
Abstract
Mid-exponential cultures of two traditional biotechnological yeast species, winery Saccharomyces cerevisiae and the less ethanol tolerant bottom-fermenting brewery Saccharomyces pastorianus, were exposed to different concentrations of added ethanol (3, 5 and 8%) The degree of ethanol-induced cell stress was assessed by measuring the cellular activity of superoxide dismutase (SOD), level of lipid peroxidation products, changes in cell lipid content and fatty acid profile. The resveratrol as an antioxidant was found to decrease the ethanol-induced rise of SOD activity and suppress the ethanol-induced decrease in cell lipids. A lower resveratrol concentration (0.5 mg/l) even reduced the extent of lipid peroxidation in cells. Resveratrol also alleviated ethanol-induced changes in cell lipid composition in both species by strongly enhancing the proportion of saturated fatty acids and contributing thereby to membrane stabilization. Lower resveratrol concentrations could thus diminish the negative effects of ethanol stress on yeast cells and improve their physiological state. These effects may be utilized to enhance yeast vitality in high-ethanol-producing fermentations or to increase the number of yeast generations in brewery.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The yeast Saccharomyces cerevisiae has been utilized by mankind for centuries for the production of bread and alcoholic beverages. Nowadays the application of yeast is widespread and plays an important role not only in food and chemical industry, but also in biomedical and environmental research. Nonetheless, ethanol remains the most valuable yeast-derived commodity (Stanley et al. 2010). The viability and fermentation performance of yeast depend on their ability to adapt to the ever-changing environment and maintain the equilibrium of intracellular physiological and chemical processes (Bauer and Pretorius 2000). This balance is threatened by multiple stressors affecting yeast cells during the fermentation process, including substrate and/or product inhibition, heat shock, oxidative stress, hyperosmolarity and exposure to toxic by-products (Zhao et al. 2014).
Among these factors, ethanol stress is a bottleneck for complete fermentation. S. cerevisiae is able to resist relatively high ethanol concentration (up to 20%) (Yamaoka et al. 2014). However, when the limit concentration is exceeded, ethanol negatively affects the growth rate of yeast, fermentation rate, and cell viability. In addition, high level of ethanol enhances other stresses, such as oxidative stress via increased production of ROS (reactive oxygen species) (Bleoanca et al. 2013; Ma and Liu 2010; Perez-Gallardo et al. 2013). ROS accumulation may lead to serious damage of basic cell biomolecules, such as nucleic acids, proteins (Cabiscol et al. 2000), and lipids (Bilinski et al. 1989).
Yeasts have developed a plethora of mechanisms in order to deal with the effects of high ethanol concentration. The negative effect of ethanol on cell membranes can be reduced by an adjustment of the membrane lipid composition. The ability of cells to alter the level of unsaturation of their membrane lipids is an important factor in cellular adaptation to environmental conditions. Reduction of membrane fluidity is associated with a higher content of saturated fatty acids, stronger hydrophobic interactions and increased content of phospholipids (Watson 1987). It has been reported by multiple authors that the increase in unsaturated/saturated fatty acids (UFAs/SFAs) ratio is instrumental for enhancing the ethanol tolerance in yeast (Ding et al. 2009; Henderson and Block 2014; You et al. 2003). However, according to Viegas et al. (2005), a high UFAs/SFAs ratio in cell lipids causes the loss of membrane integrity under stress conditions, as UFAs are susceptible to lipid peroxidation by ROS.
Oxidative damage to lipid molecules leads to lipid peroxidation and oxidation of polyunsaturated fatty acids to unstable lipid hydroperoxides, which cause oxidative damage to cell biomolecules (Halliwell and Gutteridge 2007). Even though S. cerevisiae are not capable of polyunsaturated fatty acid (PUFA) synthesis, if they are cultivated in a cultivation broth containing PUFAs, they can incorporate these molecules into their membrane structures, which increases the risk of lipid peroxidation (Bilinski et al. 1989; Farrugia and Balzan 2012). During fermentation, ethanol levels increase and cells are exposed to toxic levels of ethanol. Traditionally, the ethanol tolerance in the genus Saccharomyces is ordered from the most tolerant sake yeasts, trough wine and distillers’ yeasts to the least tolerant brewers’ yeasts (Casey and Ingledew 1986). The complex stresses which wine yeast face during the fermentation process have already been reviewed by Matallana and Aranda (2017). In the brewing process, normal fermentation conditions result in ethanol content of 3–6%, but in the case of high gravity fermentation, the ethanol level goes up to over 10%, similar to wine production (Briggs et al. 2004). The consequent ethanol stress may cause growth inhibition, reduced cell size (Canetta et al. 2006), reduced viability, reduced respiration and glucose uptake (Pascual et al. 1988), reduced fermentation (Fernandes et al. 1997), enzyme inactivation, lipid modification, and increased membrane permeability (Marza et al. 2002).
The aim of the present study was to investigate the effect of resveratrol on two yeast species, S. cerevisiae and Saccharomyces pastorianus, exposed to various concentrations of ethanol. Resveratrol is a phenolic compound naturally occurring in grapevines. It is known for its biological activities; particularly free-radical scavenging (Fremont 2000). Resveratrol has also been shown to chelate metal ions and keep them from reacting with ROS, as well as increase antioxidant enzyme activity, consequently preventing lipid peroxidation (Silva et al. 2015).
The effect of resveratrol on the modulation of ethanol stress was evaluated by measuring changes in fatty acid composition, the degree of oxidative damage (indicated by lipid peroxidation), and activity of superoxide dismutase, an important enzyme in antioxidant defense mechanism of cells.
Materials and methods
Yeast strains
Two yeast strains were used in this study: winery strain S. cerevisiae SIHA and bottom-fermenting brewery lager yeast strain S. pastorianus 95. Both strains were provided by Collection of Yeasts and Industrial Microorganisms (DBM), Department of Biochemistry and Microbiology, UCT Prague and were maintained in 50% glycerol solution at − 75 °C prior to use.
Cultivation conditions
The yeast inocula were cultivated in 100 ml of YPD medium (g/l): peptone, 20; yeast extract, 10; glucose, 20; initial pH 6.0. The cultivations were conducted in Erlenmeyer flasks on a rotary shaker at 150 rpm at 28 °C to the late exponential growth phase.
For determining the effect of resveratrol (Sigma-Aldrich), the medium was amended by resveratrol in the total concentration of 0.5 mg/l (R1) or 5 mg/l (R2) and ethanol concentration in the range of 0–8%. To prepare an inoculum the cells were harvested by centrifugation (9000g, 10 min, 15 °C), washed with phosphate buffer solution (67 mM; pH 7.0) and resuspended in the phosphate buffer.
For fatty acid analysis and biomass yield determination, superoxide dismutase and lipid peroxidation analysis, the cells were transferred into 500 ml Erlenmeyer flasks containing 200 ml YPD medium supplemented with ethanol ranging from 0 to 8% concentration or resveratrol and ethanol and incubated at temperature 28 °C and 100 rpm until they reached the late exponential growth phase.
Fatty acid composition analysis
The cells were harvested by centrifugation (9000g, 20 min, 25 °C), washed with phosphate buffer, frozen at − 75 °C and lyophilized.
The extraction and isolation of total lipids from yeast suspension was based on the method of Bligh and Dyer (1959). Briefly, the yeast suspension was ground with glass beads under liquid nitrogen and the crushed yeast was extracted according to Bligh and Dyer (1959).
Fatty acids methyl esters (FAMEs) were prepared after saponification and further esterification using BF3/MeOH. The products were dissolved in hexane and analyzed by gas chromatography–mass spectrometry (GC–MS). GC–MS of FAMEs was done on a Varian GC–MS system with the split/splitless injector (250 °C) and a SP™-2380 Capillary GC Column L × I.D. 60 m × 0.25 mm, df 0.20 µm capillary column. Helium was used as carrier gas at 1.0 ml min−1. The split/splitless injection port was maintained at 255 °C. The split ratio was 1:90, and the injection volume was 1 μl, the temperature program was as follows: 150 °C for 1 min, subsequently increasing at 20 °C min−1 to 180 °C and at 2 °C min−1 to 250 °C, which was maintained for 1 min. FAMEs were identified according to their mass spectra (Dembitsky et al. 1991; Vancura et al. 1987) and using a mixture of chemical standards obtained from Sigma-Aldrich. All experiments concerning the analysis of FAMEs and their derivatives were carried out by electron impact MS.
Superoxide dismutase (SOD) activity assay
The yeast cells were harvested by centrifugation (9000g, 10 min, 10 °C) and washed with phosphate buffer (67 mM; pH 7.0). Thereafter, the cells were ground with liquid nitrogen, resuspended in phosphate buffer and centrifuged. SOD activity was measured by inhibiting nitroblue tetrazolium reduction by O2 − generated by the xanthine/xanthine oxidase system using a commercial assay kit (Sigma-Aldrich).
Lipid peroxidation assay
Products of lipid peroxidation were determined according to Gajewska and Sklodowska (2007). One millilitre of disintegrated cell population was mixed with 1 ml of TBA (thiobarbituric acid) solution (29 mM TBA in 8.75 M acetic acid) and heated at 95 °C for 1 h. After cooling, 3.5 ml of n-butanol was added and the tubes were vigorously shaken. After centrifugation (9000g, 30 min, 15 °C) the fluorescence of organic layer was measured at 531 nm. The concentration of thiobarbituric acid reacting substances (TBARS) was estimated by referring to a standard 1,1,3,3-tetraethoxypropane and expressed as TBARS per milligram of protein.
Total soluble protein was determined by a modified method described by Bradford (1976). The disintegrated cell suspension was centrifuged (10,000g, 10 min, 15 °C), 10 μl of supernatant was added to 200 μl of Bradford’s reagent and after 5 min the absorbance was measured at 595 nm. Protein content was estimated by referring to a standard curve for bovine serum albumin. Standard deviation was calculated from five independent measurements.
Results
The effect of ethanol stress on the yeast cells S. cerevisiae and S. pastorianus and the ability of resveratrol to modulate this stress was evaluated by examining changes in fatty acid composition of the cells, activity of the antioxidative enzyme superoxide dismutase and the extent of peroxidation of lipids, which indicated the degree of oxidative damage.
Influence of growth phase
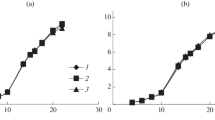
The added ethanol prolonged the lag and exponential phase (Fig. 1). Figure 2 illustrates the influence of growth phase on the degree of oxidative damage indicated by the concentration of TBARS. Biomass of the strain S. pastorianus 95 from the exponential, late exponential and stationary growth phase were collected and analyzed (7, 15 and 20 h for cells without added ethanol; 16, 24 and 33 h in case of yeast growing in 3% ethanol; and 24, 48 and 54 h in the case of cell growing in 5% ethanol). Both concentrations of ethanol made the cells more sensitive to oxidative damage. The level of TBARS in late exponential and stationary phase cells growing at 3% ethanol concentration was 135% lower and at 5% ethanol 318% lower than in exponential cells. Because of this, the protective effect of resveratrol was examined only in mid-exponential cells.

Growth characteristics of in S. cerevisiae SIHA (a, c, e) and S. pastorianus 95 (b, d, f) under ethanol stress (0, 3, 5, 8% ethanol) and addition of resveratrol (R1: 0.5 mg/l, R2: 5 mg/l)

Influence of growth phase on the degree of oxidative damage indicated by the concentration of TBARS in S. pastorianus 95 as affected by 0, 3 and 5% ethanol. Filled bar: mid-exponential phase; open bar: early stationary phase; striped bar: late stationary phase
Similar results were achieved in S. cerevisiae SIHA (data not shown). The two strains differed in their ability to tolerate ethanol since the winery strain S. cerevisiae SIHA grew also in the presence of 8% ethanol while S. pastorianus 95 failed to grow at this concentration. Therefore, the data for S. cerevisiae SIHA are given also for 8% ethanol concentration whereas the results for S. pastorianus 95 are shown only for 3 and 5% ethanol.
Effect of ethanol stress on superoxide dismutase activity
The effect of different ethanol concentrations and the presence of resveratrol on SOD activity was measured in mid-exponential cells of both strains. The strains differed in SOD activity since in the absence of ethanol and resveratrol the basic SOD activity of S. cerevisiae SIHA was about half that in S. pastorianus 95 (Fig. 3). In both strains the SOD activity in control cells in the absence of resveratrol increased with increasing ethanol concentration. In the presence of 0.5 mg/l resveratrol, the SOD activity of S. cerevisiae SIHA at any ethanol concentration was not as markedly affected as at the presence of 5 mg/l resveratrol, but both concentrations of resveratrol caused a decrease in SOD activity in comparison to cultivations without the presence of resveratrol. In S. pastorianus 95 the rise in SOD activity with increasing ethanol concentration was less pronounced. In general, both strains respond to ethanol by increasing their SOD activity and resveratrol decreases in both strains the enzyme activity.

Changes in superoxide dismutase activity in mid-exponential cells of S. cerevisiae SIHA and S. pastorianus 95 under ethanol stress (0, 3, 5, 8% ethanol) and addition of resveratrol (R1: 0.5 mg/l, R2: 5 mg/l)
Effect of ethanol and resveratrol on the extent of lipid peroxidation
With increasing ethanol concentration S. cerevisiae SIHA exhibited increasing lipid peroxidation that amounted to a fivefold increase in TBARS level at 8% ethanol (Fig. 4). Low concentrations of resveratrol (0.5 mg/l) significantly decreased the TBARS level, in the strain S. cerevisiae SIHA even in the absence of ethanol. At 8% EtOH, 0.5 mg/l resveratrol decreased lipid peroxidation by 25% and 5 mg/l resveratrol by 40%. S. pastorianus 95 showed higher lipid peroxidation levels at all ethanol concentrations (Fig. 4). Lower resveratrol concentrations slightly decreased the lipid peroxidation in this strain cultivated with 3% of ethanol.

Lipid peroxidation in mid-exponential cells of S. cerevisiae SIHA and S. pastorianus 95 under ethanol stress (0, 3, 5, 8% ethanol) and addition of resveratrol (R1: 0.5 mg/l, R2: 5 mg/l)
Effect of ethanol and resveratrol on lipid content and fatty acid profile
Cultivation of S. cerevisiae SIHA with ethanol led to a concentration-dependent drop of cell lipid content; with 8% ethanol this drop amounted to 20% (Table 1). In the presence of resveratrol, the drop was reduced to a maximum of 15% regardless of the resveratrol concentration used.
The slightly higher total lipid content of S. pastorianus 95 cells also decreased progressively with ethanol concentration (a 28% drop with 5% ethanol) (Table 2). This ethanol-induced drop in cell lipids was again reduced by resveratrol (10% drop with 5% ethanol plus 5 mg/l resveratrol).
Ethanol and resveratrol also caused changes in the fatty acid profile of both species. Under the effect of 8% ethanol the saturated fatty acids content of S. cerevisiae SIHA decreased (palmitic acid decreased by 52%, stearic acid by 63%) while the content of monounsaturated palmitoleic acid and oleic acid, which made up some 85% of total FA, increased (palmitoleic acid by 10%, oleic acid by 15%) (Table 1). In the presence of resveratrol, these changes in the fatty acid profile were reduced two- to five-fold. The content of the less abundant arachidic acid was affected neither by ethanol nor by resveratrol.
The brewery strain S. pastorianus 95 (Table 2) showed similar changes in fatty acid pattern. The most abundant fatty acids in S. pastorianus 95 were oleic acid, which made up around 46%, and palmitoleic acid around 38% of the total FA. In the presence of 5% ethanol the content of saturated FAs in the lipids again significantly decreased (33% drop in palmitic acid, 54% drop in stearic and 70% drop in arachidic acid content) while that of monounsaturated FAs increased (8% rise in palmitoleic and 14% rise in oleic acid). Resveratrol reduced both the drop in saturated FAs (10–14% drop) and the rise in monounsaturated ones (1–5% increase).
Resveratrol lowered the UFA/SFA ration in both strain when grown in the presence of ethanol (Fig. 5). Net resveratrol-induced changes in the total content of SFAs and UFAs at different ethanol concentrations relative to controls without resveratrol were evaluated in both species by summing up the values for SFAs and UFAs from Tables 1, 2 (see Fig. 6). In ethanol-free controls both concentrations of resveratrol slightly reduced the sum of SFAs and negligibly increased UFAs. However, in the presence of ethanol, resveratrol decreased the content of unsaturated FAs in both species by as much as 10% while it strongly (by as much as 80% in S. cerevisiae SIHA and by 40% in S. pastorianus 95) increased the content of saturated FAs. This increase in the level of saturated FAs caused by resveratrol in the presence of ethanol is likely to counteract the ROS-induced fluidization of cell membranes and cause thereby membrane stabilization.

UFA/SFA ratio in in S. cerevisiae SIHA (a) and S. pastorianus 95 (b) at 0, 3, 5, 8% ethanol related to control cultivations without the addition of resveratrol (R1: 0.5 mg/l, R2: 5 mg/l)

Net resveratrol-induced changes in the content of UFAs and SFAs in S. cerevisiae SIHA (a) and S. pastorianus 95 (b) at at 0, 3, 5, 8% ethanol related to control cultivations without the addition of resveratrol (R1: 0.5 mg/l, R2: 5 mg/l)
Discussion
The aim of this study was to assess the effect of the polyphenolic antioxidant resveratrol on the extent of the stress caused by increased ethanol concentration in two biotechnologically important yeast strains, the winery S. cerevisiae SIHA and the bottom-fermenting brewery S. pastorianus 95. In gauging the stress intensity and the protective effects of resveratrol against it, we focused on two features, changes in cell lipids and variations in antioxidative enzyme superoxide dismutase.
The cells of our two strains cultivated at 0, 3, 5 and 8% ethanol were exposed to two concentrations of resveratrol (0.5 and 5 mg/l) to assess its protective effect against the ethanol stress. Polyphenols such as resveratrol are preferentially incorporated into membrane lipid bilayers and act as hydrogen donors inhibiting the formation of lipid radicals (Dani et al. 2008).
In mammalian tissues such as rat liver or horse blood, resveratrol has been reported to reduce lipid peroxidation and diminish the effects of oxidative stress (Dani et al. 2008; Ememe et al. 2015; Kasdallah-Grissa et al. 2007; Silva et al. 2015). We found a similar lipid peroxidation reducing effect of resveratrol in yeast S. cerevisiae SIHA. In both yeast strains increasing ethanol concentration brought about a significantly increasing level of lipid peroxidation products, 8% ethanol increasing the TBARS level in mid-exponential cells of S. cerevisiae SIHA about fivefold while 5% ethanol caused the TBARS levels to double in both strains. In the presence of 0.5 mg/l of resveratrol, the lipid peroxidation levels in S. cerevisiae SIHA decreased and similar results were obtained for S. pastorianus 95 grown in 3% of ethanol. This concentration of resveratrol had no significant effect on S. pastorianus 95 grown in media without ethanol and with 5% of ethanol. This phenomenon can be explained, e.g., by the fact that the protective action of resveratrol in S. cerevisiae SIHA shows a hormetic dependence on ethanol concentration (Ahmad et al. 2003; Escote et al. 2012). Hormesis is a phenomena in which an organism responds biphasic (mostly as an “inverted U” curve) to a substance which is mostly toxic at higher doses (Chadwick and Maudsley 2010).
Higher levels of resveratrol (5 mg/l) seemed to have quite the opposite effect on lipid peroxidation (except for 8% of ethanol in S. cerevisiae SIHA). The antioxidant effects are in some cases dose dependent and thus might cause more oxidative damage to the cells at higher concentrations. Several studies (Cillard et al. 1980; Chen et al. 2014; Rietjens et al. 2002) already showed that the prooxidant activity of some antioxidants is dose dependent and might increase with higher concentrations.
Resveratrol exerts a protective effect against the ethanol-induced drop of total cell lipid content. With increasing ethanol concentration the percent fraction of lipids in cell dry weight in both yeast strains decreased by as much as 30% (Tables 1, 2) and even the lower resveratrol concentration used (0.5 mg/l) reduced this ethanol-induced drop to about half in S. cerevisiae SIHA and even more strongly in the more ethanol susceptible S. pastorianus 95.
As expected (Tables 1, 2), ethanol stress entailed a rise in the overall content of major UFAs in both yeast strains while the content of SFAs decreased. S. cerevisiae SIHA cultivated in the absence of ethanol had a UFA/SFA ratio on 4.1; this ratio rose to 9.8 in the presence of 8% ethanol and dropped back to 5.1 in the presence of resveratrol (5 mg/l) and 5.2 (0.5 mg/l) grown in 8% of ethanol. Similar data were obtained with S. pastorianus 95 (Fig. 5). The reason was that, in the presence of ethanol, resveratrol strongly (by as much as 80% in S. cerevisiae SIHA and by 40% in S. pastorianus 95) increased the content of SFAs relative to control samples without resveratrol, while hardly affecting the content of UFAs in both species (Fig. 6). This means that this increase in the level of SFAs caused by resveratrol in the presence of ethanol might counteract the ROS-induced fluidization of cell membranes caused by high UFAs/SFAs rations (Viegas et al. 2005) and aid in membrane stabilization. Furthermore, as seen in Tables 1, 2, in both species, resveratrol lowered the ethanol-induced rise in the content of oleic acid by more than 50%. According to You et al. (2003), oleic acid (C18:1) plays an important role in counteracting ethanol toxicity in S. cerevisiae. Palmitoleic and especially oleic acids prevent cell damage caused by increased membrane fluidity in yeast exposed to stressful levels of ethanol (Sajbidor et al. 1994; You et al. 2003). Apart from decreasing membrane fluidity by increasing the proportion of SFAs, resveratrol thus decreases the ethanol-enhanced fluidity of cell lipids by reducing the content of oleic acid.
The defences against oxidative stress include the action of antioxidant enzymes such as superoxide dismutase (Gille and Sigler 1995; Sigler et al. 1999). Loss of superoxide dismutase activity leads to a significant increase in the sensitivity of yeast cells to ethanol (Kitagaki et al. 2007). Exposure of S. cerevisiae to 7.5% ethanol has been found to result in a sixfold rise in SOD1 gene expression compared to the control (Bleoanca et al. 2013).
Consistent with these data, we found increased SOD activity in cells of both species exposed to increased ethanol concentrations (Fig. 3). Our data are at variance with these findings since S. pastorianus 95, a lager yeast strain that has a limited ability to withstand ethanol stress since lager beer contains less than 5% of ethanol, showed generally higher SOD activity than S. cerevisiae SIHA, a winery strain that is more resistant to ethanol.
Studies on different animal models showed that resveratrol increases the activity of antioxidant enzymes including SOD (Cenesiz et al. 2007; Dani et al. 2008; Kasdallah-Grissa et al. 2007; Rivera et al. 2009; Silva et al. 2015; Yang et al. 2005). In our study, both concentrations of resveratrol decreased the SOD activity of S. pastorianus 95 and S. cerevisiae SIHA cultivated in the presence of ethanol compared to cultivation without resveratrol. These findings could be explained by a possible indirect protective effect of resveratrol that could cause a lower induction of the enzyme in yeast cells. Similarly, Estruch et al. (2011) observed a decrease in SOD activity in healthy men after red wine intake, which shows that in order to save energy, the body reduces the enzyme activity when not necessary.
Resveratrol has a great potential as an antifungal and antimicrobial agent in biotechnologies. Previous studies showed that resveratrol inhibits the growth of wine spoilage yeasts, acetic acid bacteria (Pastorkova et al. 2013) and lactic acid bacteria (Garcia-Ruiz et al. 2011), which indicates that resveratrol could be a prospective agent to control the growth of wine spoilage microorganisms without the need for sulfites in winemaking. Our results suggest that low concentrations of resveratrol could be beneficial also in breweries. The protective action of resveratrol against ethanol stress improves the physiological state of yeast cells and resveratrol can thus be utilized to enhance yeast vitality in high-ethanol-producing fermentations or to increase the number of yeast generations in brewery.
References
Ahmad KA, Clement MV, Pervaiz S (2003) Pro-oxidant activity of low doses of resveratrol inhibits hydrogen peroxide-induced apoptosis. Ann N Y Acad Sci 1010:365–373. doi:10.1196/annals.1299.067
Bauer F, Pretorius IS (2000) Yeast stress response and fermentation efficiency: how to survive the making of wine-a review. S Afr J Enol Vitic 21:27–51
Bilinski T, Litwinska J, Blaszczynski M, Bajus A (1989) Superoxide dismutase deficiency and the toxicity of the products of autooxidation of polyunsaturated fatty acids in yeast. Biochim Biophys Acta 1001:102–106. doi:10.1016/0005-2760(89)90312-3
Bleoanca I, Silva AR, Pimentel C, Rodrigues-Pousada C, Menezes Rde A (2013) Relationship between ethanol and oxidative stress in laboratory and brewing yeast strains. J Biosci Bioeng 116:697–705. doi:10.1016/j.jbiosc.2013.05.037
Bligh EG, Dyer WJ (1959) A rapid method of total lipid extraction and purification. Can J Biochem Physiol 37:911–917. doi:10.1139/o59-099
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254. doi:10.1016/0003-2697(76)90527-3
Briggs DE, Brookes P, Stevens R, Boulton C (2004) Brewing: science and practice. Taylor & Francis, London
Cabiscol E, Piulats E, Echave P, Herrero E, Ros J (2000) Oxidative stress promotes specific protein damage in Saccharomyces cerevisiae. J Biol Chem 275:27393–27398. doi:10.1074/jbc.M003140200
Canetta E, Adya AK, Walker GM (2006) Atomic force microscopic study of the effects of ethanol on yeast cell surface morphology. FEMS Microbiol Lett 255:308–315. doi:10.1111/j.1574-6968.2005.00089.x
Casey GP, Ingledew WMM (1986) Ethanol tolerance in yeasts. Crit Rev Microbiol 13:219–280. doi:10.3109/10408418609108739
Cenesiz S, Yarim GF, Karabulut AB, Ara C (2007) Changing of antioxidant enzyme activity on the biliary obstructed rats treated with resveratrol. Dtsch Tierarztl Wochenschr 114:345–348. doi:10.2377/0341-6593-114-345
Chadwick W, Maudsley S (2010) Hormesis: a revolution in biology, toxicology and medicine. Springer, New York
Chen W, Lin YC, Ma XY, Jiang ZY, Lan SP (2014) High concentrations of genistein exhibit pro-oxidant effects in primary muscle cells through mechanisms involving 5-lipoxygenase-mediated production of reactive oxygen species. Food Chem Toxicol 67:72–79. doi:10.1016/j.fct.2014.02.004
Cillard J, Cillard P, Cormier M, Girre L (1980) α-Tocopherol prooxidant effect in aqueous media: increased autoxidation rate of linoleic acid. J Am Oil Chem Soc 57:252–255
Dani C, Pasquali MA, Oliveira MR et al (2008) Protective effects of purple grape juice on carbon tetrachloride-induced oxidative stress in brains of adult Wistar rats. J Med Food 11:55–61. doi:10.1089/jmf.2007.505
Dembitsky VM, Rezanka T, Bychek IA, Shustov MV (1991) Identification of fatty acids from Cladonia lichens. Phytochemistry 30:4015–4018. doi:10.1016/0031-9422(91)83455-T
Ding J, Huang X, Zhang L et al (2009) Tolerance and stress response to ethanol in the yeast Saccharomyces cerevisiae. Appl Microbiol Biotechnol 85:253–263. doi:10.1007/s00253-009-2223-1
Ememe MU, Mshelia WP, Ayo JO (2015) Ameliorative effects of resveratrol on oxidative stress biomarkers in horses. J Equine Vet Sci 35:518–523. doi:10.1016/j.jevs.2015.02.006
Escote X, Miranda M, Menoyo S et al (2012) Resveratrol induces antioxidant defence via transcription factor Yap1p. Yeast 29:251–263. doi:10.1002/yea.2903
Estruch R, Sacanella E, Mota F et al (2011) Moderate consumption of red wine, but not gin, decreases erythrocyte superoxide dismutase activity: a randomised cross-over trial. Nutr Metab Cardiovasc Dis 21:46–53. doi:10.1016/j.numecd.2009.07.006
Farrugia G, Balzan R (2012) Oxidative stress and programmed cell death in yeast. Front Oncol. doi:10.3389/fonc.2012.00064
Fernandes L, Corte-Real M, Loureiro V, Loureiro-Dias MC, Leao C (1997) Glucose respiration and fermentation in Zygosaccharomyces bailii and Saccharomyces cerevisiae express different sensitivity patterns to ethanol and acetic acid. Lett Appl Microbiol 25:249–253. doi:10.1046/j.1472-765X.1997.00214.x
Fremont L (2000) Biological effects of resveratrol. Life Sci 66:663–673. doi:10.1016/S0024-3205(99)00410-5
Gajewska E, Sklodowska M (2007) Effect of nickel on ROS content and antioxidative enzyme activities in wheat leaves. Biometals 20:27–36. doi:10.1007/s10534-006-9011-5
Garcia-Ruiz A, Moreno-Arribas MV, Martin-Alvarez PJ, Bartolome B (2011) Comparative study of the inhibitory effects of wine polyphenols on the growth of enological lactic acid bacteria. Int J Food Microbiol 145:426–431. doi:10.1016/j.ijfoodmicro.2011.01.016
Gille G, Sigler K (1995) Oxidative stress and living cells. Folia Microbiol 40:131–152. doi:10.1007/BF02815413
Halliwell B, Gutteridge J (2007) Free radicals in biology and medicine. Oxford University Press, Oxford
Henderson CM, Block DE (2014) Examining the role of membrane lipid composition in determining the ethanol tolerance of Saccharomyces cerevisiae. Appl Environ Microbiol 80:2966–2972. doi:10.1128/aem.04151-13
Kasdallah-Grissa A, Mornagui B, Aouani E et al (2007) Resveratrol, a red wine polyphenol, attenuates ethanol-induced oxidative stress in rat liver. Life Sci 80:1033–1039. doi:10.1016/j.lfs.2006.11.044
Kitagaki H, Araki Y, Funato K, Shimoi H (2007) Ethanol-induced death in yeast exhibits features of apoptosis mediated by mitochondrial fission pathway. FEBS Lett 581:2935–2942. doi:10.1016/j.febslet.2007.05.048
Ma M, Liu ZL (2010) Mechanisms of ethanol tolerance in Saccharomyces cerevisiae. Appl Microbiol Biotechnol 87:829–845. doi:10.1007/s00253-010-2594-3
Marza E, Camougrand N, Manon S (2002) Bax expression protects yeast plasma membrane against ethanol-induced permeabilization. FEBS Lett 521:47–52. doi:10.1016/S0014-5793(02)02819-3
Matallana E, Aranda A (2017) Biotechnological impact of stress response on wine yeast. Lett Appl Microbiol 64:103–110. doi:10.1111/lam.12677
Pascual C, Alonso A, Garcia I, Romay C, Kotyk A (1988) Effect of ethanol on glucose-transport, key glycolytic-enzymes, and proton extrusion in Saccharomyces cerevisiae. Biotechnol Bioeng 32:374–378. doi:10.1002/bit.260320317
Pastorkova E, Zakova T, Landa P et al (2013) Growth inhibitory effect of grape phenolics against wine spoilage yeasts and acetic acid bacteria. Int J Food Microbiol 161:209–213. doi:10.1016/j.ijfoodmicro.2012.12.018
Perez-Gallardo RV, Briones LS, Diaz-Perez AL et al (2013) Reactive oxygen species production induced by ethanol in Saccharomyces cerevisiae increases because of a dysfunctional mitochondrial iron-sulfur cluster assembly system. FEMS Yeast Res 13:804–819. doi:10.1111/1567-1364.12090
Rietjens IMCM, Boersma MG, de Haan L et al (2002) The pro-oxidant chemistry of the natural antioxidants vitamin C, vitamin E, carotenoids and flavonoids. Environ Toxicol Pharmacol 11:321–333. doi:10.1016/S1382-6689(02)00003-0
Rivera L, Morón R, Zarzuelo A, Galisteo M (2009) Long-term resveratrol administration reduces metabolic disturbances and lowers blood pressure in obese Zucker rats. Biochem Pharmacol 77:1053–1063. doi:10.1016/j.bcp.2008.11.027
Sajbidor J, Ciesarova Z, Smogrovicova D (1994) Influence of ethanol on the lipid content and fatty acid composition of Saccharomyces cerevisiae. Folia Microbiol 40:508–510. doi:10.1007/BF02814733
Sigler K, Chaloupka J, Brozmanova J, Stadler N, Hofer M (1999) Oxidative stress in microorganisms-I. Microbial vs. higher cells-damage and defenses in relation to cell aging and death. Folia Microbiol 44:587–624. doi: 10.1007/BF02825650
Silva P, Fernandes E, Carvalho F (2015) Dual effect of red wine on liver redox status: a concise and mechanistic review. Arch Toxicol 89:1681–1693. doi:10.1007/s00204-015-1538-1
Stanley D, Bandara A, Fraser S, Chambers PJ, Stanley GA (2010) The ethanol stress response and ethanol tolerance of Saccharomyces cerevisiae. J Appl Microbiol 109:13–24. doi:10.1111/j.1365-2672.2009.04657.x
Vancura A, Rezanka T, Marsalek J et al (1987) Effect of ammonium ions on the composition of fatty acids in Streptomyces fradiae, producer of tylosin. FEMS Microbiol Lett 48:357–360. doi:10.1111/j.1574-6968.1987.tb02624.x
Viegas CA, Cabral MG, Teixeira MC et al (2005) Yeast adaptation to 2,4-dichlorophenoxyacetic acid involves increased membrane fatty acid saturation degree and decreased OLE1 transcription. Biochem Biophys Res Commun 330:271–278. doi:10.1016/j.bbrc.2005.02.158
Watson K (1987) Temperature relations. In: Rose A, Harrison J (eds) The yeasts, vol 2. Academic Press, London, pp 41–72
Yamaoka C, Kurita O, Kubo T (2014) Improved ethanol tolerance of Saccharomyces cerevisiae in mixed cultures with Kluyveromyces lactis on high-sugar fermentation. Microbiol Res 169:907–914. doi:10.1016/j.micres.2014.04.007
Yang H, Lee MK, Kim YC (2005) Protective activities of stilbene glycosides from Acer mono leaves against H2O2-induced oxidative damage in primary cultured rat hepatocytes. J Agric Food Chem 53:4182–4186. doi:10.1021/jf050093+
You KM, Rosenfield CL, Knipple DC (2003) Ethanol tolerance in the yeast Saccharomyces cerevisiae is dependent on cellular oleic acid content. Appl Environ Microbiol 69:1499–1503. doi:10.1128/AEM.69.3.1499-1503.2003
Zhao H, Li J, Han B, Li X, Chen J (2014) Improvement of oxidative stress tolerance in Saccharomyces cerevisiae through global transcription machinery engineering. J Ind Microbiol Biotechnol 41:869–878. doi:10.1007/s10295-014-1421-8
Acknowledgements
The research was supported by Czech Science Foundation (GACR) project P503 17-00027S.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Gharwalova, L., Sigler, K., Dolezalova, J. et al. Resveratrol suppresses ethanol stress in winery and bottom brewery yeast by affecting superoxide dismutase, lipid peroxidation and fatty acid profile. World J Microbiol Biotechnol 33, 205 (2017). https://doi.org/10.1007/s11274-017-2371-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11274-017-2371-x