
Abstract
Knowledge of wetland vegetation spectral reflectance signatures can assist in spectral classification of remotely sensed images for monitoring of wetland hydroperiod. This study aimed at assessing the differences between wetland vegetation communities of varying species composition and density in terms of spectral reflectance. The investigation was carried out in floodplains at Nxaraga and Seronga in the Okavango Delta, Botswana. Spectral measurements were conducted during rising and receding flood stages. In each study area, 2 transects were located in homogeneous macrophyte stands, and in each transect 5–10 1 m2 quadrats were randomly set. In each quadrat, water depth and cover percentages (green leaved and senescent) of macrophyte vegetation were recorded, and a full reflectance spectrum between 325 and 1,075 nm wavelengths captured using a handheld spectroradiometer. Multiple regression analysis with stepwise selection of significant variables based on AIC was performed to determine the influence of various stand characteristics on spectral reflectance and vegetation indices (NDVI and EVI). The results showed that stand characteristics explained a large proportion of variance in spectral vegetation indices with r2 = 0.67 for NDVI and r2 = 0.75 for EVI. For NDVI, the significant explanatory variables were percentage green cover, senescent cover and water depth, while for EVI it was the total vegetation cover. Analysis of regression residuals for the various vegetation community classes showed significant differences between the classes in their NDVI, but no differences in EVI, with the exception of the Panicum repens class. Reflectance in visible wavelengths did not vary significantly between sites and seasons, but the NIR reflectance differed across sites and seasons. The results indicate that the simple vegetation indices in the Okavango Delta respond strongly to stand characteristics and thus can be used to derive these from remote sensing imagery. The scope for determination of vegetation communities from these indices is limited, however.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Aquatic productivity in wetlands leads to high production of biomass which serves as food for both aquatic and terrestrial animals, and forms the basis for rich and diverse ecosystems. Social and socio-economic values of wetlands are determined by provisioning of resources such as timber or thatching grass and fish. Wetlands are of aesthetic value and provide services such as flood protection and water purification. In the rural context, wetland vegetation is also often used as food or medicine. Knowing the values of wetland vegetation to the community means that they should be managed and monitored for any effects brought about as a result of wetland and dry land ecosystem interaction.
Managing and monitoring wetlands is difficult as some of the systems are large and inaccessible, therefore the use of satellite remote sensing may assist in understanding the system, since satellite imagery can cover a wide area at a time. Knowledge of wetland vegetation spectral reflectance signatures acquired at a close range or from ground based tools can assist in calibrating and creating spectral classification in satellite images for mapping of vegetation communities. Such mapping provides a means of monitoring wetland systems affected by stresses such as climate change, fire, wildlife and human impact, and is a necessary prerequisite to understanding their vegetation dynamics and to preserve their ecological and socio-economic functions.
Satellite imagery has been frequently used in monitoring and managing wetland ecosystems (e.g., Munyati 2000; Chopra et al. 2001). Satellite data provide a time series useful in analysis and monitoring of wetlands, especially where financial and logistical constraints make ground-based monitoring impossible. However, readily available satellite imagery is mostly of relatively coarse spatial resolution, making it difficult to identify or monitor small and narrow wetlands (Ozesmi and Bauer 2002; Adam et al. 2012). Satellites are also inadequate for mapping in detail ecological changes and maybe unsatisfactory in land cover classification due to limitations in spatial resolution. A factor further confounding the application of satellite remote sensing to monitoring the hydrology of flood pulsed wetlands is the effect of interaction between flooding and floodplain vegetation. These have been consistent problems in attempts to monitor flood extent in the Okavango Delta, a large, seasonally pulsed sub-tropical wetland in northern Botswana (Ringrose et al. 2003; McCarthy et al. 2005). In the Okavango, the flood pulse causes an annual expansion and contraction of flooded area, stimulating growth, development and decline of communities of floodplain macrophytes. The changing proportions of open water and leaf cover in the floodplains results in progressively modified spectral reflectance and vegetation indices.
Recent studies (Murray-Hudson et al. 2014a, b) in the Okavango Delta have used time series of satellite imagery to derive flood frequency and monthly flood duration, to improve the understanding of the hydrology–vegetation relationship. These studies were based on a single band (MODIS Band 1, 620–670 nm) threshold approach to classifying wet and dry areas within the wetland, but did not take into account the effects of the development of floodplain macrophytes over the course of the flood pulse cycle. Such effects may lead to the underestimation of flood extent as the growth of floodplain macrophytes progressively masks the signature of open water. More detailed knowledge of the spectral characteristics of wetland vegetation will clearly contribute to the ability to discriminate vegetated floodplains from adjacent dryland. Therefore this study aimed at assessing the differences between wetland vegetation communities of varying species composition and density in terms of spectral reflectance, by documenting the different spectral signatures and indices as the flood pulse progressed, to allow spatial and temporal comparisons to be made. This was approached by asking the following questions:
-
What observable differences are there in vegetation stand characteristics in terms of spectral reflectance?
-
What is the relationship between the stand characteristics and the spectral indices?
-
Which of the spectral indices are sensitive to species composition and density?
Materials and methods
Study site
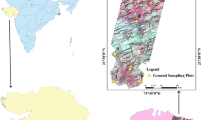
Spectral measurements were conducted at two sites: Seronga (Chaa Island) and Nxaraga Island in the Okavango Delta (Fig. 1). Seronga is situated near the apex of the Delta (the lower Panhandle region). The area has 50–70 m wide channels with water depths in the order of 2–5 m. Nxaraga is located in the distal part of the Delta. The channels are mostly narrow (5–20 m), with water depth in the order of 1.5–3 m. These two sites were selected for comparison as they occur in different positions along the hydroperiod gradient (Fig. 2). The Panhandle is a permanently, and Nxaraga is a seasonally inundated site. Both are inhabited by an assemblage of emergent grasses, sedges and floating-leaved herbaceous aquatic plants e.g. Nymphaea species.

Location map, the two study sites are shown in the lower Panhandle and middle of the Delta

Water level fluctuations in Seronga (1996) and Nxaraga (2010), rainfall and temperature in Maun (2010) (source http://www.okavangodata.ub.bw)
Data collection
Field measurements were taken in December 2012 (during receding flood conditions), March and May 2013 (during rising flood conditions; Fig. 2). An Analytical Spectral Device Field Spec® UV/VNIR Handheld Spectroradiometer, of a wavelength range from 325 to 1,075 nm with a 1.6 nm spectral sampling interval was used, covering the visible and infrared spectral wavelength ranges. Measurements were taken at knee height (40 cm) from the top of the stand, at 25° field of view. Data were collected between 10 am and 3 pm. The instrument was calibrated every 10 min in each plot with a white reference Spectralon® calibration panel, in accordance with the procedure recommended by the ASD manual (2002).
Spectral measurements were made on stands of relatively homogeneous composition and structure located along ~17–34 m long transects (Fig. 3). At each site, 2 such transects were set during rising flood and 1 transect during receding flood. Along each transect, 5–10 plots of 1 m2 were sampled, with the plots 3 m apart, making a total of 49 plots sampled; all the measured plots were set over inundated sites. Higher numbers of plots i.e. 5 or more were measured during rising flood, as the skies were mostly clear with no obscurance from cloud cover, giving us more time to measure up to 10 plots and a second transect in a day per season. A plot size of a 1 m2 was used in this study as it is an ideal plot area to be used for seasonally pulsed floodplains (Murray-Hudson et al. 2011). In each plot, 15–25 measurements were made of reflectance spectra, to match or exceed the 10–25 sets recommended by the ASD manual (2002). The following vegetation stand characteristics were recorded: estimated percentage of area covered by green leaved [green cover (%)] and senescent vegetation [senescent cover (%)], estimated percentage of area covered by water [water cover (%)] and vegetation species composition. The estimates were made from 100 % coverage of the plot and were a result of consensus from 3 to 4 investigators. Water depth was also measured using a 2.5 m dipstick.

Schematic diagram of the sampling plots. Five plots were sampled during receding flood and 9–10 plots during rising flood
Data processing
Mean spectra were then calculated from the 15–25 reflectance spectra collected per plot, using View Spec Pro Version 6.0 software. Reflectance values were taken as the ratio of target radiance to white reference radiance. Maximum reflectance values recorded in the general spectral regions—Blue (B—400–500 nm), Green (G—500–600 nm), Red (R—600–700 nm) and Near Infrared (NIR—700–1,075 nm) were used to calculate vegetation indices:
and
The latter equation was developed by Huete et al. (1997), where g = 2.5 is a gain factor, l = 1 is a canopy background adjustment factor, c1 = 6 and c2 = 7.5 are aerosol resistance coefficients, with values adopted from Gao et al. (2002).
Data analysis
As the first step of data analysis, full reflectance spectra were plotted for each site, and similarities or differences were described based on known spectral characteristics of green vegetation and water. Subsequently an analysis aimed at exploring and quantifying relationships between spectral reflectance indices derived from the full spectra and stand characteristics was carried out. Firstly, the differences of stand characteristics between seasons (rising and receding flood levels) and location were assessed using an F test (analysis of variance). Similar analyses were performed for mean reflectance in the R, G, B and NIR bands. Subsequently, reflectance and stand characteristics data were analyzed using the multiple linear regression (lm) procedure in the general statistics package of R version 2.15.0 (R Core Team 2013) under the following schematization of the sampling design:
-
Quantitative independent variables: the stand characteristics (cover values and water depth).
-
Qualitative nominal variables: season and site, considered as “treatments” or categorical variables in the multiple regressions.
-
Quantitative dependent variables: spectral reflectance indices (R, G, B, NIR, NDVI and EVI).
Under the above scheme, multiple regression analysis with stepwise selection of significant variables based on Akaike’s Information Criterion (AIC) was performed, which was aimed at finding which explanatory variables were the strongest determinants of spectral characteristics.
The measurements also included estimates of vegetation species composition (percent cover for each species) for each plot. Since the measured sites comprised a diverse set of plant species, in order to obtain an objective set of vegetation communities based purely on their species composition, hierarchical agglomerative cluster analysis was done using PC-ORD version 6.0 (McCune and Mefford 2011). Data were relativized with respect to the median to reduce chaining. Clustering was done with Euclidean distance and Ward’s method. The combination of Euclidean distance and Ward’s method improved the results as data can be manipulated, i.e. by relativization of the data as binary with respect to median. This will give the smallest value of percentage chaining. The resulting vegetation communities could, potentially, be used in the regression analysis as nominal variables, or “treatments” in addition to location and season. However, the total number of samples was not large enough to maintain statistically adequate numbers of samples for each combination of location, season and vegetation community. Thus, the regression analyses were carried out considering only two “treatments”: location and season. However, in order to assess the differences in spectral characteristics of the different vegetation community, analysis of regression residuals was performed for the various vegetation community classes.
Results
Classes of vegetation present at measurement sites
Twenty-three species were identified in both sites. Two species could not be easily identified or distinguished; they were classified as floating-leaved aquatics. The vegetation measured included emergent, floating and submerged vegetation. Four major vegetation classes were distinguished from cluster analysis (Fig. 4) and these were named after their dominant species: Cyperus pectinatus, Eleocharis dulcis, Leersia hexandra and Panicum repens.

The four major vegetation classes identified in Seronga and Nxaraga, Cyperus pectinatus, Eleocharis dulcis, Leersia hexandra and Panicum repens. The classes were named on the basis of the dominant species in the group
General characteristics of measured vegetation stands
Measured stands were characterized by vegetation cover in the ranges 0–100 %, with green vegetation in the range 3–100 %, senescent in the range 0–65 % and water depth in the range 8–149 cm. The majority of the plots presented a typical green vegetation reflectance curve (Fig. 5). The 4 exceptions to this were ambiguous, which were possibly affected by foreign materials in the instrument’s field of view such as algae, soil and sediments in water. Since the study aimed to understand and possibly quantitatively describe the relationship between aquatic vegetation and reflectance, it was decided to exclude them from the analyses as the non-vegetation factors were not our primary concern.

Mean reflectance spectra in Nxaraga and Seronga for both campaigns. Five quadrats were measured during receding flood and 9–10 quadrats were measured during rising flood. Two transects for Nxaraga are represented by Nxaraga (1, 2) and Seronga (1, 2)
In general, water depth was lower during the receding flood campaigns, and this was associated with lower proportions of water cover and higher senescent cover than during the rising flood campaigns (Fig. 6). Detailed statistical analysis confirmed these observations, and revealed that the following differences in cover characteristics and water depth between sites and seasons were statistically significant at p < 0.05 (Table 1):

Boxplots of vegetation stand characteristics (green and senescent cover, water cover and water depth) for sites and seasons. Boxes represent the upper and lower quartile, bold bars median, whiskers maximum or minimum value excluding outliers, open circle outliers
-
green vegetation cover was statistically different between sites, but not between seasons;
-
senescent vegetation cover was statistically different between seasons but not between sites;
-
water cover and water depth were statistically different between seasons and sites.
Spectral characteristics of measured sites: band reflectance and vegetation indices
For individual quadrats with typical vegetation reflectance curves, the main differences between sites in terms of full reflectance spectra were manifested in the depth of absorption troughs, the height of the green peak, and the level and slope of the NIR plateau (Fig. 5). The differences were quantitatively explored in the broad band reflectance wavelengths (R, G, B, and NIR) and vegetation indices (NDVI and EVI) (Fig. 7). The individual bands and vegetation indices showed some systematic differences between seasons and locations. They showed lower values when the flood level was rising than when it receded. During the rising flood, band reflectance and indices were generally similar for both study sites, but during receding flood, band reflectance and EVI were generally lower at Seronga than they were at Nxaraga.

Boxplots of measured broad band reflectance and derived vegetation indices for sites and seasons
Relationship between vegetation indices and stand characteristics
Results of the multiple regression analysis showed that the independent variables considered (water depth, water cover, green cover and senescent cover) explained a large proportion of the variance in the vegetation indices, and the NIR reflectance (66–75 % as indicated by r2 values ranging from 0.66 to 0.75) (Table 2). The significant variables selected by the AIC optimum model included green and senescent cover (and water depth for NDVI). The R, G and B ranges of the spectrum explained considerably less of the variance (r2 in the range of 0.29–0.5), with water cover and water depth being significant explanatory variables of reflectance in the G, the same plus senescent cover in B, and only vegetation cover in the R waveband.
Differences in spectral vegetation indices between vegetation communities
Analysis of regression residuals for the vegetation community classes was performed for the vegetation indices, NDVI and EVI (Fig. 8). The results showed that in general, vegetation classes were not different in EVI and NDVI, with possible exception of the Panicum repens class. The P. repens class had lower values of both NDVI and EVI than the other classes; however, this class was represented at only one site, thus it is possible that the result is accidental rather than systematic.

Boxplots of residuals from NDVI-stand characteristics and EVI-stand characteristics regressions, grouped by vegetation community
Discussion
The regression between vegetation stand characteristics and reflectance indices leaves 35–75 % of variance in the latter unexplained. There are a number of stand characteristics that we have not captured that may contribute to this, such as geometric form and canopy structure (Moraes et al. 2011). For example, the Seronga site was dominated by Eleocharis dulcis (both seasons) and floating-leaved aquatics (during rising flood) while Nxaraga was dominated by Leersia hexandra (both seasons) and Cyperus pectinatus (during rising flood). The canopy structure of the floating-leaved aquatic observed at Nxaraga during the rising flood (quadrat 4) exhibited a higher reflectance in the visible region (green band) and a relatively high and flat NIR plateau, hence behaving subtly differently from other quadrats. The vegetation cover was mostly green which resulted in a high reflectance with a well-defined peak observed in the green band and low reflectance in the blue and red (at 490 and 686 nm). Again, higher reflectance in the green band in Seronga site 1—Quadrat 3 was observed. Almost all the quadrats for Seronga site 2 were dominated by Eleocharis dulcis, reflected in the similar spectral response of these quadrats. In Nxaraga site 2, most quadrats were dominated by C. pectinatus while the anomalous quadrat was dominated by L. hexandra. The two species have different stem and leaf orientations which have an influence in the response pattern of the curve in the differing quadrat.
Spanglet et al. (1998) observed that a vertically structured canopy has a stronger effect on the spectral reflectance of vegetation than a spherical canopy. Vertical canopies are relatively open exposing water and possibly substrate in the background; these may affect reflectance as there will be combination of effects from shadows, soil absorption of light and leaf orientation generally contributing to a net decrease in reflectance. Water being exposed in the background may mix with plant signals resulting in a decrease in total reflectance radiation since it is known to be a good absorber mostly in the NIR wavebands (Silva et al. 2008). Spanglet et al. (1998) further argued that floating-leaved aquatics have thin leaves which causes little scattering in the NIR radiation back to the sensor, resulting in a higher reflectance in the NIR. Proctor and He (2013) who did an experimental study in Toronto found that floating macrophytes species tended to have the lowest reflectance in the visible and the green peak regions. Their results were similar to this study during rising flood levels, as a lower reflectance in the visible region was observed due to green cover (Table 3). The higher reflectance in the NIR observed in Seronga during rising flood is similar to the findings of a study in California (Searsville lake), in which the authors observed a high reflectance in NIR regions for emergent species with a dense canopy (Peñuelas et al. 1993). The difference is that in this study vegetation was sparse.
For all the dependent variables that were well explained by the stand characteristics (66–75 %), green cover and senescent cover influenced the reflectance behavior of the NIR waveband and EVI, with water depth being an additional variable with NDVI. Water cover was the only characteristic that was not significant in these variables (EVI, NDVI and NIR). This is due to the fact that water cover depicted spectral information that was different to green and senescent covers. Water cover was also equivalent to the sum of vegetation cover and therefore became insignificant in the regression.
Regression analyses have demonstrated that in the quadrat-to-quadrat reflectance, green cover and senescent cover were correlated with EVI, NIR and NDVI at p < 0.001. From the three dependent variables, NDVI was the only variable correlated with water depth (p < 0.001) in the AIC-selected model. In a study by Pettorelli et al. (2005), it was argued that water has a much lower NDVI value than do other surface features and that inundated areas can also be distinguished by changes in NDVI values before and after the flood, after eliminating the effects of other factors on NDVI. It has been observed that green vegetation strongly reflects NIR wavebands (Becker et al. 2005). It is the canopies of green foliage that have a multiple scattering effect on the NIR wavelengths, which gives optimum results as they enhance the leaf-level signal (Asner 1998). The EVI approach minimizes contamination problems such as canopy background and residual aerosol influences presented by NDVI, by providing complementary information about the spatial and temporal variations of vegetation (Pettorelli et al. 2005). High EVI values clearly indicate vegetation cover and low values of NDVI are good at distinguishing water depth, therefore using a combination of the two indices (EVI and NDVI) may help to improve flood extent mapping.
The vegetation indices showed high calculated values, ranging from −0.0086 to 0.62 for EVI, while for NDVI it ranged from −0.026 to 0.6414. Typical values of EVI and NDVI in inundated areas range from −1 to 1 with the negative values indicating the presence of open water, the positive green vegetation (Wang et al. 2003). The derived values were consistent as they showed the presence of vegetation and water. The negative values were nearly 0, implying that there was not a lot of water cover, while the positive values were close to 1 indicating abundant vegetation cover. A study by Pricope (2013) assumed that inundated vegetation can be differentiated from inundated areas if the EVI values are from 0.2 to 0.3 and that anything more than 3 should be classified as non-flooded. This assumption may be in error, since all the measured sites in this study were inundated, and values measured were high similar to those of the terrestrial reflectances.
When looking at season and vegetation cover, there are some systematic differences (Table 3). The statistical analysis showed that green vegetation cover was significantly different between sites and the senescent cover was significantly different across seasons. In general terms, during rising flood conditions (end of rainy season), when water depth in Seronga averaged 100.5 cm while that at Nxaraga averaged 75 cm, low biomass cover with most of vegetation being green was observed. During receding flood conditions (beginning of rainy season), when water depth averaged 30.5 cm in Seronga and 61.8 cm in Nxaraga, higher biomass was observed, although it was mostly senescent. In the climatic region of the Okavango Delta, where there is a very clear distinction between rainy season and dry season, highest green biomass occurs at the end of April (the end of the growing season as defined by radiation, temperature and local rainfall). This was not the case in this study, clearly suggesting that floodplain vegetation in the Okavango is strongly controlled by hydroperiod and less by local climate.
The analysis showed that the Panicum repens community was the only class differing from other communities in both vegetation indices (Fig. 8). Panicum repens occurs on sandy soils usually associated with water; in the lower or mid floodplain zones of the seasonal swamps (Ellery and Ellery 1997), which makes them facultative aquatics, while all the other vegetation types measured during our study were typical aquatic species. It is therefore possible that the Panicum class is indeed different in spectral reflectance to the other classes. However since this class was encountered at only one site, there is not enough evidence to confirm this assertion with confidence.
Conclusions
NDVI, EVI and NIR may be used to map emergent vegetation cover, this may be possible once we know from other sources what areas are inundated. The possibility for determination of vegetation community from the indices (EVI and NDVI) is limited as most of the vegetation exhibited similar spectral behavior. This may be improved by collecting more data at different phenological stages and measuring other factors such as soil and the internal structure of the leaves.
EVI responds to canopy type and structural variation and is NIR sensitive whereas NDVI is chlorophyll sensitive and responds to R variations (Pettorelli et al. 2005). This means that it will be difficult to use a single threshold value of NDVI or EVI to discriminate between inundated and non-inundated area. The results have shown that the inundated areas may have high values of NDVI and EVI, which are similar to those of dense terrestrial vegetation cover. Therefore inundation classification based on thresholds of NDVI, EVI or NIR will be uncertain.
References
Adam EM, Mutanga O, Rugege D, Ismail R (2012) Discriminating the papyrus vegetation (Cyperus papyrus L.) and its co-existent species using random forest and hyperspectral data resampled to HYMAP. Int J Remote Sens 33(2):552–569
Analytical Spectral Devices, Inc. (2002) HandHeld Spectroradiometer; User’s guide. Analytical Spectral Devices, Inc., Boulder
Asner GP (1998) Biophysical and biochemical sources of variability in canopy reflectance. Remote Sens Environ 64(3):234–253
Becker BL, Lusch DP, Qi J (2005) Identifying optimal spectral bands from in situ measurements of Great Lakes coastal wetlands using second-derivative analysis. Remote Sens Environ 97(2):238–248
Chopra R, Verma VK, Sharma PK (2001) Mapping, monitoring and conservation of Harike wetland ecosystem, Punjab, India, through remote sensing. Int J Remote Sens 22(1):89–98
Ellery K, Ellery W (1997) Plants of the Okavango Delta: a field guide. Tsaro Publishers, Durban
Gao X, Huete AR, Ni W, Miura T (2002) Optical-biophysical relationships of vegetation spectra without background contamination. Remote Sens Environ 74(3):609–620
Huete AR, Liu HQ, Batchily K, Van Leeuwen WJDA (1997) A comparison of vegetation indices over a global set of TM images for EOS-MODIS. Remote Sens Environ 59(3):440–451
McCarthy J, Gumbricht T, McCarthy TS (2005) Ecoregion classification in the Okavango Delta, Botswana from multitemporal remote sensing. Int J Remote Sens 26(19):4339–4357
McCune B, Mefford MJ (2011) PC-ORD. Multivariate analysis of ecological data. Version 6. MjM Software, Gleneden Beach
Moraes CE, Pereira G, Cardozo F, de Oliveira G, Ferreira MP (2011) Spectral response of vegetation covered surface subject to flooding due to viewing geometry. Geografia 36:188–199
Munyati C (2000) Wetland change detection on the Kafue Flats, Zambia by classification of multitemporal remote sensing image dataset. Int J Remote Sens 21(9):1787–1806
Murray-Hudson M, Combs F, Wolski P, Brown MT (2011) A vegetation-based hierarchical classification for seasonally pulsed floodplains in the Okavango Delta, Botswana. Afr J Aquat Sci 36(3):223–234
Murray-Hudson M, Wolski P, Cassidy L, Brown MT, Thito K, Kashe K, Mosimanyana E (2014a) Remote Sensing-derived hydroperiod as a predictor of floodplain vegetation composition. Wetlands Ecol Manage. doi:10.1007/s11273-014-9340-z
Murray-Hudson M, Wolski P, Murray-Hudson F, Brown MT, Kashe K (2014b) Disaggregating hydroperiod: components of the seasonal flood pulse as drivers of plant species distribution in floodplains of a tropical wetland. Wetlands. doi:10.1007/s13157-014-0554-x
Ozesmi SL, Bauer ME (2002) Satellite remote sensing of wetlands. Wetlands Ecol Manage 10(5):381–402
Peñuelas J, Gamon JA, Griffin KL, Field CB (1993) Assessing community type, plant biomass, pigment composition, and photosynthetic efficiency of aquatic vegetation from spectral reflectance. Remote Sens Environ 46(2):110–118
Pettorelli N, Vik JO, Mysterud A, Gaillard JM, Tucker CJ, Stenseth NC (2005) Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol Evol 20(9):503–510
Pricope NG (2013) Variable-source flood pulsing in a semi-arid transboundary watershed: the Chobe River, Botswana and Namibia. Environ Monit Assess 185(2):1883–1906
Proctor C, He Y (2013) Estimation of foliar pigment concentration in floating macrophytes using hyperspectral vegetation indices. Int J Remote Sens 34(22):8011–8027
R Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. ISBN 3-900051-07-0. http://www.R-project.org/
Ringrose S, Vanderpost C, Matheson W (2003) Mapping ecological conditions in the Okavango delta, Botswana using fine and coarse resolution systems including simulated SPOT vegetation imagery. Int J Remote Sens 24(5):1029–1052
Silva TS, Costa MP, Melack JM, Novo EM (2008) Remote sensing of aquatic vegetation: theory and applications. Environ Monit Assess 140(1–3):131–145
Spanglet HJ, Ustin SL, Rejmankova E (1998) Spectral reflectance characteristics of California subalpine marsh plant communities. Wetlands 18(3):307–319
Wang J, Rich PM, Price KP (2003) Temporal responses of NDVI to precipitation and temperature in the central Great Plains, USA. Int J Remote Sens 24(11):2345–2364
Acknowledgments
The authors thank the German Ministry for Education and Research (BMBF) for sponsoring this project (TFO). We also thank the field technician for their assistance in field work and herbarium unit (ORI) with their assistant in sample identification. Appreciations to Mr Tsheboeng who aided in cluster analysis. Thanks are also due to two anonymous reviewers whose comments and suggestions on an earlier version helped greatly to improve this manuscript. The work was carried out under Botswana Government Research Permit reference number: EWT 8/36/4 XVI (47).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Thito, K., Wolski, P. & Murray-Hudson, M. Spectral reflectance of floodplain vegetation communities of the Okavango Delta. Wetlands Ecol Manage 23, 637–648 (2015). https://doi.org/10.1007/s11273-014-9403-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11273-014-9403-1