
Abstract
Simulating regional variations in gross primary production (GPP) and yields of major land cover types is complex due to differences in plant physiological properties, landscape topography, and climate gradients. In our study, we analyzed the inter-annual and inter-regional variation, as well as the effect of summer drought, on gross primary production and crop yields of 9 major land uses within the state-funded Bioenergy Region Bayreuth in Germany. We developed a simulation framework using a process based model which accounts for variations in both CO2 gas exchange, and in the case of crops, growth processes. The results indicated a severe impact of summer drought on GPP, particularly of forests and grasslands. Yields of winter crops, early planted summer grain crops as well as the perennial 2nd generation biofuel crop Silphium perfoliatum, on the other hand, were buffered despite drought by comparatively mild winter and spring temperatures. We estimated regional yield increases from SW to NE, suggesting a comparative advantage for these crops in the cooler and upland part of the region. In contrast, grasslands and annual summer crops such as maize and potato did not exhibit any apparent regional pattern in the simulations. The 2nd generation bioenergy crop exhibited significantly higher GPP and yields compared to the conventional bioenergy crop maize, suggesting that cultivation of S. perfoliatum should be increased for economic and environmental reasons, but additional study of the growth of S. perfoliatum is still required.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
While forest vegetation in temperate zones is normally the natural climax, and is still regionally dominant in areas with mountainous terrain, dynamic changes in forest cover have occurred for several thousands of years as man exploited nature to derive specific ecosystem services. Large changes in forest biomass in recent centuries and decades and their impact on global carbon balance have stimulated unprecedented research on forest ecosystem processes throughout the world (Dixon et al. 1994; Luyssaert et al. 2007; Granier et al. 2008; Luyssaert et al. 2010; Muraoka et al. 2010). Yet changes in forest cover impact man in multiple ways in addition to carbon balance and forest production, modifying the overall ecosystem services that are produced by the regional social-ecological systems (SESs) within which we live. Therefore, sustainable acquisition of ecosystem services in complex SESs, their economic gains, and their management under the influence of global change are concerns that must be analyzed (Tenhunen et al. 2009; Tenhunen 2013).
With respect to modification of land use in Germany, ambitious goals to increase the use of renewable energies have led to doubling of the use of non-food biomass. Within only a 2-year period from 2004 to 2006, the production of biogas, dominantly obtained from maize, increased tenfold, induced by tax exemptions for biofuels and feed-in tariffs for power production by energy plants (Bringezu et al. 2009). Meanwhile, the indicated land use change for energy cropping competes with forest and conventional agriculture food production. In order to avoid land use conflicts, bioenergy crop production should best take place on land not required for food and fiber (Fritsche et al. 2010). Fritsche et al. (2010) stated that biomass is the “stuff of life” on this planet and that changes in biomass production, e.g. the replacement of natural vegetation with crop cultivation could have either positive or negative impacts on ecosystem services, carbon balances and via food chains also on human livelihoods. Therefore, a sustainable bioenergy production from perennials grown on prior degraded croplands or marginal lands and from waste biomass is a priority consideration that requires scientific evaluation. Use of perennials on marginal lands would minimize habitat destruction, competition with food production and carbon debts, all of which are associated with direct and indirect land clearing for biofuel production (Fargione et al. 2008).
In this context, CO2 flux studies, which are critical to understanding carbon-based services in forested landscapes and regions, should integrate and consider simultaneously the behavior of all major landscape elements which determine the performance of the social-ecological-system in question. In particular, regional scale analysis is needed in order to focus on real acquired data and integrative social-ecological measures, and on specific trade-offs and compromises that are dependent on local ecosystem and land use characteristics. To integrate point measurements, such as eddy-tower observations and biomass measurements, and to examine these with respect to variations in climate, process-based models are needed. Their application, however, often remains challenging due to the lack of coordinated regional data bases for driver variables and detailed land use maps.
In our study, we describe a process-based simulation modeling framework that allows us to examine ecosystem services provided by nine dominant landscape elements in 42 communities in the Bioenergy Region Bayreuth over a time period of 10 years (2003–2012). In detail, the goal of our study is to determine time and spatial variations in annual (i) gross carbon uptake (GPP) and (ii) total usable biomass yield of grasslands, agricultural crops such as potatoes, summer and winter grains and winter rape, as well as the 1st and 2nd generation bioenergy crops maize vs. the perennial S. perfoliatum, and (iii) to analyze the impact of the summer drought 2003 in Germany on agricultural production and yields. Additionally, we estimate variations in the GPP of coniferous and deciduous forests which are major land cover types in the region, but where variations in biomass accumulation on an annual basis are beyond the scope of the current analysis. In the case of forests, the values of integrated GPP are viewed as an indicator of relative changes in production. This study provides one basis from which we can begin to examine economically and environmentally sustainable land use distribution in the region and adaptation strategies with respect to climate change and weather extremes.
Materials and methods
Site description
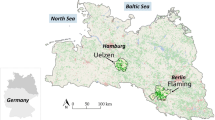
The Bioenergy Region Bayreuth is one of 25 regions in Germany, which were selected in a competition by the German Federal Ministry of Food, Agriculture and Consumer Protection (BMELV) to be supported in implementation of their regional development plans for bioenergy production. The Bioenergy Region Bayreuth represents an area of 154,455 hectare and is divided into 42 communities (Fig. 1). The altitude in the region ranges from 336 masl in Igensdorf (49°37′20.0″N, 11°13′51.7″E) located in the southwestern part of the Bioenergy Region up to 1,024 masl at the mountain Ochsenkopf in the Fichtelgebirge (50°01′50.5″N, 11°48′27.1″E) in the northeast.

Topography of the Bioenergy Region Bayreuth with 42 communities
The climate conditions in the region depend highly on topography. At Ochsenkopf, the average annual temperature is 6.4 °C and the mean annual precipitation is 1,210 mm. Accordingly, the southwestern part is characterized by higher temperatures and lower amounts of precipitation. In Weissenohe in the extreme southwest, the mean annual temperature and the mean annual precipitation sum are 8.4 °C and 665 mm, respectively. In 2003, the minimum and maximum air temperature in June and August ranged more than 4 °C above the average base line, while precipitation was low from May–September with particularly low rates in June and August (Rebetez et al. 2006). In our study, we used hourly climate data from 2003–2012 of six weather stations provided by the agrometeorological weather station network of the Bavarian State Research Centre for Agriculture (LfL), a weather station of the botanical garden (ÖBG) in Bayreuth (49°55′45″N, 11°35′10″E) as well as a weather station of the University of Bayreuth located in Bayreuth Waldstein/Weidenbrunnen (50°08′31.2″N, 11°52′00.8″E) (http://fluxnet.ornl.gov/site/405). We interpolated the climate variables global radiation, air temperature, relative humidity, wind speed and precipitation using bilinear interpolation method (triangulation) based on a raster map with a resolution of 100 × 100 m to obtain individual climate data sets for all 42 communities.
The soils in the region are classified according to the national soil classification system. In the Bayreuth region, the dominant soil types are Podzolic Cambisols and Cambisols developed on mesozoic/tertiary sands and loams/clay, respectively. In the southwestern part of the region the dominant soil types are Leptosols and Cambisols developed on calcareous rocks and marls. Soils such as Cambisols and Podsolic-Cambisols in the northeastern part of the region are mostly acidic due to its granitic parent rock material. Furthermore, and in dependence on base saturation Luvisols or Alisols, Cambisol, stagnic Cambisols and Stagnosols develop on residual loess. On agricultural field sites, agricultural management activities such as tillage and plowing often modified the natural soil structure. The plowing pan functions as a sharp interface between topsoil and underlying subsoil.
The land use of the region is characterized by a patchy distribution of agricultural fields and forest stands accounting for 45 and 43 % of the land cover, respectively. Table 1 provides an overview about the land use in the region. The agricultural area of ca. 70,000 ha is 53 % in agricultural production and 35 % in permanent pasture. Only 8 % of the area in agricultural production is currently used for bioenergy crop cultivation (Bavarian State Agency for Statistics and Data Processing 2012).
Simulation of gas exchange and annual plant growth
Model characteristics
The model PIXGRO consists of two coupled modules, the canopy flux module PROXELNEE (PROcess pixel net ecosystem exchange model) and vegetation structure module CGRO (Crop GROwth). The module PROXELNEE captures canopy processes such as gross photosynthesis (GPP), ecosystem respiration (Reco), net ecosystem CO2 exchange (NEE) and transpiration. The simulation of gross photosynthesis (GPP) is implemented in module PROXELNEE using algorithms of Farquhar and Caemmerer (1982) with modifications from Harley and Tenhunen (1991). CGRO simulates growth and development processes e.g. leaf area index (LAI) and differentiated biomass development of different plant compartments such as leaves, stems, roots and grains according to five growth stages. The simulated LAI from CGRO is passed to the PROXELNEE canopy process module and the computed fixed C fluxes are returned to CGRO, which then simulates crop growth. Dry matter accumulation rate is simulated from the hourly gross photosynthesis, Pgros (molCO2 m−2 h−1) after conversion to gross carbohydrate production rate, Pg (gCH2O m−2 h−1) and the latter reduced by plant respiration losses (Adiku et al. 2006; see detailed description in the supplementary materials). Until the end of vegetative growth, the net hourly growth is partitioned to leaves, stems, roots, and grains. The values of the partitioning coefficients change with the stage of the crop development. For instance, in the second stage (development stage) biomass partitioning coefficients for leaves are higher compared to biomass partitioning coefficients for grains. Thereafter, biomass produced is partitioned mainly to reproductive structures, e.g. to the grain or storage organ (cf. potato). Detailed descriptions of the PIXGRO model algorithms and the linkage between PROXELNEE and CGRO modules are provided in the supplementary material. The parameter settings utilized in the current application are described below after explanation of the model calibration procedures.
Model calibration for different land uses
In our modeling study, the models for varying land-uses were first calibrated against GPP flux observations of eddy covariance tower sites in Germany, France and Belgium obtained from the FLUXNET (http://www.fluxnet.ornl.gov/) data base. The climate characteristics as well as the vegetation cover of the selected tower sites are given in Table 2. Both, the coniferous and deciduous forest models were calibrated using the eddy-flux datasets of the year 2002 and were validated with the GPP flux observation datasets of the years 2003–2005. For grasslands, we used eddy covariance datasets of 2003 for calibration and validated the model with the dataset of the following year. For the agricultural crops, GPP datasets were only available for a single year, namely winter rape (2005), winter grain (2006), potato (2006) and maize (2007). For summer grain and S. perfoliatum, eddy covariance tower sites and, thus, GPP data sets easily related to conditions in our region were not available, so that these land use categories were calibrated only against biomass measurements and annual yield statistics (Table 4).
If data was available, we additionally calibrated the models against leaf area index (LAI), biomass measurements and yields. Datasets of LAI were only available for the land use categories grassland, maize and potato. For potato, aboveground and root biomass as well as tuber yield was also available. For maize, we calibrated the model against root and aboveground biomass. Grain yield was available for summer and winter grain as well as for winter rape. Table 3 provides an overview of the available datasets which were included into the calibration procedure.
The final calibration was carried out using annual yield statistics from the Bioenergy Region Bayreuth (Table 4). After calibrating the GPP fluxes, LAI, biomass and yields for varying land uses at eddy sites, all models for the different land cover types were run with the weather data of the years 2003 and 2011 from Bayreuth/Mistelbach, and checked against the observed yields according to data from experiments conducted by the Amt für Ernährung, Landwirtschaft und Forsten during these years (Dr. Friedrich Asen, personal communication). If the total yields differed significantly from the annual yield statistics, biomass partitioning coefficients were adjusted to obtain agreement.
The 10 years chosen allowed us additionally to adjust crop response for the influence of severe summer drought which occurred in 2003 across Europe. In the model, the effective leaf area in carbon uptake (GPP) is proportionally decreased in response to soil matric potential beyond a critical threshold. Reduction of the effective leaf area reflects the decreases in canopy conductance for CO2 that result from stomatal closure. Based on the GPP observations in 2003 vs. other years, a linear relationship of effective leaf area to soil matric potential was adjusted for all land uses. Plant response to drought was finally calibrated to obtain the ratio of the yields found for Bayreuth/Mistelbach in 2003 and 2011. The calibrated key parameters of PIXGRO modules CGRO (Table SM1) and PROXELNEE (Table SM2) as used in the simulations are provided in the supplementary material.
Defining the growth season across the Bioenergy Region Bayreuth
Temperature is one of the most important factors governing plant growth (Neitsch et al. 2009). The concept of heat units are typically used to predict growth and development of crops and have been successfully employed in order to develop planting schedules for horticultural crops (Brown 1989). Each plant has its individual heat requirements for optimal growth and a mean daily temperature has to exceed the base temperature in order to induce growth. According to (Brown 1989) heat unit accumulation for a given day is calculated by
where HU = accumulated heat unit on a given day, Tav = mean daily temperature (°C) and Tbase = base temperature (°C). Indirectly, the planting carried out by farmers is also related to heat sums in their subjective evaluation of when it is appropriate to seed their fields.
In this study, we used the heat sum concept to determine a start of the growth season for each of the 42 communities of the Bioenergy Region Bayreuth. In order to do so, we calculated HU at the beginning of the season at the calibration sites for all land uses except for coniferous forests. The season start for coniferous forest with evergreen needle species was set constant to DOY 120 for all years. For deciduous forests the HU concept was applied with a base temperature of 5 °C (Diekmann 1996). A base temperature of 0 °C was used for winter grain, winter rape and spring barley (Kiniry et al. 1995), while for grassland a base temperature of 4 °C was assumed (Hutchinson et al. 2000). For potato and maize, we used a base temperature of 7 and 8 °C, respectively (Kiniry et al. 1995; Hackett and Carolane 1982). The base temperature for S. perfoliatum was set to 5 °C (Titei et al. 2013). Secondly, we calculated HU for grassland and agricultural crops for each land cover and each year (2003–2012) using climate data specific for the 42 communities indicated in Fig. 1. Thus, we determined the day, when the heat sum corresponded to the heat sum calculated for planting at the calibration sites. This approach allowed us to implement individual season starts for all agricultural crops and grassland in 42 communities and specific for each of 10 years. The variation in the beginning of the growing season for maize in the years 2003 and 2010 within the Bioenergy Region Bayreuth is shown as an example in Fig. 2. Generally, the time of season start was related to the temperature gradient from southwest to northeast within the region.

Simulated regional variation in season start (DOY = day of the year) of maize cultivation for the years a 2003 and b 2010
Statistical analysis
Statistical analysis was carried out using the statistical software package R (R Core Team 2013) in order to identify significant drought effects. The simulated annual GPP and the total yields from 2003 to 2012 for all land use types were tested for normal distribution and for homogeneity of variance using the Shapiro–Wilk-test and Levene test, respectively. Afterwards, the Kruskal–Wallis-test was applied in order to identify differences in mean within all groups. Finally, we used the paired Wilcoxon-signed rank test (95 % confidence interval) as a post hoc test to determine whether the annual GPP and the total yields are significantly different between the drought year 2003 and the other years.
Results
Model calibration and validation
In Fig. 3 the annual course of observed and simulated GPP fluxes for all land use types except for S. perfoliatum and summer grain are shown. In general, the agreement between observed and simulated GPP fluxes for coniferous and deciduous forests was satisfying for the calibration year 2002. With regard to validation, the agreement of observed and simulated carbon uptake of deciduous and coniferous forests for the years 2003–2005 was satisfying with R 2 > 0.65 except for the broadleaf forest for the year 2004 (R 2 = 0.58). The validation results are fully shown in the supplementary material (Figs. 1). The GPP fluxes of deciduous forest in 2004 decreased during late summer significantly because of forest thinning (Granier et al. 2008). This management practice resulted in an overestimation of GPP fluxes during summer in comparison to the observed GPP. The calibration and validation models of grassland showed also a good agreement of GPP fluxes in comparison to the observations, although the simulated GPP fluxes in both years were slightly overestimated by the model. The GPP fluxes of the agricultural crops maize, potato and winter grain were accurately predicted and resulted in a very good agreement (R 2 > 0.88). However, the agreement between simulated and observed GPP fluxes for winter rape was low (R 2 = 0.56) in comparison to all other land uses since the simulated fluxes were overestimated in the later part of the growing season.

Simulated vs. observed GPP fluxes of a deciduous forest 2002/calibration, b deciduous forest 2003/validation, c coniferous forest 2002/calibration, d coniferous forest 2003/validation, e grassland 2003/calibration f grassland 2004/validation, g maize 2007/calibration, h potato 2006/calibration, i wintergrain 2006/calibration, and j winter rape 2005/calibration
The simulated leaf area index (LAI) was also compared to observed LAI but only for grassland, maize and potato (Fig. 4 ). For both years 2003 and 2004, the simulated LAI of grassland was consistent with the observations. The crop model of maize delivered also a good agreement although the observed LAI declined earlier compared to the simulated LAI. The comparison of the simulated and observed LAI of potato showed the expected trends but was less satisfying (R 2 = 0.25). Time-sequence LAI observations of S. perfoliatum were not available, thus the model was only adjusted to the observed LAI of 4.77 (DOY 193). In comparison, the simulated LAI on DOY 193 was 4.84.

Simulated vs. observed leaf area index (LAI) of a grassland 2003/calibration, b grassland 2004/validation, maize 2007/calibration, and d potato 2006/calibration
The PIXGRO model was calibrated using yield data of an experimental plot planted with S. perfoliatum in Bayreuth in 2012. In this year, the yield of the experimental sites was 18.3 t ha−1, while the simulated yield was 18.4 t ha−1 when using weather data of Mistelbach near Bayreuth (Fig. 5). The yield of the agricultural crops winter grain and winter rape was slightly overestimated by the model, while maize and potato was underestimated. However, generally the agreement between simulated and observed yields was satisfying as shown in Fig. 5. The model calibrated for grassland largely overestimated yield biomass. However, the yield ratio shown by the annual statistics (Table 4) as well as by the measured yields in Grillenburg corresponded well to the simulated yield ratio of the years 2003 and 2004. Additionally, however, it should be realized that the direct comparison of yields between both years is not feasible due to differences in total number of harvests and harvest timing. In 2003 e.g. grasslands were harvested only twice on DOY 167 and 287, while during 2004 the grassland site was harvested three times (DOY 177, 217, 300). The PIXGRO model was calibrated for summer grain crops using only annual yield statistics (Table 4), since no eddy flux observations and LAI data from Central Europe was available for this land use type. Therefore, we used the plant physiological parameter set of winter grain but shifted the season start to April. Subsequently, we simulated summer grain using weather data of 2003 and 2011 in Mistelbach. At the end of July (DOY = 212) the simulated yield for the years 2003 and 2011 was 5.93 and 5.97 t ha−1, respectively. In comparison to the annual yield statistics of the region (Table 4) the simulated yields matched very well.

Simulated vs. observed total yields of a1 grassland 2003, a2 grassland 2004, b potato, c winter rape, d winter grain, e maize, f S. perfoliatum
Gross primary production (GPP) of forests, agricultural and bioenergy crops across the Bioenergy Region Bayreuth
Drought effects and interannual variability of GPP
Forests and grassland
The distribution of simulated annual GPP for all land use types within 10 years are summarized in Fig. 6. For both, coniferous and deciduous forest the maximum simulated annual GPP was reached in the year 2007. In contrast, the severe drought in summer 2003 affected annual GPP of both, coniferous and deciduous forest by decreasing GPP to a 10 years minimum. In the comparison of the annual GPP of 2003 and all other years, both forest types showed significantly lower carbon uptake (paired Wilcoxon test, p < 0.001) in 2003. In agreement with the carbon uptake of forests, grasslands also showed a high interannual variation of annual GPP with the highest carbon uptake rate in 2007 as well as significantly lower annual GPP in the drought year 2003 compared to other years (paired Wilcoxon test, p < 0.001).

Simulated interannual variation of GPP from 2003 to 2012 (n = 42 communities) for a coniferous forest, b deciduous forest, c grassland, d potato, e summer grain, f winter grain, g winter rape, h maize, i S. perfoliatum. Note that y-scales vary between land uses
Agricultural crops
The group of agricultural crops, namely potato, winter and summer grain, and winter rape, showed similar annual carbon uptake rates during the time period 2003–2012 with average annual GPP sums ranging from 624 to 934 gC m−2 year−1. Only winter rape showed noticeable higher carbon uptake ranging from approx. 1,260 to 1,990 gC m−2 year−1. The interannual comparison showed further that for grain crops, highest GPP was reached in 2004, while for the root crop potato the highest carbon uptake was simulated in 2009. The summer drought 2003 did not affect carbon uptake rates of agricultural crops by decreasing it to a minimum. Instead, the simulation years 2009 and 2010 showed lowest simulated carbon uptake for grain crops and root crop, respectively. Indeed, the annual GPP of agricultural crops in 2003 was significantly different (paired Wilcoxon test, p < 0.05) compared to the majority of other years, but this was attributed to the overall high interannual variability of GPP.
Bioenergy crops
The annual carbon uptake of the bioenergy crop maize ranged from approx. 500 to 1,000 gC m−2 year−1. In comparison, the 2nd generation bioenergy crop S. perfoliatum showed remarkably high annual carbon uptake ranging from approx. 1,000 to 1,900 gC m−2 year−1. Similar to the group of grain crops, the minimum of annual GPP by the perennial S. perfoliatum was reached in 2009. In contrast, maize showed its annual minimum GPP similar to the root crop potato in the year 2010. The annual GPP of maize in the summer drought year 2003 was significantly different to all other years (paired Wilcoxon test, p < 0.001) except for 2006. However, and similar to the agricultural crops, the interannual comparison showed an average level for GPP in 2003. In comparison, the annual GPP of the 2nd generation bioenergy crop S. perfoliatum, did not significantly differ between the drought year 2003, in 2006 or in 2010 (paired Wilcoxon test, p > 0.24). Except for the year 2009, in which the annual GPP was significantly lower compared to the drought year 2003 (paired Wilcoxon test, p < 0.001), the annual GPP in the remaining years were significantly higher in comparison to 2003 (paired Wilcoxon test, p < 0.01). In comparison to maize, the interannual variability of GPP by S. perfoliatum was strikingly higher, perhaps due to its long growth period and the influences of climate during spring, summer and fall.
Interregional comparison of 10 years average GPP
In Fig. 7, maps of 10-year average annual carbon uptake for the dominant land use types within the region are shown. While the deciduous forest showed a clear GPP gradient from NE to SW, the lowest average annual GPP of coniferous forest was located in the central part of the region. The interregional comparison of average annual GPP of grassland, potato, and maize followed also the NE-SW gradient similar to deciduous forest with lower carbon uptake in northeastern part of the region and highest uptake rates in the southwestern region. In contrast, all crops which are planted in the previous year, namely winter grain, winter rape, the perennial crop S. perfoliatum as well as the early planted crop summer grain showed a gradient reversal with lower average annual GPP in the southwest and highest carbon uptake in the regions of high altitudes (NE). The average GPP of deciduous forests, grassland, potato and maize was noticeably lower in the community Pottenstein (located centrally within the southwestern region) in contrast to its surrounding communities. Similarly, the community Warmensteinach (located centrally within the outmost northeastern region) showed low annual GPP of winter and summer grain as well as for winter rape in comparison to its surrounding communities.

Simulated 10-year average annual GPP (gC m−2 year−1) for a coniferous forest, b deciduous forest, c grassland, d potato, e summer grain, f winter grain, g winter rape, h maize, i S. perfoliatum in the Bioenergy Region Bayreuth
Total yields of grasslands, agricultural and bioenergy crops
Drought effects and interannual comparison of yields
The interannual comparison of yield boxplots is shown in Fig. 8. In general, annual total yields correlated positively to annual GPP regardless for all agricultural and bioenergy crops. A detailed overview about the relationship between GPP fluxes and yields for all crop types are given in Fig. 3 in the supplementary material.

Distribution of total yields from 2003 to 2012 in 42 communities for a grassland, b potato, c summer grain, d winter grain, e winter rape, f maize, and g S. perfoliatum
Grasslands
The total yield for grassland varied between approx. 4.8 and 8 t ha−1 year−1 within the time period of 2003–2012. As it was shown in the interannual comparison of GPP, the drought year 2003 also led to significantly lower grassland yields compared to all other years (paired Wilcoxon test, p < 0.001). Accordingly, the highest yield was simulated in the year 2007. Both results are in accordance to the annual yield statistics (Table 4), which also showed lowest yields in 2003 and relatively high yields in 2007. The simulated total grassland yields in all other years were comparable as seen by a low interannual variability.
Agricultural crops
The total yield of potato ranged between 5.9 and 9.8 t ha−1 year−1 (2003–2012). As in the case of grassland, the total yield of potatoes was significantly lower in the drought year 2003 compared to all other years (paired Wilcoxon test, p < 0.001). In principle, the potato simulations showed relatively high interannual variability of total yields with highest yields in the years 2009 and 2011. For summer and winter grain, the range of total yield was approx. from 4.6 to 7.5 t ha−1 year−1 and 3.7 to 6.3 t ha−1 year−1, respectively. The simulated total yields of winter rape ranged from 4.4 to 8 t ha−1 year−1. Similarly to the annual GPP, the lowest total yield of grain crops was not due to the summer drought 2003 but was simulated for the years 2008 and 2009.
Bioenergy crops
Within the time period of 10 years, the range of total yields of maize and S. perfoliatum was 9.8 to 15.4 t ha−1 year−1 and 12.7 to 23.3 t ha−1 year−1, respectively. For maize crop, the total yield was significantly lower in the drought year 2003 compared to the other years (paired Wilcoxon test, p < 0.001) except for 2010 (paired Wilcoxon, p = 0.17). The low maize yield in in 2003 and 2010 was also shown by the annual yield statistics (Table 4). The total yield of the 2nd generation biofuel crop S. perfoliatum in 2009 was significantly lower compared to the drought year 2003 but in comparison to all other years, the yield in 2003 was still low. In general, the interannual variability of total yields was high for both, maize and S. perfoliatum.
Interregional comparison of 10 years average yields
Figure 9 gives an overview of the regional differences in 10-year average total yields for grassland, agricultural and bioenergy crops. The regional difference of grassland yield within the region was 1 t ha−1 year−1 (6.2 min.; 7.2 max. yields). Considering the maximum yield of 7.2 t ha−1 year−1 as 100 %, the minimum yield corresponds to 14 % less grassland yield. Accordingly, the average regional difference of potato and summer grain yields was 1.5 and 1.2 t ha−1 year−1, which corresponds to 17 and 19 % fewer yields in comparison to the maximum total yield. Yields of both, winter grain and winter rape showed a regional difference of 1.1 t ha−1 year−1, which is equal to only 80 and 85 % of the maximum total yield, respectively. Furthermore, the interregional difference in yields of the bioenergy crops maize and S. perfoliatum was 2.9 and 3.6 t ha−1 year−1. These differences correspond to 20 and 18 % fewer yields compared to the maximum yield of the respective crop.

Simulated 10-year average total yield (t ha−1 year−1) for a grassland, b potato, c summer grain, d winter grain, e winter rape, f maize, g S. perfoliatum in the Bioenergy Region Bayreuth
As it was shown in the interregional variation of GPP (Fig. 7), both communities Pottenstein and Warmensteinach simultaneously show invariable low yields compared to their surrounding communities (Fig. 9). The regional pattern of average annual GPP of grassland, potato and maize suggested a similar pattern of total yields within the region following a NE-SW gradient with relatively low yields in NE and relatively high yield in SW. However, such yield gradient could not be identified for these three crop types (Fig. 9). Moreover, no obvious regional pattern in yields was discernible. In contrast, the perennial crop S. perfoliatum, winter grain and winter rape as well as the early planted summer grain showed the gradient with low yields in SW and high yields in the mountainous areas in NE. This gradient was comparable with the gradient already observed in average annual GPP.
Discussion
Methodology
Levy et al. (1999) stated that the regional interpolation of point eddy-covariance measurement is extremely unreliable due to heterogeneity of the landscapes and nonlinearity inherent in ecophysiological processes. In our approach we did not scale up from eddy covariance measurements to regional scale in the usual sense by employing a raster-based regional landscape model, but used one-dimensional models to simulate GPP and yields for each individual community, varying plant physiological and plant growth parameters in combination with different season start dates and different weather input for each community. This modeling procedure allowed us to derive regionally dependent GPP and yields for 9 different land uses over 10 years. Although both, GPP and yields will not change abruptly as shown for community borders in mapped outputs and hydrological coupling is not addressed (the model application is a one-dimensional vertical formulation), the modeling approach provides a new perspective, since changes in specific ecosystem services are obtained that can be related to agricultural economic statistical data and analyses.
Care is needed to specifically simulate plant response across a region. Our study is a methodological attempt to do so, although some model limitations arise in terms of calibrating at stand level and using these parameter sets for a regional modeling approach, especially in the dimension of GPP, LAI, carbon allocation and yields. For instance, acclimation response of the individual species, soil heterogeneity as well as agricultural management e.g. conventional vs. organic fertilization influences the CO2 gas exchange and biomass production within the region. These factors had to be neglected or simplified because little is known about acclimation responses of individual species as well as small-scale climate gradients and soil heterogeneity. Thus, the criticism of Levy et al. (1999) that nonlinearity in ecophysiological processes remains a problem for regional extrapolations cannot be denied. The results of the modeling effort must be understood with this reservation. However, the PIXGRO application across the Bioenergy region Bayreuth provides a valuable tool that is sensitive to climate, especially drought as far as calibration data for drought response of prominent vegetation is available.
The potential of using the model output datasets to determine yields and to estimate risks of specific land use configurations in terms of drought impacts will support the regional economic evaluation of production, farmers’ crops choice and land use decisions. To determine whether a certain land use for forests, agriculture and bioenergy plants within a region is economically profitable and environmentally sustainable in the context of different adaption strategies to climate change will be a focus of our future research by applying specific IPCC climate change scenarios and a regional economic evaluation of adaption measures. Using natural science crop modeling approaches together with economic analysis can be useful for similar attempts at other locations (Nguyen and Tenhunen 2013).
Interannual and interregional comparison of GPP and yields
The comparison of simulated GPP and yields over a period of 10 years showed that both are highly variable for all land uses. The simulated interannual and interregional variability was due to the interplay of weather conditions, especially temperature and rainfall as well as differences in the beginning of the season. For example, in 2009 the high average monthly temperatures in April and May supported growth conditions for summer crops such as potato and maize and resulted in highest yields. Furthermore, mild temperatures in February 2004 supported high yields of winter and summer grain, winter rape and the perennial crop S. perfoliatum (Fig. 2). Affected by such weather conditions, the heat sum approach led to early season starts of winter and summer grain, winter rape and the perennial S. perfoliatum due to their low base temperatures which in turn automatically extended the growing season, thus the time to allocate carbon into individual plant parts was extended. This was in accordance to Mendham et al. (1981) who found high yields of winter oil-seed rape in dependence on mild temperatures during spring season. Since climate change scenarios forecast higher temperatures in winter and spring in future, winter annual and perennial crops species are likely to have an advantage (Olesen and Bindi 2002). However, Rötter and van de Geijn (1999) reported that e.g. higher temperatures in winter are likely to regularly exceed optimal temperatures for growth and development, and that vernalization could be reversed, which poses a risk for crop damages. Additionally, for both winter wheat and spring barley, a temperature increase in winter may lead to damages caused by pests and phytopathogens (Olesen et al. 2011). These feedback mechanisms with regard to increased temperatures could not be captured by our model simulations.
Generally, an increase in temperature and CO2 due to climate change has been predicted to be beneficial for forests, grasslands and annual summer crop because of a prolongation of the growing season, enhanced photosynthesis rates and spatially northward expansion of suitable areas for crop cultivation (Olesen and Bindi 2002; Luyssaert et al. 2007), however, the vulnerability of these ecosystems is increased due to a higher frequency of summer droughts followed by heavy rain events (Solomon et al. 2007). In our simulations, forests and grasslands as well as annual summer crop cultivations of potatoes and maize were mostly affected by low water availability during summer drought 2003, showing significantly lower GPP and yields compared to other years. This was also reported by Hussain et al. (2011) and Ciais et al. (2005), who found significantly reduced productivity of forests and grasslands in Europe and Germany turning these ecosystems into carbon sources under summer drought conditions. Furthermore, Rouault et al. (2006) reported that drought had indeed directs effects on tree physiology and growth in western European forests, but similar to winter crops the most important impact is caused by secondary factors such as insects, pathogens and fires.
1st and 2nd generation bioenergy crop cultivations
To support sustainable bioenergy production, it is necessary to investigate the potential of 2nd generation bioenergy plants in order to minimize ecological impacts such as the attrition of organic matter, soil erosion and compaction and subsequently nutrient leaching and eutrophication of ground and surface water resources which is likely to occur for instance in widespread annual bioenergy crop cultivation systems such as maize (Vadas et al. 2008). S. perfoliatum was identified to have several advantages from an ecological point of view compared to maize since it is cold-resistant; fast-growing and cultivated as a permanent crop that makes the annual soil tillage redundant and covers soils during winter season. In our study, the 2nd generation bioenergy crop S. perfoliatum showed much higher GPP and yields compared to commonly grown bioenergy crop maize. Hence, to promote sustainable bioenergy production perennials such as S. perfoliatum can either potentially substitute 1st generation bioenergy plants like maize or can be grown on marginal lands to avoid land competition between 1st generation bioenergy crops or food crops (Kang et al. 2013). Indeed, the estimation of GPP and yields of S. perfoliatum in our study is limited in validity because of lacking extensive calibration data for bioenergy crops. However, although the GPP and yield estimation might be overestimated by the model, cultivating permanent crops for bioenergy production are beneficial to reduce ecological impacts on soils and water resources. Generally only few eddy covariance studies on bioenergy cultivations exist (Shurpali et al. 2009; Zenone et al. 2011) demonstrating the need of future research in order to derive reliable estimates for simulating GPP and yields of 2nd generation bioenergy cultivations. Additionally, future field studies should address the sensitivity of production to annual climate variations as well as the suitability of marginal land as habitats for S. perfoliatum.
Conclusion
Although elevated CO2 concentrations and increasing temperatures enhance net photosynthesis and expand the growth period of forests, grasslands and annual summer crop cultivations, the impact of summer drought events, which are expected to increase in frequency under the influence of climate change, are likely to affect gross primary production and yields. Furthermore, our simulations demonstrate that mild winter and spring temperatures support increased yields of winter crops, early planted summer grain and perennial crops such as S. perfoliatum. Since future climate scenarios forecast temperature increases in the winter season, winter annuals and the perennial crop S. perfoliatum may benefit from changing climate, if we disregard secondary effects, such as pest outbreaks.
Except for coniferous forest, the GPP of deciduous forest, grassland and summer crop cultivations increased along a regional gradient from northeast to southwest, while the GPP of winter crops such as winter grain, winter rape, and early planted summer grain crops showed a gradient reversal from southwest to northeast. Yield outputs of grasslands and summer crops, however, did not show a clear regional pattern which makes it difficult to develop land use recommendations. From a regional point of view, our study suggests to give greater preference to the cultivation of winter crops, early planted summer grain crops as well as the perennial S. perfoliatum for bioenergy production in the colder and mountainous northeastern part of the region than to annually planted summer crops such as maize or potatoes.
Finally, our simulations suggest that bioenergy crop production accomplished by planting the perennial crop S. perfoliatum will likely have environmental and economic benefits due to promising plant species characteristics such as cold-resistance, rapid growth in early spring and overall higher yield when compared to the conventional annual maize cultivations. However, future investigations of CO2 gas exchange using eddy-covariance measurements and of growth of S. perfoliatum are necessary in order to underpin these conclusions with respect to its high potential for bioenergy production.
The described up-scaling approach provides a new perspective in the context of understanding ecosystem services at regional scale. First, it illustrates that the influences of regional gradients in habitat and climate conditions may be simultaneously and systematically analyzed for multiple ecosystem types that play a role in potential land cover change. It also demonstrates the need for information from different disciplines in order to support and accomplish such analyses. Finally, it provides insight with respect to the limitations that exist in our understanding at relatively local scales and in our ability to contribute to management, i.e., broader concepts are required in the planning of comparative production studies, in considering various environmental impacts, in quantifying plant growth, and in regionally building data bases for driver variables of carbon gain in order to support the construction and verification of models. If this can be accomplished, we can improve predictions by stepwise linking of production processes, expanding our view to quantify not only crop yields but regional carbon and water balances as well, and by considering a broad palette of climate scenarios. In this case, system internal feedbacks at regional scale can also be addressed.
References
Adiku S, Reichstein M, Lohila A, Dinh NQ, Aurela M, Laurila T, Lueers J, Tenhunen JD (2006) PIXGRO: a model for simulating the ecosystem CO2 exchange and growth of spring barley. Ecol Model 190:260–276
Bavarian State Agency for Statistics and Data Processing (2012) In: Regionalentwicklungskonzept der Bioenergieregion Bayreuth, Fortschreibung 2012–2015. Regionalmanagement Stadt und Landkreis Bayreuth GbR, Bayreuth, pp 3–17
Bringezu S, Schütz H, Arnold K, Merten F, Kabasci S, Borelbach P, Michels C, Reinhardt GA, Rettenmaier N (2009) Global implications of biomass and biofuel use in Germany—recent trends and future scenarios for domestic and foreign agricultural land use and resulting GHG emissions. J Clean Prod 17:S57–S68
Brown PW (1989) Cooperative extension 8915. University of Arizona, Heat units
Ciais P, Reichstein M, Viovy N, Granier A, Ogee J, Allard V, Aubinet M, Buchmann N, Bernhofer C, Carrara A, Chevallier F, de Noblet N, Friend AD, Friedlingstein P, Grunwald T, Heinesch B, Keronen P, Knohl A, Krinner G, Loustau D, Manca G, Matteucci G, Miglietta F, Ourcival JM, Papale D, Pilegaard K, Rambal S, Seufert G, Soussana JF, Sanz MJ, Schulze ED, Vesala T, Valentini R (2005) Europe-wide reduction in primary productivity caused by the heat and drought in 2003. Nature 437:529–533
Core Team R R (2013) A language and environment for statistical computing. R Foundation for statistical computing, Vienna
Diekmann M (1996) Relationship between flowering phenology of perennial herbs and meteorological data in deciduous forests of Sweden. Can J Bot 74:528–537
Dixon RK, Solomon AM, Brown S, Houghton RA, Trexier MC, Wisniewski J (1994) Carbon pools and flux of global forest ecosystems. Science 263:185–190
Fargione J, Hill J, Tilman D, Polasky S, Hawthorne P (2008) Land clearing and the biofuel carbon debt. Science 319:1235–1238
Farquhar GD, Caemmerer S (1982) Modelling of photosynthetic response to environmental conditions. In: Lange OL, Nobel PS, Osmond CB, Ziegler H (eds) Physiological plant ecology II, 12/B. Springer, Berlin Heidelberg, pp 549–587
Fritsche UR, Sims REH, Monti A (2010) Direct and indirect land-use competition issues for energy crops and their sustainable production—an overview. Biofuels Bioprod Bioref 4:692–704
Granier A, Bréda N, Longdoz B, Gross P, Ngao J (2008) Ten years of fluxes and stand growth in a young beech forest at Hesse, North-eastern France. Ann For Sci 65:704
Hackett C, Carolane J (1982) Introduction and crop profiles. In: Hackett C, Carolane J (eds) Edible horticultural crops: a compendium on information on fruit, vegetable, spice and nut species. Press, Sydney, Australia, Acad
Harley PC, Tenhunen JD (1991) Modeling the photosynthetic response of C3 leaves to environmental factors. In: Boote KJ, Loomis RS (eds) Modeling crop photosynthesis—from biochemistry to canopy. Anaheim, California, pp 17–39
Hussain MZ, Grünwald T, Tenhunen JD, Li YL, Mirzae H, Bernhofer C, Otieno D, Dinh NQ, Schmidt M, Wartinger M, Owen K (2011) Summer drought influence on CO2 and water fluxes of extensively managed grassland in Germany. Agric Ecosyst Environ 141:67–76
Hutchinson GK, Richards K, Risk WH (2000) Aspects of accumulated heat patterns (growing degree-days) and pasture growth in Southland. Proc NZ Grassl Assoc 62:81–85
Kang S, Post WM, Nichols JA, Wang D, West TO, Bandaru V, Izaurralde RC (2013) Marginal lands: concepts, assessment and management. J Agr Sci 5:129–139
Kiniry JR, Williams JR, Major DJ, Izaurralde RC, Gassman PW, Morrison M, Bergentine R, Zentner RP (1995) EPIC model parameters for cereal, oilseed, and forage crops in the northern Great Plains region. Can J Plant Sci 75:679–688
Levy PE, Grelle A, Lindroth A, Mölder M, Jarvis PG, Kruijt B, Moncrieff JB (1999) Regional-scale CO2 fluxes over central Sweden by a boundary layer budget method. Agricult Forest Meteorol 98–99:169–180
Luyssaert S, Inglima I, Jung M, Richardson AD, Reichstein M, Papale D, Piao SL, Schulze E, Wingate L, Matteucci G, Aragao L, Aubinet M, Beer C, Bernhofer C, Black KG, Bonal D, Bonnefond J, Chambers J, Ciais P, Cook B, Davis KJ, Dolman AJ, Gielen B, Goulden M, Grace J, Granier A, Grelle A, Griffis T, Grünwald T, Guidolotti G, Hanson PJ, Harding R, Hollinger DY, Hutyra LR, Kolari P, Kruijt B, Kutsch W, Lagergren F, Laurila T, Law BE, Lemaire G, Lindroth A, Loustau D, Malhi Y, Mateus J, Migliavacca M, Misson L, Montagnani L, Moncrieff J, Moors E, Munger JW, Nikinmaa E, Ollinger SV, Pita G, Rebmann C, Roupsard O, Saigusa N, Sanz MJ, Seufert G, Sierra C, Smith M, Tang J, Valentini R, Vesala T, Janssens IA (2007) CO2 balance of boreal, temperate, and tropical forests derived from a global database. Global Change Biol 13:2509–2537
Luyssaert S, Ciais P, Piao SL, Schulze E, Jung M, Zaehle S, Schelhaas MJ, Reichstein M, Churkina G, Papale D, Abril G, Beer C, Grace J, Loustau D, Matteucci G, Magnani F, Nabuurs GJ, Verbeeck H, Sulkava M, Van Der Werf GR, Janssens IA (2010) The European carbon balance. Part 3: forests. Global Change Biol 16:1429–1450
Mendham NJ, Shipway PA, Scott RK (1981) The effects of delayed sowing and weather on growth, development and yield of winter oil-seed rape (Brassica napus). J Agr Sci 96:389–416
Muraoka H, Saigusa N, Nasahara KN, Noda H, Yoshino J, Saitoh TM, Nagai S, Murayama S, Koizumi H (2010) Effects of seasonal and interannual variations in leaf photosynthesis and canopy leaf area index on gross primary production of a cool-temperate deciduous broadleaf forest in Takayama, Japan. J Plant Res 123:563–576
Neitsch SL, Arnold JG, Kiniry JR, Williams JR (2009) Soil and Water Assessment Tool. Theoretical Documentation Version 2009, Texas Water Resources Institute Technical Report No. 406. Texas, pp 1–618
Nguyen TT, Tenhunen JD (2013) Review of integrated ecological-economic analyses for bioenergy plants under climate change at local scale. Int J Clim Change Strateg Manag 5:324–343
Olesen JE, Bindi M (2002) Consequences of climate change for european agricultural productivity, land use and policy. Eur J Agron 16:239–262
Olesen JE, Trnka M, Kersebaum KC, Skjelvåg AO, Seguin B, Peltonen-Sainio P, Rossi F, Kozyra J, Micale F (2011) Impacts and adaptation of european crop production systems to climate change. Eur J Agron 34:96–112
Rebetez M, Mayer H, Dupont O, Schindler D, Gartner K, Kropp JP, Menzel A (2006) Heat and drought 2003 in Europe: a climate synthesis. Ann For Sci 63:569–577
Rötter R, van de Geijn SC (1999) Climate change effects on plant growth, crop yield and livestock. Clim Change 43:651–681
Rouault G, Candau JN, Lieutier F, Nageleisen LM, Martin JC, Warzée N (2006) Effects of drought and heat on forest insect populations in relation to the 2003 drought in Western Europe. Ann For Sci 63:613–624
Shurpali NJ, Hyvönen NP, Huttunen JT, Clement RJ, Reichstein M, Nykänen H, Biasi C, Martikainen PJ (2009) Cultivation of a perennial grass for bioenergy on a boreal organic soil—carbon sink or source? Global Change Biol Bioenergy 1:35–50
Solomon S, Qin D, Manning M, Marquis M, Averyt K, Tignor MMB, Le Roy Miller H Jr, Chen Z (2007) Climate Change 2007. The physical science basis. Contribution of working group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change (IPCC). Cambridge, Cambridge University Press
Tenhunen J (2013) Relating forest ecosystem flux studies to regional ecosystem services. In: Synthesis workshop on the carbon budget and forest ecosystem in the Asian Monitoring Network—the 20th Anniversary of the Takayama Site, Japan. Hida Earth Wisdom Center Takayama, pp 40–43
Tenhunen J, Geyer R, Adiku S, Reichstein M, Tappeiner U, Bahn M, Cernusca A, Dinh NQ, Kolcun O, Lohila A, Otieno D, Schmidt M, Schmitt M, Wang Q, Wartinger M, Wohlfahrt G (2009) Influences of changing land use and CO2 concentration on ecosystem and landscape level carbon and water balances in mountainous terrain of the Stubai Valley, Austria. Global Planet Change 67:29–43
Titei V, Teleuta A, Muntean A (2013) The perspective of cultivation and utilization of the species Silphium Perfoliatum L. Bull UASMV Ser Agric 70:160–166
Vadas PA, Barnett KH, Undersander DJ (2008) Economics and energy of ethanol production from alfalfa, corn, and switchgrass in the Upper Midwest, USA. Bioenerg Res 1:44–55
Zenone T, Chen J, Deal MW, Wilske B, Jasrotia P, Xu J, Bhardwaj AK, Hamilton SK, Philip Robertson G (2011) CO2 fluxes of transitional bioenergy crops: effect of land conversion during the first year of cultivation. Global Change Biol Bioenerg 3:401–412
Acknowledgments
This work was initiated as part of the project REGECON—Regional Economic Evaluation of Adaptation Measures in Agricultural, Forestry, and Bioenergy Production under the Influence of Climate Change funded by the Federal Ministry of Education and Research (BMBF) of Germany (Funding number: 01LA1101A). The authors also acknowledge the support of Mr. Asen of the Amt für Ernährung, Landwirtschaft und Forsten. Last but not least, we acknowledge those, who provided eddy-flux datasets to FLUXNET: Christian Bernhofer and Thomas Grünwald (TU Dresden, Germany), Andre Granier and Bernhard Longdoz (Institut Nationale de la Recherche Agronomique, Nancy, France), Marc Aubinet (University of Liege, Gembloux Agro-Bio Tech, Unit of Biosystems Physics, Belgium), Christine Moureaux, Tanguy Manise and Anne Ligne (University de Liège, Belgium).
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
About this article

Cite this article
Ruidisch, M., Nguyen, T.T., Li, Y.L. et al. Estimation of annual spatial variations in forest production and crop yields at landscape scale in temperate climate regions. Ecol Res 30, 279–292 (2015). https://doi.org/10.1007/s11284-014-1208-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11284-014-1208-4