
Abstract
The ultimate goal of threatened plant translocations is to establish self-sustaining populations. Lessons learnt from plant translocations can inform future translocations and improve conservation outcomes via adaptive management. We assessed translocation success for 76 translocations of 50 species established as part of recovery programmes between 1998 and 2016 in the Southwest Australian Floristic Region (SWAFR), a biodiversity hotspot with Mediterranean-type climate. We used a series of Bayesian hierarchical models to assess translocation success as measured by plant survival, height growth, crown growth, and reproduction (flowering and fruiting). We found that fencing to prevent vertebrate herbivory improved survival, irrigating plants over the first two summer dry periods improved the likelihood of reproduction, and where the rainfall increased above the 30-year average in the two years following planting, it positively influenced survival, growth, and reproduction. We recommend that fencing to prevent herbivory should be broadly considered in threatened plant translocations, while irrigation to improve translocation success would benefit plant species in Mediterranean-type ecosystems and other dryland environments.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
As the number of threatened plant species increases globally (IUCN 2022), the use of translocations as part of efforts to prevent extinction has steadily gained momentum (Dalrymple et al. 2012; Silcock et al. 2019). Translocation, the deliberate transfer of plants or regenerative plant material from one area to another (Silcock et al. 2019; IUCN/SSC 2013), is used to augment small or declining populations, re-establish extinct populations or establish new populations in places where a species has never been recorded. Additionally, as the impacts of climate change have become evident, there is the option to use translocation to move species beyond their current range to track their preferred climate (i.e. ‘assisted colonisation’; Hoegh-Guldberg et al. 2008; Seddon et al. 2009; Kreyling et al. 2011; Burbidge et al. 2011). While the practice of translocation is increasing (Guerrant 2012), the outcomes of these actions remain under-reported in the published literature (Godefroid et al. 2011), with a clear bias towards only publishing successful outcomes (Menges et al. 2016; Godefroid et al. 2011; Drayton and Primack 2012). Furthermore, the results of many studies go unreported due to the difficulties of publishing when sample sizes are small, which frequently occurs with threatened species. Thus, despite the increasing practice of translocation, there is still limited knowledge about successful techniques, rates of success, and whether translocation is a viable conservation strategy (Drayton and Primack 2012; Silcock et al. 2019).
There is broad support that the success of translocations should be considered in the context of plant species abundance, extent, persistence, and resilience (Pavlik 1996; Guerrant 2013). Metrics for determining success include survival of propagules, growth, reproductive output, recruitment of subsequent generations, measures of genetic diversity, and recovery from disturbance, as these are likely to offer the most insight into the likelihood of establishing a resilient, self-sustaining population (Pavlik 1996; Guerrant 1996; Menges 2008; Guerrant and Kaye 2007). However, due to the short time frames over which many translocations are reported, survival of translocated propagules is frequently the most common, and often the only, measure of success (Godefroid et al. 2011; Guerrant 2012; Liu et al. 2015). This can provide a skewed perspective on translocation success as initial losses are to be expected (Guerrant 2013; Albrecht et al. 2019). Moreover, timeframes that reflect the life history of the species are important as some species take many years to become reproductive (Maschinski and Duquesnel 2006; Monks et al. 2012; Albrecht et al. 2019), seedling recruitment and establishment may be sporadic or linked to disturbance events such as fire (Menges 2000; Yates and Broadhurst 2002; Wendelberger and Maschinski 2016), and initial translocation results may not necessarily reflect longer-term outcomes (Guerrant and Kaye 2007; Silcock et al. 2019).
Global reviews have provided useful generalisations regarding factors that influence translocation outcomes and contributed to debate around appropriate measures of translocation success (Godefroid et al. 2011; Guerrant 2012, 2013; Dalrymple et al. 2012). These are complemented by detailed field studies that focus on aspects of translocation techniques or success at a range of scales, with reviews at regional (Fenu et al. 2019), state (Guerrant and Kaye 2007) and country (Liu et al. 2015; Silcock et al. 2019; Abeli et al. 2021), levels providing useful insights into translocation success at these different spatial scales. Similarly, a review for plant functional groups has provided guidance on techniques for species that share regenerative and other relevant plant traits (Reiter et al. 2016). A recent review of translocations in Australia documented 1001 translocations of 376 taxa that have been established since the 1970s (Silcock et al. 2019). The number of founder propagules was found to be the primary determinant of population persistence, with populations founded with more than 500 individuals more likely to establish viable, persistent populations (Silcock et al. 2019). The influence of management interventions was explored using this same dataset and found protection from vertebrate herbivores increased survival, and prescribed fire and watering increase the probability of recruitment of second-generation plants (Whitehead et al. 2023).
Analysis at regional scales can provide further, more targeted insights without strong influence of variation in soils and climate. A region of global plant conservation significance and recognised Biodiversity Hotspot is the Southwest Australian Floristic Region (SWAFR) (Myers et al. 2000; Mittermeier et al. 2004, 2011). The region has a Mediterranean-type climate of cool wet winters and warm dry summers, and nutrient poor soils, on a flat, stable, and highly weathered low plateau (Hopper and Gioia 2004; Gosper et al. 2020). This region is noted for its extraordinary botanical diversity with 8,379 described vascular plant taxa, of which 47% are endemic to the region (Gioia and Hopper 2017). Additionally, much of the flora has a small geographic range with rapid species turnover between similar habitats over relatively short distances (Hopper and Gioia 2004; Gosper et al. 2020). This combined with a range of threatening processes including extensive habitat fragmentation, the pathogen Phytophthora cinnamomi, environmental weeds, grazing by introduced and native herbivores, altered hydrology, changed disturbance regimes and more recently climate change, has resulted in 399 taxa being listed as threatened, with over one third Critically Endangered (Monks et al. 2019). Many of the threatened species in SWAFR also occur in small, isolated populations and frequently have quite disjunct distributions (Gosper et al. 2022), and 72% of threatened flora populations occur outside the conservation reserve system (Gosper et al. 2022) where management is challenging as it can conflict with other land uses. The establishment of translocated populations of threatened flora is a key strategy being used to assist in recovering threatened plant species in the SWAFR (Monks et al. 2019), where the first recorded translocation occurred in 1985. Since 1998, plant translocations have been established in the SWAFR using an experimental framework to better understand the factors that influence translocation success (Monks and Coates 2002; Dillon et al. 2018), with translocation success measured by survival, growth, reproduction, and recruitment of second-generation plants. Key limitations for plant establishment and survival in the SWAFR, as well as other Mediterranean-type climates, are summer drought stress and herbivory (Rathbone and Barrett 2017; Fenu et al. 2016, 2019; Castro et al. 2005; López-Jurado et al. 2019) and effort has focused on techniques to reduce the impact of these factors, such as the use of irrigation during dry periods and the exclusion of herbivores.
Designing translocations in an experimental framework with specific hypotheses is likely to provide the best guidance regarding management interventions or environmental factors affecting success (Guerrant and Kaye 2007; Guerrant 2012; Menges et al. 2016). Not all factors can be experimentally tested in individual studies, especially when sample size and sufficient replication is limited making direct assessment of translocation success difficult to quantify. Synthesising many translocation studies offers an alternative approach to identify factors that may be critical in translocation success (Guerrant 2012, 2013; Dalrymple et al. 2011, 2012; Godefroid et al. 2011; Menges 2008). Here we evaluate 76 translocations established as part of recovery programmes between 1998 and 2016 in the SWAFR for 50 threatened plant taxa. Given the key limitations for plant establishment and survival in this region as well as other Mediterranean-type climates relating to drought stress and herbivory, we asked the following questions:
-
1.
Does the amount of rainfall in the month before planting, or in the two years following planting influence plant survival, growth, and reproduction?
-
2.
Does irrigation of translocated plants over first two dry seasons following planting influence translocation plant survival, growth, and reproduction?
-
3.
Does fencing to prevent herbivory influence translocation plant survival, growth, and reproduction?
Materials and methods
The study species
The 50 species included in this study are long-lived (> 20 years) perennial species, except the annual forb Schoenia filifolia subsp. subulifolia (Online Resource 1), reflecting that most of the threatened flora in SWAFR are long-lived perennials (Hopper and Gioia 2004). The species often took several years to become reproductive, with seed production commonly commencing three to four years following planting. Many are obligate seeders after fire (Hopper and Gioia 2004) and because of this we could not use recruitment in our analysis, as less than a quarter of the 76 sites had any second-generation plants, usually only a small number (Online Resource 1), and this was often due to a lack of fire to stimulate recruitment.
The 76 translocations analysed for this study were included in an Australian-wide meta-analysis undertaken by Silcock et al. (2019), who considered plant life form, habitat type, propagule type, and number of individuals planted when assessing survival and second-generation recruitment of 1001 plant translocations. These same translocations have also been included in an Australian-wide meta-analysis undertaken by Whitehead et al. (2023) who considered the effect of common management practices of fencing, watering, burning, weed control as well as habitat type and lifeform on translocation survival and recruitment. Both these studies only analysed outcomes at a single, final census. However, as we have undertaken additional monitoring of plant height, canopy width, and reproduction not included in these two studies, with multiple censuses, here we focus on factors that lead to greater plant survival, growth, and reproduction specifically for these 50 species that occur in the Mediterranean-type climate of the SWAFR.
Five species were filtered from the reproduction analysis due to missing or incomplete data due to the young age of the plants at time of analysis including Banksia brownii, Banksia cuneata, Calytrix breviseta subsp. breviseta, Daviesia ovata, and Latrobea colophona (Online Resource 1). Schoenia filifolia subsp. subulifolia was filtered from the growth analyses because we did not measure size for this annual species (Online Resource 1).
Translocation procedure
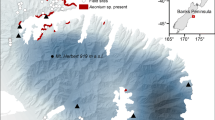
All translocations in this study were planned, planted, and monitored by the Western Australian government conservation agency (Department of Biodiversity, Conservation, and Attractions). Translocation planning usually began a year in advance of planting. Using between two and four expert opinions, translocation sites were selected based on similarity of soil, topography, and associated vegetation to wild locations of each species. Consideration was also given to sites where the land tenure was for conservation and whether threats were absent or controllable. The 76 translocation sites were spread across the SWAFR and planted between 1998 and 2018 (Fig. 1). Sites were either used for single species or multiple species, and in other cases multiple sites were used for a single species, such that there were 59 unique sites across the SWAFR (Fig. 1, Online Resource 1). In addition to some sites being used for multiple species, all sites contained varying densities of native vegetation (Online Resource 1). As all translocated species experienced some varied degree of interaction with other species (such as competition with neighbouring plants), and that individual level information about their immediate neighbourhoods was not recorded, neighbour effects were not included as a variable in the models.

Map of southwest Western Australia showing the translocation sites in this study (pink dots), the Southwest Australian Floristic Region (green area) and rainfall isohyets in mm (blue lines). Study sites and species translocated are described in Online Resource 1
Plants were propagated ex situ from seed or cutting material. Seed was wild collected and then either germinated immediately or stored at − 18 °C, 15% RH in a seedbank until required for translocation. Seed was germinated ex situ on agar (to gain viability data) before being transferred to soil and grown for approximately six months under nursery conditions. Cutting material was wild collected and then multiple plants (clones) grown from the material under nursery conditions until a sufficient root system had developed, which usually took six to twelve months. Only healthy plants were planted at the translocation sites.
Planting generally occurred in the wetter months of the year (between April and September). Plants were planted in a grid and, depending on expected adult size, between two and five metres from each other. Each plant was watered with one to two litres of water following planting and then tagged with a unique number which allowed us to track each plant individually. Monitoring of plants occurred immediately after planting (the start points for the models) and then annually thereafter where possible; however, there were breaks of several years in some cases due to funding or other logistical issues. The number of times monitored therefore varies between two and twenty-two (Online Resource 1). At each monitoring period, survival, plant height (m), crown width (m), and reproductive status were recorded for each individual. Plant height was measured as the maximum height of living canopy, crown width was the mean of widest canopy width and perpendicular to widest width of living canopy, and reproductive status was recorded as presence or absence of flowers or fruit.
Predictor variables
Four predictor variables were used in analyses: fencing, watering, rainfall in the 30 days prior to planting and the rainfall deviation from the 30-year average in the two years following planting.
Fencing is the most common management intervention used in SWAFR translocations to limit vertebrate herbivory. Fencing was carried out around either individual plants, groups of plants or the entire site, and was designed to exclude the vertebrate herbivores most common to the area where the translocation site was located, primarily rabbits (Oryctolagus cuniculus), goats (Capra hircus), and western grey kangaroos (Macropus fuliginosus). Fences were constructed of single strand wire and wire netting to 1.2 m in height. The netting also extended out 0.25 m horizontally at the base of fences, to prevent animals digging underneath. Fence size (length and width) was scaled to fit the number of plants and spatial layout and so varied between species. Cages around individual plants were made from wire netting and were 0.9 m in height and approximately 0.6 m in diameter and held in place with metal stakes.
Watering of plants was another common management intervention and was used to minimise the impact of the summer dry period (drought) in SWAFR translocations. Watering was implemented independent of whether it was likely to be a dry or wet year and independently of rainfall zones, rather it was used as resources and funding permitted. Water was applied to the root zone of plants from water tanks via a gravity-fed drip irrigation system controlled by a timer. The delivery of water was calibrated using adjustable drippers. Water was given once a week for the first two dry seasons (November to April) after planting. However, as the systems were gravity-fed, it was not possible to deliver an exact amount and, therefore, watering was assumed to deliver approximately one litre of water to irrigated plants once a week.
Rainfall, and when it occurred, was considered likely to impact translocation establishment. Planting usually occurred at the onset of reliable seasonal rainfall. However, planting sometimes occurred just prior to reliable winter rains when the soil was still dry after the summer drought, to meet financial obligations. Therefore, the impact of rainfall in the 30 days prior to planting was investigated. The amount of rainfall following planting was also considered likely to impact translocation success, as the SWAFR region can have long dry periods, especially over the hot summer period. Therefore, the deviation of rainfall (increase or decrease) in the two years following planting from the long-term mean was investigated. The two rainfall variables were collated using data from the Australian Bureau of Meteorology (BoM 2019) from the weather station geographically closest to each translocation site. A 30-year mean rainfall was calculated for the period 1970 to 1999 from the BoM climate data. Prior to 1970, rainfall in the SWAFR was 10 to 20% higher (BoM 2019); therefore, the period from 1970 to 1999 was used to calculate the average as it best represents the long-term rainfall for the sites during the period the translocations for this analysis were established and monitored.
Model development
We used a series of hierarchical models to assess translocation outcomes for all plant species. Translocation outcomes were measured by four different response variables, each modelled individually. These were the time until mortality of each individual plant (survival), height growth, and crown-width growth, and whether plants were reproductive (presence of flowers or fruit). Planting date was used as the starting point for the monitoring data used for each model.
Models for all four response variables had a similar structure, written in pseudocode: response ~ days + propagule type + predictors + (1 + predictors | species) + (1 | source pop) + (1 | site) + (1 | plant no).
These models assess the effects of age, propagule type, and the four main predictor variables (fencing, watering, rainfall deviation in two years following planting, and rainfall in 30 days prior to planting) on plant translocation outcomes. The model for rainfall deviation in the two years following planting showed a positive effect if greater than mean rainfall improved outcomes or if less than mean rainfall made outcomes worse. The rainfall deviation model showed a negative effect if less than mean rainfall improved outcomes or greater than mean rainfall made outcomes worse. Rainfall deviation and rainfall prior to planting predictors were spread consistently across fencing and watering treatments (Figure S1 Online Resource 2). The models additionally included random-effect terms to account for differences in average response among species, species-specific effects of the four main predictor variables, and differences among source populations, sites, and individual plants.
The model for survival (time until mortality) differed from the other models in two ways. First, this model did not include a random intercept for individual plants because this response (days until mortality) was observed only once for each individual. Second, the survival model required an additional term to account for censored observations, that is, plants that were recorded alive at their most recent survey.
We used different error distributions for each response variable. Survival (days until mortality) was assumed to follow a Weibull distribution, reproductive status a Bernoulli distribution, and both growth response variables were assumed to follow a lognormal distribution. We used Bayesian inference, and all models were fitted in R 4.1.2 (R Core Team 2021) with brms 2.16.3 (Bürkner 2017) and Stan 2.21.0 (Stan Development Team 2021). Model inferences were based on four chains of 20,000 iterations, discarding the first 10,000 iterations and saving every 10th iteration, resulting in a total of 4000 posterior draws. Models were assessed for appropriateness with posterior predictive checks and model fit was estimated with Bayesian R2 values (Gelman et al. 2018).
Results
Posterior predictive checks indicated that all four models were appropriately specified (Figures S1–S4 Online Resource 2). Based on Bayesian R2 values, these models explained 50% of the variation in survival outcomes (days until mortality), 45% of the variation in reproductive status, 74% of the variation in height growth, and 67% of the variation in crown growth.
The use of fences to protect translocated plants from herbivores resulted in a positive survival effect across all species and was the only response variable to have a consistently positive impact across all species. It also had one of the largest effect sizes on survival of all the predictor variables, although credible intervals were quite variable among species (Fig. 2). By contrast, fencing had, on average, negligible impact on height growth and reproduction and even a slightly negative impact on crown growth, although credible intervals were variable among species, with some quite wide (e.g. Gastrolobium lutefolium) and some quite narrow (e.g. Lambertia orbifolia subsp. orbifolia) (Fig. 2). Although individual species responded positively for the growth or reproduction response variables, these were generally not the same species across these three response variables.

Effects of fencing on plant translocation outcomes. Estimated values are the link-scale effect of fencing on survival, reproduction, and growth of each species. Positive values indicate that fencing increased a given response variable. Points are median estimated effects, thick lines bound 80% credible intervals, and thin lines bound 95% credible intervals. The dashed black line is at zero (no effect), and the dashed grey line is the mean estimated effect over all species. Colours denote the most- common rainfall zone in which each species occurs (values in mm per year)
Watering over the first two dry seasons had no effect on survival (averaged across species), although survival of several individual species was positively influenced by watering (Fig. 3). Across all species, watering had a negligible effect on height and crown growth. The largest effect of watering was on the probability of reproduction, with all species except one (Synaphea quartzitica) showing a positive response to watering. Credible intervals for each of the four response variables showed notable variation among species.

Effects of watering on plant translocation outcomes. Estimated values are the link-scale effect of watering on survival, reproduction, and growth of each species. Positive values indicate that watering following planting increased a given response variable. Points are median estimated effects, thick lines bound 80% credible intervals, and thin lines bound 95% credible intervals. The dashed black line is at zero (no effect), and the dashed grey line is the mean estimated effect over all species. Colours denote the most common rainfall zone in which each species occurs (values in mm per year)
Rainfall deviation in the 2 years post-planting was associated with a positive effect across all species for all four response variables (i.e. above-average rainfall was associated with increased survival, growth, and likelihood of reproduction) (Fig. 4). It was the only predictor to have a positive effect for all response variables. However, for the survival response variable thirty species had a positive response to rainfall deviation but a negative or neutral response to watering (Figs. 3 and 4). Conversely, 11 species had a negative survival response to rainfall deviation but a positive survival response to watering. Confounding of watering and rainfall deviation outcomes is considered unlikely as watering was applied independent of rainfall predictions or rainfall zones. Only two species (Grevillea maccutcheonii and Grevillea humifusa) responded positively for the survival response to both watering and rainfall deviation. The amount of rainfall in the 30 days prior to planting across all species had a negligible effect on survival, growth and reproduction, although some species had a small positive response to this variable (Fig. 5).

Effects of the deviation from average rainfall in two years following planting on plant translocation outcomes. Estimated values are the link-scale effect of rainfall deviation on survival, reproduction, and growth of each species. Positive values indicate that higher-than-average rainfall in the two years following planting increased a given response variable. Points are median estimated effects, thick lines bound 80% credible intervals, and thin lines bound 95% credible intervals. The dashed black line is at zero (no effect), and the dashed grey line is the mean estimated effect over all species. Colours denote the most common rainfall zone in which each species occurs (values in mm per year)

Effects of rainfall in the 30 days prior to planting on plant translocation outcomes. Estimated values are the link-scale effect of prior rainfall on survival, reproduction, and growth of each species. Positive values indicate that larger amounts of rainfall prior to planting increased a given response variable. Points are median estimated effects, thick lines bound 80% credible intervals, and thin lines bound 95% credible intervals. The dashed black line is at zero (no effect), and the dashed grey line is the mean estimated effect over all species. Colours denote the most common rainfall zone in which each species occurs (values in mm per year)
Discussion
As the establishment of new populations of threatened plants through translocation becomes an increasingly important option to prevent extinction, the factors that influence the likelihood of establishing resilient and viable populations have received increasing attention (Guerrant 2012, 2013; Dalrymple et al. 2011, 2012; Godefroid et al. 2011; Silcock et al. 2019). Our analysis of 50 translocations of threatened species established across the SWAFR showed that, in general, excluding herbivores using fences increased survival, watering plants over the first two summer dry periods following planting increased the probability of reproduction, and an increase in rainfall from the long-term average in the two years after planting increased survival, growth, and probability of reproduction. These results highlight the importance in translocation establishment of herbivory and the need to account for seasonal water availability in Mediterranean ecosystems and, more broadly, in water limited environments.
Herbivores can reduce the establishment and fitness of translocated populations by impacting plant survival, growth, and reproduction (Albrecht and Long 2019) and, as such, excluding herbivores is one of the most common post-planting management techniques used in translocations of threatened plants (Guerrant 2012). In this study, fencing to protect plants from vertebrate herbivores consistently, strongly, increased plant survival across all the species, consistent with many other studies of threatened plant translocations (Maschinski et al. 2004; Jusaitis 2005; Fenu et al. 2016; Albrecht and Long 2019). Fences physically prevent vertebrate grazers and browsers from accessing the plants and, unless the fences are breached, this type of management will have a long-lasting benefit to plant survival. However, in our study, herbivore exclusion did not benefit plant growth or reproduction, which contrasts with other studies where herbivory reduced growth (Jusaitis 2005; Buisson et al. 2015) and reproduction (Albrecht and Long 2019; Fenu et al. 2016). The translocated plants that survived herbivory outside the fences may have been protected in some way by other plants. They may grow under or through these other plants, so they are not seen, or were protected by the spines, prickles, or toxin defences of these other plants (Jusaitis 2005; Garcia and Obeso 2003). Plants that survive outside the fences in these safe havens are then able to grow and reproduce to the same extent as plants protected by the fences. For example, Acacia cochlocarpa subsp. cochlocarpa plants growing in unfenced and open sites were heavily grazed, compared to those unfenced plants that were growing under large shrubs, which remained ungrazed (L. Monks pers obs.). There was also some variation around the effect sizes for many of the species for survival, growth and reproduction. While species with large sample sizes, such as Lambertia orbifolia subsp. orbifolia, did have less variation around the effect sizes, variability in herbivory, both spatially and temporally, is well documented (Huntly 1991).
Watering of translocated plants is another commonly applied management action, particularly in arid and Mediterranean climates (Guerrant 2012; Fenu et al. 2019). Many studies show the beneficial effects of irrigation on seedling survival (Castro et al. 2005; Siles et al. 2010; Dollard and Carrington 2013; Lopez-Jurado et al. 2019). However, in our study, we did not see an overall positive effect from watering on survival and a mostly neutral to very weakly positive effect on growth across the range of habitats and rainfall zones. This suggests either that watering does not improve survival or growth, that watering was not applied at a level, or a frequency sufficient to minimise deaths or maximise growth (Siles et al. 2010), or that the watering was insufficient during events like periods of extreme heat or drought. Interestingly, despite no overall survival response to watering, just under a third of the species showed a small positive effect from supplementary watering. As these species were distributed across the different rainfall zones, it may have been temporal rainfall patterns (i.e. short-term dry periods) rather than overall annual rainfall that played a role in this positive response. It’s possible, that if watering amount and/or frequency is increased, the beneficial effect on seedling survival seen in other studies (e.g. Dollard and Carrington 2013; Lopez-Jurado et al. 2019) would occur in our species. Watering could also be combined with other treatments such as mulch (Devine et al. 2007) or nurse plants (Castro et al. 2002) to enhance its effectiveness.
The greatest benefit from supplementary watering was the probability that plants were reproductive, with all except one species showing a positive effect from watering. This is an interesting result as the likelihood of reproduction was increased long after the watering had ceased, suggesting that watering did have a beneficial effect on plant establishment, perhaps by improving root systems (Castro et al. 2005), despite this not being reflected in increased survival. Increased probability of reproduction may mean an increased probability of recruitment of second-generation plants. Watering did indeed result in improved chances of recruitment in the Australia-wide review of translocation outcomes (Whitehead et al. 2023). However, in our case, recruitment was rarely seen, likely due to the longevity of the species and time lags to recruitment, which are common in many translocated species (Albrecht et al. 2019), and lack of a recruitment trigger, such as fire (Yates and Broadhurst 2002). Nevertheless, watering increased the probability of reproduction, which should lead to larger seed bank availability when recruitment conditions are suitable and, therefore, an increased likelihood of establishing a self-sustaining population.
Where rainfall is limited, such as in arid areas, or rainfall is seasonal, such as in Mediterranean climates, the amount of rainfall during and after planting can have a large impact on translocation survival, growth, reproduction, and second-generation seedling recruitment (Maschinski et al. 2004; Guerrant 2012). In our study, rainfall in the 30 days prior to planting had a negligible impact on translocation survival, growth, and reproduction. This may indicate that soil moisture prior to planting plays a limited role in translocation establishment, or if conditions are drier than average, lack of soil moisture prior to planting can be overridden with watering during planting. This is useful knowledge when financial or logistical factors rather than just seasonal considerations influence the timing of planting. In contrast, increased rainfall in the two years following planting was the only predictor variable to positively influence all four response variables. This result is not surprising given natural recruitment and establishment of many species relies on infrequent rainfall events or particularly wet years (Jusaitis 2005; Castro et al. 2005; Wendelberger and Maschinski 2016), and reproduction can be enhanced by increased rainfall (Duncan and Moloney 2018; Janissen et al. 2021). However, this clear result is not particularly helpful to managers when translocation planning and preparation generally takes place well before accurate rainfall predictions can be made. Although there may be some benefit to using seasonal weather forecasting to plan planting around adverse weather events in the short term (< 4 months) (Hagger et al. 2018), once propagation has begun, planting cannot usually be delayed significantly to take into account longer-term weather predictions, so strategies that bet hedge against poor climatic conditions, such as watering or planting across multiple years and sites are the best option (Siles et al. 2010; Menges et al. 2016). Additionally, reduced rainfall, increased temperatures, and changes to rainfall seasonality are predicted for southwest Australia, with an increase in climate variability and extreme events predicted globally under many climate change scenarios (Indian Ocean Climate Initiative 2012; IPCC 2022). As such, the use of irrigation is likely to become more important in translocation establishment, especially if the volume and frequency can be resolved to a point where it consistently improves survival and growth in addition to reproduction across all species.
There was no consistent pattern to the rainfall zones of species that responded positively to watering or rainfall deviation following planting. Species adapted to and planted in wetter rainfall zones did not respond to rainfall deviation or watering differently to species adapted to and planted in drier rainfall zones. There was a group of 30 species that had a positive survival response to rainfall deviation but a negative or neutral survival response to watering and another group of 11 species that responded negatively to rainfall deviation but positively to watering, with both groups containing species from across the rainfall zones and across taxonomic groups. Local site conditions, habitat quality, and influence of neighbouring plants play an important role in plant establishment and population viability (Albrecht and Long 2019; Wendelberger and Maschinski 2016; Menges et al. 2016; Buisson et al. 2015) and more detailed studies at the site level may provide some insight into effects of these factors and their interactions. For example, inter-site variation in annual rainfall may have contributed to species tolerating a wide range in rainfall amounts for establishment. Indeed, rainfall reliability, rather than amounts, is suggested to be important for seedling establishment in jarrah forest within SWAFR (Standish et al. 2015). Many species may require average or above-average rainfall over one or more seasons for successful seedling establishment (Maschinski et al. 2004; Yates et al. 2011), and our findings suggest that watering is not needed to improve survival rates in the periods of above-average rainfall. Nevertheless, the long lead-in time to translocation planting does mean that the decision to irrigate, and allocation of budgets for watering is often made well in advance of knowledge of projected seasonal rainfall. Therefore, combined with the improved likelihood of reproduction with watering, our findings suggest this management action should be standard procedure, particularly given projections for a drying climate.
The value of using several measures to determine translocation success was highlighted by our study, as we found that at the broad level, across all species, there were different responses to the different management techniques or environmental conditions investigated. Survival was improved by fencing, the probability of reproduction was enhanced by watering, and survival, growth, and reproduction were increased by rainfall deviation. Survival is often the only measure of success (Godefroid et al. 2011; Guerrant 2012) and, if this was the case, our study would not have identified the value of watering in improving reproduction. In addition, as most of the species in our study were long-lived woody perennials, the long timeframe over which monitoring occurred (in some cases up to 20 years) was necessary to understand some aspects of success such as reproduction, which took several years to begin for many species. However, this timeframe was still too short to understand other aspects such as recruitment of a second generation, as many of these species have episodic recruitment linked primarily to fire, which occurred infrequently in our translocated populations.
Long-term monitoring of translocations and analysis of the factors that contribute to successful establishment of self-sustaining populations is vital to ensure we learn from our efforts (Menges 2008; Albrecht and Long 2019). In our study, the best management strategy to maximise survival was to protect translocated plants from herbivory by fencing, which aligns with results from several studies in other regions (Maschinski et al. 2004; Jusaitis 2005; Buisson et al. 2015; Fenu et al. 2016; Albrecht and Long 2019). Watering increased the likelihood of reproduction, and planting in years of above-average rainfall was beneficial to survival, growth, and reproduction. Given that rainfall amount is difficult to predict when translocation planning begins, consideration should be given to selecting translocation sites at the wetter end of species’ climatic zones to improve success. As climate change models predict increased aridity and increased frequency, intensity, longevity and severity of drought in many areas globally (Cook et al. 2022; IPCC 2022), the results of this study could well apply across other regions with Mediterranean-type climates and drylands where water is already a limiting factor.
Data Availability
Data sets from this study are available from corresponding author on reasonable request. Model code to recreate all analyses is available at https://github.com/jdyen/translocations.
References
Abeli T, D’Agostino M, Orsenigo S et al (2021) IDPlanT: the Italian database of plant translocation. Plant Biosyst 155(6):1174–1177. https://doi.org/10.1080/11263504.2021.1985004
Albrecht MA, Long QG (2019) Habitat suitability and herbivores determine reintroduction success of an endangered legume. Plant Divers 41:109–117. https://doi.org/10.1016/j.pld.2018.09.004
Albrecht MA, Osazuwa-Peters OL, Maschinski J, Bell TJ, Bowles ML, Brumback WE, Duquesnel J, Kunz M, Lange J, McCue KA, McEachern AK, Murray S, Olwell P, Pavlovic NB, Peterson CL, Possley J, Randall JL, Wright SJ (2019) Effects of life history and reproduction on recruitment time lags in reintroductions of rare plants. Conserv Biol 33:601–611. https://doi.org/10.1111/cobi.13255
BoM (2019) Australian Government Bureau of Meteorology Climate data online. http://www.bom.gov.au/climate/data/. Accessed 2019
Buisson E, Corcket E, Dutoit T (2015) Limiting processes for perennial plant reintroduction to restore dry grasslands. Restor Ecol 23:947–954. https://doi.org/10.1111/rec.12255
Burbidge AA, Byrnes M, Coates D, Garnett ST, Harris S, Hayward MW, Martin T, McDonald-Madden E, Mitchell N, Nally S, Setterfield SA (2011) Is Australia ready for assisted colonization? Policy changes required to facilitate translocations under climate change. Pac Conserv Biol 17:259–269. https://doi.org/10.1071/PC110259
Bürkner P (2017) brms: an R package for Bayesian multilevel models using Stan. J Stat Softw 80:1–28. https://doi.org/10.18637/jss.v080.i01
Castro J, Zamora R, Hódar J, Gómez JM (2002) Use of shrubs as nurse plants: a new technique for reforestation in Mediterranean mountains. Restor Ecol 10:297–305. https://doi.org/10.1046/j.1526-100X.2002.01022.x
Castro J, Zamora R, Hódar J, Gómez JM (2005) Alleviation of summer drought boosts establishment of Pinus sylvestris in a Mediterranean mountain: an experimental approach. Plant Ecol 181:191–202. https://doi.org/10.1007/s11258-005-6626-5
Cook BI, Smerdon JE, Cook ER et al (2022) Megadroughts in the common era and the anthropocene. Nat Rev Earth Environ. https://doi.org/10.1038/s43017-022-00329-1
Dalrymple SE, Stewart GB, Pullin AS (2011) Are re-introductions an effective way of mitigating against plant extinctions? CEE review 07–008 (SR32). Collab Environ Evid 008:1–63
Dalrymple S, Banks E, Stewart GB, Pullin AS (2012) A meta-analysis of threatened plant reintroductions from across the globe. In: Maschinski J, Haskins K (eds) Plant reintroduction in a changing climate: promises and perils. Island Press, Washington D.C., pp 31–50
Devine WD, Harrington CA, Lathrop LP (2007) Post-planting treatments increase growth of Oregon white oak (Quercus garryana Dougl. Ex Hook.) seedlings. Restor Ecol 15:212–222. https://doi.org/10.1111/j.1526-100X.2007.00205.x
Dillon RA, Monks LT, Coates DJ (2018) Establishment success and persistence of threatened plant translocations: an experimental approach. Aust J Bot 66:338–346. https://doi.org/10.1071/BT17187
Dollard JJ, Carrington ME (2013) Experimental reintroductions of Beach Pea (Lathyrus japonicaus) to the Indiana Dunes National Lakeshore. Ecol Restor 31:368–377. https://doi.org/10.3368/er.31.4.368
Drayton B, Primack RB (2012) Success rates for reintroductions of eight perennial plant species after 15 years. Restor Ecol 20:299–303. https://doi.org/10.1111/j.1526-100X.2011.00860.x
Duncan M, Moloney PD (2018) Comparing wild and reintroduced populations of the threatened orchid Diuris fragrantissima (Orchidaceae) in south-eastern Australia. Aust J Bot 66:459–467. https://doi.org/10.1071/BT18047
Fenu G, Cogoni D, Bacchetta G (2016) The role of fencing in the success of threatened plant species translocation. Plant Ecol 217:207–217. https://doi.org/10.1007/s11258-015-0517-1
Fenu G, Bacchetta G, Charalambos SC, Fournaraki C, del Galdo GPG, Gotsiou P, Kyratzis A, Piazza C, Vicens M, Pinna MS, de Montmollin B (2019) An early evaluation of translocation actions for endangered plant species on Mediterranean islands. Plant Divers 41:94–104. https://doi.org/10.1016/j.pld.2019.03.001
Garcia D, Obeso JR (2003) Facilitation by herbivore-mediated nurse plants in a threatened tree, Taxus baccata: local effects and landscape level consistency. Ecography 26:739–750. https://doi.org/10.1111/j.0906-7590.2003.03601.x
Gelman A, Goodrich B, Gabry J, Vehtari A (2018) R-squared for Bayesian regression models. Am Statistician 73:307–309. https://doi.org/10.1080/00031305.2018.1549100
Gioia P, Hopper SD (2017) A new phytogeographic map for the Southwest Australian Floristic Region after an exceptional decade of collection and discovery. Bot J Linn Soc 184:1–15. https://doi.org/10.1093/botlinnean/box010
Godefroid S, Piazza C, Rossi G, Buord S, Stevens A, Aquraiuja R, Cowell C, Weekley CW, Vogg G, Iriondo JM, Johnson I, Dixon B, Gordon D, Magnanon S, Valentin B, Bjureke K, Kroopman R, Vicens M, Virevaire M, Vanderborght T (2011) How successful are plant species reintroductions? Biol Conserv 144:672–682. https://doi.org/10.1016/j.biocon.2010.10.003
Gosper CR, Coates DJ, Hopper SD, Byrne M, Yates CJ (2020) The role of landscape history in the distribution and conservation of threatened flora in the Southwest Australian Floristic Region. Bot J Linn Soc 133:394–410. https://doi.org/10.1093/biolinnean/blaa141
Gosper CR, Percy-Bower JM, Byrne M, Llorens TM, Yates CJ (2022) Distribution, biogeography and characteristics of the threatened and data-deficient flora in the Southwest Australian Floristic Region. Diversity 14:493. https://doi.org/10.3390/d14060493
Guerrant EO (1996) Designing populations: demographic, genetic, and horticultural dimensions. In: Falk DA, Millar CI, Olwell M (eds) Restoring diversity. Strategies for reintroduction of endangered plants. Island Press, Covelo, pp 127–155
Guerrant EO (2012) Characterizing two decades of rare plant reintroductions. In: Maschinski J, Haskins K (eds) Plant reintroduction in a changing climate: promises and perils. Island Press, Washington D.C., pp 9–29
Guerrant EO (2013) The value and propriety of reintroduction as a conservation tool for rare plants. Botany 91:v–x. https://doi.org/10.1139/cjb-2012-0239
Guerrant EO, Kaye TN (2007) Reintroduction of rare and endangered plants: common factors, questions and approaches. Aust J Bot 55:362–370. https://doi.org/10.1071/BT06033
Hagger V, Dwyer J, Shoo L, Wilson K (2018) Use of seasonal forecasting to manage weather risk in ecological restoration. Ecol Appl 28:1797–1807. https://doi.org/10.1002/eap.1769
Hoegh-Guldberg O, Hughes L, McIntyre S, Lindenmayer DB, Parmesan C, Possingham HP, Thomas CD (2008) Assisted colonization and rapid climate change. Science 321:345–346. https://doi.org/10.1126/science.1157897
Hopper SD, Gioia P (2004) The southwest Australian floristic region: evolution and conservation of a global hot spot of biodiversity. Annu Rev Ecol Evol S 35:623–650. https://doi.org/10.1146/annurev.ecolsys.35.112202.130201
Huntly N (1991) Herbivores and the dynamics of communities and ecosystems. Annu Rev Ecol Evol S 22:477–503. https://doi.org/10.1146/annurev.es.22.110191.002401
Indian Ocean Climate Initiative (2012) Western Australia's weather and climate: a synthesis of indian ocean climate initiative stage 3 research. In: Bates B, Frederiksen C, Wormworth J (eds) CSIRO and BoM, Australia
IPCC (2022) Summary for Policymakers. Pörtner H-O, et al. (eds). In: Climate Change 2022: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change Pörtner H-O, et al. (eds). Cambridge University Press, Cambridge, UK and New York, NY, USA, pp 3–33. https://doi.org/10.1017/9781009325844.001.
IUCN (2022) The IUCN Red List of Threatened Species Version 2022–1. https://www.iucnredlist.org. Accessed 9 Sept 2022
IUCN/SSC (2013) Guidelines for Reintroductions and Other Conservation Translocations Version 10 Gland, Switzerland: IUCN Species Survival Commission
Janissen B, French G, Selby-Pham J, Lawrie AC, Huynh T (2021) Differences in emergence and flowering in wild, re-introduced and translocated populations of an endangered terrestrial orchid and the influences of climate and orchid mycorrhizal abundance. Aust J Bot 69:9–20. https://doi.org/10.1071/BT20102
Jusaitis M (2005) Translocation trials confirm specific factors affecting the establishment of three endangered plant species. Ecol Manag Restor 6:61–67. https://doi.org/10.1111/j.1442-8903.2005.00220.x
Kreyling J, Bittner T, Jaeschke A, Jentsch A, Steinbauer MJ, Thiel D, Beierkuhnlein C (2011) Assisted colonization: a question of focal units and recipient localities. Restor Ecol 19:433–440. https://doi.org/10.1111/j.1526-100X.2011.00777.x
Liu H, Ren H, Liu Q, Wen XY, Maunder M, Gao JY (2015) Translocation of threatened plants as a conservation measure in China. Conserv Biol 29:1537–1551. https://doi.org/10.1111/cobi.12585
López-Jurado J, Mateos-Naranjo E, García-Castaño JL, Balao F (2019) Conditions for translocation of a key threatened species, Dianthus inoxianus Gallego, in the southwestern Iberian Mediterranean forest. Forest Ecol Manag 446:1–9. https://doi.org/10.1016/j.foreco.2019.05.008
Maschinski J, Duquesnel J (2006) Successful reintroductions of the endangered long-lived Sargent’s cherry palm, Pseudophoenix sargentii, in the Florida Keys. Biol Conserv 134:122–129. https://doi.org/10.1016/j.biocon.2006.07.012
Maschinski J, Baggs JE, Sacchi CF (2004) Seedling recruitment and survival of an endangered limestone endemic in its natural habitat and experimental reintroduction sites. Am J Bot 91:689–698. https://doi.org/10.3732/ajb.91.5.689
Menges ES (2000) Population viability analyses in plants: challenges and opportunities. Trends Ecol Evol 15:51–56. https://doi.org/10.1016/S0169-5347(99)01763-2
Menges ES (2008) Turner Review No 16 Restoration demography and genetics of plants: when is a translocation successful? Aust J Bot 56:187–189. https://doi.org/10.1071/BT07173
Menges ES, Smith SA, Weekley CW (2016) Adaptive introductions: how multiple experiments and comparisons to wild populations provide insights into requirements for long-term introduction success of an endangered shrub. Plant Divers 38:238–246. https://doi.org/10.1016/j.pld.2016.09.004
Mittermeier RA, Turner WR, Larsen FW, Brooks TM, Gascon C (2011) Global biodiversity conservation: the critical role of hotspots. In: Zachos E, Habel JC (eds) Biodiversity hotspots: distribution and protection of conservation priority areas. Springer, Berlin, pp 3–22
Mittermeier RA, Gil PR, Hoffmann M, Pilgrim J, Brooks T, Mittermeier CG, Lamoreux J, da Fonseca GAB (2004) Hotspots revisited. Earth’s biologically richest and most endangered terrestrial ecoregions. Cemex, Mexico City, pp 29–33
Monks L, Coates D (2002) The translocation of two critically endangered Acacia species. Conserv Sci West Aust 4:54–61
Monks LT, Coates DJ, Bell T, Bowles M (2012) Determining success criteria for reintroductions of threatened long-lived plants. In: Maschinski J, Haskins KE (eds) Plant reintroduction in a changing climate: promises and perils. Island Press, Washington D.C., pp 189–208
Monks L, Barrett S, Beecham B, Byrne M, Chant A, Coates D, Cochrane JA, Crawford A, Dillon R, Yates C (2019) Recovery of threatened plant species and their habitats in the biodiversity hotspot of the Southwest Australian Floristic Region. Plant Divers 41:59–74. https://doi.org/10.1016/jpld201809006
Myers N, Mittermeier RA, Mittermeier CG, da Fonseca GAB, Kent J (2000) Biodiversity hotspots for conservation priorities. Nature 403:853–858. https://doi.org/10.1038/35002501
Pavlik BM (1996) Defining and measuring success. In: Falk DA, Millar CI, Olwell M (eds) Restoring diversity. Strategies for reintroduction of endangered plants. Island Press, Covelo, pp 127–155
R Core Team (2021) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Rathbone DA, Barrett S (2017) Vertebrate browsing impacts in a threatened montane plant community and implications for management. Ecol Manag Restor 18:164–171. https://doi.org/10.1111/emr.12259
Reiter N, Whitfield J, Pollard G, Bedggood W, Argall M, Dixon K, Davis B, Swarts N (2016) Orchid re-introductions: an evaluation of success and ecological considerations using key comparative studies from Australia. Plant Ecol 217:81–95. https://doi.org/10.1007/s11258-015-0561-x
Seddon PJ, Armstrong DP, Soorae P, Launay F, Walker S, Ruiz-Miranda CR, Molur S, Koldewey H, Kleiman DG (2009) The risks of assisted colonization. Conserv Biol 23:788–789. https://doi.org/10.1111/j.1526-100X.2010.00724.x
Silcock JL, Simmons CL, Monks L, Dillon R, Reiter N, Jusaitis M, Vesk PA, Byrne M, Coates DJ (2019) Threatened plant translocation in Australia: a review. Biol Conserv 236:211–222. https://doi.org/10.1016/j.biocon.2019.05.002
Siles G, Rey PJ, Alcántara JM, Bastida JM, Herreros JL (2010) Effects of soil enrichment, watering and seedling age on establishment of Mediterranean woody species. Acta Oecol 36:357–364. https://doi.org/10.1016/j.actao.2010.03.002
Stan Development Team (2021) Stan modeling language users guide and reference manual, 2.21.0. https://mc-stan.org
Standish RJ, Daws MI, Gove AD, Didham RK, Grigg AH, Koch JM, Hobbs RJ (2015) Long-term data suggest jarrah-forest establishment at restored mine sites is resistant to climate variability. J Ecol 103:78–89. https://doi.org/10.1111/1365-2745.12301
Wendelberger KS, Maschinski J (2016) Assessing microsite and regeneration niche preferences through experimental reintroduction of the rare plant Tephrosia angustissima var corallicola. Plant Ecol 217:155–167. https://doi.org/10.1007/s11258-015-0521-5
Whitehead M, Silcock JL, Simmons CL, Monks L, Dillon R, Reiter N, Jusaitis M, Coates DJ, Byrne M, Vesk PA (2023) Effects of common management practices on threatened plant translocations. Biol Conserv. Accepted 13 March 2023.
Yates CJ, Broadhurst LM (2002) Assessing limitations on population growth in two critically endangered Acacia taxa. Biol Conserv 108:13–26. https://doi.org/10.1016/S0006-3207(02)00084-8
Yates CJ, Gibson N, Pettit NE, Dillon R, Palmer R (2011) The ecological relationships and demography of restricted ironstone endemic plant species: implications for conservation. Aust J Bot 59:692–700. https://doi.org/10.1071/BT11199
Acknowledgements
We would like to thank the staff and volunteers of the Department of Biodiversity, Conservation, and Attractions who have assisted with planning, planting, and monitoring of the translocations included in this study. Specifically, we would like to highlight and acknowledge the assistance of Alanna Chant, Amanda Shade, Andrew Crawford, Anne Cochrane, Bree Phillips, and Sarah Barrett who have inspired, supported, encouraged, and assisted with this work over the past two decades.
Funding
Funding for this study was received from the Australian government’s National Environmental Science Program Threatened Species Recovery Hub.
Author information
Authors and Affiliations
Contributions
LM, DC, and RD contributed to planning, planting, and monitoring all the translocations; all authors contributed to development of research; LM, RD, DC, and RS contributed to data collection; LM and RD contributed to data collation; JY, PV, and LM contributed to model development; LM wrote the manuscript; and all authors edited the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Communicated by Thomas Abeli.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Monks, L., Yen, J., Dillon, R. et al. Herbivore exclusion and water availability improve success across 76 translocations of 50 threatened plant species in a biodiversity hotspot with a Mediterranean climate. Plant Ecol 224, 817–830 (2023). https://doi.org/10.1007/s11258-023-01313-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-023-01313-5