shoot adsorption
The adsorption on the surface of the plant tissues was calculated as the difference between the metal bioaccumulation for the Milli-Q-washed and the acidified water (pH-3.5)-washed root or shoot. For the purposes of this assay, the assumption was made that washing with water acidified to pH 3.5 would displace surface-bound metal cations.
Plant growth measurements
Different plant growth parameters were recorded at the start of the experiment and 3 weeks after the addition of specific metal treatments in each single-element tub trial. The mass of all the 10 plants in each tub was recorded at the start (week 0 or day 0) and end of the experiment (week 3 or day 21). In addition, the lengths of leaf number 2 (leaf-2 petiole) and of the root were measured on three randomly selected plants per tub. The youngest petioles at the center (petiole number 1) of each of two plants in each tub were tagged at the start of the experiment, and the position of that leaf was recorded at the end of the experiment to evaluate the rate of leaf production per plant per week. The numbers of ramets per plant were counted from all the plants in each tub. A total of nine leaves per treatment (three leaf-2 from each tub) were also traced in outline onto an A4 paper, and the area of each leaf was measured from a cutout of that outline using a LI-3100 Area Meter (LI-COR, Inc., Lincoln, Nebraska, USA 68504).
Data analysis
Comparisons of the same plant parameters were made between the start and end of the experiment. These were tested by one-way ANOVA (analysis of variance) followed by Fisher’s least significant difference (LSD) post hoc test, except for the root length data, where Kruskal–Wallis one-way ANOVA was used to test the difference between the two measuring events. This is because the P value for the normality test was <0.05, unlike in all the other growth plant parameters. Comparisons of metal treatments with the control treatment were also analyzed using a Mann–Whitney non-parametric U test, comparing two independent sets of samples. STATISTICA and Six Sigma (StatSoft Release 7, 2006) and Microsoft Office Excel 2007 were the computer packages used for the data analysis (Table 1).
Results
Removal of metals from the water
Analysis of metal concentrations in the water samples showed that 7 of the 12 heavy metal treatments showed a significant decrease in their water concentration after 3 weeks in contact with water hyacinth, compared to their initial water concentration at the beginning of the experiment, just after the addition of the metal treatments (P < 0.001). These include Au, Cu, Hg, Mn-L, Mn-M, Mn-H, and Zn (Table 2). The final concentrations of Au, Hg, Zn, and Mn-L in the tub water were not significantly different from the respective metal concentrations in the control treatment (Table 2). The concentrations of the metals initially added to the tubs dropped by over 79 % in the remaining water after plant harvest at the end of week 3, with the exception of the iron dose-response treatments (Fe-L, Fe-M, and Fe-H) and the arsenic treatment. The greatest percentage reduction was found for Hg followed by Mn-H, Mn-M, and Mn-L in descending order.
Adsorption, assimilation, and allocation of metals by E. crassipes
A significant amount of each metal was removed from water by precipitation on the surface of the roots. Copper, with 52 % adsorption on the surface of the roots from the total amount of metal removed by water hyacinth roots, ranked the highest of all, while the others, with the exception of As and Fe, fell between a range of 31 to 47 % adsorption onto the root surfaces (Table 3). In contrast, 73–98 % of the total amount of metal or metalloid assimilated by the whole plant was sequestered in the roots, with both Cu and Hg assimilating the highest (97 and 98 %, respectively). Thus, this study showed that over 80 % of the metals removed from water were allocated in the roots, with most of the Au and U metals being accumulated in the roots, since their concentration in the shoots was below the detectable limits (Table 3).
In the case of Fe and Mn dose-response treatments, their assimilation by the roots and shoots of E. crassipes increased with the increase of each metal concentration in the water, although the amount of assimilation of both metals was significantly lower in the shoots compared to the roots (Table 4). No metal was added to the control treatment other than those included in the Hoagland solution. Nevertheless, the pattern of Cu, Fe, Mn, and Zn concentrations found in the roots or shoots of the controls was the same as that in the metal treatments where the root/shoot allocation of the metals was significantly higher in the roots than in the shoots, with the exception of Cu in the control (F (1, 4) = 3.3284, P < 0.142). Arsenic, Au, and U concentrations in the shoots of the metal treatments were below the detectable limit of the OES-ICP method of analysis (Table 4).
Bioconcentration factor of E. crassipes
The BCF was higher in the iron dose-response treatments than all the other heavy metal treatments in the tubs, despite the fact that the initial Fe concentration in the water did not show any significant change even after their exposure to water hyacinth plants for 3 weeks (see Table 2). Nevertheless, their BCF pattern decreased with the increase in Fe concentration in the water and the highest BCF was recorded from the low (Fe-L) iron treatment (Table 3). In contrast, the BCF in the manganese dose-response treatment increased with the increase of concentration from low to high (Mn-L to Mn-H) treatments. In addition, the BCF for Au, Cu, Hg, and Zn treatments was over 1000, whereas that of arsenic was the lowest of all. Although U concentration in the shoots was below the detectable limit, the BCF was calculated from the U concentration in the roots, and it was 670 (Table 3).
Metal impact on plant growth parameters
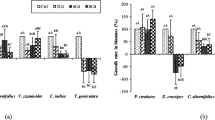
The mean fresh weight of water hyacinth plant biomass increased significantly after 3 weeks of metal treatment between the different treatments (F (12, 26) = 2.2039, P < 0.045) (Fig. 1a). An increase in biomass weight of between 41 and 75 % was observed at the end of the experiment in week 3 from the initial weight of 1.2 kg/tub, before the addition of metal treatments into the tubs at the beginning of the experiment (week 0). However, the increase was significantly less only in Cu, Hg, and Zn treatments compared to the control treatment (Fig. 1a). Considering the dose-response treatments of iron and manganese metals, no significant difference was found in the fresh biomass weight as a response to the varying concentrations of the two metal treatments. Similarly, the mean area of leaf-2 showed a significant difference between treatments at the end of week 3 after the plant exposure to the metal/metalloid treatments (F (12, 26) = 4.9338, P < 0.001) and a leaf area increase of up to 37 % was observed in most treatments, except in Cu, Hg, Mn-M, and Mn-H treatments, where a decrease of up to 46 % was found in contrast to their initial leaf area in the respective treatments before their exposure to the metal treatments in Wk0 (Fig. 1b). Unlike in the fresh biomass weight, the dose-response treatments of manganese and iron showed significant changes in the mean leaf area at the end of the experiment in week 3 (Fig. 1b). The leaf area decreased with the increase of metal concentrations in the growth medium of the water hyacinth plants.

The percentage change in plant growth parameters of water hyacinth before and after the exposure of plants to each metal treatment in a single-element tub trial for 3 weeks. a Percentage change in mean fresh weight of plant biomass. b Percentage change in mean area of leaf number 2. c, d Percentage changes in mean lengths of leaf-2 petiole and roots, respectively. Means compared by one-way ANOVA and those followed by the same letter(s) are not significantly different (P > 0.05; Fisher’s LSD test). Ctrl denotes the control treatment without metal addition, and the suffixes L, M, and H denote low, medium, and high concentrations, respectively
The length of leaf-2 also showed a significant increase of up to 21 % 3 weeks after the metal treatments, except in Cu, Au, and Fe-L treatments, which showed a decrease contrary to the other treatments (F (12, 65) = 1.9328, P < 0.046) (Fig. 1c). The Cu-treated plants have the greatest decrease (by 16 %) of all the other treatments.
The roots were the only plant parameter that did not show any significant change between the treatments, although there was an increase of between 53 and 152 % in length at the end of the experiment in week 3 (KW-H(12,78) = 5.382, P > 0.943) (Fig. 1d). The root length in the Cu treatment increased by 66 % from the initial root length of 16.1 cm (in week 0) as opposed to 111 % in the control treatment from an initial root length of 13 cm.
The leaf production recorded per plant at the end of week 3 did not show any significant difference between the treatments (F (12, 65) = 1.0556, P > 0.411) (Fig. 2a). The mean number of ramets per plant, however, showed a significant difference between treatments in week 3 (F (12, 65) = 2.4819, P < 0.009) (Fig. 2b). The Cu and Hg treatments followed by Au, Mn-M, and Mn-H treatment revealed significantly lower numbers of ramets than the control treatment at week 3. The number of ramets did not show significant differences between the Fe dose-response treatments, unlike in the manganese treatments. The number of ramets in the high manganese concentration (Mn-H) treatment was significantly lower than that in the low concentration (Mn-L) treatment (Fig. 1b).

The effect of heavy metals on plant growth parameters of water hyacinth in a single-element tub trial 3 weeks (week 3) after the addition of metal treatments. a Mean number of leaf production per plant per week. b Mean number of daughter plants (ramets) per plant. Means compared by one-way ANOVA and those followed by the same letter(s) are not significantly different (P > 0.05; Fisher’s LSD test). NB: Ctrl denotes the control treatment without metal addition, and the suffixes L, M, and H denote low, medium, and high concentrations, respectively
Discussion
The phytoremediation capacity of E. crassipes
Removal of metals from the water
Over 62 % of all the initial metal concentrations in the water were removed by water hyacinth after 3 weeks, with the exception of Fe, which increased in the water in the low and medium Fe dose treatments, as opposed to being removed by water hyacinth. The initial and final pH of the growth medium was between 6.2 and 7.5 (not presented here), which is within the suitable range of pH for optimum adsorption of metal ions on the roots of water hyacinth (Jayaweera et al. 2008). Thus, the increased Fe concentration in the growth medium after being incubated with water hyacinth for 3 weeks suggests Fe leakage from the plant to the medium. Eichhornia crassipes has the ability to leak excess iron into the growth medium to avoid iron toxicity (Sutcliffe 1962). Iron is a micronutrient and plants require low concentrations of Fe (0.6 mg/L in Hoagland solution). Tolerant plants retain most heavy metals in their roots, where the toxicity is minimal, while others are adapted to reduce shoot metal toxicity by active excretion of cations into the medium. Win et al. (2002) showed an increased rate of an iron uptake in water hyacinth plants with iron deficiency and a decreased rate as the plant cells were saturated with iron, with a possible iron leakage into the medium in the case of iron oversaturation.
The total amount of Fe assimilated to the shoots of E. crassipes in the control treatment was 13,865 mg/kg (the sum of total root and shoot up take of Fe-L) (Table 4). This Fe was taken up into the water hyacinth shoots while they were still in the common culture pool treated with technical fertilizers and iron chelates (Fe 11 %), prior to their transfer into the tubs for the experiment. Jayaweera et al. (2008) found a progressive decline in the removal of Fe by water hyacinth from a heavy metal concentration of 9.3 mg/L Fe when the nutrient levels of total nitrogen (TN) and total phosphorous (TP) in the solution were altered to six different regimes, from 0 to nutrient concentrations of 28 and 7.7 mg/L, respectively. They found a maximum removal of 6707 mg/kg Fe under the nutrient starving conditions (no N and P). The current study was conducted under relatively higher nutrient conditions with concentrations of 126 mg/L N and 31 mg/L P (Table 1) compared to the above literature. Thus, the insignificant changes in the initial Fe concentration of water after its incubation with water hyacinth for 3 weeks could be the result of the nutrient level in the growth medium and the readily oversaturated Fe level in the tissue of water hyacinth prior to the start of the experiment. The slight increase observed in the initial Fe concentration of the water at the end of the experiment (Table 2) can therefore be due to iron leakage from the plants into the test medium. Moreover, the release of iron into the medium could also be from decaying root and shoot tissues that detached from the mother plant, either due to metal toxicity or senescence. In another study, a 40 % increase of Fe concentration in water, 4 weeks after water hyacinth showed effective removal of Fe in the 6th week, was concluded to be due to effluxing and leaching from tissues affected by severe necrotic and chlorotic stresses due to Fe toxicity (Jayaweera et al. 2008). Similarly, Mishra et al. (2008) found a slight increase in Hg and As concentrations in the growth medium at 25 days compared to their values at day 20, which, as they also concluded, was a result of metal discharge from the decaying water hyacinth plant tissues. A prolonged exposure of plants to excess heavy metals eventually leads to the adoption of different physiological and molecular mechanisms that enhance plant tolerance to metal toxicity. Among these are adaptive strategies such as exclusion of heavy metals from plants, cellular exclusion of heavy metals, and complexation and compartmentations of heavy metals in cell walls and cell vacuoles, respectively (Hossain et al. 2012). Thus, it was no surprise to see a spike in the Fe concentration as opposed to their removal in the Fe dose-response treatment of this study as a result of Fe leakage through the roots or leaching from decayed or stressed leaves of water hyacinth.
The arsenic analysis was repeated by three different accredited laboratories, which all presented the same results. The initial concentration of arsenic in the water, collected just after the addition of the metal, could not be matched to the amount of arsenic (1 ppm) originally added to the tubs at the start of the experiment. The initial and final concentrations of arsenic did not show a significant difference between them. However, Mishra et al. (2008) found that the removal of arsenic by water hyacinth exposed to coal mine effluent for 21 days was 80 %. The unchanging arsenic concentrations found in this study could therefore be due to the ICP-OES analytical method being inappropriate. The ICP-MS analysis is the generally preferred method for lower or trace metal concentrations in water (Dunn 2007). Nevertheless, arsenic analysis, even with ICP-MS, is fraught with difficulties in establishing accurate results from samples with concentrations below 1 ppm (Dunn 2007). In addition, As has a strong affinity towards iron plaques, on the surface of the water hyacinth roots, and this could impede its uptake from the surface of the roots of water hyacinth. The As affinity to the iron plaque, however, depends on its species. The As(V) species is a characteristic feature of oxic conditions, unlike the reduced form of As, the arsenite species (As(III)), which is more soluble and toxic to plants (Kim et al. 2002). The tubs in this trial were equipped with submersible pumps, ensuring that the water was well aerated, enough to oxidize the As(III) added to the tubs, to As(V). This would result in adsorption of As(V) by the iron plaques on the detached root and other plant materials at the base of the tub and the reduction of the As uptake by plants and its transportation into the aerial parts (Rahman and Hasegawa 2011).
The root/shoot metal allocation of E. crassipes
Over 80 % of the total amount of metal removed from water by the whole water hyacinth plant in this study was located in the roots. For instance, 99 % of the Cu removed was accumulated in or on the roots. This also agrees with the results of Kay et al. (1984) who found 92 % Cu accumulation in the roots of water hyacinth after exposure to a concentration of 2.5 mg/L for 3 weeks. The disparity between the two Cu concentrations in the roots of water hyacinth could however be due to the difference in the initial Cu levels that the plants were exposed to, in the respective growth medium of the two studies. This is because the overall removal of heavy metals generally increases with the decrease of their concentrations in the growth medium (Liao and Chang 2004). This can be illustrated by the removal pattern of Mn observed in Table 3 where its removal by the roots of water hyacinth decreased with the increase of Mn concentration in the solution. The bioconcentration of both Hg and Zn in the roots were also 98 and 90 %, respectively. Riddle et al. (2002) also found an accumulation of 79 % Hg in the roots of water hyacinth when the plant was exposed to similar Hg (1 L Hg) and nutrient (one fourth of Hoagland solution) concentration as in the current study, although the difference in the incubation period could have accounted for the disparity observed in Hg accumulations between the two studies. The Zn bioconcentration in the roots also agrees with those found in the literature. For instance, Liao and Chang (2004) found that the Zn accumulation in the roots of water hyacinth was up to five times that of the shoot.
Over 97 % of the overall Cu absorbed by water hyacinth plant was located inside the roots. According to Vesk et al. (1999,) heavy metals such as Cu and Zn are localized in cell walls, cell vacuoles, and epidermal cell granules in association with the anionic P and/or S elements which eventually are complexed with phytochelatin, and their further encroachments into the interior cells of the bundle tissue are minimized or inhibited. Thus, only 3 % of Cu was assimilated by the shoots of water hyacinth. Furthermore, a record of up to 52 % precipitation of the heavy metals on the root surface from the total amount of metals removed from water by roots of water hyacinth was found in this study, and the maximum adsorption was observed in Cu followed by Mn (up to 47 %) and Zn (40 %) (Table 3). Although root plaques are the common characteristics of plants grown in flooded sediments of wetlands, free-floating aquatic macrophytes such as water hyacinth are also known to form iron root plaques (Vesk et al. 1999). Thus, we think that the high amount of Cu, Mn, and Zn removal by adsorption is partly attributed to the fact that root plaques are also known to contain these metals besides its main Fe component (Ye et al. 1997; Otte et al. 1989). This is also probably true for uranium uptake, whose overall accumulation in the roots of water hyacinth was 1340 mg/kg dw, of which 31 % was adsorbed on the external root surface (Tables 3 and 4). There is no literature found on the U uptake to compare this U concentration in water hyacinth with other studies. The fact that U accumulation in the shoots was below the detectable limit and no uranium-induced plant stress was also observed suggests that water hyacinth exclusion of U or other metals is part of the plant’s adaptive strategy that enables it to overcome phytotoxicity from excess metal uptake.
The bioconcentration factor of E. crassipes
Half of the metal treatments in this study exceeded the minimum BCF limit of 1000, as demarcated by Zhu et al. (1999) as plant selection criteria in terms of feasibility for phytoremediation, and this includes Au, Cu, Hg, and Zn. The BCF for Fe in the iron dose treatments shown in Table 3 does not represent the actual BCF of Fe by water hyacinth, since no Fe was removed from the water in any of the three dose-response treatments. In fact, the initial Fe concentration in each of the Fe treatments remained either unchanged or was slightly greater than the originally starting value in the water. Thus, although the high level of Fe (13,865 mg/kg) accumulated in the water hyacinth plant tissues as indicated in the control plant in Table 4 (the sum of the last two columns against the Fe-L treatment) indicates the enormous capacity of water hyacinth in the phytoremediation of Fe contaminant, it happened prior to the start of the experiment while the plants were still in the growth and preparation phase in a common iron-fertilized pool.
The BCF of all the manganese dose-response treatments was below 1000. However, it increased with increasing Mn concentration in the water, suggesting that the plants could be an effective accumulator at concentrations greater than those used in this trial (4 mg/L Mn). This also agrees with the Mn results of Tejeda et al. (2010), although their Mn concentration in the solution was lower than those in the current studies (Table 5). Similarly, the comparison between the BCF results of Hg in this study and those of Lenka et al. (1992) in Table 5 shows that the BCF for Hg in the literature with an Hg concentration of 0.004 mg/L was 2.5 times greater than that in the current study with an Hg concentration of 1.05 mg/Lin the water hyacinth growth medium. This is also true for the BCFs of As, Cu, and Zn when compared with the respective BCFs in the literature in Table 5, where lower metal concentration means higher index of BCF. Such trend of BCF is confined not only to water hyacinth but also to other aquatic macrophytes such as Oenanthe javanica (Bl.) DC., which showed an increasing trend of BCF for Hg with the increase of Hg concentration in the growth medium (Wang et al. 2002). Nevertheless, this is not always true and the BCF trend depends on the target metal species and its concentration in the growth medium and the period of incubation (Lu et al. 2004). For instance, the BCF for Cd in water hyacinth and the duckweed, Lemna minor L., decreased with the increase of Cd concentration in the solution. Therefore, our results indicate that the determination of potential aquatic plants for phytoremediation based on the BCF criteria alone, as set by Zhu et al. (1999), is not inadequate and unreliable. This is because the bioconcentration of heavy metals depend on their concentrations as well as the nutrient conditions of the growth medium among others.
The effect of heavy metals on water hyacinth plant growth
Generally, the water hyacinth plants continued to grow under most heavy metal treatments, although different levels of stress were observed. Copper followed by Hg reduced the growth of E. crassipes. The leaf production, however, remained unaffected under all the metal treatments with an average production of one leaf per plant per week.
The prominent consequence of excess heavy metal uptake by plants is excessive production of reactive oxygen species (ROS) that destabilizes the overall plant metabolic processes and leads to severe plant stresses (Fernandes and Henriques 1991). Although plants have adapted to boost the production of counteractive enzymes such as peroxidase (POX) to eliminate ROS, most of them succumb to the challenges of excessive ROS generated as a result of enormous amount of heavy metal uptake (Malar et al. 2014). Thus, different plants have different tolerance mechanisms. One such example is the exclusion of heavy metals either from the plant or the inner root cells by localizing them in root cell walls and epidermal and vacuolar granules bound to phytochelatins (Jayaweera et al. 2008). The removal of heavy metals in this study by root surface adsorption of up to 52 % from the overall metal removal by the roots and metal removal by root absorption of up to 98 % from the overall metal assimilated in the whole plant tissues provides evidence of the metal exclusion adaptive strategies of water hyacinth developed to overcome severe stress from excessive metal uptake. This particularly makes sense the fact that only less than 2 and 3 % of the total Cu and Hg assimilation by water hyacinth plant, respectively, were translocated in the shoots. Thus, the water hyacinth plants seem to generally tolerate the metal uptake in this study, although some symptoms of stress were observed in some plant parameters. This could however be partly due to the effect of heavy metals in the uptake of some nutrient elements, because of competition or their replacement once they enter the root cells. For instance, uranium and copper decreased the movement of potassium (K) into Azolla filiculoides and reduced their content inside the root cells of the plant, which concluded to be due to leakage of K from the plant into the growth medium (Sela et al. 1988).
The root length in the Hg treatment was significantly reduced compared to some of the metal treatments, among which were Zn, Fe-M, Fe-H, Mn-L, and Mn-H. Unlike these metals, however, Hg does not have any vital role in plant metabolism (Dunn 2007). The Hg concentration in the roots was 58 times greater than the Hg concentration in the shoot system (Table 4). The roots of water hyacinth have impressive ability to bind and accumulate Hg (Wolverton and McDonald 1975; Mishra et al. 2008; Chattopadhyay et al. 2012). Although accumulation of heavy metals in the roots of most aquatic plants is a strategy for avoiding phytotoxicity, their effect on the root permeability, by altering the uptake of nutrient elements, is unavoidable. Excess Cu in roots can also damage the cell wall and cell membrane and compromise the root’s selective permeability, enhancing passive flows of some metals into the root tissues (Fernandes and Henriques 1991). After the addition of the metals to the tub water, only plants in the Cu treatment showed significantly smaller increases in root length compared to the control treatment. This suggests that the roots of water hyacinth are sensitive to the toxic effects of Cu. Kay et al. (1984) also showed similar results where Cu at concentrations of 2.5 mg/L in water inhibited the growth of new water hyacinth roots and disrupted the root functions. Although the concentration of Cu in water in this study was lower, 2 mg/L, Cu still inhibited the root growth. Such effects on root growth was also reported by Hasan et al. (2007), where the growth of new roots was inhibited when water hyacinth was exposed to Cd and Zn at concentrations of 1 and >4 mg/L, respectively, for 16 days. Thus, this study confirms that Cu concentrations less than those used in literature (≥2.5 mg/L) leads to severe plant stress and inhibits the growth of water hyacinth plants. However, the zinc concentrations of 4 mg/L used in this study agree with results in the literature. For instance, Lu et al. (2004) found an increase in the relative growth rather than symptoms of stress in water hyacinth plants with treatments of 5 and 10 mg/L Zn incubated between 8 and 12 days. Nevertheless, the effects of some metals occur over an extended period of exposure to plants. A reduction of 30 % in biomass weight was observed when water hyacinth was incubated at a concentration of 9 mg/L for 24 days (Delgado et al. 1993).
Copper and Hg were the only two treatments that showed a significant reduction in the mean area of leaf-2 and the plant biomass fresh weight at the end of the experiment compared to the control treatments. For instance, the final Cu concentration in the shoots was more than twice the normal range of 3–20 mg Cu/kg dw for most plant species (Nriagu 1979; Clarkson and Hanson 1980). Therefore, it was not surprising to see that most of the plant parameters revealed stunted and stressed water hyacinth due to the Cu phytotoxicity. These results also agree with those of Yruela (2005), Xiong et al. (2006), and Burkhead et al. (2009) who also showed that an increased ionic Cu concentration in the shoot system resulted in stunted root growth, reduced shoot development, leaf chlorosis, and disruption of plant photosynthesis in different plant species. Nevertheless, this study gives evidence of Cu toxicity at concentrations less than those indicated in the above literature. For instance, the extreme Cu-induced stress observed in the Chinese cabbage, Brassica pekinensis Rupr., by Xiong et al. (2006) in aquatic culture experiment was at a concentration 10.3 μmol/L of Cu.
Despite the negative effect of Cu and Hg on several plant parameters, the leaf production rate was unaffected. The fact that the water hyacinth plant was able to maintain the normal rate of leaf production (1 leaf/plant/week (Center and Spencer 1981; Byrne et al. 2010)), across the different heavy metal treatments regardless of the metal toxicity level, is an evidence of its wide adaptation and resilience to grow under polluted water systems. Water hyacinth sheds a leaf (older leaf) with the growth of a new one every week (Center and Spencer 1981). In addition, metal-contaminated leaves show early chlorotic and necrotic symptoms which cause decay and detaching of leaves from the mother plant. This is indicated by the fact that the fresh weight of plant biomass in the Cu and Hg treatments in the current trial was the lowest of all the treatments, which suggests that shedding of more contaminated leaves was a result of heavy metal toxicity, and probably shedding of leaves is faster in the severely stressed plants than those with little or no toxicity stress. This, however, contradicts with the findings of Xiong et al. (2006) who tested different plants, where the number of leaves produced per plant was significantly lower in the Chinese cabbage B. pekinensis exposed to a concentration of 10.3 μmol/L of Cu for 14 days. This study therefore concludes that different plants have different levels of tolerance to metal toxicity and water hyacinth is generally tolerant to most of the metals tested.
Conclusion
The assimilation of 73–98 % of the total amount of metal absorbed by E. crassipes was in the roots, of which 30–52 % was adsorbed onto the external root surface. This suggests a mechanism by which the plant is generally tolerant to metal toxicity. Nevertheless, Cu and Hg were the most stressful metals to the plant’s growth because of their toxic effects on the photosynthetic and metabolic processes even at trace amounts in the plant shoots. While generally the efficiency with which water hyacinth removes heavy metals from water is indicated to increase when metal concentrations in water are low, or in trace amounts, the results of this study from the Mn dose-response treatments confirm otherwise. Although four (Cu, Hg, Au, and Zn) out of eight metals for which their BCF was calculated showed an index of >1000, a demarcating criterion for selection of potential phytoremediating plants according to the literature, the fact that the BCF index for Mn increased with the increase of their concentration in the growth medium suggests that using the BCF index alone to determine potential plants for phytoremediation could be insufficient. Thus, this study indicates that water hyacinth is generally an effective accumulator of these metals including U, As, Fe, and Mn and can be used for phytoremediation of metal-contaminated waters under controlled culture conditions or if the weed infestation already pre-exists in the water system targeted for phytoremediation. Understanding whether metals are adsorbed on or assimilated within the plant tissues and understanding their allocation between root and shoot can inform on decisions on how to re-treat biomass for metal recovery, or the mode of biomass reduction for safe disposal after phytoremediation.
References
Bennicelli R, Banach A, Szajnocha K, Ostrowski J (2004) The ability of Azolla caroliniana to remove heavy metals (Hg(II), Cr(III), Cr(VI)) from municipal wastewater. Chemosphere 55:141–146
Burkhead JL, Reynolds KA, Abdel-Ghany SE, Cohu CM, Pilon M (2009) Copper homeostasis. New Phytol 182:799–816
Byrne MJ, Hill MP, Robertson M, King A, Jadhav A, Katembo N, Wilson J, Brudvig R, Fisher J (2010) Integrated management of water hyacinth in South Africa: development of an integrated management plan for water hyacinth control, combining biological control, herbicidal control and nutrient control, tailored to the climatic regions of South Africa. Report to the Water Research Commission, Pretoria, South Africa.
Center TD, Spencer NR (1981) The phenology and growth of water hyacinth (Eichhornia crassipes (Mart.) Solms) in a eutrophic North-Central Florida lake. Aquat Bot 10:1–32
Chaney RL (1989) Toxic element accumulation in soils and crops: protecting soil fertility and agricultural food-chains. In: Bar-Yosef B, Barrow NJ, Goldshmid J (eds) Inorganic contaminants in the Vadose Zone. Springer, Berlin
Chattopadhyay S, Fimmen RL, Yates BJ, Lal V, Randall P (2012) Phytoremediation of mercury and methyl mercury-contaminated sediments by water hyacinth (Eichhornia crassipes). Int J Phytorem 14:142–161
Clarkson DT, Hanson JB (1980) The mineral nutrition of higher plants. Ann Rev Plant Physiol 31:239–298
Delgado M, Bigeriego M, Guardiola E (1993) Uptake of zinc, chromium and cadmium by water hyacinth. Wat Res 27:269–272
Dunn CE (2007) Biogeochemistry in mineral exploration. In: Hale M. (Ed), January, The Netherlands. 21–26, 232–324.
Fernandes JC, Henriques FS (1991) Biochemical, physiological, and structural effects of excess copper in plants. Bot Rev 57(3):246–273
Gerhardt KE, Huang X-D, Glick BR, Greenberg BM (2009) Phytoremediation and rhizoremediation of organic soil contaminants: potential and challenges. Plant Sci 176(1):2–30
Hasan SH, Talat M, Rai S (2007) Sorption of cadmium and zinc from aqueous solutions by water hyacinth (Eichchornia crassipes). Bioresour Technol 98:918–928
Henry JR (2000) In an overview of the phytoremediation of lead and mercury. NNEMS Report. Washington, D.C. Pp. 3–9.
Hossain MA, Piyatida P, da Silva JAT, Fujita M (2012) Molecular mechanism of heavy metal toxicity and tolerance in plants: central role of glutathione in detoxification of reactive oxygen species and methylglyoxal and in heavy metal chelation. J Bot 37 pages.
Hussain ST, Mahmood T, Malik SA (2010) Phytoremediation technologies for Ni++ by water hyacinth. Afr J Biotechnol 9(50):8648–8660
Jayaweera MW, Kasturiarachchi JC, Kularatne RKA, Wijeyekoon SLJ (2008) Contribution of water hyacinth (Eichhornia crassipes (Mart.) Solms) grown under different nutrient conditions to Fe-removal mechanisms in constructed wetlands. J Environ Manag 87:450–460
Kamal M, Ghalya AE, Mahmouda N, Cote R (2004) Phytoaccumulation of heavy metals by aquatic plants. Environ Int 29:x1029–x1039
Kay SH, Haller WT, Garrard LA (1984) Effects of heavy metal on water hyacinths (Eichhornia crassipes (Mart) Solms). Aquat Toxicol 5:117–128
Kim MJ, Ahn KH, Jung Y (2002) Distribution of inorganic arsenic species in mine tailings of abandoned mines from Korea. Chemosphere 49:307–312
Lenka M, Panda KK, Panda BB (1992) Monitoring and assessment of mercury pollution in the vicinity of a chloralkali plant. IV. Bioconcentration of mercury in in situ aquatic and terrestrial plants at Ganjam, India. Arch Environ Contam Toxicol 22:195–202
Lösch R, Köhl KI (1999) Plant respiration under the influence of heavy metals. In: Prasad MNV, Hagemeyer J (eds) Heavy metal stress in plants—from molecules to ecosystems. Springer, Berlin, pp. 139–156
Liao S-W, Chang W-L (2004) Heavy metal phytoremediation by water hyacinth at constructed wetlands in Taiwan. J Aquat Plant Manag 42:60–68
Lu X, Kruatrachue M, Pokethitiyook P, Homyok K (2004) Removal of cadmium and zinc by water hyacinth, Eichhornia crassipes. Sci Asia 30:93–103
Malar S, Vikram SS, Favas PJC, Perumal V (2014) Lead heavy metal toxicity induced changes on growth and antioxidative enzymes level in water hyacinths [Eichhornia crassipes (Mart.). Bot Stud 55:54
Malik A (2007) Environmental challenge vis a vis opportunity: the case of water hyacinth. Environ Int 33:122–138
Mishra VK, Upadhyay AR, Pathak V, Tripathi BD (2008) Phytoremediation of mercury and arsenic from tropical opencast coalmine effluent through naturally occurring aquatic macrophytes. Water Air Soil Pollut 192:303–314
Newete SW, Byrne MJ (2016) The capacity of aquatic macrophytes for phytoremediation and their disposal with specific reference to water hyacinth. J Environ Sci and Research Pollut. doi:10.1007/s11356-016-6329-6
Nriagu JO (1979) The global copper cycle. In: Nriagu JO (ed) Copper in the environment. Part I: ecological cycling. Wiley, New York, pp. 1–17
Otte ML, Rozema J, Koster L, Haarsma MS, Broekman RA (1989) Iron plaque on roots of Aster tripolium L.: interaction with zinc uptake. New Phytol 111: 309–317.
Paz-Alberto AM, Sigua GC (2013) Phytoremediation: a green technology to remove environmental pollutants. Am J Clim Chang 2:71–86
Prasad MNV, Malec P, Waloszek A, Bojko M, Strzałka K (2001) Physiological responses of Lemna trisulca L. (duckweed) to cadmium and copper bioaccumulation. Plant Sci 161:881–889
Rahman MA, Hasegawa H (2011) Aquatic arsenic: phytoremediation using floating macrophytes. Chemosphere 83:633–646
Rajan M, Darrow J, Hua M, Barnett B, Mendoza M, Greenfield BK, Andrews JC (2008) Hg L3 XANES study of mercury methylation in shredded Eichhornia crassipes. Environ Sci Technol 42:5568–5573
Riddle SG, Tran HH, Dewitt JG, Andrews JC (2002) Field, laboratory, and x-ray absorption spectroscopic studies of mercury accumulation by water hyacinths. Environ Sci Technol 36:1965–1970
Roldán G (2002) Treating industrial wastes in Colombia using water hyacinth. Waterlines 21:6–8
Sasmaz A, Obek E (2009) The accumulation of arsenic, uranium, and boron in Lemna gibba L. exposed to secondary effluents. Ecol Eng 35:1564–1567
Sela M, Tel-Or E, Fritz E, Huttermann A (1988) Localization and toxic effects of cadmium, copper, and uranium in Azolla. Plant Physiol 88:30–36
Smolders AJP, Roelofs JM (1996) The roles of internal iron hydroxide precipitation, sulphide toxicity and oxidizing ability in the survival of Stratiotes aboides roots at different iron concentrations in sediment pore water. New Phytol 133:253–260
Stiborová M, Doubravová M, Brezinová A, Friedrich A (1986) Effect of heavy metal ions on growth and biochemical characteristics of photosynthesis of barley (Hordeum vulgare L.). Photosynthetica 20:418–425
Sutcliffe JF (1962) Mineral salts absorption in plants. Pergamon Press, London
Tejeda S, Zarazúa G, Ávila-Pérez P, Carapia-Morales L, Martínez T (2010) Total reflection X-ray fluorescence spectrometric determination of elements in water hyacinth from the Lerma River. Spectrochim Acta Part B 65:483–488
Vaillant N, Monnet F, Sallanon H, Coudret A, Hitmi A (2004) Use of commercial plant species in a hydroponic system to treat domestic wastewaters. J Environ Qual 33(2):695–702
Vesk PA, Nockolds CE, Allaway WG (1999) Metal localization in water hyacinth roots from an urban wetland. Plant Cell Environ 22:149–158
Wang Q, Cui Y, Dong Y (2002) Phytoremediation of polluted waters: potentials and prospects of wetland plants. Acta Biotechnol 22(1–2):199–208
Weiss JD, Hondzo M, Semmens M (2006) Storm water detention ponds: modeling heavy metal removal by plant species and sediments. J Environ Eng 132(9):1034–1042
Win DT, Than MM, Tun S (2002) Iron removal from industrial waters by water hyacinth. AU J Technol 6(2):55–60
Xiong Z-T, Liu C, Geng B (2006) Phytotoxic effects of copper on nitrogen metabolism and plant growth in Brassica pekinensis Rupr. Ecotoxicol Environ Saf 64:273–280
Ye ZH, Baker AJM, Wong MH, Willis AJ (1997) Copper and nickel uptake, accumulation and tolerance in Typha latifolia with and without iron plaque on the root surface. New Phytol 136:481–488
Yruela I (2005) Copper in plants. Braz Plant Physiol 17(1):145–156
Zhu YL, Zayed AM, Qian J-H, de Souza M, Terry M (1999) Phytoaccumulation of trace elements by wetland plants: II. Water hyacinth. J Environ Qual 28:339–344
Acknowledgments
We would like to thank, and we are greatly indebted by, Dr. Sashnee Raja for her continuous advice and for facilitating logistical support. We would like also to thank Azmera Mebrahtu and Lutendo Mugwedi for their help during data collection. This study was supported by AngloGold Ashanti Ltd., S.A. Region, the Department of Trade and Industry (DTI), the National Research Foundation (NRF) of South Africa (THRIP fund awarded to I.M. Weiersbye and E.T.F. Witkowski), and the Water Research Commission (WRC-Pretoria: fund awarded to M.J. Byrne). Last but not least, we would like to thank the Water Research Commission (WRC-Pretoria) and AngloGold Ashanti Ltd. for collectively funding this study.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible editor: Elena Maestri
Electronic supplementary material
ESM 1
(DOCX 38 kb)
Rights and permissions
About this article

Cite this article
Newete, S.W., Erasmus, B.F., Weiersbye, I.M. et al. Sequestration of precious and pollutant metals in biomass of cultured water hyacinth (Eichhornia crassipes). Environ Sci Pollut Res 23, 20805–20818 (2016). https://doi.org/10.1007/s11356-016-7292-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-016-7292-y