
Abstract
The raccoon Procyon lotor (Linnaeus 1758) is a native species of North and Central America, but it can be also found as an alien species in parts of Europe and Asia. The raccoon’s flexibility and capability in coping with environmental changes have made it the most successful invader compared with other species. In 1991, a raccoon was recorded in Iran for the first time. Since then, knowledge about the distribution, expansion range, and influences of this species on native ecosystem has remained unclear. The importance of environmental variables for the distribution of raccoons might vary widely across different spatial resolutions; therefore, we developed a robust statistical framework to predict the raccoon distribution across different spatial resolutions in Iran. Our predictions made by species distribution models (ENFA, GARP, and MaxEnt) highlighted the importance of food availability at a small spatial resolution and water resources, settlements, and population density at a large spatial resolution. The models indicate particular areas, mostly located in forests and rangelands near to the Caspian Sea, that can be a potential habitat for raccoons. We observed that the distribution of the raccoon population in Iran almost entirely overlaps with protected areas, which consequently brings about some conflicts with conservation practices and management plans. It is therefore imperative that the local knowledge be strengthened to adequately respond to the threats posed by raccoon invasions of special protected lands.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Alien species cause ecological, economic, and social strains. Knowledge about the invasion range of these species plays a vital role in understanding the ecology of invasive species and in creating a sustainable conservation and management plan. Predictive models, such as species distribution models (SDMs), have become an essential tool for researchers to predict and assess the distribution of alien species by quantifying species–environment relationships (Guisan and Thuiller 2005; Robertson et al. 2004). Different types of modeling techniques are used to fit different types of biological information recorded at each sample site. These techniques include (1) presence-only: where occurrences of the target species are recorded, and (2) presence/absence: when each sample site is carefully monitored to assert with sufficient certainty whether the species is present or absent. Absence data are either not available, unreliable (for most cryptic or rare species), or of limited use since certain sites may be suitable but not yet reached by the invading alien species (Hirzel et al. 2002). Thus, presence-only models are suitable tools for predicting the potential spread of invasive species. The presence-only methods search for characteristics of the species’ presence-point in order to extrapolate the remaining areas under study. Examples of these alternative techniques, often called profile or envelope methods, are bioclimatic prediction systems (BPS), support vector machines (SVM), ecological niche factor analyses (ENFA), genetic algorithm for rule-set production (GARP), and maximum entropy (MaxEnt) (Guisan et al. 2007). Choosing the wrong spatial resolution in the modeling process may cause an incorrect prediction of habitat patterns and failure to reproduce ecological processes (Tamis and Van’t Zelfde 1998; Naugle et al. 1999; Meyer 2007; Storch et al. 2007; Convertino et al. 2009; McGill 2010). The challenge is in defining a pattern of biodiversity at different spatial resolutions (Levin 1992; Storch et al. 2007; Convertino et al. 2009) and in detecting critical bioecological spatial resolutions (Levin 1992; Rahbek 2005; Storch et al. 2007; McGill 2010) or sets of commanding processes (Graves and Rahbek 2005). Moreover, there is always an upper threshold for spatial resolution above which the SDMs can no longer predict species distribution correctly (Hurlbert and Jetz 2007; Seo et al. 2009; Yamakita and Nakaoka 2009; Pineda and Lobo 2012).
The raccoon Procyon lotor (Linnaeus 1758) is a native species of North and Central America. Presently, the raccoon can be observed in some parts of Europe and Asia as well (Fig. 1). Over the first 60 years of the raccoon’s presence in Europe, the distribution range of this species was only restricted to Germany and France and the number of raccoons in these areas remained stable. The German raccoon has started to increase in number and distribution since the 1980s. Currently, an increase in the raccoon population is expected, given that the species is still colonizing new countries (Hohmann and Bartussek 2001; Michler et al. 2004; Farashi et al. 2013). The raccoon’s capability in coping with environmental changes has brought about a great invasion success. Raccoons usually prefer old deciduous forests close to water; however, they also settle in various types of habitats ranging from partly open and marshy grounds to urbanized areas (Sanderson 1987; Zeveloff 2002). The raccoon was introduced in Russia in 1936 for its valuable fur (Aliev and Sanderson 1966). Raccoon was recorded in Iran in 1991 for the first time near the Iran–Azerbaijan border, where it most likely had migrated from Azerbaijan. Although it has been a long time since the raccoon was seen for the first time in Iran, the current knowledge on the distribution, expansion range, and impacts of this species on natural environments remains extremely poor. Here, we propose a multiple spatial resolution approach to predict the expansion range of the raccoon in Iran. The objectives of our study are (1) to develop a robust statistical framework to predict the raccoon distribution in Iran, and (2) to investigate the performance of the prediction model in Iran across multiple spatial resolutions.

Methods
We used ENFA, GARP, and MaxEnt to predict the expansion range of the raccoon in Iran.
Analytical/statistical procedure
ENFA (Biomapper v4.0) transforms a number of correlated environmental variables into niche factors (Hutchinson 1957). The first extracted ENFA factor maximizes the absolute value of the marginality of the species, defined as the ecological distance between the species optimum and the mean available habitat. The higher the coefficient’s absolute value is, the farther the species distribution from the mean available habitat for that particular variable will be. The positive coefficients indicate a higher preference with high values, while the negative coefficients indicate a higher preference for the mean values. The remaining factors (i.e., the species specialization) are defined as the ratio of the ecological variance of the available habitat to that observed for the species. The higher positive or negative specialization values indicate that the species distribution is more narrowly focused with regard to the corresponding variable (Hirzel et al. 2002). The ENFA factors are also used to compute global marginality (M, indicating some degree of marginality when it is greater than 1), specialization (S, varying generally from 0 to ∞), and global tolerance that is the inverse of specialization (T, varying generally from 0 to 1) (Hirzel et al. 2002, 2004). In the present study, a certain number of factors are retained to produce the raccoon distribution map based on a comparison with MacArthur’s “broken-stick” distribution (Hirzel et al. 2002). The median, harmonic mean, and geometric mean algorithms were applied to calculate the raccoon distribution map.
GARP (Desktop GARP v1.1.6) is a non-parametric, machine-learning species distribution model, which is used to predict species’ potential geographical distribution based on a genetic algorithm (Stockwell and Noble1991; Stockwell and Peters 1999). Using an iterative process of rule selection, evaluation, testing, and incorporation or rejection, GARP looks for the non-random relationship between the environmental characteristics of the known species presence and the whole study area (Peterson et al. 2002). First, a random population is selected by combining initial prediction rules, which may show proper solutions for the problem. Each pixel is then evaluated for the degree of fit to the characteristics of the population in the search space. The rule will be kept for further runs of the algorithm if its performance is sufficiently based on the rule’s significance measure. This procedure continues until the maximum number of iterations is satisfied (Stockwell and Peters 1999).
MaxEnt is the other general-purpose machine-learning method for species distribution modeling (Phillips et al. 2006). MaxEnt estimates niches by finding the probability distribution of maximum entropy, given the constraint that feature values match their empirical average (Phillips et al. 2006). The importance of environmental variables in MaxEnt can be evaluated by jackknife tests (Elith et al. 2011). In order to evaluate the average behavior of the models, 10 random partitions were used in the three models (Phillips et al. 2006; Tittensor et al. 2009). Each partition was generated by randomly dividing the species occurrence into a calibration (75%) and validation (25%) data set. Model evaluation was conducted on the basis of three different criteria: (1) the true skill statistic (TSS) which corresponds to the sum of sensitivity and specificity minus one and is independent of prevalence (Lobo et al. 2008); (2) the sensitivity, ‘true positives’; and (3) specificity, ‘true negatives’ (Thuiller et al. 2009; Barbet-Massin et al. 2012).
Presence points
The presence points of the raccoon were collected from two sources, the Iranian Department of Environment and the regional inventories. In total, 21 observations and 40 signs of the raccoon have been recognized between 1991 and 2013. The presence points of the raccoon were scanned by searching for any sign of their presence, captured and photographed by cage trap and camera trap, respectively.
Environmental predictor variables
An extensive literature review was conducted to select the important variables which are involved in determining the distribution of the raccoon (Stuewer 1943; Sanderson 1987; Gehrt and Fritzell 1998; Wilson and Nielsen 2007; Newbury and Nelson 2007; Farashi et al. 2013). The selected environmental variables were land cover/land use characteristics, climatic, and topographic variables. These variables were extracted from three different spatial resolutions, i.e., 30, 100, and 1000 m (Table 1). Land cover/land use data were derived from 30-m Landsat Enhanced Thematic Mapper Plus (ETM+) imagery in 2010 [Iranian Forests, Range and Watershed Management Organization (IFRWO)]. The statistics of population density for 2011 were obtained from the Statistical Center of Iran. Topographic variables were obtained from a digital elevation model (DEM) originally produced by the National Cartographic Center of Iran (NCC) on 1:25,000 scale. The normalized difference vegetation index (NDVI) for 2011 was derived from the conterminous 30-m Landsat TM imagery. We obtained bioclimatic variables from the Ministry of Energy in Iran. The spatial analysis tool of ArcGIS 9.3 (ESRI 2009) was used to resample the data layers to different spatial resolutions (i.e., 30, 100, and 1000 m). The multicollinearity test was conducted using Pearson correlation coefficient (R) to examine the cross-correlation between variables. The variables with a cross-correlation coefficient value larger than ±0.8 were excluded from further analysis (see Table 1).
Results
Model evaluation
Table 1 presents the results of the remaining environmental variables after the multicollinearity test. In general, the invasion predictions made by ENFA, GARP, and MaxEnt for the raccoon were significantly better than those made by the random performance (Table 2). We found that MaxEnt outperformed GARP and ENFA models at the three spatial resolutions as evaluated by TSS, sensitivity, and specificity values (Table 2). Moreover, the evaluation results indicated that the 30-m spatial resolution performs better than other spatial resolutions in all three models.
Importance of environmental variables
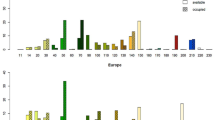
The results of a jackknife test of variable importance in MaxEnt are shown in Fig. 2. Accordingly, the relative importance of environmental variables in determining the raccoon distribution varies according to the spatial scale.

Importance of environmental variables for MaxEnt models (spatial resolution a 30 m, b 100 m, c 1000 m). Models with each variable omitted black bars, models containing each variable in isolation gray bars, models with all variables red bars, AUC area under curve (AUC is a measure of rank correlation). The AUC varies between 0 (worse model), 0.5 (random model) and 1 (best model) (color figure online)
Distance from settlements and population density in both urban and rural areas, and distance from roads, lakes, rivers, and rocky areas were the most important environmental variables affecting the distribution of the raccoon at 1000-m spatial resolution. On the other hand, the annual precipitation and the distance from forests, ranges, and rivers were the most important predictors in explaining the geographic distribution of the raccoon at 30-m and 100-m spatial resolution scales. The results of the ENFA are presented in Table 3. Three ENFA factors were retained after comparing with a broken-stick distribution of the invasion prediction. The first three retained factors accounted for 89, 86, and 88% of specialization variance of the raccoon distribution at 1000-m, 100-m, and 30-m spatial resolutions, respectively. Moreover, the first factor explained the 100% of marginality of the raccoon distribution at all three spatial resolutions. The first factor shows the importance of each environmental variable for the raccoon distribution (Table 3). ENFA and MaxEnt indicated the same importance for environmental variables.
Predicted potential invasion range
The raccoon was recorded at 31 sites in the study area (Fig. 3). The MaxEnt probability map of the raccoon occurrence at three different spatial resolutions is shown in Fig. 3.

Protected areas and potential distribution map of the raccoon in Iran made by MaxEnt at 30-m, 100-m, and 1000-m special resolutions
The results indicate that a large part of Iran can be a suitable habitat for the raccoon. The predicted suitable habitats for the raccoon were almost the same among all three models. However, the results of MaxEnt were generally more precise, compared to ENFA and GARP, with higher levels of predictive power and accuracy at all three spatial resolutions (Table 2). The MaxEnt analysis at 1000-m spatial resolution showed that areas with a high probability of invasion risk were mostly located in north, central, and west Iran. North, northwest, and west Iran were predicted to be the potential areas for invasion risk at 30-m and 100-m spatial resolutions. Furthermore, the MaxEnt results showed that 15.7, 10.8, and 11.1% of Iran can be considered as suitable habitats for the raccoon at 1000-m, 100-m, and 30-m spatial resolutions, respectively. On the basis of Fig. 3, the areas at risk of invasion by raccoon had 79, 76, and 79% overlap with protected areas at 1000-m, 100-m, and 30-m spatial resolutions, respectively.
Discussion
The MaxEnt model outperformed ENFA and GARP, which is consistent with the previous studies (Elith et al. 2011; Li et al. 2009; Phillips et al. 2006; Tittensor et al. 2009; Tong et al. 2013). In other words, MaxEnt seems to be a more suitable technique for predicting species distribution in comparison with ENFA and GARP.
In recent years, spatial resolution-dependent effects have received more attention in species distribution modeling (Pineda and Lobo 2012). Using the pixel aggregate method, Ferrier and Watson (1997) resampled 0.2-km grid cells to the coarse spatial resolution of 5 km, but they did not find any significant effect on the prediction performance of the presence-only model. However, Seo et al. (2009) found a decline in the model accuracy and spatial output agreement when increasing the grid size, as the accelerated decline generally happened between 8 and 16 times the initial grid. They also showed that the accuracy of the prediction was inversely related to the spatial resolution size, and decreased whenever the spatial resolution size increased in habitat analysis. There is no strict rule about which spatial resolution is appropriate for species distribution modeling. The choice of proper spatial resolution can be changed depending on the species being studied and the aim of the investigation (Boyce et al. 2003). As a result of the management goals we pursued and the existing little knowledge about the invasive situation in Iran, we proposed to carry out the study at different spatial resolutions. Moreover, all three distribution models, i.e., MaxEnt, ENFA, and GARP, showed the highest performance at the 30-m spatial resolution. Both global marginality and global specialization indicated that the raccoon had the same niche breadth at three different spatial resolutions (Table 3). As the raccoon occupies a narrow niche, it can be considered a specialized species in the occupied area. The relative importance of environmental variables in determining the raccoon distributions varies according to spatial resolution. Our findings illustrated that environmental variables including food availability and water resources at small spatial resolutions, as well as anthropogenic variables such as settlements and population density at large spatial resolutions, were important for the raccoon distribution. These results are in line with other studies that found positive relationships between the raccoon distribution and the closeness to water and food resources at small spatial resolutions (Stuewer 1943; Sanderson 1987; Gehrt and Fritzell 1998; Wilson and Nielsen 2007; Farashi et al. 2013). Many studies have been carried out on the habitat selection of the raccoon (Wilson and Nielsen 2007; Newbury and Nelson 2007; Farashi et al. 2013), while only a few investigated the raccoon distribution across multiple spatial resolutions (Farashi et al. 2013). The raccoon population was shown to spread in the forests and rangelands near the Caspian Sea and some parts of central and west Iran. It is important to emphasize that the raccoon distribution in Iran almost entirely overlapped the protected areas (Fig. 3). This may cause conflicts with the existing conservation and management plans because of the negative impacts of this species on indigenous biodiversity. More effort should be made in enhancing management or protection in these protected areas, since in the near future they may become important corridors for the expansion range of the raccoon in Iran.
This study is the first step towards a better understanding of the interactions between the raccoon distribution and environmental variables in Iran. The results of our study provide important tools to prioritize areas needing special action to control exotic species.
References
Aliev FF, Sanderson GC (1966) Distribution and status of the raccoon in Soviet Union. J Wildl Manage 30:497–502
Barbet-Massin M, Jiguet F, Albert CH, Thuiller W (2012) Selecting pseudo-absences for species distribution models: how, where and how many? Methods Ecol Evol 3:327–338
Boyce MS, Mao JS, Merrill EH, Fortin D, Turner MG, Fryxell J, Turchin P (2003) Scale and heterogeneity in habitat selection by elk in Yellowstone National Park. Ecoscience 10:421–432
Convertino M, Muneepeerakul R, Azaele A, Bertuzzo E, Rinaldo A, Rodriguez-Iturbe I (2009) On neutral meta community patterns of river basins at different scales of aggregation. Water Resour Res 45:W08424
Elith J, Phillips SJ, Hastie T, Dudík M, Chee YE, Yates CJ (2011) A statistical explanation of MaxEnt for ecologists. Divers Distrib 17:43–57
Farashi A, Kaboli M, Karami M (2013) Predicting range expansion of invasive raccoons in northern Iran using ENFA model at two different scales. Ecol Inform 15:96–102
Ferrier S, Watson G (1997) An evaluation of the effectiveness of environmental surrogates and modelling techniques in predicting the distribution of biological diversity. Environment Australia, Canberra
Gehrt SD, Fritzell EK (1998) Resource distribution, female home range dispersion and male spatial interactions: group structure in a solitary carnivore. Anim Behav 55:1211–1227
Graves GR, Rahbek C (2005) Source pool geometry and the assembly of continental avifaunas. Proc Natl Acad Sci USA 102(May):7871–7876
Guisan A, Thuiller W (2005) Predicting species distribution: offering more than simple habitat models. Ecol Lett 8:993–1009
Guisan A, Graham CH, Elith J, Huettmann F, NCEAS Working Group (2007) Sensitivity of predictive species distribution models to change in grain size. Divers Distrib 13:332–340
Hirzel AH, Hausser J, Chessel D, Perrin N (2002) Ecological-niche factor analysis: how to compute habitat-suitability maps without absence data? Ecology 83:2027–2036
Hirzel A, Posse B, Oggier PA, Crettenand Y, Glenz C, Arlettaz R (2004) Ecological requirements of reintroduced species and the implications for release policy: the case of the bearded vulture. Appl Ecol 41:1103–1116
Hohmann U, Bartussek I (2001) Der Waschbar. Oertel and sporer, Reutlin-gen
Hurlbert AH, Jetz W (2007) Species richness, hotspots, and the scale dependence of range maps in ecology and conservation. Proc Natl Acad Sci USA 104:13384–13389
Hutchinson GE (1957) Concluding remarks. Cold Spring Harb Symp Quant Biol 22:415–427
Ikeda T, Asano M, Matoba Y, Abi G (2004) Present status of invasive alien raccoon and its impact in Japan. Global Environ Res 8(2):125–131
Levin SA (1992) The problem of pattern and scale in ecology: the Robert H. MacArthur Award Lecture. Ecology 73:1943–1967
Li B, Ma J, Hu X, Liu H, Zhang R (2009) Potential geographical distributions of the fruit flies Ceratitis capitata, Ceratitis cosyra, and Ceratitis rosa in China. J Econ Entomol 102:1781–1790
Lobo JM, Jiménez-valverde A, Real R (2008) AUC: erratum: predicting species distribution: offering more than simple habitat models. Global Ecol Biogeogr 17:145–151
McGill BJ (2010) Matters of scale. Science 328:575–576
Meyer CB (2007) Does scale matter in predicting species distributions? Case study with the marbled murrelet. Ecol Appl 17(5):1474–1483
Michler FUF, Hohmann U, Stubbe M (2004) Investigation of home range, daytime resting site selection and social system of raccoons Procyon lotor in an urban habitat in Kassel. Beiträge zur Jagd- und Wildforchung, Bd 29:257–273 (In German with English summary)
Naugle DE, Higgins KF, Nusser SM, Johnson WC (1999) Scale-dependent habitat use in three species of prairie wetland birds. Landscape Ecol 14:267–276
Newbury RK, Nelson TA (2007) Habitat selection and movements of raccoons on a grassland reserve managed for imperiled birds. J Mammol 88:1082–1089
Peterson AT, Stockwell DRB, Kluza DA (2002) Distributional prediction based on ecological niche modeling of primary occurrence data. In: Scott JM, Heglund PJ, Morrison ML (eds) Predicting species occurrences: issues of scale and accuracy. Island, Washington, p 617–623
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecol Model 190:231–259
Pineda E, Lobo JM (2012) The performance of range maps and species distribution models representing the geographic variation of species richness at different resolutions. Global Ecol Biogeogr 21:935–944
Rahbek C (2005) The role of spatial scale and the perception of large-scale species richness patterns. Ecol Lett 8:224–239
Robertson MP, Villet MH, Palmer AR (2004) A fuzzy classification technique for predicting species’ distributions: applications using invasive alien plants and indigenous insects. Divers Distrib 10:461–474
Sanderson GC (1987) Raccoon. In: Noval M, Baker JA, Obbard ME, Malloch B (eds) Wild furbearer management and conservation in North America. Ontario Trappers Association, North Bay, pp 486–499
Seo C, Thorne JH, Hannah L, Thuiller W (2009) Scale effects in species distribution models: implications for conservation planning under climate change. Biol Lett 5(1):39–43
Stockwell DRB, Noble IR (1991) Induction of sets of rules from animal distribution data: a robust and informative method of analysis. Math Comput Simulat 33:385–390
Stockwell D, Peters D (1999) The GARP modelling system: problems and solutions to automated spatial prediction. Int J Geogr Inf Sci 13:143–158
Storch D, Marquet P, Brown J (2007) Scaling biodiversity. In: Storch D, Marquet P, Brown J (eds) Ecological reviews. Cambridge University, Cambridge
Stuewer FW (1943) Raccoons: their habits and management in Michigan. Ecol Monogr 13:203–257
Tamis WLM, Van’t Zelfde M (1998) An expert habitat suitability model for the disaggregation of bird survey data: bird counts in the Netherlands downscaled from atlas block to kilometer cell. Landscape Urban Plan 40(4):269–282
Thuiller W, Lafourcade B, Araújo M (2009) Mod operating manual for BIOMOD. Université Joseph Fourier, Laboratoire D’Ecologie Alpine, Grenoble
Tittensor DP, Baco AR, Brewin PE, Clark MR, Consalvey M, Hall-Spencer J, Rowden AA, Schlacher T, Stocks KI, Rogers AD (2009) Predicting global habitat suitability for stony corals on seamounts. J Biogeogr 36:1111–1128
Tong R, Purser A, Guinan J, Unnithan V (2013) Modeling the habitat suitability for deep-water gorgonian corals based on terrain variables. Ecol Inform 13:123–132
Wilson SE, Nielsen CK (2007) Habitat characteristics of raccoon daytime resting sites in southern Illinois. Am Midl Nat 157:175–186
Winter M (2009) Procyon lotor. In: AISIE (ed) Handbook of alien species in Europe, Springer, Dordrecht, p 368
Yamakita T, Nakaoka M (2009) Scale dependency in seagrass dynamics: how does the neighboring effect vary with grain of observation? Popul Ecol 51:33–40
Zeveloff SI (2002) Raccoons: a natural history. Smithsonian Institution, Washington
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Farashi, A., Naderi, M. Predicting invasion risk of raccoon Procyon lotor in Iran using environmental niche models. Landscape Ecol Eng 13, 229–236 (2017). https://doi.org/10.1007/s11355-016-0320-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11355-016-0320-8