
Abstract
Many reports on the effects of conventional salvage logging—the removal of fallen and damaged trees after a catastrophic windthrow—on subsequent forest restoration have focused on short-term results occurring over less than 20 years; however, this time scale is inadequate, especially for boreal forests, because of the time required for tree growth. Here, we examine the long-term effects of salvage logging after a catastrophic windthrow event in 1954 on the resilience of a boreal forest by assessing the continuous recruitment of coniferous trees, dominance of typical coniferous tree species, and potential for future recruitment. We targeted two regions with different proportions of coniferous trees that were subject to three disturbance and management histories: windthrow (WT: fallen trees left intact), windthrow and salvage (WT+SL: salvage logged after the windthrow), and old growth (OG: not affected by the windthrow). In both regions, past salvaging has had serious negative impacts on the continuous recruitment of coniferous trees and potential for future recruitment. Negative impacts on the dominance of typical coniferous tree species were only observed in mixed forests. Our results suggest that in comparison to the coniferous forest, the mixed forest was less resilient, i.e.; the capability of a forest to maintain its identity as assessed by the dominance and recruitment of typical conifer species after wind disturbance and salvage logging. We found that salvage logging could affect forest structure, even 60 years later, by destroying advanced growth, including potential mother trees, and nursery beds for seedlings of typical conifer tree species.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Boreal forest is a circumpolar biome of cold-tolerant coniferous and deciduous tree species that covers over 14.7 million km2 or 11% of the terrestrial surface of the Earth (Bonan and Shugart 1989). The major genera of conifers include Picea, Abies, Larix, and Pinus, depending on the soil moisture and the specific region. Windthrow is a common natural disturbance in boreal forests in Northern Europe, North America, and East Asia (Schelhaas et al. 2003; Kamimura and Shiraishi 2007; D’Amato et al. 2017), and the threat of such events is increasing as a result of climate change (IPCC Working Group I Technical Support Unit 2013). Salvage logging, that is, the removal of dead or damaged trees to compensate for economic value that would otherwise be lost due to such events, has been the typical response following catastrophic windthrow events in regions where timber production is a priority (see Table 1.2 in Lindenmayer et al. 2008). However, evaluations of forest resilience, i.e., the ability of a forest to maintain its identity (Cumming et al. 2005) in the face of wind disturbance and salvage logging, based on long-term observations are especially necessary for boreal forests because of the dominance of slow-growing conifer species.
Many studies have investigated the effects of salvage logging on forest restoration and biodiversity worldwide (Royo et al. 2016; Thorn et al. 2018) based on field measurements covering a few years to two decades after salvage operations following windthrow events. Pioneer tree species dominate within a few years after the completion of salvage logging (Elliott et al. 2002; Rumbaitis del Rio 2006; Ilisson et al. 2007), whereas the dominance of these species decreases by approximately eight years after salvage (Royo et al. 2016), with mid- and late-successional tree species dominating 10–20 years later (Bottero et al. 2013; Kramer et al. 2014). However, no studies have reported complete forest recovery to the original species composition within 25 years (Lang et al. 2009).
Indeed, there is a considerable lack of studies of the long-term (over 25 years) effects of salvage logging on forest structure and species composition (Lang et al. 2009). A rare case study by Mabry and Korsgren (1998) found that the species composition of a temperate mixed forest recovered within 53 years after salvage following a windthrow event, but no comparable study has been conducted on boreal forests, which have a limited growing season (Bonan and Shugart 1989). The long-term effects (60 years) of windthrow on the boreal forest in Japan have been investigated by Kosugi et al. (2016), who suggest that the successional composition of tree species during the recovery process depends on the pre-disturbance stand conditions. In addition, they found that the forests were still in the stem-exclusion stage, during which younger stems cannot establish due to light limitation from crown closure, and thus, species that vary in terms of light tolerance and growth rate compete against each other (Oliver 1980-1981).
In 1954, Typhoon No. 15, named Marie, caused tremendous damage, especially in northern Japan where it impacted 651,844 ha of forests (Scientific investigation group of the wind-damaged forests in Hokkaido 1959). Most fallen trees were salvage logged over many years following the catastrophic windthrow; however, some reserves were left, including fallen trees in several regions for scientific investigation. In this study, we examined the long-term effects of salvage logging after this catastrophic windthrow on the resilience of boreal forests using permanent research plots in reserves. We targeted two regions with different proportions of conifer species to analyze resilience from the perspective of the continuous recruitment of coniferous trees, the dominance of typical coniferous tree species, and the potential for their future recruitment.
Methods
Typhoon event
In 1954, the violent storm Typhoon No. 15 (Marie, Toyamaru) caused serious damage, especially in Hokkaido in northern Japan. The volume of fallen trees and the windthrow area in Hokkaido were 21.1 × 106 m3 and 651,844 ha, respectively (Scientific investigation group of the wind-damaged forests in Hokkaido 1959). In most of the damaged forests, salvage logging was conducted for several years using chain saws, extraction with horses and skidders, and transportation of the logs in trucks (Editorial Board for the 50-year Anniversary of Forest Restoration from the Toyamaru Typhoon 2005). After Typhoon No. 15, another major wind disturbance, Typhoon No. 18 (Songda), occurred in 2004, disturbing 36,956 ha of forested area in Hokkaido (Hokkaido Branch of Forestry and Forest Products Research Institute 2004). Of the total windthrow area, 30% was concentrated in two adjacent cities: Chitose City and Tomakomai City (Tsushima and Saitoh 2003).
Study area
Boreal forests in Hokkaido are generally dominated by A. sachalinensis (F.Schmidt) Mast. and P. jezoensis Carr. and characterized by a dense undergrowth of dwarf bamboo (Sasa spp.) (Kira 1991). The dominance of these conifer species and other broad-leaved deciduous species largely depends on elevation. In lowland Hokkaido, broad-leaved species such as Quercus crispula Blume, Tilia, Acer and Ulmus are distributed with A. sachalinensis (Kira 1991); A. sachalinensis and P. jezoensis become dominant at higher elevations or in areas where the warmth index (WI) < 5 (Kira 1991). Therefore, variation in the dominance of conifer species is a characteristic feature of boreal forests in Hokkaido (Ishizuka and Sugawara 1986).
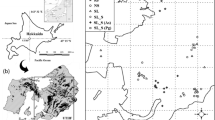
We targeted two regions in Hokkaido, Japan: Shikotsu (42.7N 141.3E, elevation 270–330 m, with an annual mean temperature of 6.7 °C and annual precipitation of 1766 mm at the Shikotsukohan meteorological station located 6 km from the reserve area) and Tokachi-Taisetsu (43.5N 143.1E, elevation 690–987 m, with an annual mean temperature of 3.7 °C, annual precipitation of 1315 mm, and annual snowfall of 103 cm at the Nukabira meteorological station located 19 km from the reserve area) (Fig. 1).
Shikotsu is a boreal mixed forest dominated by P. jezoensis, A. sachalinensis, Tilia japonica (Miq.) Simonk., and Acer amoenum Carrière var. matsumurae (Koidz.) K. Ogata, and the soil is volcanic in origin and immature. This region has been conserved as a national park since 1949, prior to which it had been protected as the emperor’s forest since 1889 and has, thus, been protected from cutting for more than 60 years. However, the typhoon in 1954 removed 85% of the standing trees (diameter at breast height [DBH] ≥ 5 cm) and 94% of the basal area (BA), representing a stand-replacing event (Kosugi et al. 2016), and except for those in the reserve area, most of the damaged trees were salvaged.
Tokachi-Taisetsu is a boreal coniferous forest dominated by P. jezoensis, A. sachalinensis, and Picea glehnii Masters, and the soil is volcanic in origin and immature. This region was a primary forest until the 1954 typhoon, after which the damage in terms of volume was estimated at 80–89% (Ishibashi et al. 2018), representing a stand-replacing event. The fallen trees in the Tokachi-Taisetsu region accounted for 695,000 m3, and except in the reserve area, more than 80% of them were logged in the subsequent 5 years, which led to economic activation of the region (Obihiro-City et al. 1987).
Quadrat setting
In both regions, a large quadrat of 0.5 ha (50 × 100 m) was established in each reserve area in 1956, after which no salvage logging was performed. The species name was recorded for both standing and fallen trees with DBH ≥ 5 cm, and the DBH of standing trees was measured with an accuracy of within 2 cm (Kosugi et al. 2016; Ishibashi et al. 2018). These data were published, but the quadrats remained for further investigation. Thus, we decided to take full advantage and defined them as “windthrow” (WT: fallen trees left intact).
To select sites that have not experienced disturbances other than Typhoon No. 15 (Marie, Toyamaru) and subsequent salvage logging and sites that have not experienced any major typhoons and any forest management, we reviewed aerial photographs taken before (1944–1948) and after the typhoon (in 1955 and 1962) in the two regions along with management histories by the Forestry Agency of the Japanese Government and a map of the forest gaps created by the typhoon (Tamate et al. 1977). We were able to avoid selecting stands affected by Typhoon No. 18 in 2004 (Songda) and by any artificial disturbances, such as cutting and thinning, as based on the recorded management history. We carefully selected several candidate sites for the “windthrow and salvage” (WT+SL: salvage logged after the windthrow of 1954) and “old growth” (OG: not affected by Typhoon No. 15 in 1954 and Typhoon No. 18 in 2004) categories, so that they would be representative of the regions.
To narrow down the sites for investigation, we visited several candidates in June 2017 with foresters of the Hokkaido Forest Bureau and confirmed the height and DBH of the dominant species, tree density, state of decay of fallen trees, and accessibility. To avoid bias due to the physical environment, including the slope angle and elevation among quadrats, we selected stands with a gentle slope ( < 10°) and minimal difference in elevation ( < 300 m). Regarding the Shikotsu region, we targeted the area with sparse Sasa species in the southeastern part of lake Shikotsu (Hokkaido Branch of Forestry and Forest Products Research Institute 1983) (Supplementary Material 1).
In the two regions, we defined three disturbance histories, i.e., WT, WT+SL, and OG (Fig. 1), and each included a 0.5-ha (50 × 100 m) quadrat. The WT quadrat was established in the reserve area in 1956, two years after the windthrow. The WT+SL and OG quadrats were established in 2017 as near to the WT quadrat as possible, with the goal that the site conditions be consistent with those in the WT. The OG quadrat that was established inside the study sites for long-term monitoring was not affected by Typhoon No. 15 (Nishimura et al. 2009). The distances between the WT+SL and WT quadrats were 5.4 km in Shikotsu and 2 km in Tokachi-Taisetsu. As it was difficult to locate the OG quadrats near the WT quadrats because the surrounding areas had been severely damaged by the windthrow and salvaging, the distances between the WT and OG quadrats were 7.5 km in Shikotsu and 15 km in Tokachi-Taisetsu.

Locations of the study area and quadrats
Field measurement and analysis
In the WT, WT+SL and OG quadrats, trees with a DBH ≥ 5 cm were identified (without shrub and vine species), and DBH was measured with an accuracy of within 0.1 cm in September 2017 (Supplementary Material 1). Tree height was selectively measured for at least eight (if present) A. sachalinensis and P. jezoensis individuals in each quadrat. The diameter and decay class of the fallen logs (> 7.6 cm) of both of conifer and broad-leaved tree species crossing the four lines in a quadrat (Fig. 1) were measured. Dead fallen logs were classified into five decay classes from I (least decayed) to V (most decayed) based on Graham and Cromack (1982). Wood in decay classes IV, with well-decayed sapwood (partly absent) and partly decayed heartwood, and V, deeply decayed wood that can be easily broken by hand, are suitable for seedling establishment (Table 1).
The distribution of DBH at 5-cm intervals was drawn for conifer and broad-leaved tree species. We regarded the trees composing the canopy layer of stands as potential mother trees that can disperse seeds, and the potential mother trees of the typical conifer species A. sachalinensis and P. jezoensis were judged based on their DBH in each WT, WT+SL, and OG quadrat. The size of trees that reached the canopy layer was estimated from the relationship between DBH and height (H = aDb; H: height, D: DBH, a and b: constants). Individual A. sachalinensis and P. jezoensis larger than D ≥ 15 cm in the WT, D ≥ 20 cm in the WT+SL, and D ≥ 30 cm in the OG were assumed to be mother trees. The sum of the BA of mother trees was calculated for the three types of disturbance and management history in each region.
Species composition (i.e., the BA of each species) in the three types of disturbance and management history in each region was compared using global non-metric multidimensional scaling (nMDS) with the program PC-ORD version 7 (McCune and Mefford 2016). The Bray–Curtis distance was applied to measure the distance between quadrats. The number of runs with real data was 250, with a maximum number of iterations of 500 and 250 randomized runs. The nMDS runs were repeated 50 times with random starting configurations in one–six dimensions, and the solution with the lowest amount of stress (< 0.001 of the final stress) was chosen. In all ordinations, one-dimensional solutions proved to be the best, and the coefficients of determination were 0.943 in Shikotsu and 0.999 in Taisetsu. The dried weight of downed logs (W kg m−2) in decay classes IV and V were estimated using Eq. (1), where D (cm) is the diameter of dead wood, Lns (m) is the horizontal length of the north–south line, Lew (m) is the horizontal length of the east–west line (Fig. 1), and M (g cm−3) is the wood density of each species category and decay class (Ugawa et al. 2012).
We define resilience in this study as the ability of a boreal forest to maintain its identity in the face of wind disturbances and salvage logging, according to Cumming et al. (2005). We determined the identity of the target boreal forests as the “continuous recruitment of typical conifer species”, “dominance of typical coniferous tree species”, and “potential for future recruitment of typical conifer species”. This study focuses on conifer tree species because dominance variation is a characteristic feature of boreal forests in Hokkaido, and such variations depend on stand elevation. We consider the boreal forest to be resilient if (1) the DBH distribution expresses sustainable recruitment of A. sachalinensis and P. jezoensis (2) the tree species ordination expresses the dominance of A. sachalinensis and P. jezoensis, and (3) potential mother trees and potential nursery beds exist to support future recruitment of A. sachalinensis and P. jezoensis in the WT (63 years after windthrow), WT+SL (63 years after salvage logging followed by windthrow), and OG (no disturbance for decades) quadrats.
Results
DBH histogram
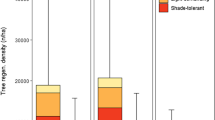
In the boreal mixed forest, the number of conifers was much lower in WT+SL than in WT and OG (Fig. 2a). In WT, both conifer and broad-leaved trees exhibited a reverse J-shaped distribution, but only broad-leaved trees showed that distribution in WT+SL and OG. Recruitment of conifers was constrained in WT+SL and OG in the boreal mixed forest. Potential mother trees of conifer species did not exist in WT+SL. The total BA of potential A. sachalinensis and P. jezoensis mother trees was higher in the OG than in WT.

Histogram of the coniferous and broad-leaved trees for each disturbance and management history in a boreal mixed forest and b boreal coniferous forest. Arrows indicate the DBH of potential A. sachalinensis and P. jezoensis mother trees. The sum of the BA of potential mother trees is shown under the arrows. No arrows mean no potential mother trees
In contrast to the mixed forest, in the boreal coniferous forest, conifer tree species survived at in much lower numbers in WT+SL than in WT (Fig. 2b). In addition, coniferous trees in WT+SL were most abundant in the 15-cm ≤ D < 20-cm size range, showing poorer recruitment in recent years. In contrast, the histogram of conifers in WT and OG mostly exhibited a reverse J-shaped distribution, although the number of coniferous trees was much lower in the OG quadrat than the WT quadrat. Recruitment of conifers in WT+SL was slightly constrained in the boreal mixed forest, and potential mother trees of A. sachalinensis and P. jezoensis were mostly equally distributed in WT+SL, WT and OG.
Species composition
In the boreal mixed forest, no typical conifer species characterized WT+SL, WT, or OG (Fig. 3a). All species that had a strong relation to disturbance and management histories were broad-leaved species, except T. cuspitada (TC), which characterized OG and WT+SL. In the boreal coniferous forest, P. jezoensis (PJ) was placed in the center of the ordination among the WT, WT+SL, and OG quadrats, and it was especially strongly related to OG (Fig. 3b). Conversely, A. sachalinensis was not related to any disturbance or management history. The WT and WT+SL quadrats were primarily characterized by broad-leaved species.

Ordination biplot by nMDS for a boreal mixed forest and b boreal coniferous forest. The vertical scale indicates the score on the first axis. Species with Pearson’s r-squared correlations with the first ordination axis > 0.6 are placed in ascending order by their score. Species names are shown using the abbreviation of their scientific names. Conifer tree species: PG, Picea glehnii Masters; PJ, Picea jezoensis Carr.; TC, Taxus cuspidata Siebold et Zucc., Broad-leaved tree species: AH, Alnus hirsuta (Spach) Turcz. ex Rupr. var. hirsute; AM, Acer pictum Thunb. subsp. mayrii (Schwer.) H. Ohashi; AP, Acer pictum Thunb. subsp. mono (Maxim.) H. Ohashi; BE, Betula ermanii Cham.; CC, Cornus controversa Hemsl. ex Prain; CeS, Cerasus sargentii (Rehder) H. Ohba; CJ, Cercidiphyllum japonicum Siebold et Zucc. ex Hoffm. et Schult.; CM, Cerasus maximowiczii (Rupr.) Kom.; CS, Chengiopanax sciadophylloides (Franch. et Sav.) C. B. Shang et J. Y. Huang; FM, Fraxinus mandshurica Rupr.; KS, Kalopanax septemlobus (Thunb.) Koidz.; MO, Magnolia obovata Thunb.; PS, Padus ssiori (F. Schmidt) C. K. Schneid.; QC, Quercus crispula Blume; SA, Sorbus alnifolia C. Koch; SoC, Sorbus commixta Hedl.; UD, Ulmus davidiana Planch. var. japonica (Rehder) Nakai. Species names of conifer trees are shown in the box
Downed logs
The dried weight of downed logs in decay classes IV and V in WT+SL, WT, and OG in the boreal mixed forest (Table 1) was generally the same, exhibiting the same potential as a nursery for conifer seedlings. In the boreal coniferous forest, this potential was highest in OG followed by WT and then WT+SL (Table 1), which was half that of WT and one–fifth that of OG.
Discussion
In this study, we exerted every effort to utilize the valuable data and quadrats from 63 years ago. This dataset does not contain replications of forest types or disturbance and management history types and thus does not permit statistical analysis. However, due to our careful selection of the WT+SL and OG sites and the establishment of quadrats equal to those in WT, we can provide a careful discussion for understanding the resilience of boreal forests. Our results suggest that the mixed forest is less resilient than the coniferous forest in terms of our definition of resilience, i.e., the capability of a forest to maintain its identity as assessed by the dominance and recruitment of typical conifer species in the face of wind disturbance and salvage logging. We found that salvage logging could affect the forest structure, even 60 years later, by destroying advanced growth, including potential mother trees, and nursery beds for seedlings of typical conifer tree species.
Effect of salvage logging on boreal forests
Continuous recruitment of typical conifer species
Continuous recruitment of conifers was observed in the WT in both boreal mixed forest and boreal coniferous forest, though the total BA and frequency were much lower in the mixed forest (Fig. 2). The reason for the continuous recruitment of coniferous trees would be the survival of saplings and seedlings on the forest floor after wind disturbance. Advanced growth left undamaged has been shown to play a major role in the early stage of forest regeneration after wind disturbances (Morimoto et al. 2011; Royo et al. 2016), and seedlings and juveniles left undamaged on the forest floor at the time, i.e., 63 years since the windthrow, should have grown and become saplings and adult trees in the WT quadrat in both the mixed and coniferous forests. The reason for the lower total BA and coniferous tree frequency in the mixed forests is likely due to the lower number of coniferous saplings and adult trees left undamaged after the windthrow. In the boreal mixed forest of WT, only 10 coniferous trees (DBH ≥ 5 cm) survived (in the 0.5-ha quadrat), and this number decreased to 1 by 1959 (Appendix S1 in Kosugi et al. 2016). In contrast, 424 (/ha) coniferous trees (DBH ≥ 5 cm) survived in the boreal coniferous forest in WT in 1959, including potential mother trees (Ishibashi et al. 2018). These data suggest that more coniferous trees, including potential mother trees, survived in WT in the boreal coniferous forest, which was likely due to the higher original density of these trees than in WT in the boreal mixed forest.
Salvage logging should have completely damaged a small number of undamaged coniferous trees that survived the windthrow, resulting in stands with few coniferous trees in WT+SL in the boreal mixed forest. In the boreal coniferous forest in the WT+SL quadrat, however, regeneration of coniferous trees might have been possible due to advanced growth of trees that survived both the windthrow and salvaging.
Discontinuous distribution of conifers in OG in the boreal mixed forest and the lower BA and frequency of coniferous trees in OG than in WT and WT+SL in the boreal coniferous forest indicate that a prolonged closed canopy inhibits seedling development. These results support the theory that natural disturbance is fundamental to the structure and function of forest ecosystems (Attiwill 1994).
Dominance of typical coniferous tree species
Boreal coniferous forest exhibiting sustainable recruitment (Fig. 2b) was characterized by P. jezoensis in all disturbance and management histories, i.e., WT+SL, WT, and OG (Fig. 3b), but no typical conifer species characterized the boreal mixed forest (Fig. 3a). Instead, broad-leaved tree species characterized the boreal and mixed forests in the WT and WT+SL quadrats.
Pits are holes that can form after trees are uprooted, which occurred in both the WT and WT+SL quadrats. In addition to these areas, the area scarred by salvage logging emerged in WT+SL. The mineral soil that appeared in both the pits and salvaged area would have provided good opportunities for the germination and establishment of small-seeded pioneer tree species (Peterson and Pickett 2000; Ulanova 2000) such as Alnus hirsuta (AH) and Betula ermanii (BE) in the initial recovery stage, followed by shade-tolerant broad-leaved species (Kramer et al. 2014; Royo et al. 2016) such as Acer pictum Thunb. subsp. mono (AP), A. pictum Thunb. subsp. Mayrii (AM), Cerasus maximowiczii (CM), and Cerasus sargentii (CeS). However, both WT+SL and WT were markedly different from OG, consistent with a study in a Tsuga canadensis-Pinus strobus forest 78 years after a windthrow event (Sass et al. 2018) that showed a strong shift in species composition compared to before a hurricane characterized by a tremendous decrease in conifer species (P. strobus and T. canadensis) and an increase in broad-leaved species (Betula and Acer spp.), regardless of the application of salvage logging.
Potential for future recruitment of typical conifer species
Interestingly, the amount of potential nursery habitat in boreal mixed forest was the same in WT+SL as in WT and OG, yet that in WT+SL was limited compared to that in WT and OG in the boreal coniferous forest (Table 1). The more moderate climate in the boreal mixed forest might have rapidly decomposed the dead wood supplied 63 years ago in WT to reach a stable equilibrium.
The WT+SL history in boreal mixed forest had no potential mother trees, which should be the major factor limiting the future recruitment of A. sachalinensis and P. jezoensis. In contrast, there is no serious obstacle to the future recruitment of coniferous trees in boreal coniferous forest because of the existence of potential mother trees and nursery beds for all types of disturbance and management history.
In the boreal mixed forest, even in WT, which contained potential mother trees and potential nursery beds, decades will be required for coniferous trees to become dominant due to the slow process of conifer regeneration, which is constrained by growth–limiting climate factors (Noble and Alexander 1977; Antos et al. 2000).
What determines the resilience of boreal forests?
Conifer species in a boreal forest, especially at the initial stage of establishment, have strict microenvironmental requirements (Harvey et al. 1987; Sugita and Nagaike 2005; Iijima and Shibuya 2010). Downed logs can provide a suitable nursery because they support the survival of conifer seedlings by protecting them from pathogens (Wall 1984) as well as from competition with fast-growing species such as broad-leaved trees and herbaceous species (Bottero et al. 2013). Sasa also becomes the major obstacle to the establishment of these species in the subfrigid zone in East Asia (Narukawa and Yamamoto 2002), where downed logs are a crucial element for the recruitment of canopy tree seedlings. Decades are required for fallen logs to decay and become suitable seedbeds (Zielonka and White 2006), and this nursery role can only be fulfilled when potential mother trees survive windthrow and salvaging. Hence, the primary propagule source for restoration immediately after a windthrow is vegetation that survived on the forest floor rather than widely dispersed seeds (Roberts 2004). Therefore, the intensity of salvaging influences its effect on the recovery of biomass and species composition. For example, within 2 years after moderate-intensity salvage logging, vegetation was observed to return to its original condition (Peterson and Leach 2008), though the effect of severe salvaging that caused a 41–82% decline in sapling abundance was extended to 5 years (Royo et al. 2016). This observation means that the amount of surviving vegetation determines the period necessary for the recovery of forests.
When a boreal forest is salvaged after a windthrow event, it loses two important features that contribute to resilience: the advanced growth of typical conifer species, including potential mother trees, and future nursery beds that support the establishment of conifer seedlings. Thus, post-disturbance salvage logging can greatly reduce the resilience of a forest, especially when no potential mother trees remain. Forests dominated by conifer species require more than 200 years after a stand-replacing disturbance to survive to the old-growth stage (Angelstam and Kuuluvainen 2004). Salvaging can extend the duration of succession, and unexpectedly, old-growth species composition might be completely different from that before a disturbance.
Long-term observations in a boreal forest revealed that the negative effects of salvage operations on the ability of the forest to maintain its identity in the face of wind disturbance and salvage logging can persist for over 60 years, suggesting that in contrast to temperate forests, boreal forests require more time to restore their species compositions (Mabry and Korsgren 1998) because the major tree species in the boreal forests are coniferous and require specific conditions and a sustained period of time to establish. Additional long-term data and case studies are required to validate our conclusions, which are based on limited replication.
References
Angelstam P, Kuuluvainen T (2004) Boreal forest disturbance regimes, successional dynamics and landscape structures: a European perspective. Ecol Bull :117–136
Antos JA, Parish R, Conley K (2000) Age structure and growth of the tree-seedling bank in subalpine spruce-fir forests of south-central British Columbia. Am Midl Nat 143:342–354
Attiwill PM (1994) The disturbance of forest ecosystems: the ecological basis for conservative management. For Ecol Manag 63:247–300
Bonan GB, Shugart HH (1989) Environmental factors and ecological processes in boreal forests. Ann Rev Ecol Syst 20:1–28
Bottero A, Garbarino M, Long JN, Motta R (2013) The interacting ecological effects of large-scale disturbances and salvage logging on montane spruce forest regeneration in the western European Alps. For Ecol Manag 292:19–28
Cumming GS, Barnes G, Perz S, Schmink M, Sieving KE, Southworth J, Binford M, Holt RD, Stickler C, Van HT (2005) An exploratory framework for the empirical measurement of resilience. Ecosystems 8:975–987
D'Amato AW, Orwig DA, Foster DR, Barker PA, Schoonmaker PK, Wagner MR (2017) Long-term structural and biomass dynamics of virgin Tsuga canadensis–Pinus strobus forests after hurricane disturbance. Ecology 98:721–733
Editorial Board for the 50-Year Anniversary of Forest Restoration From Toyamaru Typhoon 2005 (2005) Entrusting the revived forest to the future; 50 years after the hit of Toyamaru typhoon. iWord. Inc, Sapporo (in Japanese)
Elliott KJ, Hitchcock SL, Krueger L (2002) Vegetation response to large scale disturbance in a southern Appalachian forest: Hurricane Opal and salvage logging. J Torrey Bot Soc 129:48–59
Graham RL, Cromack K Jr (1982) Mass, nutrient, and decay rate of dead boles in rain forest of Olympic National Park. Can J For Res 12:511–521
Harvey AE, Jurgensen MF, Larsen MJ, Graham RT (1987) Relationships among soil microsite, ectomycorrhizae, and natural conifer regeneration of old-growth forests in western Montana. Can J For Res 17:58–62
Hokkaido Branch of Forestry and Forest Products Research Institute (1983) Distribution map of Sasa group in Hokkaido. Hokkaido Branch of Forestry and Forest Products Research Institute, Bibai (in Japanese)
Hokkaido Branch of Forestry and Forest Products Research Institute (2004) Preliminary report on damage caused by Typhoon No.18 (Summary). Hokkaido branch of forestry and forest products research institute, Bibai (in Japanese)
Iijima H, Shibuya M (2010) Evaluation of suitable conditions for natural regeneration of Picea jezoensis on fallen logs. J For Res 15:46–54
Ilisson T, Köster K, Vodde F, Jõgiste K (2007) Regeneration development 4–5 years after a storm in Norway spruce dominated forests, Estonia. For Ecol Manag 250:17–24
IPCC Working Group I Technical Support Unit (2013) Climate change 2013 the physical science basis working group I contribution to the fifth assessment report of the intergovernmental panel on climate change
Ishibashi S, Furuya N, Sasaki S, Takahashi M (2018) Sixty-year stand dynamics of a forest stand damaged by Typhoon Toyamaru in Higashitaisetsu natural forest, central Hokkaido, Japan. Bull FFPRI 17:83–88 (in Japanese)
Ishizuka M, Sugawara S (1986) Composition and structure of natural mixed forests in central Hokkaido (I) composition differences and species characteristics by elevation and from disturbances. J Jpn For Soc 68:79–86
Kamimura K, Shiraishi N (2007) A review of strategies for wind damage assessment in Japanese forests. J For Res 12:162–176
Kira T (1991) Forest ecosystems of east and southeast Asia in a global perspective. Ecol Res 6:185–200
Kosugi R, Shibuya M, Ishibashi S (2016) Sixty-year post-windthrow study of stand dynamics in two natural forests differing in pre-disturbance composition. Ecosphere 7:e01571
Kramer K, Brang P, Bachofen H, Bugmann H, Wohlgemuth T (2014) Site factors are more important than salvage logging for tree regeneration after wind disturbance in Central European forests. For Ecol Manag 331:116–128
Lang KD, Schulte LA, Guntenspergen GR (2009) Windthrow and salvage logging in an old-growth hemlock-northern hardwoods forest. For Ecol Manag 259:56–64
Lindenmayer DB, Burton PJ, Franklin JF (2008) Salvage logging and its ecological consequences. Island Press, Washington
Mabry C, Korsgren T (1998) A permanent plot study of vegetation and vegetation-site factors fifty-three years following disturbance in central New England, U.S.A. Écoscience 5:232–240
McCune B, Mefford. MJ (2016) Pc-ord. Multivariate analysis of ecological data. Version 7.03. MjM Software, Gleneden Beach
Morimoto J, Morimoto M, Nakamura F (2011) Initial vegetation recovery following a blowdown of a conifer plantation in monsoonal East Asia: impacts of legacy retention, salvaging, site preparation, and weeding. For Ecol Manag 261(8):1353–1361
Narukawa Y, Yamamoto S (2002) Effects of dwarf bamboo (Sasa sp.) and forest floor microsites on conifer seedling recruitment in a subalpine forest. Japan. For Ecol Manag 163:61–70
Nishimura N, Kato K, Sumida A, Ono K, Tanouchi H, Iida S, Hoshino D, Yamamoto S, Hara T (2009) Effects of life history strategies and tree competition on species coexistence in a sub-boreal coniferous forest of Japan. Plant Ecol 206:29–40
Noble DL, Alexander RR (1977) Environmental factors affecting natural regeneration Engelmann spruce in the central Rocky Mountains. For Sci 23:420–429
Obihiro-City, Otofuke-Town, Kamishihoro-Town (1987) Northern line: 63 years of Shihoro-line. Toyo Co., Obihiro (in Japanese)
Oliver CD (1980/1981) Forest development in North America following major disturbances. For Ecol Manag 3: 153−168
Peterson CJ, Pickett STA (2000) Patch type influences on regeneration in a western Pennsylvania, USA, catastrophic windthrow. Oikos 90:489–500
Peterson CJ, Leach AD (2008) Salvage logging after windthrow alters microsite diversity, abundance and environment, but not vegetation. Forestry 81:361–376
Roberts MR (2004) Response of the herbaceous layer to natural disturbance in North American forests. Can J Bot 82:1273–1283
Royo AA, Peterson CJ, Stanovick JS, Carson WP (2016) Evaluating the ecological impacts of salvage logging: can natural and anthropogenic disturbances promote coexistence? Ecology 97:1566–1582
Rumbaitis del Rio CM (2006) Changes in understory composition following catastrophic windthrow and salvage logging in a subalpine forest ecosystem. Can J For Res 36:2943–2954
Sass EM, D'Amato AW, Foster DR (2018) Lasting legacies of historical clearcutting, wind, and salvage logging on old-growth Tsuga canadensis-Pinus strobus forests. For Ecol Manag 419–420:31–41
Schelhaas MJ, Nabuurs GJ, Schuck A (2003) Natural disturbances in the European forests in the 19th and 20th centuries. Glob Change Biol 9:1620–1633
Scientific Investigation Group of the Wind-damaged Forests in Hokkaido (1959) A report of the scientific investigations of the forests wind-damaged in 1954, Hokkaido. Japan, Japan Forest Technical Association
Sugita H, Nagaike T (2005) Microsites for seedling establishment of subalpine conifers in a forest with moss-type undergrowth on Mt. Fuji, central Honshu. Japan. Ecol Res 20:678–685
Tamate S, Kashiyma T, Sasanuma T, Takahashi K, Matsuoka H (1977) On the distribution maps of forest wind damage by Typhoon No.15, 1954 in Hokkaido. Bull Gov For Exp Stat 2:43–67
Thorn S, Bassler C, Brandl R, Burton PJ, Cahall R, Campbell JL, Castro J, Choi CY, Cobb T, Donato DC, Durska E, Fontaine JB, Gauthier S, Hebert C, Hothorn T, Hutto RL, Lee EJ, Leverkus AB, Lindenmayer DB, Obrist MK et al (2018) Impacts of salvage logging on biodiversity: a meta-analysis. J Appl Ecol 55:279–289
Tsushima T, Saitoh K (2003) Background to the windthrow by typhoon No.18. In: Research Group on Forest Disasters in Hokkaido (eds) Reports on the Analyses of the Windthrow by Remote Sensing
Ugawa S, Takahashi M, Morisada K, Takeuchi M, Matsuura Y, Yoshinaga S, Araki M, Tanaka N, Ikeda S, Miura S, Ishizuka S, Kobayashi M, Inagaki M, Imaya A, Nanko K, Hashimoto S, Aizawa S, Hirai K, Okamoto T, Mizoguchi T, Torii A, Sakai H, Ohnuki Y, Kaneko S (2012) Carbon stocks of dead wood, litter, and soil in the forest sector of Japan: general description of the national forest soil carbon inventory. Bull FFPRI 425:207–221
Ulanova NG (2000) The effects of windthrow on forests at different spatial scales: a review. For Ecol Manag 135:155–167
Wall RE (1984) Effects of recently incorporated organic amendments on damping-off of conifer seedlings. Plant Dis 68:59–60
Zielonka T, White PS (2006) When does dead wood turn into a substrate for spruce replacement? J Veg Sci 17:739–746
Acknowledgements
Funding this research was supported by a KAKENHI grant from the Japan Society for the Promotion of Science (Grant Number 17H01516); the Science and Technology Research Promotion Program for Agriculture, Forestry, Fisheries and Food Industry of the Environment Research and Technology Development Fund (S-15) of the Ministry of the Environment, Japan; and the Grant for Joint Research Program of the Institute of Low Temperature Science, Hokkaido University.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.


Rights and permissions
About this article

{kind=link}
{kind=link}
Cite this article
Morimoto, J., Umebayashi, T., Suzuki, S.N. et al. Long-term effects of salvage logging after a catastrophic wind disturbance on forest structure in northern Japan. Landscape Ecol Eng 15, 133–141 (2019). https://doi.org/10.1007/s11355-019-00375-w
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11355-019-00375-w