
Abstract
The objective of this study was to analyze tree response to environmental pollution using basal area increment (BAI) tree ring stable isotopes and radiocarbon. Scots pine (Pinus sylvestris L.) was assessed from three fresh mixed coniferous forest habitat sites within highly urbanized and populated areas of Silesia subject to high levels of atmospheric pollution and were compared with trees from a site in Silesia where atmospheric emissions were comparatively low. The combined analysis of tree ring width and isotopic data allowed the identification of tree adaptation to environmental pollutants. Changes in BAI revealed a clear long-term decrease in wood increment from 1960 to 1980. We also observed depletion rates of carbon isotopes (14C and δ13C) and increased water use efficiency related to atmospheric CO2 emissions from fossil fuel combustion.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Emissions from human activities connected with home heating, motor vehicles, and industrial activities, including low and high stack emissions, impact the environment. In Poland, most electricity is generated by burning coal. The extensive use of coal as a primary energy source produces significant emissions of pollutants such as sulfur oxides, carbon oxides, particulate matter, and B(a)P (Kaleta, 2014). Between 1960 and 1980, large increases in the emission of industrial pollutants occurred in Silesia, which resulted in the introduction of industrial pollution abatement measures. However, despite actions to reduce pollution, according to the European Environment Agency (EEA, 2021), in the past decade, some cities in Silesia were among the most polluted in the EU. Atmospheric pollution not only can affect tree vigor, but its effects also can form a permanent record captured in the carbon isotope composition of tree rings. Variations in atmospheric carbon isotopic composition can be due to many factors, for example, from an increase in anthropogenic CO2 emissions, biomass burning, releases from nuclear reactors, and the exchange of carbon within and between different natural carbon reservoirs. Fossil fuel combustion affects atmospheric carbon isotope levels through the Suess effect, in which the admixture of carbon from fossil fuel combustion dilutes the proportion of 14C and 13C in the atmosphere (Keeling, 1973; Rakowski, 2011; Suess, 1955).
Trees can be used as bioindicators of ecosystem function. For example, α-cellulose has been considered a reliable component of the annual tree ring for examining carbon isotopes because it is directly related to the formation of glucose and cellulose by the plant during photosynthesis (Park & Epstein, 1961; Rakowski, 2011; Saurer & Siegwolf, 2007; Sensuła & Pazdur, 2013; Sensuła & Wilczyński, 2018; Sensuła et al., 2011). In addition, when cellulose forms, further carbon exchange between saccharide molecules and the atmosphere is blocked (Białobok et al., 1993). Pine tree rings have been used to evaluate responses to anthropogenic and natural climatic factors and have been used to document changes in tree growth and carbon isotopic composition of Scots pine (P.sylvestris L.) (for example, Nöjd & Reams, 1996; Krąpiec & Szychowska-Krąpiec, 2001; Wilczyński, 2006; Stravinskiene et al., 2013; Sensuła et al., 2011; Sensuła, Opała, et al., 2015; 2018; Sensuła and Pazdur, 2013).
Elevated CO2 induces changes in stomatal conductivity (gs) and photosynthetic rates (Amax), which can increase plant intrinsic water use efficiency (iWUE = Amax/gs). Furthermore, changes in WUE are associated with quantitative changes in isotopic carbon and oxygen ratios in plants, as described by Scheidegger et al. (2000) and Saurer et al. (2004). Emissions of other pollutants may also influence tree growth and carbon isotope composition of tree rings; however, information on such emissions is unavailable due to a lack of reporting by government and industry.
The influence of climate on the isotopic composition of wood has been the subject of previous research and is beyond the scope of this study. Industrial pollution reduces tree vigor and growth and increases sensitivity to meteorological factors, increasing the heterogeneity of their annual incremental responses and weakening the strength of the climatic signal contained in tree rings and in tree basal area increment (BAI). Previous dendrochronological studies indicate that Scots pine is very sensitive to industrial pollution (Nöjd & Reams, 1996; Krąpiec & Szychowska-Krąpiec, 2001; Juknys et al., 2002; Wilczyński, 2006; Malik et al., 2012; Stravinskiene et al., 2013; Sensuła, Wilczynski, et al., 2015; Sensuła et al., 2017;, 2018; Sensuła & Wilczyński, 2018). However, most studies of tree rings evaluated changes in radial growth, which in Scots pine is characterized by a clear decrease with age (the so-called age trend), which can potentially mask changes in tree condition. In contrast to radial growth, basal area increment (BAI) is characterized by a long-term upward trend with age in healthy trees, peaking at an age of several dozen years (Erteld & Hengst, 1966; Wilczyński, 2020).
This study examines the responses of Scots pine to environmental change in four forest areas in Silesia (south part of Poland). Specifically, we assess whether BAI in younger trees is a sensitive indicator of tree vigor, and we examine the use of carbon isotopes in tree rings as a long-term record of historical changes in environment and tree condition.
2 Material and Methods
The research was carried out in four forest areas, three of which have high levels of emissions of airborne pollutants (near the towns of Łaziska (lab code: LAP), Kędzierzyn-Koźle (lab code: ZKC), and Dąbrowa Górnicza (lab code: HKF2)). The fourth forest site was in Olesno (lab code: OLE) and served as a non-polluted control, located about 100 km from large industrial areas (Fig. 1).

Sampling sites localized nearby Łaziska, Kedzierzyn-Koźle, Dąbrowa Górnicza, and Olesno
2.1 BAI
Fifteen 100-year-old Scots pine trees were selected for study in each of the four stands (Olesno, OLE– the control site; Łaziska, Kędzierzyn-Koźle, and Dąbrowa Górnicza– the sites affected by industrial emissions). Two increment cores were taken 1.3 m above ground level from each sample tree. Tree ring widths on increment cores were measured to the nearest 0.01 mm using the CoRecorder and CDendro programs (www.cybis.se) and were dated and rechecked using the Cofecha computer program (Holmes, 1983). The average radial increment in each year (r) was calculated using the two cores from each tree (r1 and r2), where r = (r1 + r2)/2. Annual basal area increment (BAI) was calculated from the average radius values as follows:
where BAIi is basal area increment in year i, ri is radial increment in year i, and ri−1 is radial increment in year i − 1.
At each site, the annual BAI of the 15 sampled trees was averaged for each year. In this way, site BAI chronologies were constructed. In addition, the BAI series of each tree was subjected to indexing and autoregressive modeling using the ARSTAN program (Cook & Holmes, 1986). The program fits twice a negative exponential curve or trend line to each BAI series. The BAI indices (BAII) for each year are calculated using the formula:
BAIIi = Ri/Yi, where Ri is the BAI in year i and Yi is the value of a fit curve in year i.
Then each indexed series was subjected to autoregressive modeling in order to eliminate autocorrelation. To calculate rbar, the residual series created by the ARSTAN program were used. This removed trends, long-term fluctuations, and autocorrelation in the BAI series. For each site, the mean inter-series correlation coefficient (rbar) was estimated to quantify the confidence interval of the indexed BAI chronology for each site and to measure the common variance between individual series (Briffa & Jones, 1990).
The running rbar indices were calculated and plotted for 15-year periods (Fig. 2). The average correlation coefficient of the indexed series determines the degree of homogeneity of the annual growth responses of trees from year to year. It, therefore, gives an insight into the characteristics of inter-tree variability within a stand and the occurrence of environmental disturbances (Briffa et al., 1987).

The BAI chronologies (thin line) and 15-year running values of rbar (thick line). Consecutive values of rbar are plotted at the end of each 15-year period – the first period ranged 1941–1955 and the last period 1998–2012. Łaziska, Kędzierzyn-Koźle, Dąbrowa Górnicza, and Olesno
2.2 Stable Isotopes and Radiocarbon
The standard protocol described by Pazdur et al. (2013) was used for α-cellulose extraction from annual tree rings. The isotopic composition of tree rings is described by δ (in permilles), where δ = (Rsample/Rstandard − 1)*1000, and R is the ratio of the heavier to lighter isotope in the sample and in the standard.
Intrinsic WUE, directly linked to the ratio of intercellular to atmospheric CO2, was calculated using carbon isotopes in tree rings according to Saurer et al. (2004):
where δ13Ccel and δ13Cair are respectively carbon isotope composition of α-cellulose and the air, ca is atmospheric CO2 concentration, a ~ 4.4‰ is isotope fractionation during CO2 diffusion through stomata, b ~ 27‰ is isotope fractionation during fixation by Rubisco, Amax is photosynthesis rate, and gs is stomata conductance, respectively.
Determination of carbon and oxygen isotopes in α-cellulose was carried out to assess differences in photosynthetic efficiency and stomatal conductivity from 1975 to 2012. Carbon and oxygen stable isotope compositions were measured at the Mass Spectrometry Laboratory of the Institute of Physics at the Silesian University of Technology, Poland, using continuous-flow isotope ratio mass spectrometry (ISOPRIME, GV Instruments, Manchester, UK). Samples consisted of 60 µg of α-cellulose for δ13C measurements and 90 µg of α-cellulose for δ18O determinations (Sensuła et al., 2017, 2018). The detailed analysis of the oxygen isotopes fractionation has already been discussed (Sensuła & Wilczyński, 2018) and is out of the scope of current research. In the current investigation, δ18O data have been used to analyze the changes in WUE, as described by Scheidegger et al. (2000) and (Saurer and Siegwolf, 2007).
The α-cellulose samples were converted to graphite according to procedures described by Piotrowska (2013). The Δ14C in graphite was determined by the Rafter Radiocarbon Laboratory (Lower Hutt, New Zealand) or the DirectAMS Laboratory (Bothell, WA, USA). The NIST Oxalic Acid II standard was used for normalization, and black coal was used as a blank material. Values of Δ14C (‰) were calculated according to van der Plicht and Hogg (2006), in which.
where F14C is the normalized radiocarbon concentration, λ is the decay constant for radiocarbon isotopes (equal to 8267 yr−1), and Ti is calendar year.
The analysis of isotopes composition of tree rings was conducted with annual resolution over the years 1975–2012 (stable isotopes), with 5-year resolution between 1975 and 2000, and with annual resolution from 2000 to 2012 (radiocarbon). A detailed description of those results was presented by Sensuła et al., (2018, 2020).
3 Results and Discussion
Trees can be biomonitors of the environment. In this regard, annual tree ring properties can be important in the analysis of local and regional changes in the environment, including the effects of industrial pollution.
3.1 Dendrochronology
Tree rings from the four research plots showed dynamic changes in basal area increment and rbar values over time (Fig. 2). In the years 1935–1955 at Kędzierzyn-Koźle, Dąbrowa Górnicza, and Łaziska, there was a significant increase in BAI. In this period, rbar values remained high. However, in Olesno (control) forest area, high BAI growth and rbar values were maintained only up to the mid-1970s (Fig. 2). The strong decrease of rbar values at Kędzierzyn-Koźle, Dąbrowa Górnicza, and Łaziska occurred in the years 1960–1990, i.e., when emissions of industrial pollutants peaked. At Olesno (OLE), decreasing BAI and much weaker rbar occurred primarily in the 1970s and 1980s (Fig. 2). At the beginning of the 1990s, BAI and rbar increased in all forest areas in the study, with the largest increase in both indicators at Kędzierzyn-Koźle, Dąbrowa Górnicza, and Łaziska (Fig. 2).
Industrial pollution weakens the physiological activity of trees (Emberson, 2003; L’Hirondelle & Addison, 1985; McLaughlin et al., 2002; Percy & Ferretti, 2004), reducing their vitality and growth (Juknys et al., 2002; Stravinskiene et al., 2013; Wilczyński, 2006). Pollution also decreases sensitivity to meteorological factors, increases the heterogeneity of annual growth, and weakens the climatic signal in tree ring chronologies (Wilczyński, 2006, 2020). Changes in BAI revealed a clear long-term decrease in wood increment in study trees. This period marked a peak in emissions of industrial pollutants in Silesia. In those years, the annual variability of BAI and the tree-to-tree uniformity of BAI responses to other factors decreased (Fig. 2), which may be a result of pollutants. Early in the 1990s, following a reduction in industrial pollutant emissions, Scots pine resumed increased wood growth rates. In this period also, the annual variability of BAI and the homogeneity incremental reactions of trees increased (rbar). These results show that Scots pine can recover from damage caused by heavy industrial pollution.
These results confirm the usefulness of BAI and the degree of homogeneity of growth responses (rbar) for evaluating the effects of industrial pollution on trees. Scots pine growing in areas of heavy pollution had, in the period of greatest emissions, clearly reduced BAI and increased heterogeneity in annual growth. Following the reduction of pollutant emissions, pine trees quickly increased both wood growth and the homogeneity of annual growth. In the early 2000s, trees in this study reached their natural maximum BAI, after which the homogeneity of annual incremental responses returned to values seen before the prior century’s period of increased pollution.
3.2 Carbon Isotopes
Variations in carbon isotope concentrations in tree rings (Figs. 3 and 4) can be due to the admixture of atmospheric CO2 with CO2 from the burning of fossil fuels, as well as the exchange of CO2 between natural carbon reservoirs. From 1975 to 2012, Δ14C in tree rings decreased exponentially from year to year, from 409 to 32‰ in Scots pine at Kędzierzyn-Koźle, from 399 to33‰ at Dąbrowa Górnicza, from 375 to 34‰ at Łaziska, and from 380 to 34‰ at Olesno. The admixture of atmospheric carbon with carbon from biomass burning or with carbon from other emission sources enriched in 14C in this region can be significant. In this regard, the most significant local Suess effect was observed in Łaziska.

The comparison of radiocarbon concentrations in Scots pine tree rings (1975–2012) growing in four forests in Silesia and atmospheric radiocarbon concentration measured at the Alpine research station in Jungfraujoch (JFJ) (Hammer & Levin, 2017), an area considered unaffected by local anthropogenic effects

a Variation in iWUE in α-cellulose extracted from annual pine tree rings and increases atmospheric carbon concentration. b Theoretical, quantitative link between isotopic carbon and oxygen in plants due to changes in stomatal conductivity (gs) and rates of CO2 assimilation (Amax), as described by (Saurer and Siegwolf, 2007) and Scheidegger et al. (2000); direction of the arrow presents a direction of the changes; “*” means no changes. c Variation in oxygen and carbon isotopes
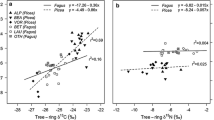
Between 1975 and 2012, the concentration of atmospheric CO2 increased from 330 to 394 ppm (Boden et al., 2016), while over the same period, iWUE varied between ca. 90 µmol/mol and 128 µmol/mol, a significant increase in iWUE of about 42%. According to the quantitative model described by Suarer and Scheidegger (Fig. 4b and c), pines react to environmental stress mostly by reducing stomatal conductivity. With climate change, warmer temperatures, especially in concert with reduced rainfall, could produce water stress resulting in reduced stomatal conductivity. In locations where there are large emissions of CO2 from fossil fuel combustion, local depletion of δ13C may be observed due to admixture of CO2 from different sources (Ferrio et al., 2003). In an area where there is not a strong local Suess effect, δ13C in atmospheric CO2 is equal to ~ − 8.5‰ (White et al., 2015). We observed that δ13C in trees ranged from − 24.4 to − 23.1‰ in Łaziska, from − 25.3 to − 22.9‰ in Dąbrowa Górnicza, from − 24.6 to − 23.2‰ in Kędzierzyn-Koźle, and from − 25.1 to − 23.5‰ in Olesno. Mean δ13C across all sites was − 23.9‰. According to Zimnoch et al. (2012), in southern Poland, the δ13C in coal is ~ − 24‰ and in gasoline is ~ − 31‰. When these sources mix with CO2 in the atmosphere, the result is depletion of δ13C in atmospheric CO2 and in the biosphere.
4 Conclusions
In conclusion, our study indicates that Scots pine can be useful for biomonitoring environmental conditions. In particular, tree conditions can be important in the analysis of local and regional changes in the environment affected by pollution. The combined analysis of tree ring width and isotopic data allowed the identification of tree adaptation to environmental pollutants. Changes in BAI revealed a clear long-term decrease in wood increment in the years of the culmination of industrial pollution (1960–1990) and an increase in the following years. Based on these results, we believe that Scots pine is able to regenerate rapidly after reducing harmful emissions. We also observed depletion rates of carbon isotopes (14C and δ13C) and an increase in the water use efficiency related to atmospheric CO2 emissions from fossil fuel combustion. In the region where most energy has been produced by burning coal, the result has been that during photosynthesis, CO2 fixed by trees will be influenced by depletion of δ13C caused by the Suess effect.
Data Availability
The datasets (BAI and stable isotopes composition of pine tree rings) analyzed during this study are included in this published article and its supplementary information files. Radiocarbon datasets are available from the authors upon reasonable request.
References
Boden, T.A., Marland, G. & Andres, R.J. (2016). Global, regional and national fossil-fuel CO2 emissions. Carbon Dioxide Information Analysis Center, Oak Ridge National Laboratory, U.S. Department of Energy, Oak Ridge, Tenn., U.S.A. https://doi.org/10.3334/CDIAC/00001_V2010
Hammer, S. & Levin, I. (2017). Monthly mean atmospheric 14CO2 at JungfraujochandSchauinsland from 1986 to 2016 [data set, www document]. University Library Heidelberg https://doi.org/10.11588/data/10100
Białobok, S., Boratynski, W., & Bugała, W. (1993). Biologia sosny zwyczajnej. Polish Academy of Sciences publications.
Briffa, K.R.& Jones, P.D. (1990). Basic chronology statistics and assessment. In: Cook ER, Kairiukstis LA, editors. Methods of Dendrochronology. Applications in the Environmental Sciences. Kluwer Academic Publishers, Dordrecht, 137–152.Cook, E.R., & Holmes, R.L. (1986). Users manual for computer program ARSTAN: Tree-ring chronologies of western North America: California, eastern Oregon and northern great basin. Chronology Series 6. Tucson, University of Arizona, 50–65.
Briffa K.R., Wigley T.M.L & Jones P.D. (1987). Standardization and the preparation of chronologies some contrasting approaches. Towards an objective approach to standardization. In: Kairiukstis L., Bednarz Z., Feliksik E. Methods of dendrochronology. Proceedings of the Task Force Meeting on Methodology of Dendrochronology: East/West Approaches, 2–6 June, Krakow, Poland, 69–86.
Cook, E. R., & Holmes, R. L. (1986). User manual for ARSTAN Laboratory of Tree Ring Research. University of Arizona, Tucson.
Emberson, L. (2003). Air pollution impacts on crops and forests: An introduction. In L. Emberson, M. Ashmore, & F. Murray (Eds.), Air Pollution Impacts on Crops and Forests: A Global Assessment (pp. 3–29). Imperial College Press.
European Environment Agency (2021). https://www.eea.europa.eu/themes/air/urban-air-quality/european-city-air-quality-viewer
Erteld, W., & Hengst, E. (1966). Waldertragslehre. Neumann Verlag.
Ferrio, J., Voltas, J., & Araus, J. (2003). Use of carbon isotope composition in monitoring environmental changes. Management of Environmental Quality, 14, 82–98.
Holmes, R. L. (1983). Computer−assisted quality control in tree-ring dating and measurement. Tree-Ring Bulletin, 43, 69–78.
Juknys, R., Stravinskiene, V., & Vencloviene, J. (2002). Tree-ring analysis for the assessment of anthropogenic changes and trends. Environmental Monitoring and Assessment, 77, 81–97.
Kaleta, D. (2014). State of air pollution in Silesia province including low emission sources. Architecture Civil Engineering Environment, 7(4), 79–87.
Keeling, C. D. (1973). Industrial production of carbon dioxide from fossil fuel and limestone. Tellus, 25, 174–198.
Krąpiec, M., & Szychowska-Krąpiec, E. (2001). Tree-ring estimation of the effect of industrial pollution on pine (Pinus sylvestris) and fir (Abies alba) in the Ojców National Park (southern Poland). Nature Conservation, 58(1), 33–42.
L’Hirondelle, S. J., & Addison, P. A. (1985). Effects of SO2 on leaf conductance, Xylem tension, Fructose and sulphur levels of Jack pine seedlings. EnvironmentalPollution, 39(A), 373–386.
Malik, I., Danek, M., Marchwińska-Wyrwał, E., Danek, T., Wistuba, M., & Krąpiec, M. (2012). Scots pine (Pinus sylvestris L.) growth suppression and adverse effects on human health due to air pollution in the Upper Silesian Industrial District (USID), southern Poland. Water, Air and Soil Pollution, 223, 3345–3364.
McLaughlin, S. B., Shortle, W. C., & Smith, K. T. (2002). Dendroecological applications in air pollution and environmental chemistry: Research needs. Dendrochronologia, 20(1–2), 133–157.
Nöjd, P., & Reams, G. A. (1996). Growth variation of Scots pine across a pollution gradient on the Kola Peninsula, Russia. Environmental Pollution, 93, 313–325.
Park, R., & Epstein, S. (1961). Metabolic fractionation of C13 & C12 in plants. Plant Physiology, 36(2), 133.
Pazdur, A., Kuc, T., Pawełczyk, S., Piotrowska, N., Sensuła, B. M., & Różański, K. (2013). Carbon isotope composition of atmospheric carbon dioxide in southern Poland: Imprint of anthropogenic CO2 emissions in regional biosphere. Radiocarbon, 55(2–3), 848–864.
Percy, K. E., & Ferretti, M. (2004). Air pollution and forest health: Toward new monitoring concepts. Environmental Pollution, 130(1), 113–126.
Piotrowska, N. (2013). Status report of AMS sample preparation laboratory at GADAM Centre, Gliwice, Poland. Nuclear Instruments and Methods in Physics Research Section B, 294, 176–181.
Rakowski, A. (2011). Radiocarbon method in monitoring of fossil fuel emission. Geochronometria, 38(4), 314–332.
Saurer, M., & Siegwolf, R. (2007). Human impacts on tree-ring growth reconstructed from stable isotopes. In T. E. Dawson & R. T. F. Siegwolf (Eds.), Stable Isotopes as Indicators of Ecological Change (pp. 49–62). Terrestrial Ecology Series, Elsevier, Amsterdam.
Saurer, M., Siegwolf, R., & Schweingruber, F. (2004). Carbon isotope discrimination indicates improving water-use efficiency of trees in northern Eurasia over the last 100 years. Global Change Biology, 10, 2109–2120. https://doi.org/10.1111/j.1365-2486.2004.00869.x
Scheidegger, Y., Saurer, M., Bahn, M., & Siegwolf, R. (2000). Linking stable oxygen and carbon isotopes with stomatal conductance and photosynthetic capacity: A conceptual model. Oecologia, 125, 350–357.
Sensuła, B., & Pazdur, A. (2013). Stable carbon isotopes of glucose received from pine tree-rings as bioindicators of local industrial emission of CO2 in Niepolomice Forest (1950–2000). Isotopes in Environmental and Health Studie, 49(4), 532–541.
Sensuła, B., & Piotrowska, N. (2020). Carbon isotopic research of Pinus Sylvestris L. Growing in the Southern Poland (near Kędzierzyn-Koźle, Dąbrowa Górnicza Katowice and Olesno). Radiocarbon, 62(2), 497–502. https://doi.org/10.1017/RDC.2019.130
Sensuła, B., & Wilczyński, S. (2017). Climatic signals in tree-ring width and stable isotopes composition of Pinus sylvestris L. growing in the industrialized area nearby Kędzierzyn-Koźle. Geochronometria, 44, 240–255. https://doi.org/10.1515/geochr-2015-0070
Sensuła, B., & Wilczyński, S. (2018). Tree-ring widths and the stable isotope composition of pine tree-rings as climate indicators in the most industrialised part of Poland during CO2 elevation. Geochronometria, 45, 130–145. https://doi.org/10.1515/geochr-2015-0094
Sensuła, B., Pazdur, A., Bickerton, J., & Derrick, P. J. (2011). Probing palaeoclimatology through quantitation by mass spectrometry of the products of enzyme hydrolysis of alpha-cellulose. Cellulose, 18(2), 461–468.
Sensuła, B., Opała, M., Wilczyński, S., & Pawełczyk, S. (2015). Long- and short-term incremental response of Pinus sylvestris L. from industrial area nearby steelworks in Silesian Upland. Poland. Dendrochronologia, 36, 1–12. https://doi.org/10.1016/j.dendro.2015.08.001
Sensuła, B., Wilczynski, S., Opala, M. (2015b). Tree growth and climate relationship: Dynamics of Scots pine (Pinus Sylvestris L.) growing in the near-source region of the combined heat and power plant during the development of the pro-ecological strategy in Poland. Water Air and Soil Pollution, 226, 7, https://doi.org/10.1007/s11270-015-2477-4
Sensuła, B., Wilczyński, S., Monin, L., Allan, M., Pazdur, A., & Fagel, N. (2017). Variations of tree ring width and chemical composition of wood of pine growing in the area nearby chemical factories. Geochronometria, 44, 226–239. https://doi.org/10.1515/Geochr-2015-0064
Sensuła, B., Michczyński, A., Piotrowska, N., & Wilczyński, S. (2018). Anthropogenic CO2 Emission Records in Scots Pine Growing in the Most Industrialized Region of Poland from 1975 to 2014. Radiocarbon, 60(4), 1041–1053.
Stravinskiene, V., Bartkevicius, E., & Plausinyte, E. (2013). Dendrochronological research of Scots pine (Pinus sylvestris L.) radial growth in vicinity of industrial pollution. Dendrochronologia, 31, 179–186.
Suess, H. E. (1955). Radiocarbon concentration in modern wood. Science, 122(3166), 415–417.
van der Plicht, J., & Hogg, A. (2006). A note on reporting radiocarbon. Quaternary Geochronology, 1(4), 237–240.
White, J.W.C., Vaughn, B.H. & Michel, S.E. (2015). University of Colorado, Institute of Arctic and Alpine Research (INSTAAR), Stable isotopic composition of atmospheric carbon dioxide (13C and 18O) from the NOAA ESRL Carbon Cycle Cooperative Global Air Sampling Network, 1990–2014, Path: https://aftp.cmdl.noaa.gov/data/trace_gases/co2c13/flask/
Wilczyński, S. (2006). The variation of tree-ring widths of Scots pine (Pinus sylvestris L.) affected by air pollution. European Journal of Forest Research, 125(3), 213–219. https://doi.org/10.1007/s10342-005-0106-2
Wilczyński, S. (2020). Record of the changes in environment by Scots pine and Jack pine trees. Sylwan, 164(7), 583–593. https://doi.org/10.26202/sylwan.2020040
Zimnoch, M., Jelen, D., Galkowski, M., Kuc, T., Necki, J., Chmura, L., Gorczyca, Z., Jasek, A., & Różański, K. (2012). Partitioning of atmospheric carbon dioxide over central Europe: Composition. Isotopes in Environmental and Health Studies, 48(3), 421–433.
Acknowledgements
This project was a part of the BIOPOL project entitled “Trees as bioindicators of industrial air pollution during implementation of the pro-environmental policy in the Silesia region,” funded by the National Science Centre and allocated on the basis of decision number DEC-2011/03/D/ST10/05251. The authors express their gratitude to everyone from the Silesian University of Technology who contributed to making these investigations possible.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Sensuła, B., Wilczyński, S. Records of Anthropogenic Pollution in Silesia Captured in Scots Pine Tree Rings: Analysis by Radiocarbon, Stable Isotopes, and Basal Area Increment Analysis. Water Air Soil Pollut 233, 143 (2022). https://doi.org/10.1007/s11270-022-05617-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11270-022-05617-6