
Abstract
Plant species diversity could enhance plant productivity and pollutant removal efficiency in constructed wetlands (CWs). However, the potential importance of plant density for ecosystem functioning has largely been neglected. In this study, we conducted a factorial experiment in which three common plant species were planted in a gradient of species richness (one, two, and three) and seven species compositions at two densities (six and twelve individuals per microcosm). Plant total biomass and total organic carbon (TOC) and total inorganic nitrogen (TIN) removal efficiency were measured to explore the effect of plant species diversity and density on the ecosystem functioning of CWs. Results showed that (1) plant species richness had no significant effect on plant total biomass and TOC and TIN removal efficiency under high and low plant density. (2) There were significant differences in TIN removal efficiency among seven species compositions under low plant density; especially, the presence of Canna indica reduced the TIN removal efficiency. In contrast, species composition and species identity had no significant effect on ecosystem functioning under high plant density. (3) High plant density increased plant total biomass of C. indica monocultures, and also enhanced TIN removal efficiency in mixtures of two species. These results indicated C. indica alone may not be an ideal species for enhancing pollutant removal in constructed wetlands but planting at high density could mitigate its negative effect on ecosystem functioning.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Rapid growth of urbanization and industrial development are often associated with substantial wastewater production that has become a serious problem affecting water quality and human health (Galloway et al. 2008; Gu et al. 2013). Excessive nitrogen (N) loading to aquatic ecosystems may contribute to nutrient enrichment or eutrophication (Conley et al. 2009; Chen et al. 2011). Untreated organic carbon often depletes dissolved oxygen in aquatic ecosystems, causing widespread mortality of aquatic organisms (Saeed and Sun 2012). The treatment of wastewater with high N and organic carbon loading remains a challenge around the world.
As a green treatment technology, constructed wetlands (CWs) have been widely used for wastewater treatment due to low operational and maintenance requirements (Liu et al. 2012; Vymazal 2014). Nitrogen and organic carbon removal are the major target services of CWs (Caselles-Osorio and García 2006; Wu et al., 2014). Nitrification, denitrification, and plant uptake are the major pathway for nitrogen removal in CWs (Vymazal et al., 2007; Saeed and Sun 2012); aerobic degradation and anaerobic degradation are the major pathway for organic removal in CWs (Saeed and Sun 2012). Plants as the important component of CWs could direct uptake of nitrogen from the wastewater; plants also indirectly influence nitrogen and organic removal through providing oxygen, carbon source, and habitats to microorganisms (Brix 1994; Picek et al., 2007; Vymazal 2013). As plant species may have specific nutrient uptake ability and growth pattern, the activity and relative abundance of rhizosphere microbial communities are also species-dependent (Hartmann et al. 2009). Such a difference may affect the pollutant removal efficiency in the CWs. Optimizing the plant community structure of CWs is an important aspect to enhance nitrogen and organic removal efficiency.
Plant diversity is perhaps a major determinant of ecosystem functions (Tilman et al. 2014). The effect of plant diversity on ecosystem functioning in CWs have gained tremendous attention in recent years (Engelhardt and Ritchie 2002; Fraser et al. 2004; Han et al. 2016; Luo et al. 2020). In CWs, plant species richness often positively affects plant productivity (Engelhardt and Ritchie 2002; Han et al. 2016) and the removal efficiencies of pollutants (Han et al. 2018; Geng et al. 2019). Species composition and identity also affected ecosystem functioning of CWs (Ge et al. 2015; Han et al. 2019). The presence of Coix lacryma-jobi increased both plant biomass and N removal, whereas the presence of Aeschynomene indica increased N removal but decreased plant biomass (Ge et al. 2015). However, initial plant density was kept constant at a particular level in previous experiments, and the potential importance of plant density for ecosystem functioning has largely been neglected.
As an important aspect of plant community structure, plant density determines the intensity competition for available resources and space acquisition (Wei et al. 2018; Hecht et al. 2019). Changing plant density may also modify any biodiversity effect via changed plant-plant interactions (He et al. 2005; Polley et al. 2010; Stachova et al. 2013). For example, the diversity-productivity relationship can be flatter at higher than at lower density in grassland (He et al. 2005). In CWs, the detention time of CWs may increase with the increase of plant density (Jaddhav and Buchberger 1995). However, the effect of plant density on the ecosystem functioning (such as plant biomass and N and organic carbon removal efficiency) and the relationship of plant species diversity and ecosystem functioning in CWs remain uncertain.
In this study, microcosms were established to simulate CWs and were fed with simulated wastewater with high N and organic carbon loading. Three common species were planted in a gradient of species richness (one, two, and three) and seven species compositions at each of two densities (six and twelve individuals per microcosm). Our objectives were to (1) test the effects of plant density and plant species diversity on plant biomass and N and organic carbon removal efficiency; and (2) determine a combination of plant species and plant density that provide high ecosystem functioning in CWs.
2 Materials and Methods
2.1 Experiment Design
The experiment was set up in a semi-open greenhouse at the campus of Zhejiang Normal University (29° 8′ 19″ N, 119° 38′ 2″ E, Jinhua City, Zhejiang province, China). The greenhouse with a transparent plastic roof was used to keep off rainwater, and no walls were used to ensure that light intensity, air temperature, and humidity inside the greenhouse were similar to those of the external environment. A total of 42 microcosms were constructed using ceramic tubs (45 cm length × 34 cm width × 20 cm height) and a piece of polyethylene planting bed in March 2019 (Fig. 1).

The vertical cross sections of the microcosms of constructed wetlands (45 cm length × 34 cm width × 20 cm height)
A two-factor experiment design was used in this study. One factor was plant density. One factor was plant density, with 21 microcosms planted with twelve individuals (78 plants m−2, high density), another 21 microcosms planted with six individuals (39 plants m−2, low density). Under each plant density treatment, plant diversity treatments included three species richness levels (one, two, and three) and all possible species compositions: three monocultures, three two-species mixtures, and one three-species mixture. Three true replicates were established for each density and composition treatment.
Three local species with similar size and as the dominate species were selected for the experiment: Oenanthe javanica (Blume) DC, Acorus calamus L., and Canna indica L. These three species have been widely used in the construction of CWs (Vymazal 2011, 2013). In March of 2019, seedlings of these three species of similar size (10 cm height) were transplanted into the microcosms. According to the plant density and diversity treatments, an equal number of individuals were assigned to each species and uniformly distributed in each microcosm. The experiment was conducted using a randomized complete block design with three blocks (three replicates per treatment).
The simulated wastewater was the Hoagland nutrient solution (Hoagland and Arnon 1950) with a minor modification (Table 1). The N concentration was 112 mg L−1 with mixed ammonium-N and nitrate-N (NH4+-N: NO3−-N = 1:1). Nutrient removal efficiency has been shown to peak in CWs when the chemical oxygen demand (COD)/N ratio was five (Wu et al. 2009). Accordingly, sucrose was added to create this COD to N ratio (5:1). The simulated wastewater (23 L) was supplied once every 10 days from late March to early June, simulating the intermittent water operation mode of CWs (Faulwetter et al. 2009). Before each irrigation event, all microcosms were emptied of water.
2.2 Sampling and Measurements
Effluent samples (250 ml) were collected in each microcosm on the tenth day after the last supply of simulated wastewater (9 June 2019) and were stored at − 18 °C. Each water sample was filtered using a membrane syringe filter (pore size 0.45 μm) before analysis. The total organic carbon (TOC) concentration in effluent was measured by the non-dispersion infrared method with a total organic carbon analyzer (Torch TOC, Teledyne Tekmar Corporation, USA). The NH4+-N and the NO3−-N were measured by spectrophotometric method (Clescerl et al. 1999). The sum of NH4+-N and the NO3−-N is the total inorganic nitrogen (TIN) concentration. Following water sampling, all plants were harvested by species, cleaned, and oven dried at 65 °C for 72 h. Then, plant materials were weighed for total biomass per species.
2.3 Parameter Calculations
The TOC and N removal efficiency (RE) in the last batch of simulated wastewater was calculated based on mass balance:
where Ci is the concentration of influent TOC or TIN of each microcosm (mg L−1); Ce is the concentration of effluent TOC or TIN of each microcosm (mg L−1).
2.4 Statistical Analysis
Two-way ANOVA was applied to test the effects of plant species diversity (species richness and species composition) and plant density on the variables (total plant biomass, TOC removal efficiency, and N removal efficiency). General linear regression analysis was performed to address potential relationships between species richness and variables under high and low plant density treatments. One-way ANOVA was applied to test for difference in variables among seven plant species compositions under the same plant density treatment. If the treatment effects proved to be significant, Tukey’s method was used to ascertain differences among treatment levels. An independent t test was used to find differences between the means of variables when a species was present and when it was absent in the system. An independent t test was also used to detect the difference in variables between high and low plant density treatments. All the statistical analyses were performed with R ver. 3.4.3 (R Core Team 2017). We set the statistical significance level at α = 0.05. All values were expressed as mean ± standard error.
3 Results
3.1 Plant Total Biomass
There was no significant correlation between plant species richness and plant total biomass under high and low plant density treatments (Table 1 (a); Fig. 2a). Species composition also had no significant effect on plant total biomass (Table 1 (b); Fig. 3a), but species identity had significant effect (Table 2). Under the low plant density treatment, the presence of O. javanica significantly increased plant total biomass (Table 2 (a)); but the presence of any species did not affect plant total biomass under the high plant density treatment (Table 2 (b)). Plant density only significantly affected plant total biomass for one of the species: the plant total biomass of C. indica monoculture under high plant density was significantly higher than that of low plant density (Fig. 3a).

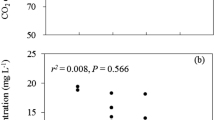
Relationships between plant species richness and plant total biomass (a), TOC removal efficiency (b), and TIN removal efficiency (c). Closed circles: high plant density; open circles: low plant density. Asterisks denote significant difference between high and low plant density treatments (P < 0.05)

Plant total biomass (a), TOC removal efficiency (b), and TIN removal efficiency (c) in relation to plant community. Species abbreviations are: Oj, O. javanica; Ac, A. calamus; Ci, C. indica. Black bars: high plant density; white bars: low plant density. Values are mean ± SE. Significant difference is indicated by different letters (P < 0.05)
3.2 TOC Removal Efficiency
Plant species richness and plant density both had no significant effect on TOC removal efficiency (Table 1; Fig. 2b). Likewise, plant species composition and species identity both had no significant effect on TOC removal efficiency under both high and low plant density treatments (Fig. 3b; Tables 1 and 2).
3.3 TIN Removal Efficiency
Plant species richness had no significant effect on TIN removal efficiency under both high and low plant density treatments (Table 1; Fig. 2c). In the species richness level of two, TIN removal efficiency under high plant density treatment was significantly higher than that of the low plant density treatment, but no significant difference was found in the species richness levels of one and three (Fig. 2c).
Species composition significantly affected TIN removal efficiency (Table 1; Fig. 3c). The TIN removal efficiency of A. calamus monoculture was significantly higher than that of C. indica monoculture (Fig. 3c), and the presence of C. indica led to significantly less efficient TIN removal under low plant density treatment (Table 2). However, no significant difference was found under high plant density treatment (Fig. 3c). Plant density significantly affected TIN removal efficiency (Table 1), and the TIN removal of O. javanica monoculture was more efficient under low than high plant density (Fig. 3c).
4 Discussion
4.1 Effects of Plant Species Diversity and Density on Plant Total Biomass
lant species diversity, as a key determinant of ecosystem functioning, has been studied for some decade years (Tilman et al. 2014; van der Plas 2019), while plant density has been known longer for its role in community productivity (Harper 1977). When sowing density is included as a factor in biodiversity studies, density can affect ecosystem functioning and may also alter the direction of diversity effects (He et al. 2005; Polley et al. 2010; Stachova et al. 2013). However, our study showed plant species richness had no significant effect on plant total biomass after 3 months under both high and low density (Fig. 2). The reason may be previous studies were conducted in grassland, in which density may enhance intraspecific competition for soil resources and alter diversity-productivity relationships (He et al. 2005; Li et al. 2011). In contrast, nutrient competition would be reduced in CWs ecosystems with high N loading, especially in the ecosystem supplied with equal NH4+-N and NO3−-N. Our previous study also found no significant correlations between plant total biomass and species richness under mixed NH4+-N/NO3−-N treatment (Ge et al. 2015). In addition, this study only used three species, which would limit the species richness effect.
Plant species identity is thought to be an important component that affects ecosystem functioning (Abalos et al. 2014; Ge et al. 2015; Han et al. 2019). In this study, we found plant density altered the effect of plant species identity on plant total biomass (Table 2). The presence of O. javanica significantly increased plant total biomass under low plant density treatment (Table 2a), but no significant effect was found under the high plant density treatment (Table 2 (b)). The reason may be that O. javanica yielded the highest plant total biomass among three monocultures (Fig. 3), and also was a dominant species in mixtures, thus increasing community productivity under low plant density. High plant density could affect the species identity effect on productivity through changing species abundance and performances. For example, high plant density increased plant total biomass of C. indica (Fig. 3). In addition, competitive interactions between species can increase under high plant density (Li et al. 2011) and then weaken the positive effect of O. javanica on community productivity. These results indicated plant density may in part compensate for an effect of plant species identity on productivity.
4.2 Effects of Plant Species Diversity and Density on TOC and TIN Removal Efficiency
N removal is one of the major objectives of CWs (Vymazal 2014). Previous studies have reported plant species diversity had positive effect on N removal efficiency in CWs (Ge et al. 2015; Han et al. 2016; Geng et al. 2019), but plant density was kept constant at a particular level in these experiments. Our study is the first focused on the effect of plant species diversity and plant density simultaneously on N removal efficiency in CWs with high N loading. We found plant species richness had no significant effect on TIN removal efficiency under both high and low plant density (Fig. 2). It is possible that the chemical oxygen demand (COD/N = 5) in this study was at a level that makes N removal less sensitive to species richness. Although this ratio leads to efficient N removal (Wu et al. 2009), the addition of carbon may have as much role in improving N removal as does increasing plant species richness (Han et al. 2016). In addition, we only set up three plant species richness levels (one, two, and three), which may limit the effect of plant species richness on N removal. Notably, high plant density increased TIN removal within the species richness level of two (Fig. 3). Denitrification and plant uptake are the major pathways of N removal from wastewater in CWs without substrate (Vymazal 2007; Saeed and Sun 2012). In this study, plant density had no significant effect on plant total biomass within any species richness level (Fig. 2), indicating that plant uptake was not the dominant cause for high plant density enhancing N removal in two-species treatments. High plant density may increase root length density and specific root length (Hecht et al. 2019) and provide more microbial attachment sites, then enhancing denitrification and N removal efficiency in CWs.
Plant species identity is also an important component that affects pollutant removal efficiency in CWs (Ge et al. 2015; Han et al. 2016). In this study, the presence of C. indica significantly decreased TIN removal efficiency under low plant density treatment (Table 2 (a)) but had no significant effect under high plant density treatment (Table 2 (b)). Additionally, C. indica yielded the lowest plant total biomass in monocultures under low plant density (Fig. 3). Logically, low biomass production may decrease the utilization of underground nutrient resources (Palmborg et al. 2005). High plant density could increase the plant total biomass of C. indica (Fig. 3) and then improve the N removal efficiency.
Organic carbon removal is also the target ecosystem service of CWs (Wu et al., 2014). In this study, we found plant species diversity and density both had no significant effect on TOC removal efficiency (Fig. 2; Table 2). Organic carbon removal mechanisms in CWs include aerobic degradation and anaerobic degradation (Saeed and Sun 2012). Availability of oxygen was one of the major environmental factors influencing organic carbon removal. Oxygen for aerobic degradation can be supplied by atmospheric oxygen diffusion, and/or macrophyte root transfer into the plant rhizosphere (Cooper et al. 1996). The three species, O. javanica, A. calamus, and C. indica, all had a well-developed aerenchyma tissue which could transfer oxygen into the plant rhizosphere and enhance aerobic degradation (Stottmeister et al. 2003). Further study should select additional species to explore the response of TOC removal efficiency to plant species diversity and density.
4.3 Recommendations for Achieving High Net Ecosystem Services of CWs
Nitrogen and organic carbon removal are the major target services of CWs (Caselles-Osorio and García 2006; Wu et al., 2014); plant biomass also could be used as biofuels (Liu et al. 2012). In this study, plant species richness did not affect plant biomass and nitrogen and organic carbon removal under both high and low plant density treatment (Fig. 2). Nevertheless, under low plant density, plant species composition and species identity significantly affected these functions (Fig. 3 and Table 2). Among seven species compositions, C. indica showed the lowest TIN removal efficiency (Fig. 3), the total biomass and TOC removal efficiency also lower than other species compositions (Fig. 3). The presence of C. indica also decreased the TIN removal efficiency (Table 2). But no significant difference in these functions was found under high plant density. These results indicate that C. indica alone may not be ideal species for CWs, planting high plant density could mitigate its negative effect on ecosystem functioning. It is worth noting that the presence of O. javanica significantly increased plant total biomass (Table 2); the TOC and TIN removal efficiency of O. javanica monoculture were not lower than other species compositions (Fig. 3). These results indicate O. javanica may be the ideal species for CWs. Of course, only two levels of plant density were explored in this study, and plant mortality would increase at higher plant densities (Weiner et al. 2001). In addition, our study was controlled experiment; the results may do not reflect their performance in natural environment. Further research should carry out experiments across a wider density range to explore the proper density and plant species diversity to maintain high ecosystem multifunctionality in real CWs with high N and organic carbon loading.
5 Conclusions
This study was the first to explore the effect of plant species diversity and plant density on ecosystem functioning in CWs with high N loading. We found plant density altered the relationship of plant species diversity and ecosystem functioning. Under low plant density, there were significant differences in TIN removal efficiency among seven species compositions; the presence of C. indica had a negative effect on TIN removal efficiency. However, plant species richness, species composition, and species identity all had no significant effect on ecosystem functions under high plant density. C. indica alone may not be an ideal species for enhancing ecosystem functioning but planting high plant density could mitigate its negative effect on ecosystem functioning. Further research should investigate additional plant species and plant density to find more appropriate species and plant density for ecosystem functioning in CWs with high N and organic carbon loading.
References
Abalos, D., Deyn, G. B., Kuyper, T. W., & van Groenigen, J. W. (2014). Plant species identity surpasses species richness as a key driver of N2O emissions from grassland. Global Change Biology, 20, 265–275.
Brix, H. (1994). Functions of macrophytes in constructed wetlands. Water Sciences Technology, 29, 71–78.
Caselles-Osorio, A., & García, J. (2006). Performance of experimental horizontal subsurface flow constructed wetlands fed with dissolved or particulate organic matter. Water Research, 40, 3603–3611.
Chen, Y., Wen, Y., Cheng, J., Xue, C. H., & Zhou, Q. (2011). Effects of dissolved oxygen on extracellular enzymes activities and transformation of carbon sources from plant biomass: implications for denitrification in constructed wetlands. Bioresource Technology, 102, 2433–2440.
Clescerl, L. S., Greenberg, A. E., & Eaton, A. D. (1999). Standard method for the examination of water and wastewater. American Washington: American Public Health Association.
Conley, D. J., Paerl, H. W., Howarth, R. W., Boesch, D. F., Seitzinger, S. P., & Havens, K. E. (2009). Controlling eutrophication: nitrogen and phosphorus. Science, 323, 1014–1015.
Engelhardt, K. A. M., & Ritchie, M. E. (2002). The effect of aquatic plant species richness on wetland ecosystem processes. Ecology, 83, 2911–2914.
Faulwetter, J. L., Gagnon, V., Sundberg, C., Chazarenc, F., Burr, M. D., Brisson, J., et al. (2009). Microbial processes influencing performance of treatment wetlands: a review. Ecological Engineering, 35, 987–1004.
Fraser, L. H., Carty, S. M., & Steer, D. (2004). A test of four plant species to reduce total nitrogen and total phosphorus from soil leachate in subsurface wetland microcosms. Bioresource Technology, 94, 185–192.
Galloway, J. N., Townsend, A. R., Erisman, J. W., Bekunda, M., Cai, Z. C., Freney, J. R., et al. (2008). Transformation of the nitrogen cycle: recent trends, questions, and potential solutions. Science, 320, 889–892.
Ge, Y., Han, W. J., Huang, C. C., Wang, H., Liu, D., Chang, S. X., et al. (2015). Positive effects of plant diversity on nitrogen removal in microcosms of constructed wetlands with high ammonium loading. Ecological Engineering, 82, 614–623.
Geng Y., Ge Y., Luo B., Chen Z.X., Min Y., Schmid B., et al. (2019) Plant diversity increases N removal in constructed wetlands when multiple rather than single N processes are considered. Ecological Applications, e01965.
Gu, B. J., Leach, A. M., Ma, L., Galloway, J. M., Chang, S. X., Ge, Y., et al. (2013). Nitrogen footprint in China: food, energy, and nonfood goods. Environmental Science and Technology, 47, 9217–9224.
Han, W. J., Chang, J., Fan, X., Du, Y. Y., Chang, S. X., et al. (2016). Plant species diversity impacts nitrogen removal and nitrous oxide emissions as much as carbon addition in constructed wetland microcosms. Ecological Engineering, 93, 144–151.
Han, W. J., Ge, Y., Ren, Y., Luo, B., Du, Y. Y., Chang, J., et al. (2018). Removal of metals and their pools in plant in response to plant diversity in microcosms of floating constructed wetlands. Ecological Engineering, 113, 65–73.
Han, W. J., Luo, G. Y., Luo, B., Yu, C. C., Wang, H., Chang, J., et al. (2019). Effects of plant diversity on greenhouse gas emissions in microcosms simulating vertical constructed wetlands with high ammonium loading. Journal of Environmental Sciences, 77, 229–237.
Harper, J. L. (1977). Population biology of plants. New York: Academic Press.
Hartmann, A., Schmid, M., Tuinen, D. V., & Berg, G. (2009). Plant-driven selection of microbes. Plant and Soil, 321, 235–257.
He, J. S., Wolfe-Bellin, K. S., Schmid, B., & Bazzaz, F. A. (2005). Density may alter diversity–productivity relationships in experimental plant communities. Basic and Applied Ecology, 6, 505–517.
Hecht, V. L., Temperton, V. M., Nagel, K. A., Pude, R. R., & Postma, J. A. (2019). Plant density modifies root system architecture in spring barley (Hordeum vulgare L.) through a change in nodal root number. Plant and Soil, 439, 179–200.
Hoagland, D. R., & Arnon, D. J. (1950). The water culture method for growing plants without soil. California Agriculture, 347, 1–32.
Jaddhav, R. S., & Buchberger, S. G. (1995). Effects of vegetation on flow through free water surface wetlands. Ecological Engineering, 5, 481–496.
Li, A., Niu, K. C., & Du, G. Z. (2011). Resource availability, species composition and sown density effects on productivity of experimental plant communities. Plant and Soil, 344, 177–186.
Liu, D., Wu, X., Chang, J., Gu, B. J., Min, Y., Ge, Y., et al. (2012). Constructed wetlands as biofuel production systems. Nature Climate Change, 2, 190–194.
Luo, B., Du, Y. Y., Han, W. J., Geng, Y., Wang, Q., Duan, Y. Y., et al. (2020). Reduce health damage cost of greenhouse gas and ammonia emissions by assembling plant diversity in floating constructed wetlands treating wastewater. Journal of Cleaner Production, 244, 118927.
Polley, H. W., Wilsey, B. J., & Derner, J. D. (2010). Do species evenness and plant density influence the magnitude of selection and complementarity effects in annual plant species mixtures? Ecology Letters, 6, 248–256.
R Core Team (2017) R: A Language and Environment for Statistical Computing. Vienna, Austria: R Foundation for Statistical Computing. https://www.R-project.org/ (30 November 2017, date last accessed).
Saeed, T., & Sun, G. (2012). A review on nitrogen and organics removal mechanisms in subsurface flow constructed wetlands: dependency on environmental parameters, operating conditions and supporting media. Journal of Environment Management, 112, 429–448.
Stachova, T., Fibich, P., & Leps, J. (2013). Plant density affects measures of biodiversity effects. Journal of Plant Ecology, 6, 1–11.
Stottmeister, U., Wiessner, A., Kuschk, P., Kappelmeyer, U., Kaestner, M., et al. (2003). Effects of plants and microorganisms in constructed wetlands for wastewater treatment. Biotechnology Advance, 22, 93–117.
Tilman, D., Isbell, F., & Cowles, M. J. (2014). Biodiversity and ecosystem functioning. Annual Review of Ecology and Systematics, 45, 471–493.
van der Plas, F. (2019). Biodiversity and ecosystem functioning in naturally assembled communities. Biological Reviews, 94, 1220–1245.
Vymazal, J. (2007). Removal of nutrients in various types of constructed wetlands. Science of Total Environment, 380, 48–65.
Vymazal, J. (2011). Plants used in constructed wetlands with horizontal subsurface flow: a review. Hydrobiologia, 674, 133–156.
Vymazal, J. (2013). Emergent plants used in free water surface constructed wetlands: a review. Ecological Engineering, 61, 582–592.
Vymazal, J. (2014). Constructed wetlands for treatment of industrial wastewaters: a review. Ecological Engineering, 73, 724–751.
Wei, X., Martijn, B. T., & Frank, B. (2018). Density-dependency and plant-soil feedback: former plant abundance influences competitive interactions between two grassland plant species through plant-soil feedbacks. Plant and Soil, 428, 441–452.
Weiner, J., Stoll, P., & Muller-Landau, H. (2001). The effects of density, spatial pattern and competitive symmetry on size variation in simulated plant populations. The American Naturalist, 158, 438–450.
Wu, J., Zhang, J., Jia, W. L., Xie, H. J., Gu, R. R., Li, C., et al. (2009). Impact of COD/N ratio on nitrous oxide emission from microcosm wetlands and their performance in removing nitrogen from wastewater. Bioresource Technology, 100, 2910–2917.
Funding
This work was funded by the National Natural Science Foundation of China (31901213) and the Natural Science Foundation of Zhejiang Province (LQ18C030001).
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
ESM 1
(DOCX 14 kb)
Rights and permissions
About this article

Cite this article
Han, W., Sheng, X., Shao, J. et al. Effects of Plant Diversity and Plant Density on Ecosystem Functions in Floating Constructed Wetlands. Water Air Soil Pollut 231, 544 (2020). https://doi.org/10.1007/s11270-020-04913-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11270-020-04913-3