
Abstract
Persoonia longifolia is a common mid-storey species that is difficult to return to post-mining environments. This study aimed to quantify in situ emergence of P. longifolia seeds on restored areas, investigate seed cueing prior to use in restoration and assess different tree guards for increasing seedling survival and health. Initial investigations found that <1 % of seeds buried or scattered on restored areas produced seedlings. However, if seeds were cued through burial in surrounding forest, retrieved and sown on restored areas, seedling emergence increased to 24 %. Significantly more seeds emerged as seedlings when buried (14.6 %) compared to those scattered on the soil surface (2.7 %). There was no significant difference in survival between seedlings planted at 2-3 weeks of age compared with those planted at 12 months of age after 20 months in situ growth. Additionally, those seedlings planted when younger were significantly taller (29.0 ± 2.9 cm) than those that were planted at 12 months of age (4.7 ± 0.3 cm). Use of “onion bag” guards improved survival from 58.1 ± 4.0 % (no guard) to 70.8 ± 3.4 % with an onion bag guard. The use of shade cloth guards did not significantly improve survival, however plant height did increase substantially after 32 months growth (22 cm compared with 7.2 cm for no guard). These data demonstrate that consideration needs to be given to specific species requirements to improve seedling emergence and survival when attempting to return difficult to germinate species to the post-mining environment.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Restoration of mine sites requires the return of a diverse range of species commonly found in the pre-mining environment in order to return it to a self-sustaining and resilient ecosystem (Ward et al. 1996; Norman et al. 2006; SERA Standards Reference Group 2016). Germination, growth and return of many of these indigenous species to restored areas can be problematic but is often a requirement of various government authorities in different parts of the world (Bielecka and Król-Korczak 2010). Consequently mining companies are now turning their attention to developing successful restoration techniques for returning plant communities rather than individual species to disturbed areas following mining, though different species may require contrasting reintroduction approaches (Koch 2007b; Todd et al. 2009).
The jarrah forest of southern Western Australia is part of a biodiversity hotspot of international significance (Myers et al. 2000) and is subjected to localised mining for the production of bauxite. Many species found within this area are difficult to germinate, propagate and return to restored areas of these mines and considerable effort is being made to understand the ecology of these species in order to return the forest to a self-sustaining ecosystem reflective of its original composition and function.
Persoonia longifolia, a small, mid-storey resprouter tree found in sandy or lateritic soils in the jarrah forest of southern Western Australia, has considerable economic importance for the mining, horticulture and floriculture industries. The distinctive size and shape make it desirable for the horticulture trade and its leaves are used as fillers in floral arrangements most of which is internationally exported to places such as Europe, Japan and the United States after wild collection (D. Growns pers. comm.). The relative abundance of P. longifolia in the pre-mining environment ranges from 185 to 440 plants/ha (Mullins et al. 2002; Norman and Koch 2008) (Stokes pers. comm.) making it an important species for insects (Ptyssoptera tryphera and a variety of bee species) and vertebrates such as kangaroos (Macropus fuliginosus), wallabies (Macropus spp.), cockatoos (Calyptorhynchus spp.) and emus (Dromaius novaehollandiae) (Stingemore and Krauss 2012; Chia et al. 2015). However, following mine site restoration, recorded plant densities are less than 10 % of the pre-mined levels (Mullins et al. 2002). The abundance and importance of this species in the pre-mining environment results in it being considered a priority species for return to restored areas following mining. Reliable propagation via vegetative means (cuttings) or through seed germination has proven to be exceptionally difficult until recently (Mullins et al. 2002; Norman and Koch 2008; Chia et al. 2016). Cuttings generally fail to produce roots while the seed is contained within a hard woody indehiscent endocarp similar to plums (Prunus domestica), dates (Phoenix dactylifera), and olives (Olea europaea), a germination structure found throughout different vegetation zones from the tropics to the boreal/subalpine and noted for their variable germination response (Baskin and Baskin 2014).
Investigations have been undertaken over the past 15 years to find a practical and effective means of returning significant numbers of P. longifolia to restored areas through topsoil return, inclusion in seed broadcasting mixes or as greenstock, with limited success to date (Mullins et al. 2002; Norman and Koch 2005, 2008). Complex seed dormancy is likely to be the principal reason that P. longifolia fails to germinate and establish under ex situ and in situ conditions (Chia et al. 2015). For problematic species such as P. longifolia burying seeds in situ (in undisturbed forest) to initially break dormancy, then retrieving and re-sowing under different conditions on restored post mining areas may provide a simple cost effective way to cue seeds for germination prior to direct seeding.
Persoonia longifolia, typically occurs in relatively shaded areas beneath large over-storey trees such as Eucalyptus marginata and Corymbia calophylla (Marchant et al. 1987). In contrast, the post- mining environment shortly after restoration works is extremely exposed and very different from the surrounding forest. However, competition is greatly reduced and therefore water, light and nutrient availability is higher allowing some species to rapidly establish. Predation by herbivores may be more of a problem in these largely denuded landscapes as animals actively target the limited vegetation that may be present (Koch et al. 2004; Stanton-Clements et al. 2013; Daws and Koch 2015; Nield et al. 2015). Thus the survival and growth of young P. longifolia plants could potentially be affected by the age of the restoration sites (microclimate effects), grazing from highly mobile herbivores and the age (size) of the seedling at the time of planting.
In order to protect seedlings against herbivory and environmental stresses, tree guards of various styles may be used (Lai and Wong 2005; Close et al. 2009; Stanton-Clements et al. 2013). Whilst plastic tree guards are suitable for use in temperate or tropical environs (Lai and Wong 2005), they have been found to have mixed results for plant establishment in Mediterranean type environments which regularly experience very high temperatures (40 °C+) which are greatly exacerbated by these closed plastic environments (Close et al. 2009; Stanton-Clements et al. 2013). Shade cloth and open weave tree guards by comparison may provide significant benefits by minimising extreme temperatures, reducing wind speeds, retaining soil moisture through reducing evaporation and transpiration as well as providing some level of protection against browsers leading to a decrease in mortality and improved plant growth and health (Close et al. 2009).
To improve restoration outcomes, we hypothesised that burial of fruits in restored areas would greatly improve germination and that it would be possible to cue seeds through soil burial to enhance in situ emergence without the requirement to germinate and maintain seedlings under nursery conditions prior to in situ planting. To test these hypotheses our aims were to: (1) quantify seedling emergence in scattered and buried fruits on restored areas and (2) cue seeds through a period of soil burial to break seed dormancy, then retrieve and re-sow onto restored sites.
In addition, the ability of P. longifolia seedlings to survive and grow once planted is currently unknown and in order to maximise survival and growth, we also hypothesised that seedlings planted immediately following germination would survive and grow better than seedlings held under nursery conditions over the summer months and that survival and growth would be enhanced through the use of tree guards.
Methods
Germination on restored areas of different ages
Persoonia longifolia fruit was collected from Dwellingup, Western Australia in August 2010 and sown onto two different restored areas at Alcoa’s Huntly bauxite mine site near Dwellingup several weeks later in late August. The exocarp and mesocarp were retained around the endocarp (and seed within) and the fruits were sown whole (i.e. not cleaned). Fruit numbers for this trial were limited due to the particularly poor seed set during 2010 and this is reflected in the experimental design.
The restored areas, into which the fruits were sown were located within newly restored mine pits. Bauxite is mined in shallow pits and restoration involves re-contouring, respreading topsoil before ripping to reduce compaction from mining, then spreading the seed mix. Four replicates of 20 fruits were sown onto a newly restored pit where earthworks and seed sowing had recently been completed (2010 restoration site) and an older 3 year old restored pit (restored 2007). Trees, shrubs and ground covers were already well established on the three year old restoration site at the time that the P. longifolia fruits were sown. Fruits were buried to a depth of approximately 2 cm inside wire cages (40 cm long × 50 cm wide × 30 cm tall; mesh size 2 cm) to protect germinating seedlings from herbivores. Twenty fruits were also scattered directly on the soil surface within the cages and 20 fruits were scattered on the soil surface adjacent to the cages to investigate the impacts of herbivory. The areas were examined on a monthly basis for the first year (from August 2010 to August 2011) to check for germination and emergence. After the first year, these areas were examined three times in 2012 (January, April and July). Access to this area then became difficult due to ongoing mining operations and consequently these areas were only re-examined one more time (after the summer period) in April 2013.
Synthetic seed bank trial
Fruit was collected in November 2010 from near Haddleton Nature Reserve, Western Australia. Fruits were used as whole dispersal units (i.e. not cleaned) and four replicates of 25 fruits were examined by x-ray analysis (MX-20 Digital X-ray, Faxitron, USA) to determine mean seed fill at the commencement of the experiment.
To determine whether different in situ environments and sowing approaches enhance or suppress seed cueing for germination, freshly collected fruits were initially sown in December 2010 at the Worsley Boddington Bauxite Mine site in Western Australia. Fruits were either shallow buried (2 cm deep) or scattered on the soil surface in natural unburnt jarrah forest or in a newly restored pit (2010 restoration site). Fruits that were scattered on the restored soil surface as part of this initial treatment were left in place for the entire experiment. These fruits acted as a control as this treatment is similar to methods employed in the past when including P. longifolia into native seed mixes used for restoration (i.e. uncleaned fruits scattered on the surface). Fruits were also buried and left in situ in both jarrah forest sites and restoration areas. Fruits that were to be retrieved at a later date were buried in mesh bags and wire cages were placed over the bags to prevent disturbance by animals.
At the end of April 2012, fruits were retrieved (i.e. after 16 months burial and prior to the onset of the winter season) and x-rayed (as previously described) to ascertain seed fill. Fruits with no seed were removed from the replicate and the remaining fruits were returned to the jarrah forest or restoration site. Half of the fruits retrieved from the jarrah forest were returned to the forest and either buried or scattered on the soil surface. The other half were either scattered or buried on the most recently restored area (2011 restoration site) of the mine. Likewise, fruits retrieved from the restored areas of the mine were either returned to new restoration sites on the mine (2011 restoration site) or placed in a natural bushland (i.e. jarrah forest) environment (either scattered on the soil surface or buried). This resulted in 11 different treatments, comprising initial burial area (forest vs restoration site), return burial area (forest vs restoration site) and fruit sowing treatment (surface scattered vs soil burial) each with four replicates. These replicates were initially of 20 fruits however after X-ray analysis, the numbers of fruits in each treatment varied depending on seed fill, as all non filled fruits were discarded.
After reburial, the trial was assessed for seedling emergence in October 2012 and then quarterly (late December 2012, April 2013, July 2013, October 2013) for 1 year. It was then reassessed in April 2014 and again in August 2014.
Some of the results from this trial raised the question that differences in soil temperatures and moisture levels between natural jarrah forest sites and mine restoration areas were driving some of the disparity in emergence patterns observed. Hobo Microstation data loggers (Onset Computer Corporation, Cape Cod Massachusetts) with soil moisture probes (Soil Moisture Smart Sensors) and soil temperature probes (12-bit Temperature Smart Sensor) were placed in a freshly restored area (restored in 2013) and in an adjacent natural bushland area in October 2013 to quantify soil conditions at approximately 2 cm below the soil surface, within the two different environments. Measurements were made on an hourly basis and data loggers remained in place for approximately 1 year.
Seedling survival on restored areas
Forty seedlings were available during 2010 and given the small numbers it was decided to use these seedlings in a preliminary study of seedling survival at several different sites. Seedlings germinated in early August 2010 and were pricked out into biodegradable pots filled with Native Potting Mix (Baileys Fertilizer, Kwinana, Western Australia) shortly afterwards. They were left to recover from the initial transplant shock in Alcoa’s Marrinup Nursery, Dwellingup for 2 weeks before planting out into different sites in late August 2010. The seedlings were planted directly onto the various areas in the biodegradable pots so no additional disturbance to the roots occurred during the planting process. Ten seedlings were planted, with 2 cm wire mesh cages (to prevent grazing), into each of the following areas:
-
newly established restoration site with no vegetation (restored 2010);
-
3 year old restoration site with some recently established over-storey vegetation (restored 2007);
-
an area in natural jarrah forest burnt in spring 2009 (i.e. no leaf litter but surrounding trees still present) and;
-
unburnt natural jarrah forest with an extensive layer of leaf litter.
Seedling survival and height (from ground level to the apical meristem) were measured on a monthly basis from September 2010 to April 2011 and then quarterly until April 2012 (a period which covered two summers).
Seedling age and tree guard trial
A second trial was established to investigate the impacts of using guards to protect seedlings from herbivores and to determine whether plants of different ages perform differently when planted into freshly restored areas. Seedlings for this trial were germinated late July/early August 2011 and were pricked into biodegradable pots as previously described. Half of the potted plants were planted directly into the restored area within the biodegradable pots, in late August 2011 (several weeks after emergence).
At the time of planting, seedlings were either planted without a tree guard (control plants), or with an “onion bag” tree guard (Bug-it-off netting, Buono Net Australia Pty Ltd. Homebush, NSW) or a purpose built shade cloth (50 % green shade cloth) guard. Both types of guards were held in place with three bamboo poles to pull them tort. Altogether, 40 seedlings were allocated to each treatment and each treatment was replicated in four different pits (i.e. 10 seedlings per treatment per pit).
Onion bag tree guards (32 cm tall with sides of approximately 15 cm) provide protection from grazing and given their loose weave allowed some natural air movement (Fig. 1). Eight months after installation they had begun to deteriorate and disintegrate due to their UV sensitivity. The shade cloth tree guards were made up to a similar size to commercially available plastic tree guards (i.e. 46 cm tall and stretched to a triangle with sides of approximately 23 cm).

Tree guards used to protect seedlings during trials on restored areas of the mine. a Onion bag tree guard. b Shade cloth tree guard. Photos by K. Chia
A second batch of plants that germinated at a similar time (July/August 2011) that had been grown in forestry tubes (5 cm × 5 cm × 12 cm) were planted into freshly restored areas (i.e. 2012 rehabilitation), in August 2012, and were 12 months old at the time of planting. Given the limited numbers, there were only sufficient plants for installing two guarding treatments; no protection (control) and onion bag guards (which were considered the most economical of the two different guards treatments assessed in terms of price and availability). Fourteen seedlings were allocated to each treatment and each treatment was replicated in three different pits.
Data analysis
Synthetic seed bank emergence data at the final retrieval date were analysed using a Generalised Linear Model (GLM) with mixed effects and a logit link function to determine the effects of different treatments. Four blocks were nested within the forest and restoration sites under wire cages. At the time of initial retrieval the seed bags were allocated to either reburial or scattering over a forest or restoration site, meaning reburial location and treatment were modelled as crossed treatment factors. The synthetic seed bank data were analysed for the effects of burial treatment (buried vs surface scattering), initial burial location (either restored or bush), and return burial location (either restored or bush). A random effect for the pre-burial block was also included.
Survival data for the trial investigating guarding was firstly analysed using the Cox proportional hazard model with mixed effects to quantify the effects of age of seedling at planting (i.e. late vs early plant), tree guarding type (either no guard, onion bag or shade cloth), and months after planting. Pit number was included as the random (frailty) effect. Sample mean and standard errors were used for the plotting of effects, with an equivalent binomial GLM model applied to give a Tukey HSD interval for pairwise comparisons between treatment effects at each month after planting.
Height data for both seedling trials were analysed using a linear mixed effects model. The fixed effects were again the age of seedling at planting, tree guarding type and months after planting. The random effects were a tree identifier nested within pit number. A Tukey HSD interval was applied to each pairwise comparison, and sample means and standard errors were used for plotting the data.
Soil temperature and moisture data was analysed with a paired t test using the mean daily maximum and minimum soil values for the natural bushland and restoration sites.
All data analyses were undertaken in the statistical program R (R Core Team 2013) using the lme4 package.
Results
Germination on restored areas of different ages
Very low emergence (<1 %) was observed from fruits either in the cages or adjacent to the cages on either the 3 year old (2007) or newly established (2010) restored areas during the course of the study. Two germinants were observed in April 2013 and these were recorded from fruits that had been buried inside the cages on the newly established restoration site. It is likely that these seedlings germinated in August or September 2012 approximately 24 months after initial burial. This estimate of germination date is based on the size of the seedlings when observed and observations of timing of seedling germination made during phenological studies (Chia et al. 2015). However, of the 480 fruits placed out on both the 2010 and 2007 restoration areas, this represents <1.0 % emergence after 2 years.
Synthetic seed bank trial
Seed fill at the commencement of the study in December 2010 was 83.0 ± 3.8 %. After 16 months in situ placement seed fill in fruits retrieved from the restoration area (76.1 ± 1.4 %) and the natural forest (77.5 ± 3.0 %) in April 2012 were not significantly different from the initial seed fill or from each other.
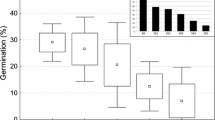
Modelling (using GLM analysis) indicated that emergence success was significantly affected by all three treatments (i.e. location of initial burial, location of return burial and sowing treatment) (Table 1). The best emergence (24.0 ± 2.7 % after 26 months) was observed for fruits that had been initially buried in natural bushland, then retrieved and reburied in the restored areas (Fig. 2c, ANOVA, P < 0.001). Once the fruits had been retrieved, burial (rather than scattering) was the best means of obtaining emergence. Those fruits that were scattered on top of the soil germinated very poorly compared to all the other treatments assessed (Table 1; Fig. 2c).

Cumulative germination percentage (±SE) recorded in each treatment following retrieval and re-burial/scattering. Treatments with different letters are significantly different. a Germination of fruits initially buried in the bush. b Germination of fruits initially buried in the restoration. c Germination of fruits that were left in position for the duration of the experiment
A total of 64 seedlings was recorded in this experiment over the period of the study. Eight of these seedlings were recorded in the last assessment (August 2014) and as a result their survival and growth beyond this point is unknown. However, survival of seedlings from previous assessments was generally poor with only 6.4 % of seedlings (i.e. three seedlings) observed from previous seasons (2012 and 2013) surviving until August 2014. Two of these seedlings were located in the natural bushland site and one was located on the mine restoration area (Fig. 3a). Seedling deaths most likely occurred as a result of desiccation (Fig. 3b) or as a result of heavy insect damage (Fig. 3c). Only a few seedlings were observed to have been grazed by herbivores (Fig. 3d).

Germinants from the synthetic seed bank trial growing in restoration areas a healthy seedling that survived to the end of the trial (2014), b healthy seedling adjacent to a desiccated seedling, c seedling with insect damage adjacent to a healthy seedling and d seedling grazed by unknown herbivores. Photos K. Chia
Interestingly, both the minimum and maximum soil water contents were generally more extreme in the restored areas compared to the natural bushland sites, and overall, restoration sites generally had higher soil moisture during the summer months than the surrounding bushland (Fig. 4a, b, paired t test, P < 0.001). Maximum temperatures recorded at 2 cm below the soil surface were higher on the restored area than in the adjacent natural bushland site (Fig. 4c, paired t test, P < 0.001). There were several days during the summer months when the soil temperature difference between the two different locations was as high as 13 °C. During the winter months maximum temperatures on the restored area were generally 5 °C higher than in the adjacent natural bushland site.

Comparison of soil temperatures and moisture levels on a restored pit at Boddington bauxite mine and the adjacent jarrah forest at 2 cm depth. a Mean maximum water content. b Mean minimum water content. c mean maximum temperature, d mean minimum temperature
While minimum temperatures were variable between sites they were still generally a little higher on the restored areas than in the natural bushland sites (Fig. 4d, paired t test, P < 0.001). However there were occasions during winter when the temperatures recorded on the restoration sites were lower than in the natural bushland sites. While the data loggers were in the ground for a full 12 months, due to technical difficulties no records were made from 14 August 2014 to 30 October 2014, during the transition from late winter into spring.
Effects of age of restoration site and bush planting on seedling survival
The majority of deaths occurred during the first summer (November to March) following planting. All deaths within the newly established restoration sites occurred within the first 2 months (August to October) of planting (Fig. 5). Initially, survival of seedlings in the 3 year old restoration site was similar to that in the newly established restoration site but as the summer progressed and soil moisture dropped seedling survival in the 3 year old restoration site was reduced to 50 %. After the second summer this then dropped to only 40 % survival.

Cumulative survival of seedlings following planting onto different post mining restoration and bush with different fire histories around the McCoy bauxite mine operated by Alcoa near Dwellingup. Treatments with the same letter are not significantly different
Those seedlings planted in the burnt bush showed similar survival to those seedlings planted in the restored areas of the mine site with 70 % surviving the first summer and two more deaths resulting in a final survival of 50 %. No seedlings in the natural unburnt bush survived past the first summer (Fig. 5).
A major outcome of this preliminary trial was the rapid and pronounced increase in height of those seedlings planted into the newly established restoration site when compared with all other sites assessed (Fig. 6, Tukey HSD pairwise comparison, P < 0.001 in all cases). This sizeable height difference was evident from monitoring undertaken in January 2012 which was the second summer after the seedlings were planted (17 months after planting) with seedlings growing rapidly between October 2011 and January 2012 (Fig. 6).

Mean height (cm ± SE) of surviving Persoonia longifolia plants when placed into different environments around Dwellingup. Treatments with same letters are not significantly different
Seedling age and tree guard trial
The survival analysis indicated there was no significant difference between the overall survival of seedlings planted at 2 weeks of age compared with those planted at 12 months of age (Table 2). However closer inspection of the data at each time after planting (using a GLM with time after planting included as a predictor thus ignoring dependence amongst the data through time) showed that at 3 months after planting, seedlings planted at 12 months of age survived better than those that were planted at 2 weeks of age after 3 months in the ground (Fig. 7a). However, 8 months after planting the seedlings, these differences were no longer significant. Similarly, 20 months after planting it did not matter if the seedlings had been planted at 2–4 weeks of age or 12 months of age as there was no significant difference in survival between the five treatments (Fig. 7a).

a Mean survival (% ± SE) in relation to the months after planting of seedlings onto mining restoration sites. Treatments within the 3 month assessment with the same letters are not significantly different (P = 0.005). None of the treatments assessed at eight and 20 months after planting were significantly different from one another. b Seedling height in relation to tree age at the time of assessment. Treatments within each age group with the same letters are not significantly different (P = 0.005)
When comparing the various guarding treatments, survival was greatest in those seedlings planted in the onion bag tree guards (Table 2). However, whilst use of the shade cloth tree guards did not significantly improve survival of the seedlings, it did result in a significant increase in height when compared with those seedlings in either the onion bag tree guards or not guarded in any way (Table 2).
When comparing the height of the seedlings in each of the treatments at different ages (using a GLM), the difference between those seedlings planted at 2–4 weeks of age and those planted at 12 months of age began to be evident once the plants had attained 14 months of age. This is only 3 months after the 12 month old seedlings were planted into the restored areas. Seedlings that were kept in the nursery for 12 months were similar in size to 3 month old seedlings planted out early (there was no significant difference between late plant 12 months old seedlings and the early plant 3 month old seedlings), indicating that the seedlings grew relatively little under nursery conditions.
Those seedlings planted at 2 weeks of age in shade cloth tree guards were significantly taller than all other seedlings by the time the seedlings were 20 months of age (ANOVA, P < 0.001, Fig. 7b). This trend was still evident following 32 months of growth (Figs. 7b, 8).

Thirty two month old seedlings surviving on the restored areas of a bauxite mine at Dwellingup. These two seedlings were adjacent to each other on the restoration site. a Seedling protected by onion bag guard (onion bag had disintegrated at the time photo was taken in April 2014). b Seedling protected by shade cloth guard. Photos K Chia
Records of insect/herbivore damage show that initially (the first 14 months after planting), there was very little insect damage evident on any of the P. longifolia plants (Table 3). However by the time the seedlings were 20 months old insect herbivory was clearly evident. Only one seedling was noted as having been grazed by a larger herbivore during this experiment. Seedlings in shade cloth guards experienced less insect damage compared with those seedlings left unguarded or those protected by onion bag tree guards, although this was not statistically significant (Tukey’s HSD pairwise comparisons, P > 0.05).
Discussion
The return of complex ecosystems to post-disturbed areas that reflect the diversity and functionality of the pre-disturbed system has now become a priority for many mining companies around the world (Koch 2007b; Bielecka and Król-Korczak 2010; Macdonald et al. 2015) However, many species within these ecosystems continue to prove difficult to return despite their commonality in the original native vegetation. Therefore, to reinstate a broad representation of the local native species a range of techniques and approaches is generally required such as direct return of topsoil, supplementary direct seeding, and the planting of actively growing greenstock derived from different sources including plant tissue culture, cuttings, seeds and rhizomes (Koch 2007a, b). In this context, the approaches outlined in this study fit neatly within current restoration activities undertaken by mining companies and complement their existing programs.
Use of seeds for restoration
The use of seeds is the cheapest and most effective means of returning species to restored areas when germination occurs readily or seed dormancy is easily and effectively overcome (Koch 2007a, b). However, if germination does not occur when standard seeding methods are used then other means of returning these species must be considered (Koch 2007b). Persoonia longifolia is one such species that does not readily germinate on post mining restoration sites when incorporated into a standard seed mix that is scattered over the soil surface during the restoration process (Mullins et al. 2002; Norman and Koch 2008). This was confirmed with the low emergence observed in this study (0.4 %) in both newly restored areas and 3 year old restoration sites and was similar to emergence (0.3 %) achieved in other field broadcast trials undertaken in previous research (Norman and Koch 2005).
The creation of a synthetic seed bank, whereby fresh seeds are initially buried then later exhumed and sown onto restoration sites, is a novel approach for returning difficult to germinate species and is presented here for the first time. By burying the fruits in their natural environment (the surrounding jarrah forest in this case) it is possible to cue the seed for germination through breaking dormancy within the seed before retrieving the fruits and sowing these at a later time (once dormancy has been partially alleviated) into restoration sites.
Results from this trial indicate that emergence of P. longifolia was enhanced when fruits or endocarps were buried in the soil. Many studies have shown that seed burial between 0 and 5 cm deep improves germination in many different ecosystems (Blackshaw 1990; Jurado and Westoby 1992; Ren et al. 2002). For example, seed burial for 12 Western Australian jarrah forest species resulted in a high level of emergence from 2 cm burial with the optimum germination occurring between 0 and 5 cm for all species and in this context P. longifolia is another species that germinates much more readily when buried (Grant et al. 1996). Consequently, scattering fruits on the soil surface, results in very low germination and based on these results is a highly ineffective way of returning P. longifolia to restoration sites following mining. Indeed, burying dormant endocarps in the forest environment exposes them to the natural conditions that break dormancy and so cues seeds for germination, and burial for up to two summers does not appear to affect seed viability in P. longifolia.
In these experiments, the fleshy mesocarp was retained on the endocarp as this would be the least expensive way to sow in situ P. longifolia seeds for restoration purposes as removal is somewhat specialised and takes several hours to effectively complete (Chia Pers Obs.). However, results from other trials undertaken (Chia 2015, unpublished data) show that removal of the mesocarp can also greatly improve germination (from 17.5 % up to 44.2 %) especially when combined with soil burial so given the relative cost of P. longifolia fruits and the importance of returning it to restoration sites, the extra resources required to depulp the endocarps prior to their utilisation is worth the extra investment in time and resources.
However, whilst using a synthetic seed bank as described in this study, substantially increased the numbers of P. longifolia emerging in situ in restoration sites, the survival of seedlings that did germinate was still very poor and methods of improving survival require further investigation.
Use of seedlings for restoration
Indeed, in cases where survival of in situ seedlings is poor and where the cost of the seed is at a premium or if affordable, relatively difficult to come by in sufficient quantities, the germination of seedlings under nursery conditions and then planting onto restored areas can result in better survival and in some cases may be a more effective use of expensive or limited seed resources.
Planting out seedlings has not been done regularly with P. longifolia largely as a result of a lack of seedling availability. Being a mid-storey tree, P. longifolia is likely to germinate in a shaded and somewhat protected environment which is very different from the open and barren post mining landscape. The initial working hypothesis for this trial was that areas of the mine requiring restoration are so fundamentally different from the surrounding forest environment (in terms of temperature, moisture, wind, relative humidity and herbivory), that P. longifolia cannot easily germinate, and seedling survival and growth for any that do manage to emerge is very difficult at best. However, this did not prove to be the case, with survival of seedlings on the newly established restored sites greater than in the surrounding natural forest or even on the 3 year old restoration sites. This is likely to be related to the significant difference in soil temperature and moisture identified between the newly restored areas and the surrounding forest. In fact, the increased availability of water in newly restored areas compared with the surrounding natural forest appears to be somewhat advantageous for P. longifolia and it is likely that this is related to a lower level of competition from surrounding vegetation on the newly restored sites. Reducing competition has been found to also improve survival and growth of many other jarrah forest species during early growth and development and has lead Alcoa to adopt a one-pass approach to restoration whereby all restoration works involving the reintroduction of native plants is completed within a short window following the completion of all earthworks and landscaping (Abbott 1984; Daws and Koch 2015).
Seedling deaths were greatest in the first few months following planting in both seedling trials, indicating that most deaths are likely to occur as a result of the profound change in conditions as they transition from the nursery environment to in situ conditions. Transplant shock or seedling damage is less likely to have been an issue for the younger plants, as seedlings were placed directly into the ground in biodegradable pots with minimal root disturbance whilst the later planting did involve the removal of plants from forestry tubes prior to planting which may have unduly impacted or disturbed their root system to some degree. Rainfall during September and October of 2010 was well below average (Dwellingup weather station no. 009538) (Bureau of Meteorology 2015), however, despite this very low rainfall, seedling survival in the first trial was still 60 % after 3 months which is similar to survival experienced with other jarrah forest species planted into restored areas in normal rainfall years (Daws and Koch 2015). However, seedling survival when planted in the surrounding jarrah forest was variable. Large amounts of deep leaf litter present in unburnt forest may have inhibited water penetration even during heavy rainfall and high levels of competition may explain the lack of seedling survival in this environment. In the burnt forest, where leaf litter and competition from surrounding vegetation was greatly reduced, survival was similar to that on the newly established restoration sites. However, unlike restoration sites, the over-storey of tall E. marginata and C. calophylla trees was still present albeit with a reduced canopy initially.
Whilst survival is clearly reduced if seedlings are planted out when 2 weeks old (55 % compared with 85 % for seedlings planted out when 12 months old), deaths under nursery conditions were also common, but not recorded during this study. To date, no data is available on seedling survival and growth under nursery conditions. Greater in situ survival at 3 months after planting of the older seedlings is an indication that transplant shock is less of an issue for older plants. However, by the time the seedlings had been growing in situ for 20 months there was no difference in survival between those P. longifolia planted at only a few weeks of age and those planted 12 months later (but were, at that point, 32 months old).
Anecdotal evidence (Chia pers. obs.) indicates that P. longifolia seedlings are palatable to herbivores and results from the guarding experiments found that planting seedlings with some form of tree guard improved seedling survival. Whilst open weave onion bag tree guards resulted in the best plant survival, those seedlings placed in shade cloth tree guards grew significantly taller and appeared to be in optimal health. The exact reasons for this rapid burst of growth are currently unknown but could be due to the reduction in insect damage to the plants that was noted (Table 3) or variations in microclimate (reduced light intensity, elevated humidity or reduced wind speed) within the guards. Nevertheless, while microclimatic variation was not measured as part of this study, Close et al. (2009) found that shade cloth tree guards provide several tangible benefits to young plants by preventing browsing, minimising temperature effects on seedlings and creating a microenvironment that significantly elevates the levels of photosynthesis in spring which, in turn results in greater growth of seedlings. Other species have also been observed to increase in height and spread with the installation of tree guards including several jarrah forest species that are sympatric with P. longifolia (Daws and Koch 2015).
This study of P. longifolia highlights the inadequacies of a number seeding techniques currently used in restoration (e.g. scattering of seeds on soil surface and retention of outer layers on the reproductive unit) within the mining industry, particularly for species with larger drupaceous indehiscent fruits such as P. longifolia, that could be revised to improve germination, seedling emergence, survival and growth. Clearly surface seeding of uncleaned P. longifolia fruits is not an effective way to re-establish this species onto post mining restoration sites. Likewise, breaking seed dormancy through burial prior to use in restoration sites is beneficial and improves restoration success. The results presented here also clearly show the advantages of using of tree guards for improving survival and growth of newly planted greenstock which, for important priority species is worthy of serious consideration. Given the many years of research, indeed decades that have been spent working on developing effective methodologies for propagating and re-establishing P. longifolia into disturbed environments we believe the results presented here greatly assist with not only this species but other equally problematic species as well as they occur in other parts of the world.
References
Abbott I (1984) Emergence, early survival, and growth of seedlings of six tree species in Mediterranean forest of Western Australia. For Ecol Manage 9:51–66. doi:10.1016/0378-1127(84)90037-9
Baskin CC, Baskin JM (2014) Seeds: ecology, biogeography, and evolution of dormancy and germination, 2nd edn. Elsevier Inc
Bielecka M, Król-Korczak J (2010) Hybrid expert system aiding design of post-mining regions restoration. Ecol Eng 36:1232–1241. doi:10.1016/j.ecoleng.2010.04.023
Blackshaw RE (1990) Influence of soil temperature, soil moisture, and seed burial depth on the emergence of round-leaved Mallow (Malva pusilla). Weed Sci 38:518–521. doi:10.2307/4045066
Bureau of Meteorology (2015) Climate data online. Commonwealth of Australia. http://www.bom.gov.au/climate/data/index.shtml. Accessed 14 Dec 2015
Chia KA, Koch JM, Sadler R, Turner SR (2015) Developmental phenology of Persoonia longifolia (Proteaceae) and the impact of fire on these events. Aust J Bot 63:415–425. doi:10.1071/BT14315
Chia KA, Sadler R, Turner SR, Baskin CC (2016) Seasonal conditions required for dormancy break of Persoonia longifolia (Protecaeae), a species with a woody indehiscent endocarp. Ann Bot (in press)
Close DC, Ruthrof KX, Turner S, Rokich DP, Dixon KW (2009) Ecophysiology of species with distinct leaf morphologies: effects of plastic and shadecloth tree guards. Restor Ecol 17:33–41. doi:10.1111/j.1526-100X.2007.00330.x
Daws MI, Koch JM (2015) Long-term restoration success of re-sprouter understorey species is facilitated by protection from herbivory and a reduction in competition. Plant Ecol 1–12 doi:10.1007/s11258-015-0459-7
Grant CD, Bell DT, Koch JM, Loneragan WA (1996) Implications of seedling emergence to site restoration following bauxite mining in Western Australia. Restor Ecol 4:146–154. doi:10.1111/j.1526-100X.1996.tb00115.x
SERA Standards Reference Group (2016) National standards for the practice of ecological restoration in Australia. Soc for Ecol Restor Aust. http://www.seraustralasia.com/standards/contents.html
Jurado E, Westoby M (1992) Seedling growth in relation to seed size among species of arid Australia. J Ecol 80:407–416. doi:10.2307/2260686
Koch JM (2007a) Alcoa’s mining and restoration process in south Western Australia. Restor Ecol 15:S11–S16
Koch JM (2007b) Restoring a jarrah forest understorey vegetation after bauxite mining in Western Australia. Restor Ecol 15:S26–S39
Koch JM, Richardson J, Lamont BB (2004) Grazing by kangaroos limits the establishment of the grass trees Xanthorrhoea gracilis and X. preissii in restored bauxite mines in eucalypt forest of southwestern Australia. Restor Ecol 12:297–305. doi:10.1111/j.1061-2971.2004.00335.x
Lai PCC, Wong BSF (2005) Effects of tree guards and weed mats on the establishment of native tree seedlings: implications for forest restoration in Hong Kong, China. Restor Ecol 13:138–143. doi:10.1111/j.1526-100X.2005.00016.x
Macdonald SE et al (2015) Forest restoration following surface mining disturbance: challenges and solutions. New Forest 46:703–732. doi:10.1007/s11056-015-9506-4
Marchant NG, Wheeler JR, Rye BL, Bennett EM, Lander NS, Macfarlane TD (eds) (1987) Flora of the Perth Region part one, vol 1. Western Australian Herbarium, Department of Agriculture, Western Australia
Mullins RG, Koch JM, Ward SC (2002) Practical method of germination for a key jarrah forest species: snottygobble (Persoonia longifolia). Ecol Manage Restor 3:97–103
Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858
Nield AP, Monaco S, Birnbaum C, Enright NJ (2015) Regeneration failure threatens persistence of Persoonia elliptica (Proteaceae) in Western Australian jarrah forests. Plant Ecol 216:189–198. doi:10.1007/s11258-014-0427-7
Norman MA, Koch JM (2005) Differences in species abundance between sites rehabilitated with direct return and stockpiled soil. Aloca World Alumina Australia, Perth
Norman MA, Koch JM (2008) The effect of in situ seed burial on dormancy break in three woody-fruited species (Ericaceae and Proteaceae) endemic to Western Australia. Aust J Bot 56:493–500
Norman MA, Koch JM, Grant CD, Morald TK, Ward SC (2006) Vegetation succession after bauxite mining in Western Australia. Restor Ecol 14:278–288. doi:10.1111/j.1526-100X.2006.00130.x
Ren J, Tao L, Liu X-M (2002) Effect of sand burial depth on seed germination and seedling emergence of Calligonum L. species. J Arid Environ 51:603–611. doi:10.1006/jare.2001.0979
Stanton-Clements EM, Koch JM, Daws MI (2013) Effectiveness of plant guards in reducing grazing of Tetraria capillaris in restored bauxite mines in Western Australia. S Afr J Bot 87:4–8. doi:10.1016/j.sajb.2013.03.006
Stingemore JA, Krauss SL (2012) Genetic Delineation of local provenance in Persoonia longifolia: implications for seed sourcing for ecological restoration. Restor Ecol 21:49–57. doi:10.1111/j.1526-100X.2011.00861.x
R Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing. http://www.R-project.org/
Todd AJ, Rufaut CG, Craw D, Begbie MA (2009) Indigenous Plant Species Establishment during Rehabilitation of an Opencast Coal Mine, South-East Otago, New Zealand. N Z J For Sci 39:81–98
Ward SC, Koch JM, Ainsworth GL (1996) The effect of timing of rehabilitation procedures on the establishment of a jarrah forest after bauxite mining. Restor Ecol 4:19–24
Acknowledgments
The authors wish to thank the Australian Government for providing an Australian Postgraduate Award for this study, and University of Western Australia for providing a top up scholarship. Thanks also go to Alcoa of Australia Limited and South32 Worsley Alumina for providing financial and practical industry assistance for this study and also the Mining Research Institute of Western Australia for its financial contribution to the project. We also acknowledge the valued comments of two anonymous reviewers whose suggestions greatly improved this paper.
Author information
Authors and Affiliations
Corresponding author
About this article

Cite this article
Chia, K.A., Koch, J.M., Sadler, R. et al. Re-establishing the mid-storey tree Persoonia longifolia (Proteaceae) in restored forest following bauxite mining in southern Western Australia. Ecol Res 31, 627–638 (2016). https://doi.org/10.1007/s11284-016-1370-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11284-016-1370-y