
Abstract
Grassland communities occupy about 25 % of global land area and global warming could alter grassland plant community coverage. A field study was conducted to investigate the impact of soil warming (surface soil temperature increased by 1.3 °C) and nitrogen addition (100 kg N ha−1 year−1) on a desert steppe community in Inner Mongolia, China, from 2006 to 2011. Although there were strong intra- and inter-annual variations, in general, warming had little effect on coverage of the four major species (two cool season C3 species: Convolvulus ammannii and Stipa breviflora, and two warm season C4 species: Cleistogenes songorica and Kochia prostrata) over six growing seasons. The C3/C4 coverage ratio consistently decreased with warming, with the decrease being statistically significant for one-third of sampling dates. Warming caused a reduction (from 27.6 to 25.1 %) in peak season total plant community coverage. N addition did not affect the four main species, four functional groups (perennial grass, perennial forb, shrub and annual-biennial) or the total plant community over the 6 years, except when precipitation was high (231 mm in 2008). The small reduction in plant community coverage demonstrated the resilience of desert steppe to warming. There could be a shift of C3 to C4 species because of warming, as indicated by the decrease in C3/C4 ratio, but confirmation of this trend requires further study.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Climate change has become a worldwide concern in recent years, including its impact on plant productivity (Knapp and Smith 2001; Reynolds et al. 2004). Global mean air temperature has increased by 0.76 °C since 1850, and is predicted to rise an additional 1.8–4.0 °C by the end of this century (Solomon et al. 2007). Numerous studies, such as long-term observations (Chapin et al. 1995), model simulations (Botkin et al. 2007), warming manipulation experiments (Cornelissen et al. 2007; Rustad 2008) and meta-analyses (Walker et al. 2006; Lin et al. 2010a), have indicated that plant communities and species composition are very sensitive to temperature increases. Research has also shown that there are no consistent responses among plant species or functional groups to increasing temperature (Van Wijk et al. 2004; Luo 2007).
Changes in plant species and functional groups in response to climate warming could modify species relations (Niu and Wan 2008). For example, warming might change species relationships directly by affecting the proportion of dominant species in the community (Kardol et al. 2010). Warming might also indirectly change the composition of plant community by affecting abiotic factors such as soil moisture, which might benefit plant growth in temperature-limiting regions and accelerate net nitrogen (N) mineralization (Rustad et al. 2001; Peñuelas et al. 2007; Yang et al. 2011).
Lower temperatures are reported to enhance growth of cool season C3 species, whereas higher temperatures might benefit warm season C4 species (Winslow et al. 2003). Competition between C3 and C4 photosynthetic types is dictated by temperature during the plant growing season (Auerswald et al. 2009; Xu et al. 2014). Climate change, such as change in seasonal precipitation (e.g., in the early part of summer) and increases in temperature, have been shown to benefit the expansion of C4 species over C3 species (Murphy and Bowman 2007). Warming has more variable effects on plant functional groups and generally stimulates shrub and forb biomass accumulation (Walker et al. 2006; Xia et al. 2009) and reduces grass biomass (Zavaleta et al. 2003; Yang et al. 2011).
N input is an important driver of changes in terrestrial plant community composition (Vitousek and Howarth 1991). N addition can cause divergence of plant communities by promoting the dominance of a few species (Tilman 1993; Craine et al. 2003). Plant functional groups have been used to categorize plant responses to global changes by N addition (Cleland et al. 2006; Bai et al. 2010). For example, Xia and Wan (2008) showed that the response of grass to N addition was more sensitive than that of forbs and semi-shrubs. A large number of studies have found that N addition or atmospheric N deposition affects the growth of both C3 species and C4 species, but in different fashions (Yuan et al. 2007; Niu et al. 2008). N use and resorption by C4 species are higher in the warm summer and lower in the cool spring and winter seasons, whereas the opposite is true for C3 species (Yuan et al. 2007).
Responses of plant communities and species composition to warming are the result of many complex, congruent processes, and it is difficult to predict plant community dynamics in the context of climate change. A field experiment was conducted to examine the effects of warming, N addition and their interaction on plant community coverage in a desert steppe. The following hypotheses were tested: (1) warming affects the plant community structure and composition by reducing C3 species and increasing C4 species; (2) N addition increases plant community productivity by promoting the growth of grasses over that of forbs, similar to that reported for temperate steppe by Xia and Wan (2008); and (3) a shift from C3 to C4 might occur under a warming environment.
Materials and methods
Study site
The study was conducted in a desert steppe in Siziwang Banner (41°46′43.6″ N, 111°53′41.7″ E, 1456 m a.s.l), Inner Mongolia, China, which has brown chestnut soil (Haplic Calcisols according to the FAO classification). The 50-year mean annual temperature in the study area is 3.4 °C and the annual precipitation is approximately 280 mm (Lin et al. 2010b). The 50-year mean growing season (May to September) temperature is 15.6 °C and precipitation is approximately 243 mm. The dominant plant species in the desert steppe include Stipa breviflora Griseb (S. breviflora) and Cleistogenes songorica Roshev. (C. songorica), with a primary productivity of 10–100 g m−2 year−1. The main co-dominant species are Convolvulus ammanni Desr. (C. ammanni), Artemisia frigida Willd. (A. frigida), Salsola collina Pall. (S. collina), Leymus chinensis (Trin.) Tzvel. (L. chinensis), Allium tenuissimum (A. tenuissimum), Kochia prostrata (L.) Schrad. (K. prostrata) and Caragana stenophylla Pall. (C. stenophylla). In the experimental plots, two C3 species (S. breviflora, C. ammanni) and two C4 species (C. songorica, K. prostrata) were the dominant plant species, accounting for 80 % of total plant coverage.
Experimental design
The experiment used a paired, split-plot design with warming as the main plot and N addition as the subplot. There were six pairs of 3 × 4-m main plots (each pair forming a block), in which one plot was assigned for the warming treatment (W+) and the other one for control treatment (W0). The plot was divided into two 2 × 3-m subplots, one of which was randomly assigned for no N addition (N0) and the other for N addition treatment (N+). For the N addition treatment, N (NH4NO3) was broadcast at a rate of 10 g N m−2 year−1 (or 100 kg N ha−1 year−1) and applied on June 23 2006, July 1 2007, June 26 2008, June 30 2009, July 2 2010 and June 28 2011.
The warming plots had been heated continuously since May 2006 by using 165 × 15-cm MSR-2420 infrared radiators (Kalgo Electronics, Bethlehem, PA, USA, power: 2000 W), which were suspended 2.25 m above the ground. ‘Dummy’ heaters (not powered) of the same shape and size as the infrared radiator were installed in the control plots to ensure that both the treatment and control plots were subjected to the same shading effect by the heaters. The surface soil temperature (0 cm) in the W+ treatment was 1.3 °C higher than the control (W0) (Wang et al. 2011).
Vegetation sampling
One permanent 1 × 1-m quadrat was established in each of the 24 subplots in June 2006. Plant species community was surveyed in each quadrat every month from May to September each year. A 1 × 1-m frame, subdivided into 10 × 10-cm grids, was placed during measurement on top of the canopy in each permanent quadrat to measure vegetation coverage. In all permanent quadrats, the percent coverage of each species was estimated by summing the coverage of that species in each 10 × 10-cm grid. Plant species in the subplots were grouped into four plant functional communities: forbs, grasses, semi-shrubs, and annual-biennials (short–lived <2 years; Jiao et al. 2006; Bai et al. 2008).
Data analysis
The coverage ratio of C3 to C4 species was calculated as the sum of coverage of S. breviflora and C. ammanni divided by the sum of coverage of C. songorica and K. prostrata. Coverage data were analyzed for dominant species, functional groups, and the total coverage. Analyses were conducted separately for each sampling date, using a split plot design in PROC MIXED of SAS (SAS Institute Inc., 2010) with warming, N addition and their interaction as fixed effects. Data from August, the month in which annual maximum coverage is normally reached, was used to examine the effect of experimental year, with year treated as a repeated measurement using the heterogeneous auto-regressive covariance structure in the model. The UNIVARIATE procedure (SAS Institute Inc., 2010) was used to check the residuals for normality and potential outliers, and outliers were removed prior to all statistical analyses. The Kenward-Rogers method was used to adjust denominator degree of freedom. When fixed effects were significant, means were identified using the protected least significant difference (LSD) test of the LSMEANS procedure. Simple correlation was used to assess relationships between monthly coverage, monthly cumulative precipitation, and mean temperature.
Results
Precipitation and temperature
During the experimental period, precipitation in the growing season (May–September) was 133, 129, 210, 151, 164, and 130 mm for the years from 2006 to 2011, respectively, which were 53–86 % of the long-term average (243 mm). The average growing season temperatures were 17.3, 17.0, 15.3, 15.9, 18.0 and 15.9 °C from 2006 to 2011, respectively, and ranged from 0.3 °C below to 2.4 °C above the long-term average (15.6 °C; Fig. 1a).

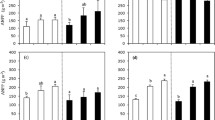
Growing season monthly temperature and precipitation and responses of seasonal coverage of two C3 species (Stipa breviflora Griseb., Convolvulus ammanni Desr.) and two C4 species (Cleistogenes songorica Roshev., Kochia prostrata L. Schrad.) to warming (W), nitrogen addition (N) and their interactions from 2006 to 2011 (W0N0 = control and no N addition, W+N0 = warming and no N addition, W0N+ = control and N addition, W+N+ = warming and N addition)
Coverage of two main C3 species
S. breviflora coverage ranged from 0.9 % (June 2011) to 9.2 % (June 2009) and was affected by warming, N addition and their interactions on 17 out of 29 sampling dates (Fig. 1b). Warming significantly decreased S. breviflora coverage in July 2007, July and September, 2008, July and August, 2009, June 2010 and July 2011 (P < 0.05). N addition significantly decreased S. breviflora coverage in August and September 2010, and May and August 2011 (P < 0.05, Fig. 1b), whereas S. breviflora coverage increased with N addition in July 2009 (Fig. 1a). The combination (interaction) of warming and N addition significantly decreased S. breviflora coverage in August 2006, August 2009, June and September 2010 and June 2011. S. breviflora coverage at peak production was significantly different and was affected by sampling year and the interaction of N addition and sampling year (Table 1). The peak coverage was greatest in 2008 and lowest in 2011 (Table 1). N addition decreased coverage of S. breviflora under warming conditions in 2006 (Table 2).
C. ammannii coverage ranged from 0.3 % (May 2010) to 14.0 % (August 2006) (Fig. 1c). Warming only significantly decreased C. ammannii coverage in August, 2008 (P < 0.05, Fig. 1c). Peak production C. ammannii coverage was significantly different in different sampling years, with the highest values in 2006 (8.7 %) and the lowest values in 2011 (2.5 %) (Table 1).
Coverage of two main warm season C4 species
C. songorica coverage ranged from 0.0 % (September 2010) to 10.9 % (September 2006) and was generally not significantly affected by warming, N addition and their interactions, except on six sampling dates (Fig. 1d). C. songorica coverage increased significantly with warming in May and September 2010, but decreased with warming in May 2011 (P < 0.05). C. songorica coverage increased with N addition in September 2011 (P < 0.05, Fig. 1d). C. songorica coverage at peak production was significantly different in different years, with values ranked in the order: 2006 (8.4 %) and 2008 (7.9 %) >2007 (3.9 %) and 2010 (4.5 %) >2009 (1.7 %), and 2011 (1.7 %; P < 0.05, Table 1).
K. prostrata coverage was less variable (0.2–7.9 %) than C. songorica and was only significantly affected by warming and N addition interaction on one sampling date (July 2010; P < 0.05, Fig. 1e). Similar to C. songorica coverage, the peak K. prostrata coverage varied from year to year, with the largest values in 2006 (3.8 %) and the lowest in 2007 (1.2 %; Table 1).
C3/C4 coverage ratio
The ratios for W+ were significantly lower than for the W0 treatment in July and September 2006, September 2007, July 2008, June 2009 and May, June, July and September 2010 (P < 0.05, Table 3). There was an exception in May 2011 when the C3/C4 ratio of W+ was higher than that of W0. Compared with the effects of warming, the C3/C4 ratio was less affected by N addition; the ratio increased with N addition on two occasions (September 2007 and June 2009) and decreased with N addition on three occasions (September 2006, 2009 and 2010; Table 3).
Functional groups and plant community coverage
Perennial grass coverage was significantly affected by warming, N addition and their interaction on seven out of 29 sampling dates (P < 0.05, Fig. 2a). For example, perennial grass coverage decreased by 2–4 % with warming in July 2007, September 2008, and July and August 2009, and by 0.9 % with N addition in July 2011 (Fig. 2a). The interaction of warming and N addition significantly decreased perennial grass coverage in August 2006 and June 2010 (Fig. 2a). Perennial grass coverage at peak production (i.e., August each year) was not affected by warming or N addition, but was significantly different in different years, with the greatest value in 2008 (15.6 %) and lowest value in 2011 (3.8 %; P < 0.05, Table 4).

Functional group and total plant community responses to warming (W), nitrogen addition (N) and their interactions from 2006 to 2011 (W0N0 = control and no N addition, W+N0 = warming and no N addition, W0N+ = control and N addition, W+N+ = warming and N addition)
Perennial forb coverage ranged from 1.9 % (June 2007) to 21.1 % (August 2006). Perennial forb coverage was significantly affected by warming, N addition and their interactions on only four out of 29 sampling dates over the 6 years (P < 0.05, Fig. 2b). Perennial forb coverage decreased significantly with warming in September 2007 (1.4 %), but increased in May 2010 (0.9 %; P < 0.05). Perennial forb coverage also increased with N addition in August 2010 (P < 0.05, Fig. 2b). Perennial forb coverage at peak production varied significantly from year to year, with the greatest value in 2006 (18.6 %) and the lowest in 2007 and 2009 (6.0 %; P < 0.05, Table 4).
Shrub coverage was less variable in comparison with perennial forbs, ranging from 0.2 % (May 2010) to 6.9 % (September 2011) over the 6 years, and significant interaction effect of warming and N addition was observed only once out of 29 sampling dates (July 2010; Fig. 2c). The magnitude of variation at peak production (August each year) was small, ranging from 1.5 % in 2007 to 4.6 % in 2006 (P < 0.05, Table 4). Similarly, annual-biennial coverage was less than 5.3 % on most sampling dates, except in August and September 2008 when coverage was between 11.0 and 21.3 %, coinciding with the above-average precipitation in August of that year. Annual-biennial coverage was also not significantly affected by warming, N addition or their interaction (Fig. 2d), but varied significantly with year (P < 0.05, Table 4). Annual-biennial coverage at peak production was highest in 2008 (17.3 %) and lowest in 2009 (0.1 %; Table 4).
The total plant coverage ranged from as low as 4.6 % in May 2011 to as high as 52.2 % in August 2008 (Fig. 2e). The total plant coverage was significantly affected by warming on four occasions and by N addition on two occasions (P < 0.05). Warming significantly decreased plant community coverage in May and July 2008 and in June and July 2009, whereas N addition significantly increased plant community coverage in August and September 2008 (Fig. 2e), when monthly precipitation was above the normal (Fig. 1a). The total coverage at peak production was significantly affected by warming and sampling year, with an overall average of 25.1 % coverage for warming treatments, which was 2.5 % lower than without warming (27.6 % coverage; P < 0.05). Over the 6 years, total plant coverage followed the order of: 2008 (47.8 %), 2006 (37.5 %), 2010 (22.9 %), 2007 (18.2 %), 2009 (15.8 %), and 2011 (15.8 %; Table 4), which is consistent with the amount of precipitation over the growing season in each year (Fig. 1a).
Impact of weather conditions
Simple regression analysis revealed that the four main species, four functional groups and total community monthly coverage were not significantly correlated to monthly average temperature (r < 0.296, P > 0.05, n = 29). Coverage of the three main species (C. songorica, C. ammannii, and K. prostrata), functional groups, and monthly total community coverage were positively correlated to monthly precipitation (r = 0.349 to 0.547, P < 0.05, n = 29). The monthly S. breviflora coverage was not correlated to monthly precipitation (r < 0.290, n = 29). The changes in coverage induced by N addition were positively correlated to monthly precipitation for annual-biennials (r = 0.335, P < 0.05, n = 29).
Discussion
Plant species responses
The small reduction in coverage of the two main C3 species (S. breviflora and C. ammannii) caused by elevated temperature over 6 years (Table 1) supported our hypothesis that warming reduced C3 species growth. Increased coverage of two C4 species (C. songorica and K. prostrata) was reported in two previous studies in Inner Mongolian steppes (Auerswald et al. 2009; Hou et al. 2013), however, the increase in C4 species was only observed on two out of 29 sampling dates over the 6 years in this study. This insignificant warming effect on C4 species coverage was possibly because of the below-normal precipitation over the 6-year period (159–231 mm compared with the long-term average of 280 mm).
Both the increase in temperature (ground temperature increased by 1.3 °C) and low annual precipitation over the 6-year study period might have co-limited expansion of C4 plant species in the plant community. N addition did not generally enhance the coverage of either C3 or C4 plant species. This might also be attributed to the relatively low rainfall. It was observed that S. breviflora (14.6 %, C3) and C. songorica (14.9 %, C4) coverages were increased by warming only when rainfall was higher in 2008, suggesting that N effects on biomass production were limited by water availability in this desert steppe (Fig. 1).
The observed decreases in C3/C4 ratio with warming in the early and late growing seasons are consistent with previous results (Murphy and Bowman 2007; Auerswald et al. 2009), indicating that climate warming might play an important role in shifting the C3/C4 ratio in desert steppes. Although N addition had less effect on the C3/C4 ratio than precipitation, precipitation could be weakly correlated to the C3/C4 ratio. The increases in C3/C4 ratio caused by N addition were observed on two occasions that were associated with relatively higher precipitation (35.2 mm in September 2008 and 46.6 mm in June 2009), whereas decreases in the C3/C4 ratio with N addition corresponded to relatively low precipitation (18.8 mm in September 2006, 23.1 mm in September 2009 and 32 mm in September 2010, respectively, Fig. 1a). It seemed that precipitation and temperature co-limited changes in the C3/C4 ratio in the early and late growing seasons in the desert steppe. Our observations revealed the importance of precipitation in moderating the response of the C3/C4 ratio to N addition. The measured C3/C4 ratios in the early and late growing seasons from 2006 to 2010 (Table 3) suggested that the C3/C4 ratio could be used as an indicator for the response of desert steppes to global warming. These warming-induced changes in species composition were consistent with a previous study conducted in grasslands in northern China (Xu et al. 2014).
Plant functional-group responses
On four (July 2007, September 2008, and July and August 2009) out of 29 sampling dates where perennial grass coverage decreased (2–4 %; Fig. 2a), the reduction in coverage mainly reflected decreases in S. breviflora coverage (Fig. 1b). The lack of warming effects on shrubs and annual-biennials on most sampling dates, and on some sampling dates for perennial grass and forbs over the study period reflected the resilient nature of desert steppe. This observation was similar to results reported by Jiao et al. (2006). Warming had insignificant effects for 29 sampling dates on the coverage of perennial forbs and shrubs, such as C. ammannii (forb, except in September 2007) and K. prostrata (semi-shrub).
In months with relatively low precipitation (except in 2008), annual-biennials were unable to take advantage of N addition, because of insufficient soil water content which affected germination. In contrast, drought-tolerant perennial plants, such as perennial grasses and perennial forbs, became increasingly dominant with high recruitment rates (Fig. 2a, b or Table 4). These plants benefited most from N addition (Bai et al. 2010). In relatively high precipitation months (e.g., 84 mm in August 2008), N addition accelerated the expansion of annual-biennial plant species (Bai et al. 2010), which became more dominant than perennial plant functional groups (Fig. 2). This observation indicated that temporal variation in rainfall could interact with N addition to affect species composition.
Community responses
The seasonal variations in total plant community coverage observed over 6 years (2006–2011) and the positive correlation between monthly coverage and precipitation demonstrated that precipitation determined annual variation of plant community coverage in the desert steppe. Seasonal variation of four main species also showed a similar trend. Our results were in agreement with previous studies for temperate steppes in China (Xia et al. 2009; Niu et al. 2010) and elsewhere (Aires et al. 2008), in which annual mean precipitation was associated with large year-to-year fluctuations in plant community coverage (Heisler-White et al. 2008). Therefore, the observations in our study confirmed the importance of precipitation in regulating plant community coverage in desert steppes.
A small reduction (2.5 %) in plant community coverage with warming (ground temperature increased by 1.3 °C) over 6 years in the desert steppe was consistent with results reported by others for temperate grasslands (Harmens et al. 2004; Xia et al. 2009) and for arctic and subarctic ecosystems (Weltzin et al. 2003). However, contrary to a recent study on temperate grasslands (Yang et al. 2011; Walker et al. 2006; Post and Pedersen 2008), our results showed a decrease in plant community coverage linked to dramatic reduction in dominant species coverage. The observed small reductions in plant community coverage (2.5 %) with warming mainly reflected changes in S. breviflora coverage over the 6 years. Our results suggested that the plant functional group or key species was a good indicator to predict community responses to climate warming in the desert steppes where species diversity is low.
N addition had variable impact on plant community coverage in the desert steppe mainly because of variation in the amount of precipitation over the growing season. N addition led to increases in plant community coverage when precipitation was close to or above the long-term average, whereas N addition did not affect or even reduced community coverage when monthly precipitation was below the long-term average. The effects of N addition were precipitation-dependent because plant community coverage relies on the availability of water (Harpole et al. 2007). Our results were consistent with previous reports for a temperate steppe (Xia et al. 2009; Niu et al. 2010). Our results also suggested that the effect of N addition on key plant species contributed to variation in plant community coverage in the desert steppe. In summary, our data collected over 6 years partially confirmed the hypothesis that N addition would increase plant community coverage by improving soil fertility, but only when there was sufficient precipitation.
Management implications
Climate change has the potential to alter competitive interactions among plant species and dominance hierarchies because of the different responses of plant species and functional groups to warming. For the four main species assessed on 29 sampling dates over 6 years, warming reduced coverage on seven (7) sampling dates for S. breviflora, two (2) sampling dates for C. songorica and one (1) sampling date for C. ammannii, while there was no effect on K. prostrata. Warming induced marginal changes in high nutritional value herbages (i.e., S. breviflora, C. songorica and K. prostrata). The small (2.5 %) reduction in plant community coverage over the 6 years suggested that warming (temperature increased by 1.3 °C) will have minimum impact on herbage supply.
Although N is severely deficient in desert steppes, N utilization strongly depends on the amount of precipitation in the growing season. N addition was increased immensely plant community coverage in 2008 when precipitation was relatively high, however plant community coverage did not respond to N addition in other years, suggesting that supplying extra N will not necessarily increase herbage supply.
Conclusions
Warming over 6 years had no significant effect on shrubs and annual-biennials, but resulted in small reductions in perennial forbs and grasses and total plant community coverage. Our results suggested that global warming would cause a minimal reduction in plant coverage in desert steppes, as this ecosystem is very resilient. The effect of N addition on plant coverage was minimal, and small increases were observed only when monthly precipitation was above or close to the long-term average. Warming could shift C3 to C4 species in this desert steppe, as indicated by the decrease in C3/C4 ratio with warming, but confirmation of this trend requires further investigation in future studies.
References
Aires LMI, Pio CA, Pereira JS (2008) Carbon dioxide exchange above a Mediterranean C3/C4 grassland during two climatologically contrasting years. Glob Change Biol 14:539–555
Auerswald K, Wittmer MHOM, Männel TT, Bai YF, Schäufele R, Schnyder H (2009) Large regional-scale variation in C3/C4 distribution pattern of Inner Mongolia steppe is revealed by grazer wool carbon isotope composition. Biogeosci Discuss 6:545–574
Bai YF, Wu JG, Xing Q, Pan QM, Huang JH, Yang DL, Han XG (2008) Primary production and rain use efficiency across a precipitation gradient on the Mongolia plateau. Ecology 89:2140–2153
Bai Y, Wu J, Clark CM, Naeem S, Pan Q, Huang J, Zhang L, Han X (2010) Tradeoffs and thresholds in the effects of nitrogen addition on biodiversity and ecosystem functioning: evidence from inner Mongolia Grasslands. Glob Change Biol 16:358–372
Botkin DB, Saxe H, AraÚJo MB, Betts R, Bradshaw RHW, Cedhagen T, Chesson P, Dawson TP, Etterson JR, Faith DP, Ferrier S, Guisan A, Hansen AS, Hilbert DW, Loehle C, Margules C, New M, Sobel MJ, Stockwell DRB (2007) Forecasting the effects of global warming on biodiversity. Bioscience 57:227–236
Chapin FS, Shaver GR, Giblin AE, Nadelhoffer KJ, Laundre JA (1995) Responses of arctic tundra to Experimental and observed changes in climate. Ecology 76:694–711
Cleland EE, Chiariello NR, Loarie SR, Mooney HA, Field CB (2006) Diverse responses of phenology to global changes in a grassland ecosystem. Proc Natl Acad Sci 103:13740–13744
Cornelissen JHC, Van Bodegom PM, Aerts R, Callaghan TV, Van Logtestijn RSP, Alatalo J, Stuart Chapin F, Gerdol R, Gudmundsson J, Gwynn-Jones D, Hartley AE, Hik DS, Hofgaard A, Jónsdóttir IS, Karlsson S, Klein JA, Laundre J, Magnusson B, Michelsen A, Molau U, Onipchenko VG, Quested HM, Sandvik SM, Schmidt IK, Shaver GR, Solheim B, Soudzilovskaia NA, Stenström A, Tolvanen A, Totland Ø, Wada N, Welker JM, Zhao X, Team MOL (2007) Global negative vegetation feedback to climate warming responses of leaf litter decomposition rates in cold biomes. Ecol Lett 10:619–627
Craine JM, Reich PB, David Tilman G, Ellsworth D, Fargione J, Knops J, Naeem S (2003) The role of plant species in biomass production and response to elevated CO2 and N. Ecol Lett 6:623–625
Harmens H, Williams PD, Peters SL, Bambrick MT, Hopkins A, Ashenden TW (2004) Impacts of elevated atmospheric CO2 and temperature on plant community structure of a temperate grassland are modulated by cutting frequency. Grass Forage Sci 59:144–156
Harpole WS, Potts DL, Suding KN (2007) Ecosystem responses to water and nitrogen amendment in a California grassland. Glob Change Biol 13:2341–2348
Heisler-White J, Knapp A, Kelly E (2008) Increasing precipitation event size increases aboveground net primary productivity in a semi-arid grassland. Oecologia 158:129–140
Hou YH, Zhou GS, Xu ZZ, Liu T, Zhang XS (2013) Interactive effects of warming and increased precipitation on community structure and composition in an Annual forb Dominant Desert Steppe. PLoS One 8(7):e70114
Jiao SY, Han GD, Zhao ML, Liu QQ, Yang WB (2006) The effects of different stocking rates on grassland functional group characteristics and diversities in desert steppe area. J Arid Land Res Environ 20:161–165
Kardol P, Campany CE, Souza L, Norby RJ, Weltzin JF, Classen AT (2010) Climate change effects on plant biomass alter dominance patterns and community evenness in an experimental old-field ecosystem. Glob Change Biol 16:2676–2687
Knapp AK, Smith MD (2001) Variation among biomes in temporal dynamics of aboveground primary production. Science 291:481–484
Lin Y, Hong M, Han GD, Zhao ML, Chang SX (2010a) Grazing intensity affected spatial patterns of vegetation and soil fertility in a desert steppe. Agric Ecosyst Environ 138:282–292
Lin D, Xia J, Wan S (2010b) Climate warming and biomass accumulation of terrestrial plants: a meta-analysis. New Phytol 188:187–198
Luo Y (2007) Terrestrial carbon–cycle feedback to climate warming. Ann Rev Ecol Evol Syst. 38:683–712
Murphy BR, Bowman DMJS (2007) Seasonal water availability predicts the relative abundance of C3 and C4 grasses in Australia. Glob Ecol Biogeogr 16:160–169
Niu S, Wan S (2008) Warming changes plant competitive hierarchy in a temperate steppe in northern China. J Plant Ecol 1:103–110
Niu S, Liu W, Wan S (2008) Different growth responses of C3 and C4 grasses to seasonal water and nitrogen regimes and competition in a pot experiment. J Exp Bot 59:1431–1439
Niu S, Wu M, Han YI, Xia J, Zhang ZHE, Yang H, Wan S (2010) Nitrogen effects on net ecosystem carbon exchange in a temperate steppe. Glob Change Biol 16:144–155
Peñuelas J, Prieto P, Beier C, Cesaraccio C, De Angelis P, De Dato G, Emmett BA, Estiarte M, Garadnai J, Gorissen A, Láng EK, Kröel-Dulay G, Llorens L, Pellizzaro G, Riis-Nielsen T, Schmidt IK, Sirca C, Sowerby A, Spano D, Tietema A (2007) Response of plant species richness and primary productivity in shrublands along a north–south gradient in Europe to 7 years of experimental warming and drought: reductions in primary productivity in the heat and drought year of 2003. Glob Change Biol 13:2563–2581
Post E, Pedersen C (2008) Opposing plant community responses to warming with and without herbivores. Proc Natl Acad Sci 105:12353–12358
Reynolds JF, Kemp PR, Ogle K, Fernández RJ (2004) Modifying the ‘pulse–reserve’ paradigm for deserts of North America: precipitation pulses, soil water, and plant responses. Oecologia 141:194–210
Rustad LE (2008) The response of terrestrial ecosystems to global climate change: towards an integrated approach. Sci Total Environ 404:222–235
Rustad LR, Campbell JC, Marion GM, Norby RN, Mitchell MM, Hartley AH, Cornelissen JC, Gurevitch JG, Gcte-News G-N (2001) A meta-analysis of the response of soil respiration, net nitrogen mineralization, and aboveground plant growth to experimental ecosystem warming. Oecologia 126:543–562
Solomon S, Qin D, Manning M, Chen Z, Marquis M, Averyt KB, Tignor M, Miller HL (2007) IPCC, 2007: Climate change 2007: The physical science basis. In: Contribution of Working Group I to the fourth assessment report of the Intergovernmental Panel on Climate Change. Cambridge University Press, New York
Tilman D (1993) Species richness of experimental productivity gradients: How important is colonization limitation? Ecology 74:2179–2191
Van Wijk MT, Clemmensen KE, Shaver GR, Williams M, Callaghan TV, Chapin FS, Cornelissen JHC, Gough L, Hobbie SE, Jonasson S, Lee JA, Michelsen A, Press MC, Richardson SJ, Rueth H (2004) Long-term ecosystem level experiments at Toolik Lake, Alaska, and at Abisko, Northern Sweden: generalizations and differences in ecosystem and plant type responses to global change. Glob Change Biol 10:105–123
Vitousek PM, Howarth RW (1991) Nitrogen limitation on land and in the sea: How can it occur? Biogeochemistry 13:87–115
Walker MD, Wahren CH, Hollister RD, Henry GHR, Ahlquist LE, Alatalo JM, Bret-Harte MS, Calef MP, Callaghan TV, Carroll AB, Epstein HE, Jónsdóttir IS, Klein JA, Magnússon B.ó, Molau U, Oberbauer SF, Rewa SP, Robinson CH, Shaver GR, Suding KN, Thompson CC, Tolvanen A, Totland Ø, Turner PL, Tweedie CE, Webber PJ, Wookey PA (2006) Plant community responses to experimental warming across the tundra biome. Proc Natl Acad Sci USA 103:1342–1346
Wang Z, Hao X, Shan D, Han GD, Zhao ML, Willms WD, Wang Z, Ha X (2011) Influence of increasing temperature and nitrogen input on greenhouse gas emissions from a desert steppe soil in Inner Mongolia. Soil Sci Plant Nutr 57:508–518
Weltzin JF, Bridgham SD, Pastor J, Chen J, Harth C (2003) Potential effects of warming and drying on peatland plant community composition. Glob Change Biol 9:141–151
Winslow JC, Hunt ER Jr, Piper SC (2003) The influence of seasonal water availability on global C3 versus C4 grassland biomass and its implications for climate change research. Ecol Model 163:153–173
Xia J, Wan S (2008) Global response patterns of terrestrial plant species to nitrogen addition. New Phytol 179:428–439
Xia J, Niu S, Wan S (2009) Response of ecosystem carbon exchange to warming and nitrogen addition during two hydrologically contrasting growing seasons in a temperate steppe. Glob Change Biol 15:1544–1556
Xu Z, Shimizu H, Ito S, Yagasaki Y, Zou C, Zheng Y (2014) Effects of elevated CO2, warming and precipitation change on plant growth, photosynthesis and peroxidation in dominant species from North China grassland. Planta 239:421–435
Yang H, Wu M, Liu W, Zhang ZHE, Zhang N, Wan S (2011) Community structure and composition in response to climate change in a temperate steppe. Glob Change Biol 17:452–465
Yuan Z, Liu W, Niu S, Wan S (2007) Plant nitrogen dynamics and nitrogen-use strategies under altered nitrogen seasonality and competition. Ann Bot 100:821–830
Zavaleta ES, Thomas BD, Chiariello NR, Asner GP, Shaw MR, Field CB (2003) Plants reverse warming effect on ecosystem water balance. Proc Natl Acad Sci USA 100:9892–9893
Acknowledgments
We thank Xinjie Zhang, Pei Zhou, Zhongwu Wang, Qi’er An and Xiaoliang Wang for their help in field measurements, and Guogang Zhang for his helpful comments. This study was funded by the Innovative Research Team of MOE (IRT 1259), the National Natural Science Foundation of China (31270502), the National Key Basic Research Program of China (2014CB138801) and innovation project (CAAS-ASTIP-IGR 2015-05-17).
Author information
Authors and Affiliations
Corresponding author
About this article

Cite this article
Wang, Z., Li, Y., Hao, X. et al. Responses of plant community coverage to simulated warming and nitrogen addition in a desert steppe in Northern China. Ecol Res 30, 605–614 (2015). https://doi.org/10.1007/s11284-015-1265-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11284-015-1265-3