
Abstract
Herbaceous understory vegetation is an important part of temperate forested ecosystems, the diversity and composition of which are strongly dependent on the conditions of the forest stand and the landscape. The aim of this study was to find the most important environmental drivers influencing understory herb layer species composition (explored with multivariate analysis), and richness and cover (analysed by linear modelling) in managed mixed forests in West Hungary. Our detailed inventory showed that the most important factors increasing the diversity and cover of the understory are light, tree species richness, and landscape diversity. Composition is also mainly influenced by light conditions and tree species richness, with minor effects of tree species composition, soil texture, and moss cover. As the strongest influencing factors are closely linked to stand structure and tree species composition, they can either directly or indirectly be altered by forest management. In the studied region, heterogeneous light conditions and canopy structure, the maintenance of tree species richness and forest continuity are key elements for the conservation of forest herbs. Forestry that maintains continuous forest cover and the tree selection management system can better provide these conditions than the presently widely used shelterwood management system.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Forest herbs make an important contribution to the biodiversity (e.g. Gilliam 2007; Hart and Chen 2006; Wayman and North 2007), ecosystem functioning (Allen et al. 2002; Nilsson and Wardle 2005), nutrient cycling (Muller 2014), and even biomass (Gonzalez et al. 2013) and seedling community (George and Bazzaz 2014) of temperate forests. The factors influencing their composition and diversity are therefore widely studied (e.g. Hutchinson et al. 1999; Vockenhuber et al. 2011). Although the results of these studies are remarkably heterogeneous, stand structure, light conditions, edaphic factors, site history and landscape characteristics are generally assumed to be of importance.
In managed stands, the features most directly affected by human activity are stand structure and tree species composition. These, in turn, affect understory light conditions (Lochhead and Comeau 2012; Tinya et al. 2009a) and edaphic conditions such as soil water content, pH, temperature and nutrient availability (Barbier et al. 2008; Bartels and Chen 2013; Heithecker and Halpern 2006). Although obviously not the object of harvesting practices, understory vegetation is also directly affected by forest management (Duguid and Ashton 2013).
In Hungary and across much of Europe, after the abandonment of traditional management practices in the twentieth century (Hédl et al. 2010), the shelterwood forestry system has been the dominant forest management method until recently. This management type resulted in even-aged stands with homogenous structure on a relatively coarse scale (5–10 ha; Matthews 1991; Savill 2004). In the last decades, efforts have been made to change the management structure in favour of more natural practices, such as group and stem selection systems (Rosenvald and Lohmus 2008; Bauhus et al. 2013). Sustainable management practices aim at mimicking the natural disturbance regime as much as possible, thus maintaining the natural processes of a forest (Bauhus et al. 2013). Professional debate on possible ways to achieve this aim is intense. In some cases, even small scale clear-cutting is argued to be an efficient way of forming and harvesting forests (Heinrichs and Schmidt 2009); however, some species indicate the effects of clear-cuts as far as 150 m inside the remaining forest (Godefroid et al. 2005). Intensive management types cause diverse changes in the species richness of ecologically distinct species groups (on the basis of ecological traits, Kenderes and Standovár 2003), similarly to changes in plant traits during the succession of a forest (Catorci et al. 2012).
Most of the forests in West Hungary have been used in ways fars from their natural disturbance regime: clear-cut for wood or to gain space for pastures and arable areas (Tímár et al. 2002). These actions have long-term effects and leave their mark both on understory and overstory (e.g. Hermy and Verheyen 2007; Wulf 2003). Ancient (chronologically continuous forest cover since first written record) and recent (area utilised differently sometime throughout recorded history) woodlands host markedly different herbaceous vegetation (e.g. Hermy and Verheyen 2007; Kelemen et al. 2014). The exact type and intensity of previous land utilisation of the area also has important effects (Verheyen et al. 2003), which may be detectable for millennia (Dupouey et al. 2002). The inclusion of this feature into analyses can be achieved via old maps, army surveys, and common knowledge of the area. In some cases, previous land use seems to have an even more important influence on the current condition of the forest floor than present stand structure or microtopography (Ito et al. 2004). According to a theory, management may act through past minimum and maximum canopy covers, which work as bottlenecks (Økland et al. 2003). Present management obviously holds the key to the characteristics of a forest; in many cases, the typical aspects of which can be assessed on the basis of the ownership of the stand (state-owned vs. private forests, Berges et al. 2013; Schaich and Plieninger 2013; Zmihorski et al. 2010). In our study area, in private forests, management techniques are much closer to the natural disturbance regime than in state-owned stands.
Edaphic factors, which are also strongly influenced by stand characteristics (von Arx et al. 2012), naturally have strong effects on ground vegetation. The surrounding landscape, serving as the species pool for the studied habitat, is also an important influential factor, and can, in some cases, serve as the basis for assessing the biodiversity of poorly explored, high diversity areas (Priego-Santander et al. 2013).
However, the diversity of understory vegetation in itself may be misleading and is not equal to forest naturalness. In disturbed or otherwise altered stands, non-forest species could considerably increase these values (Hermy and Verheyen 2007; Mikoláš et al. 2014; Paillet et al. 2010). Therefore, it is reasonable to handle closed-forest herbs separately from the total species pool (Kelemen et al. 2014). The aim of this study is to produce a detailed analysis of the understory herb layer of the studied forests, which can be used effectively in conservation and management. Our questions were (1) which environmental background factors influence the composition of the understory herb layer (multivariate analysis); (2) which are the most important factors that enhance or hinder species richness and abundance of the understory (linear modelling); (3) is there a marked difference between the environmental needs of closed-forest and non-forest species; and (iv) from the most influential background factors, which could be altered favourably by forest management.
Materials and methods
Study area
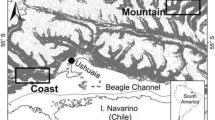
Our study was carried out on 34 plots in the Őrség region, West Hungary (N46°51–55′ and W16°07–23′; Fig. 1). Elevation of the study area is between 250 and 350 m above sea level. The bedrock in the area is alluviated gravel mixed with loess, and its topography consists of hills and wide valleys. On hills, mostly pseudogleyic brown forest soil can be found (planosols or luvisols), while in the valleys mire and meadow soils (gleysols), they are common (Stefanovits et al. 1999). The soil is acidic (pH 4.0–4.8, measured in water, in the upper 10 cm) and nutrient poor. Average annual mean temperature is 9.0–9.5 °C, with a mean annual precipitation of 700–800 mm (Dövényi 2010). In the area, we can find forests of various species composition and stand structure on similar forest sites, namely on similar climatic, topographical, and bedrock conditions. The study area is dominated by beech (Fagus sylvatica L.), sessile and pedunculate oak (Quercus petraea L. and Q. robur L.), hornbeam (Carpinus betulus L.), Scots pine (Pinus sylvestris L.), and Norway spruce (Picea abies (L.) Karst.), forming monodominant and mixed stands as well. The proportion of various, subordinant tree species (birch—Betula pendula Roth., aspen—Populus tremula L., chestnut—Castanea sativa Mill., wild cherry—Prunus avium L., etc.) is relatively high (Tímár et al. 2002). The present diverse state of the forests is partly due to the special landscape history of the area. After the twelfth century, extensive farming and other activities, such as litter collection and ridging (a special form of tillage), led to the deforestation and acidification of the area and the erosion of soil. From the nineteenth century, extensive farming was repressed, and a reforestation of the area took place, mainly by Scots pine and pioneer tree species linked to it (birch, aspen). Later, selective cutting has been applied by farmers, alongside the industrial shelterwood system in state-owned forests (Gyöngyössy 2008; Tímár et al. 2002). The conditions which developed owing to these events were favourable to tree, herb, and moss species that prefer nutrient poor and disturbed conditions. Current existing forests are a fine-scale mixture of ancient and recent stands. The mixture of these pioneer species and typical species of the mesic forests together create a remarkably rich and various species composition in the region.

Study area: Őrség region, West Hungary (N46°51–55′ and W16°07–23′); the dots show our plots
Data collection
We collected our data in 34 forest stands (Fig. 1). These were selected by stratified random sampling from the forest stand database of the Hungarian National Forest Service. All of these stands were located on relatively level ground, devoid of direct water influence, and the age of dominant tree layer was between 70 and 100 years. The stratification criterion was tree species composition: these stands represent different combinations of the main tree species (sessile and pedunculate oak, beech and Scots pine) of the area. Within the categories—based upon tree species composition—random selection was carried out.
In all the stands, one 40 m × 40 m plot was selected, where all tree individuals (including snags) above 5 cm diameter at breast height (DBH) were mapped. Species identity, DBH, and height of these trees were also recorded. The proportion of tree species (beech, hornbeam, oak, Scots pine, Norway spruce and subordinate trees) was expressed, based on volume. The volume of the trees was calculated by species specific equations, based on DBH and tree height (Sopp and Kolozs 2000). Quercus petraea, Q. robur and Q. cerris were merged as oaks, and rare tree species were merged as subordinate trees. Tree species diversity was calculated as Shannon diversity H’, based on the relative volume of tree species, using natural logarithm (Shannon and Weaver 1949). The volume of snags (standing dead wood) was calculated, based on the measurement of individuals within the plots. For dead trees the same equations were used as for living trees, for broken snags and stumps, the volume was calculated as a cylinder (measuring mean diameter and height). For lying dead wood, the mean diameter and the length of logs larger than 5 cm in diameter were also recorded, and log volume (per ha) was calculated by the cylinder formula. Saplings and shrubs below 5 cm DBH but above 0.5 m height were counted, and by species, their densities (unit per ha) were calculated.
The inventory of forest herbs was carried out in quadrats of 30 m × 30 m, positioned in the centre of the 40 m × 40 m tree plot in June and July of 2006. This means the early spring geophytes were excluded from the sample (they are relatively rare in the region). Absolute cover (dm2) was estimated for every recorded species, the nomenclature of vascular plants followed Tutin et al. (1964–1993). The cover of mineral soil and bryophytes were also estimated within the quadrats.
Relative diffuse light was calculated based on parallel measurements carried out in the quadrats and in nearby open fields with LAI-2000 Plant Canopy Analyzer instruments (LICOR Inc. 1992a, b; Tinya et al. 2009a, b). In each quadrate, 36 light measurements were taken in a systematically arranged grid, at 1.3 m height, using 5 m intervals. For quadrate level light conditions, the mean and coefficient of variation of the 36 relative diffuse light values were calculated.
Land cover types in a 300 m radius area around each plot were estimated with the help of aerial photos, maps and the forest stand database. Regenerating forests (<20 years), forests (older than 20 years) and non-forested areas (meadows and arable lands) were distinguished. Landscape diversity was calculated as the Shannon diversity H’ of land cover types. The land use history of the plots and their surroundings (300 m radius) was estimated, based on the Second Military Survey of the Habsburg Empire from 1853 (Arcanum 2006). The existence of forests in the plots (as a presence/absence variable) was recorded, and the proportion of forested areas in the historical landscape (in a circle of 300 m radius) was calculated.
Litter was collected from five systematically arranged, 30 × 30 cm areas from all quadrats. Litter weight, proportion of coniferous litter, litter pH (in water), organic carbon content and nitrogen content were measured. In the same points, soil samples were collected from the 0–10 cm layer. The following variables were measured from soil samples: pH in water using hydrolytic and exchangeable acidity measured by titration (Bellér 1997); clay (<0.002 mm), silt (0.002–0.02 mm) and sand (0.02–2 mm) fractions determined by sedimentation process (Cools and De Vos 2010); organic carbon and nitrogen content analysed by dry combustion elementary analysis using Elementar vario EL III CNS equipment (Elementar Analysensysteme GmbH 2000); ammonium-lactate/acetic-acid (AL-) extractable phosphorus and potassium content (Bellér 1997).
Air humidity and temperature were measured in the middle of the quadrats, at 1.3 m height, using Voltcraft DL-120 TH data loggers, in 24-h measurements with a 5-min recording frequency. The measurements of all quadrats were carried out within a 5-day period. During this period, two reference plots were measured permanently. Eight temperature and air humidity measurements were carried out during three vegetation periods (June and October 2009; June, August, September and October 2010; March and May 2011). For air humidity and temperature, differences were calculated from the two reference plots. Relative daily mean and range values were expressed for both variables and averaged over the eight measurements. The geographical positions of the plots were given in metres, based on the Hungarian Geographical Projection (EOV). For a summary of the potential explanatory variables, see Table 1.
Data analysis
Data structure was explored by ordination (Podani 2000; ter Braak and Smilauer 2002). In order to gain insight into the relations of the species and sites, we carried out principal component analysis (PCA) and, to determine the gradient length along the axes, detrended correspondence analysis (DCA). As the gradient lengths were less than three standard deviation units, redundancy analysis (RDA) was conducted, in order to identify the effects of explanatory variables on species composition (Table 1).
We carried out all analyses with log-transformed cover data of herbaceous species. Only species with a frequency larger than three were included. PCA and RDA were centred by species. Some explanatory variables were ln-transformed before the analyses, to fulfil normality conditions (proportions of tree species, light variables), and all explanatory variables were standardised (zero mean, one standard deviation). Within the RDA model, the explanatory variables were forward selected manually, by F statistics via Monte Carlo simulation (number of permutations was 499), and only significant (p < 0.05) variables were selected for the model. The significance of canonical axes was tested by similar Monte Carlo simulations. Latitude data, determining the geographical position of the plots, was used as a covariable within the RDA model.
To explore the effect of the explanatory variables on species richness and cover, general linear modelling (GLM) was used (Faraway 2005), using identity link function and Gaussian error structure. Ruderal and meadow-inhabiting species were separated from closed-forest herbs during the analysis; this classification is indicated in Table 1 in Online Resource 1. The classification was based on the habitat preference characterisation of the species of the Hungarian Flora (Simon 1992). Total species richness and species richness of closed-forest species were analysed separately, and the same approach was used for cover.
The dependent variables of the GLM models were in-transformed before the analyses, for a better fit of the model residuals to normality conditions. Linearity between the dependent and explanatory variables and constancy of the residual error variance were also checked after model selection. Before the statistical selection procedure, pairwise correlations and visual relationships between dependent and explanatory variables were investigated, and intercorrelations among explanatory variables were also considered. For the statistical selection procedure, only those explanatory variables were selected which showed a strong and consistent relationship with the dependent variable, and the intercorrelations with other explanatory variables were weak (r abs < 0.5). Although Poisson error structure is widely used for species richness models, we have chosen the Gaussian error structure because of better model diagnostics and coefficients of determination (Faraway 2005).
For multivariate analyses, Canoco for Windows 4.5 (ter Braak and Smilauer 2002), for all other analyses, R version 2.11.1 (R Development Core Team 2011) was used.
Results
We recorded a total of 134 species, 99 of which were labelled ‘closed-forest species’ (Simon 1992, Online Resource 1). Mean cover of the herbaceous layer in the plots was 4.08 %, with high standard deviation (7.22 % dm2). Species with the highest cover values were common woodland species, such as Rubus fruticosus, Oxalis acetosella, Pteridium aquilinum, Galeopsis pubescens, Galium odoratum and Ajuga reptans.
The PCA (see Figure 1 in Online Resource 3) revealed that most species are placed in one direction, in accordance with a distinction between species-rich and species-poor sites. The first axis explains 31.0 % of the total variance, the second 12.4 %.
In the RDA, the amount of diffuse light was the most important factor, but tree species richness, the relative volume of hornbeam, the proportion of fine-texture particles of the soil and moss cover were also significant (Table 2; Fig. 2). The first RDA axis explained 19.1 %, the second 8.9 % of the species variance, respectively, the first (F = 6.25, p = 0.002) and all other canonical axes (F = 3.67, p = 0.002) had significant effects.

Species (black) and environmental variables (red) biplot of the redundancy analysis (RDA). (Color figure online)
Herbaceous species preferring open areas (meadows, as Juncus effusus and Agrostis stolonifera, or clear-cuts, as Calamagrostis epigejos) and several acidophilus species (e.g. Luzula pilosa, Melampyrum pratense) correlate most strongly with light (Fig. 2). Many acidophilus species, however, are most strongly correlated with moss cover (Calluna vulgaris, Sieglingia decumbens and Carex pilulifera), itself also influenced by light availability. The variables referring to tree species composition (i.e. tree species richness and the amount of hornbeam) seemed to have a stronger effect on the herb species of closed forests (such as Sanicula europaea, Ajuga reptans, Viola reichenbachiana, Maianthemum bifolium and Athyrium filix-femina). The proportion of fine-texture ingredients in the soil act in the opposite direction to tree species richness, and hardly any species prefer the sites with a high clay and silt ratio (Veronica chamaedrys, Luzula luzuloides and Epipactis helleborine).
In the linear regression models, significant background factors were principally the same for all four explored-dependent variables (species richness and cover of all the species and closed-forest species; Table 3). The dependent variables were strongly correlated (after log-transformation, total cover and species richness: r = 0.845; p < 0.001; cover and species richness of closed-forest species: r = 0.843; p < 0.001; illustrated in Figures 6 and 7 in Online Resource 8 and 9, respectively). In the model for both total and closed-forest herb cover, the relative amount of diffuse light, tree species richness and landscape diversity proved significant with a positive sign, while the proportion of clay and silt in the soil had a negative effect. The explained variance, by chance, was 46 % for both of these models. In the case of species richness, again the same explanatory variables were the most effective for both studied groups (all and closed-forest species), namely the amount of relative diffuse light, tree species richness and landscape diversity, all with positive signs. The explained variance is 45 % for total species richness and 41 % for closed-forest species richness.
Scatterplots with correlation coefficients between the dependent and the selected explanatory variables and a table of the correlations can be found in Table 2 in Online Resource 2, and in Figures 2–5 in Online Resource 4–7, respectively.
Discussion
Direct and indirect effects of the significant environmental drivers
Our results show that from all the background factors examined in our study, the amount of diffuse light, tree species richness, the relative volume of hornbeam, silt and clay ratio of the soil, moss cover and landscape diversity are the most important factors correlating with the diversity and cover of the herbaceous layer. This means that the different aspects of the understory herb layer, i.e. species composition (explored by RDA), and species richness and cover (examined by GLM) are all chiefly influenced by the same environmental background factors. Most of these are not only directly operating agents, but also rather factors influenced by the same mechanisms as understory vegetation (Roberts and Gilliam 2014).
In our study, the most important driver was light. Light conditions on deciduous forest floors are evidently determined by canopy structure, or, more specifically, canopy openness. Whereas in such environments light is the most important limiting factor (Neufeld and Young 2014), its measurement is loaded with technical difficulties (e.g. Tinya et al. 2009a, b). For this study, we used LAI measurements, where the relative proportion of light is calculated by a comparison between values recorded in open areas and the forest site, at the same time.
The effect of light is not straightforward, because the abundance or diversity of the understory is not necessarily in direct proportion to light availability. In the study of Härdtle et al. (2003), light conditions were the most important drivers in acidophytic beech-oak forests, while on more neutral forest sites, different soil properties proved more determinant. Other studies have also come to the conclusion that the effects of light—or canopy openness—conditions depend on edaphic conditions (e.g. Tyler 1989; Van Couwenberghe et al. 2011). In our case, however, light was more important than soil conditions; the reason for this may lie, at least in part, in our sampling arrangements (stratified random method, resulting in similar edaphic conditions at all sites). Besides, the quality of soil itself is not independent of light availability (Strandberg et al. 2005; Van Calster et al. 2007) either.
Our studies were carried out in slightly acidic forests, and acidofrequent (e.g. Luzula pilosa, Melampyrum pratense) and non-forest (e.g. Juncus effusus, Calamagrostis stolonifera) species showed the strongest connection with light conditions. In our linear analyses, we found that light had a greater effect on total cover than on the cover of closed-forest species; however, even in the case of closed-forest species, light was the most important background factor, positively influencing both cover and species richness. This result is slightly contradictory. Studies on the subject tend to show that whereas total forest floor herb cover is increased by light, a denser canopy increases the proportion of forest dwelling species, which are more shade-tolerant (e.g. Schmidt et al. 2014; Vockenhuber et al. 2011). In our study area, most of the stands are highly closed (managed, even-aged stands, many of them dominated by beech), thus light availability is generally so low that, even for closed-forest species, this is the most important limiting factor. Similar findings have been reported elsewhere: Plue et al. (2013) conclude that even though the production of closed-forest species tends to improve with decreasing light conditions, this is only valid up to a given point; from then on, even these shade-tolerant species react strongly to small, favourable changes in light conditions. In the course of our project, Tinya et al. (2009a) explored in detail the effects of light on different understory species groups in the same forest stands. Their results show that a group of species, termed light-flexible species—mainly closed-forest species—correspond positively to light availability on a relatively small spatial scale. These species may be important in emphasising the importance of light availability in our models as well.
In the studied area, more acidic soils are linked to sparse pine forests, where ground floor light conditions are better. In these stands, ground floor mosses are also more abundant (Márialigeti et al. 2009). In the same stands, light conditions proved to be determinant in the diversity and composition of epiphytic lichens (Király et al. 2013; Nascimbene et al. 2012; Ódor et al. 2013) as well.
Terricolous bryophyte cover correlates strongly and positively with herb cover and species richness. Bryophyte and herbaceous species may act similarly to the same background conditions, apart from light availability (see also Tinya et al. 2009a), soil and litter properties, and microclimate. We recorded a number of features of these edaphic conditions (see Table 1). However, their correlations with the examined characteristics of the understory were weak and not significant. This implies that if understory herbs and bryophytes do indeed react to similar background factors, then these factors must be more complex than the ones we could measure.
It is possible that bryophytes directly facilitate the growth of vascular plants, by making the microenvironment more favourable. This has been known in more nutrient-limited habitats (peatlands—Groeneveld et al. 2007, cave entrances—Ren et al. 2010), and the same mechanisms may be at work in temperate forests as well.
Historic reasons may also be significant. Plots with former litter collection and foraging had become more acidic, which was beneficial to many bryophyte and herb species, and usually implied better light conditions as well (because of the presence of coniferous tree species).
Apart from light, other microclimatic conditions (such as temperature and air humidity) were not highly influential for understory herbs. These factors are more effective in the case of cryptogamic organisms, such as epiphytic lichens and bryophytes (Király et al. 2013; Ódor et al. 2013) and terricolous saprotrophic and mycorhizza fungi (Kutszegi et al. 2015).
Light availability may be the most important among the influencing factors determined by stand structure, but it is not the only one. We found that other characteristics of the stand structure also play important roles in the cover and species richness of the understory vegetation. As our sites host a wide variety of tree species and a broad range of their different compositions, we were able to study their influence.
Tree species richness acts as a positive factor for herb composition, species richness and cover. In the multivariate analysis, the presence of hornbeam is also a significant factor. Hornbeam is economically not preferred, but in many of our sites, it is nearly the only species which forms a second canopy layer. It is possible that the presence of the second canopy layer, and not hornbeam itself has relevant effects. Cook (2015), although admitting to the scarcity of literary data, hints that the abundance of canopy layers (shrubs, saplings or midstory) may create diverse resource limitations, thus enhancing the ecological possibilities of understory herbs. Although a second canopy layer reduces forest floor light availability and contributes to higher litter production, its presence or absence on a sub-patch scale creates highly variable microenvironments.
Stand structure and tree species composition directly determine litter properties and strongly influence the soil (e.g. Arno et al. 2012; Augusto et al. 2003) and microclimate of the site. The amount and composition of litter are fundamental for understory vegetation. In most of our forest patches, beech was dominant: on many locations, the plots consisted of even-aged beech trees, with vast amounts of litter and practically no understory (either herbaceous or bryophyte). Mölder et al. (2008), facing similar problems, found no significant effects of light conditions on the herb layer (its diversity or productivity), whereas the proportion of beech was crucial. They assumed that beech has indirect effects on ground floor vegetation through soil pH and litter production. Mölder et al. (2014) also point out that the growing proportion of beech, induced by the cessation of traditional management methods (in their case, coppicing with standards) acts negatively on the ground floor vegetation. Durak (2012) also notes the negative effect of beech and the strong connections between understory and overstory diversity. These findings correspond well to the conditions in our research sites: in our case, the unique variability of stand structure and composition has been brought about by centuries of various management types and is jeopardised by the introduction of the shelterwood system.
Vockenhuber et al. (2011) also examined both understory herbaceous plant species richness and cover, and their results highly correspond with ours. Both cover and species richness were enhanced by tree species diversity, and both responded negatively to increasing canopy cover.
Based on our study, it is expected that mixed forest stands help to maintain the biodiversity of the herb layer. Tree species richness was a crucial factor both for the total and only forest herb species richness. The importance of the heterogeneity of the tree species is also stressed in Macdonald and Fenniak (2007). They show that understory vegetation is linked to canopy composition and also edaphic conditions, with mixed and conifer forests differing significantly from broadleaved forests. Many studies show the great importance of canopy structure in maintaining understory diversity (e.g. Chávez and Macdonald 2010, 2012). As patches with different canopy types maintain various understory vegetation, all canopy patch types are important to maintain overall understory diversity.
Whereas soil properties are usually found among the most important factors for understory vegetation (e.g. Chávez and Macdonald 2010, 2012; Härdtle et al. 2003; Ikauniece et al. 2013; Van Couwenberghe et al. 2011), in our study, their significance was low. Although, as mentioned above, we have recorded several characteristics of the soil (including potassium and phosphorous content), these have not shown significant correlations with the examined variables. The reason for this may be the complexity of the interactions between trees and herbs, or the agency of some other edaphic factor, e.g. manganese, which, according to Muller (2014), may be a good indicator of general soil fertility. Finally, again, as in the case of bryophytes, it is also possible that herbs and trees react similarly to background factors (Gilliam and Roberts 2014).
In the case of cover (both total and closed-forest species), however, silt and clay content of the soil was a significant and negative background factor. The reason for this may be that increasing silt and clay content cause pseudogenization and poorer water conditions, which is not favourable to herb cover. This background factor had no significant effect on species richness, but a few species favour sites with higher silt and clay ratio. De Keersmaeker et al. (2004) suggest that soil properties (carbon, moisture, nitrogen and phosphorous) may be related to forest age. The forest stands in which our investigation has been carried out were selected to be more or less of the same age (70–100 years), so this variation is absent from our analyses; this may in part account for the lack of correlations.
Tree species composition also affects soil properties: Arno et al. (2012) demonstrated the varying effects of oak and poplar on the development of soil and understory vegetation in an experiment. They found that the soil differs under these two species, establishing the dissimilarities in understory vegetation. The positive effect of tree species richness on understory vegetation may also be conveyed through mixed litters, which are known to decompose more rapidly than litter from a single species (Gartner and Cardon 2004). The resulting fast nutrient cycling enhances the resources available to the understory layer.
Landscape diversity was an important factor for both cover and species richness for all and wood inhabiting species as well. Landscape diversity is generally thought to enhance species richness by acting as species pool for various species groups. High landscape diversity means that the forests are surrounded by other landscape types (mainly meadows and arable lands). The vicinity of these landscape elements can increase the species richness of the forests, adding many (primarily not forest specialist) species to the species pool. It is interesting that understory cover showed even stronger correlations with landscape diversity than species richness; the reason for this is not known.
Land use history was not an influential factor in species composition. It is possible that during the periods of intensive utilisation, the species sensitive to these effects partly disappeared [as suggested in the case of epiphytes in Király et al. (2013)], which means that the continuity of the forest (important for several, closed-forest species, e.g. Hermy et al. 1999) had been broken in the past (bottleneck effect, Økland et al. 2003), and its effects cannot be properly shown today.
Implications for conservation and management
The effects of changes in forest management are widely felt throughout the forests of Central Europe. The effects of the cessation of traditional management (e.g. coppicing in Germany, Mölder et al. 2014, and the Czech Republic, Vild et al. 2013, Hédl et al. 2010; leaving up chestnut forests in Bulgaria, Zlatanov et al. 2013) are revealed by several studies (this phenomenon is valid not only in forests, but other ecosystems as well, e.g. hayfields in Romania, Baur et al. 2006). The exact ecological mechanisms of the changes are often unclear; however, Kopecký et al. (2013) have demonstrated non-random extinction from the former species pool; a process that is hardly reversible. In our case, the unique versatility of the studied region is largely the result of past management practices (felling and using as arable lands and fodder removal). Diversity is largely kept up by present, close-to-nature management practices, typically in small, private forests (Schaich and Plieninger 2013). On the other hand, in our region, the shelterwood system has caused and is causing the homogenisation of stands (species composition as well as stand structure), which, according to our results, is disadvantageous for the diversity and productivity of the herbaceous vegetation.
Our results show that the most important factors affecting the composition, species richness and cover of understory herbs act on the stand level. We have also shown that the amount (and variability) of light is important, along with the presence of the second canopy layer and the diversity of tree species. Most of the relevant factors, such as canopy cover and tree species composition, can be directly affected by forest management. The analysis of species composition revealed that although light increases the diversity of forest herbs, too open conditions are favoured mainly by non-forest species (plants of the meadows and weeds of arable lands and clear-cuts). For the biodiversity of forest herbs, forest management should maintain heterogeneous light conditions including gaps and canopy openings, but the general light regime should be characterised as a high forest, to prevent the dominance of non-forest species. Management should also maintain high tree species diversity, including the mixed occurrence of the dominant species (Scots pine, beech and oaks) and a high proportion of non-dominant tree species, so as to enhance microenvironmental diversity (e.g. Arno et al. 2012) and, via litter mixing, promote nutrient cycling (Gartner and Cardon 2004). The presence of the secondary canopy layer (dominated by hornbeam) is also very important, as it also adds to the diversity of available resources (Cook 2015). These conditions can be provided by various types of management maintaining continuous forest cover, such as tree selection or group selection management systems (Matthews 1991).
Abbreviations
- LAI:
-
Leaf area index
- DBH:
-
Diameter at breast height
- RDA:
-
Redundancy analysis
- PCA:
-
Principal component analysis
- GLM:
-
General linear modelling
References
Allen CD, Savage M, Falk DA, Suckling KF, Swetnam TW, Schulke T, Stacey PB, Morgan P, Hoffman M, Klingel JT (2002) Ecological restoration of southwestern ponderosa pine ecosystems: a broad perspective. Ecol Appl 12:1418–1433. doi:10.2307/3099981
Arcanum (2006) Digitized maps of the Habsburg Empire. The Second Military Survey 1806–1869. DVD-Rom. Arcanum Kft., Budapest
Arno T, De Keersmaeker L, Van Calster H, De Schrijver A, Vandekerkhove K, Verstraeten G, Verheyen K (2012) Diverging effects of two contrasting tree species on soil and herb layer development in a chronosequence of post-agricultural forest. For Ecol Manag 278:90–100. doi:10.1016/j.foreco.2012.04.026
Augusto L, Dupouey J-L, Ranger J (2003) Effects of tree species on understory vegetation and environmental conditions in temperate forests. Ann For Sci 60:823–831. doi:10.1051/forest:2003077
Barbier S, Gosselin F, Balandier P (2008) Influence of tree species on understory vegetation diversity and mechanisms involved—a critical review for temperate and boreal forests. For Ecol Manag 254:1–15. doi:10.1016/j.foreco.2007.09.038
Bartels SF, Chen HYH (2013) Interactions between overstorey and understorey vegetation along an overstorey compositional gradient. J Veg Sci 24:543–552. doi:10.1111/j.1654-1103.2012.01479.x
Bauhus J, Puettmann KJ, Kühne C (2013) Close-to-nature forest management in Europe: does it support complexity and adaptability of forest ecosystems? In: Messier C, Puettmann KJ, Coates KD (eds) Managing forests as complex adaptive systems: building resilience to the challenge of global change. Routledge, The Earthscan Forest Library, pp 187–213. ISBN 978-0-415-51977-9
Baur B, Cremene C, Groza G, Rakosy L, Schileyko AA, Baur A, Stoll P, Erhardt A (2006) Effects of abandonment of subalpine hay meadows on plant and invertebrate diversity in Transylvania, Romania. Biol Conserv 132:261–273. doi:10.1016/j.biocon.2006.04.018
Bellér P (1997) Talajvizsgalati modszerek [Methods of soil analysis.] Egyetemi jegyzet, Soproni Egyetem, Erdőmérnöki Kar. Termőhelyismerettani Tanszék, Sopron
Berges L, Avon C, Verheyen K, Dupouey JL (2013) Landownership is an unexplored determinant of forest understory plant composition in Northern France. For Ecol Manag 306:281–291. doi:10.1016/j.foreco.2013.06.064
Catorci A, Vitanzi A, Tardella FM, Hršak V (2012) Trait variations along a regenerative chronosequence in the herb layer of submediterranean forests. Acta Oecol 43:29–41. doi:10.1016/j.actao.2012.05.007
Chávez V, Macdonald SE (2010) The influence of canopy patch mosaics on understory plant community composition in boreal mixedwood forest. For Ecol Manag 259:1067–1075. doi:10.1016/j.foreco.2009.12.013
Chávez V, Macdonald SE (2012) Partitioning vascular understory diversity in mixedwood boreal forests: the importance of mixed canopies for diversity conservation. For Ecol Manag 271:19–26. doi:10.1016/j.foreco.2011.12.038
Cook JE (2015) Structural effects on understory attributes in second-growth forests of northern Wisconsin, USA. For Ecol Manag 347:188–199. doi:10.1016/j.foreco.2015.03.027
Cools N, De Vos B (2010) Sampling and analysis of soil. Manual Part X. In: Manual on methods and criteria for harmonized sampling, assessment, monitoring and analysis of the effects of air pollution on forests, UNECE, ICP Forests, Hamburg. ISBN: 978-3-926301-03-1. http://www.icp-forests.org/Manual.htm. Accessed 2 Oct 2015
De Keersmaeker L, Martens L, Verheyen K, Hermy M, De Schrijver A, Lust N (2004) Impact of soil fertility and insolation on diversity of herbaceous woodland species colonizing afforestations in Muizen forest (Belgium). For Ecol Manag 188:291–304. doi:10.1016/j.foreco.2003.07.025
Dövényi Z (ed) (2010) Magyarország kistájainak katesztere [Cadastre of Hungarian regions]. MTA Földrajztudományi Intézet, Budapest
Duguid MC, Ashton MS (2013) A meta-analysis of the effect of forest management for timber on understory plant species diversity in temperate forests. For Ecol Manag 303:81–90. doi:10.1016/j.foreco.2013.04.009
Dupouey JL, Dambrine E, Laffite JD, Moares C (2002) Irreversible impact of past land use on forest soils and biodiversity. Ecology 83:2978–2984. doi:10.2307/3071833
Durak T (2012) Changes in diversity of the mountain beech forest herb layer as a function of the forest management method. For Ecol Manag 276:154–164. doi:10.1016/j.foreco.2012.03.027
Elementar Analysensysteme GmbH (2000) Brochure: http://www.elementar.de/fileadmin/user_upload/Elementar_Website/Downloads/Application_Notes/vario_EL_cube/AN-A-090609-D-01_01.pdf. Accessed 24 Sept 2015
Faraway JJ (2005) Linear models with R. Chapman and Hall, London
Gartner TB, Cardon ZG (2004) Decomposition dynamics in mixed-species leaf litter. Oikos 104:230–246. doi:10.1111/j.0030-1299.2004.12738.x
George LO, Bazzaz FA (2014) The herbaceous layer as a filter determining spatial pattern in forest tree regeneration. In: Gilliam FS (ed) The herbaceous layer in forests of Eastern North America. Oxford University Press, New York, pp 340–355
Gilliam FS (2007) The ecological significance of the herbaceous layer in temperate forest ecosystems. Bioscience 57:845–858. doi:10.1641/B571007
Gilliam FS, Roberts MR (2014) Interactions between the herbaceous layer and overstory canopy of Eastern forests. A mechanism for linkage. In: Gilliam FS (ed) The herbaceous layer in forests of Eastern North America. Oxford University Press, New York, pp 233–254
Godefroid S, Rucquoij S, Koedam N (2005) To what extent do forest herbs recover after clearcutting in beech forests? For Ecol Manag 210:39–53. doi:10.1016/j.foreco.2005.02.020
Gonzalez M, Augusto L, Gallet-Budynek A, Xue J, Yauschew-Raguenes N, Guyon D, Trichet P, Delerue F, Niollet S, Andreasson F, Achat DL, Bakker MR (2013) Contribution of understory species to total ecosystem aboveground and belowground biomass in temperate Pinus pinaster Ait. forests. For Ecol Manag 289:38–47. doi:10.1016/j.foreco.2012.10.026
Groeneveld EVG, Massé A, Rochefort L (2007) Polytrichum strictum as a nurse-plant in peatland restoration. Restor Ecol 15:709–719. doi:10.1111/j.1526-100X.2007.00283.x
Gyöngyössy P (2008) Gyantásország. Történeti adatok az őrségi erdők erdészeti és természetvédelmi értékeléséhez [Historical data to value forests in Őrség with a view to forest management and nature conservation]. Kerekerdő Alapítvány, Szombathely
Härdtle W, von Oheimb G, Westphal C (2003) The effects of light and soil conditions on the species richness of the ground vegetation of deciduous forests in northern Germany (Schleswig-Holstein). For Ecol Manag 182:327–338. doi:10.1016/S0378-1127(03)00091-4
Hart SA, Chen HYH (2006) Understory vegetation dynamics of north American Boreal Forests. Crit Rev Plant Sci 25:381–397. doi:10.1080/07352680600819286
Hédl R, Kopecký M, Komárek J (2010) Half a century of succession in a temperate oakwood: from species-rich community to mesic forest. Divers Distrib 16:267–276. doi:10.1111/j.1472-4642.2010.00637.x
Heinrichs S, Schmidt W (2009) Short-term effects of selection and clear cutting on the shrub and herb layer vegetation during the conversion of even-aged Norway spruce stands into mixed stands. For Ecol Manag 258:667–678. doi:10.1016/j.foreco.2009.04.037
Heithecker TD, Halpern CD (2006) Variation in microclimate associated with dispersed-retention harvests in coniferous forests of western Washington. For Ecol Manag 226:60–71. doi:10.1016/j.foreco.2006.01.024
Hermy M, Verheyen K (2007) Legacies of the past in the present-day forest biodiversity: a review of past land-use effects on forest plant species composition and diversity. Ecol Res 22:361–371. doi:10.1007/s11284-007-0354-3
Hermy M, Honnay O, Firbank L, Grashof-Bokdam C, Lawesson JE (1999) An ecological comparison between ancient and other forest plant species of Europe, and the implications for forest conservation. Biol Conserv 91:9–22. doi:10.1016/S0006-3207(99)00045-2
Hutchinson TF, Boerner REJ, Iverson LR, Sutherland S, Kennedy Sutherland E (1999) Landscape patterns of understory composition and richness across a moisture and nitrogen mineralization gradient in Ohio (U.S.A.) Quercus forests. Plant Ecol 144:177–189
Ikauniece S, Brūmelis G, Kasparinskis R, Nikodemus O, Straupe I, Zariņš J (2013) Effect of soil and canopy factors on vegetation of Quercus robur woodland in the boreonemoral zone: a plant-trait based approach. For Ecol Manag 295:43–50. doi:10.1016/j.foreco.2013.01.019
Ito S, Nakayama R, Buckley GP (2004) Effects of previous land-use on plant species diversity in semi-natural and plantation forests in a warm-temperate region in southeastern Kyushu, Japan. For Ecol Manag 196:213–225. doi:10.1016/j.foreco.2004.02.050
Kelemen K, Kriván A, Standovár T (2014) Effects of land-use history and current management on ancient woodland herbs in Western Hungary. J Veg Sci 25:172–183. doi:10.1111/jvs.12046
Kenderes K, Standovár T (2003) The impact of forest management on forest floor vegetation evaluated by species traits. Community Ecol 4(1):51–62. doi:10.1556/ComEc.4.2003.1.8
Király I, Nascimbene J, Tinya F, Ódor P (2013) Factors influencing epiphytic bryophyte and lichen species richness at different spatial scales in managed temperate forests. Biodivers Conserv 22:209–223. doi:10.1007/s10531-012-0415-y
Kopecký M, Hédl R, Szabó P (2013) Non-random extinctions dominate plant community changes in abandoned coppices. J Appl Ecol 50:79–87. doi:10.1111/1365-2664.12010
Kutszegi G, Siller I, Dima B, Takács K, Zs Merényi, Varga T, Turcsányi G, Bidló A, Ódor P (2015) Drivers of macrofungal species composition in temperate forests, West Hungary: functional groups compared. Fungal Ecol 17:69–83. doi:10.1016/j.funeco.2015.05.009
LI-COR Inc (1992a) LAI-2000 plant canopy analyzer instruction manual. LI-COR Inc., Lincoln
LI-COR Inc (1992b) 2000-90 Support software for the LAI-2000 plant canopy analyzer. LI-COR Inc., Lincoln
Lochhead KD, Comeau PG (2012) Relationships between forest structure, understorey light and regeneration in complex Douglas-fir dominated stands in south-eastern British Columbia. For Ecol Manag 284:12–22. doi:10.1016/j.foreco.2012.07.029
Macdonald SE, Fenniak TE (2007) Understory plant communities of boreal mixedwood forests in western Canada: natural patterns and response to variable-retention harvesting. For Ecol Manag 242:34–48. doi:10.1016/j.foreco.2007.01.029
Márialigeti S, Németh B, Tinya F, Ódor P (2009) The effects of stand structure on ground-floor bryophyte assemblages in temperate mixed forests. Biodivers Conserv 18:2223–2241. doi:10.1007/s10531-009-9586-6
Matthews JD (1991) Silvicultural systems. Oxford University Press, Oxford
Mikoláš M, Svoboda M, Pouska V, Morrissey RC, Donato DC, Keeton WS, Nagel TA, Popescu VD, Müller J, Bässler C, Knorn J, Rozylowicz L, Enescu CM, Trotsiuk V, Janda P, Mrhalová H, Michalová Z, Krumm F, Kraus D (2014) Comment on “Opinion paper: forest management and biodiversity”: the role of protected areas is greater than the sum of its number of species. Web Ecol 14:61–64. doi:10.5194/we-14-61-2014
Mölder A, Bernhardt-Römermann M, Schmidt W (2008) Herb-layer diversity in deciduous forests: raised by tree richness or beaten by beech? For Ecol Manag 256:272–281. doi:10.1016/j.foreco.2008.04.012
Mölder A, Streit M, Schmidt W (2014) When beech strikes back: how strict nature conservation reduces herb-layer diversity and productivity in Central European deciduous forests. For Ecol Manag 319:51–61. doi:10.1016/j.foreco.2014.01.049
Muller RN (2014) Nutrient relation of the herbaceous layer in deciduous forest ecosystems. In: Gilliam FS (ed) The herbaceous layer in forests of Eastern North America. Oxford University Press, New York, pp 13–34
Nascimbene J, Marini L, Ódor P (2012) Drivers of lichen species richness at multiple spatial scales in temperate forests. Plant Ecol Divers 5:355–363. doi:10.1080/17550874.2012.735715
Neufeld HS, Young DR (2014) Ecophysiology of the herbaceous layer in temperate deciduous forests. In: Gilliam FS (ed) The herbaceous layer in forests of Eastern North America. Oxford University Press, New York, pp 35–95
Nilsson MC, Wardle DA (2005) Understory vegetation as a forest ecosystem driver: evidence from the northern Swedish boreal forest. Front Ecol Environ 3:421–428
Ódor P, Király I, Tinya F, Bortignon F, Nascimbene J (2013) Patterns and drivers of species composition of epiphytic bryophytes and lichens in managed temperate forests. For Ecol Manag 306:256–265. doi:10.1016/j.foreco.2013.07.001
Økland T, Rydgren K, Halvorsen Økland R, Storaunet KO, Rolstad J (2003) Variation in environmental conditions, understorey species number, abundance and composition among natural and managed Picea abies forest stands. For Ecol Manag 177:17–37. doi:10.1016/S0378-1127(02)00331-6
Paillet Y, Berges L, Hjalten J, Odor P, Avon C, Bernhardt-Romermann M, Bijlsma R-J, De Bruyn L, Fuhr M, Grandin U, Kanka R, Lundin L, Luque S, Magura T, Matesanz S, Meszaros I, Sebastia M-T, Schmidt W, Standovar T, Tothmeresz B, Uotila A, Valladares F, Vellak K, Virtanen R (2010) Biodiversity differences between managed and unmanaged forests: metaanalysis of species richness in Europe. Conserv Biol 24:101–112. doi:10.1111/j.1523-1739.2009.01399.x
Plue J, Van Gils B, De Schrijver A, Peppler-Lisbach C, Verheyen K, Hermy M (2013) Forest herb layer response to long-term light deficit along a forest developmental series. Acta Oecol 53:63–72. doi:10.1016/j.actao.2013.09.005
Podani J (2000) Introduction to the exploration of multivariate biological data. Backhuys Publishers, Leiden
Priego-Santander AG, Campos M, Bocco G, Ramírez-Sánchez LG (2013) Relationship between landscape heterogeneity and plant species richness on the Mexican Pacific coast. Appl Geogr 40:171–178. doi:10.1016/j.apgeog.2013.02.013
R Development Core Team (2011) R. 2.14.0. A language and environment. www.rproject.org
Ren H, Ma G, Zhang Q, Guo Q, Wang J, Wang Z (2010) Moss is a key nurse plant for reintroduction of the endangered herb, Primulina tabacum Hance. Plant Ecol 209:313–320. doi:10.1007/s11258-010-9754-5
Roberts MR, Gilliam FS (2014) Response of the herbaceous layer to disturbance in eastern forests. In: Gilliam FS (ed) The herbaceous layer in forests of Eastern North America. Oxford University Press, New York, pp 321–339
Rosenvald R, Lohmus A (2008) For what, when, and where is green-tree retention better than clear-cutting? A review of the biodiversity aspects. For Ecol Manag 255:1–15. doi:10.1016/j.foreco.2007.09.016
Savill P (2004) Sylvicultural systems. In: Burley J, Evans J, Youngquist JA (eds) Enciclopedia of forest sciences. Elsevier, Amsterdam, pp 1003–1011
Schaich H, Plieninger T (2013) Land ownership drives stand structure and carbon storage of deciduous temperate forests. For Ecol Manag 305:146–157. doi:10.1016/j.foreco.2013.05.013
Schmidt M, Mölder A, Schönfelder E, Engel F, Schmiedel I, Culmsee H (2014) Determining ancient woodland indicator plants for practical use: a new approach developed in northwest Germany. For Ecol Manag 330:228–239. doi:10.1016/j.foreco.2014.06.043
Shannon CE, Weaver W (1949) The mathematical theory of communication. University of Illinois Press, Urbana
Simon T (1992) A magyarországi edényes flóra határozója. Tankönyvkiadó, Budapest 892 pp
Sopp L, Kolozs L (2000) Fatömegszámítási táblázatok. [Tables for calculating wood volume.] Állami Erdészeti Szolgálat, Budapest
Stefanovits P (ed), Filep Gy, Füleky Gy (1999) Talajtan. [Soil science] Mezőgazda Kiadó, Budapest
Strandberg B, Kristiansen SM, Tybirk K (2005) Dynamic oak-scrub to forest succession: effects of management on understorey vegetation, humus forms and soils. For Ecol Manag 211:318–328. doi:10.1016/j.foreco.2005.02.051
ter Braak CJF, Smilauer P (2002) Canoco 4.5. Biometris, Ceske Budejovice
Tímár G, Ódor P, Bodonczi L (2002) The characteristics of forest vegetation of the Őrség landscape protected area. Kanitzia 10:109–136
Tinya F, Márialigeti S, Király I, Németh B, Ódor P (2009a) The effect of light conditions on herbs, bryophytes and seedlings of temperate mixed forests in Őrség, Western Hungary. Plant Ecol 204:69–81. doi:10.1007/s11258-008-9566-z
Tinya F, Mihók B, Márialigeti S, Mag Z, Ódor P (2009b) A comparison of three indirect methods for estimating understory light at different spatial scales in temperate mixed forests. Community Ecol 10(1):81–90. doi:10.1556/ComEc.10.2009.1.10
Tyler G (1989) Interacting effects of soil acidity and canopy cover on the species composition of field-layer vegetation in oak/hornbeam forests. For Ecol Manag 28:101–114. doi:10.1016/0378-1127(89)90063-7
Tutin TG et al (eds) (1964–1993) Flora Europaea. Cambridge University Press, Cambridge
Van Calster H, Baeten L, De Schrijver A, De Keersmaeker L, Rogister JE, Verheyen K, Hermy M (2007) Management driven changes (1967–2005) in soil acidity and the understorey plant community following conversion of a coppice-with-standards forest. For Ecol Manag 241:258–271. doi:10.1016/j.foreco.2007.01.007
Van Couwenberghe R, Collet C, Lacombe E, Gégout JC (2011) Abundance response of western European forest species along canopy openness and soil pH gradients. For Ecol Manag 262:1483–1490. doi:10.1016/j.foreco.2011.06.049
Verheyen K, Guntenspergen GR, Biesbrouck B, Hermy M (2003) An integrated analysis of the effects of past land use on forest herb colonization at the landscape scale. J Ecol 91:731–742. doi:10.1046/j.1365-2745.2003.00807.x
Vild O, Roleček J, Hédl R, Kopecký M, Utinek D (2013) Experimental restoration of coppice-with-standards: response of understorey vegetation from the conservation perspective. For Ecol Manag 310:234–241. doi:10.1016/j.foreco.2013.07.056
Vockenhuber EA, Scherber C, Langenbruch C, Meißner M, Seidel D, Tscharntke T (2011) Tree diversity and environmental context predict herb species richness and cover in Germany’s largest connected deciduous forest. Perspect Plant Ecol Evol Syst 13:111–119. doi:10.1016/j.ppees.2011.02.004
von Arx G, Dobbertin M, Rebetez M (2012) Spatio-temporal effects of forest canopy on understory microclimate in a long-term experiment in Switzerland. Agric For Meteorol 166–167:144–155. doi:10.1016/j.agrformet.2012.07.018
Wayman RB, North M (2007) Initial response of mixed-conifer understory plant community to burning and thinning restoration treatments. For Ecol Manag 239:32–44. doi:10.1016/j.foreco.2006.11.011
Wulf M (2003) Preference of plant species for woodlands with differing habitat continuities. Flora 198:444–460. doi:10.1078/0367-2530-00118
Zlatanov T, Schleppi P, Velichkov I, Hinkov G, Georgieva M, Eggertsson O, Zlatanova M, Vacik H (2013) Structural diversity of abandoned chestnut (Castanea sativa Mill.) dominated forests: implications for forest management. For Ecol Manag 291:326–335. doi:10.1016/j.foreco.2012.11.015
Zmihorski M, Chylarecki P, Rejt L, Mazgajski TD (2010) The effects of forest patch size and ownership structure on tree stand characteristics in a highly deforested landscape of central Poland. Eur J For Res 129:393–400. doi:10.1007/s10342-009-0344-9
Acknowledgments
We thank László Bodonczi, Zsuzsa Mag, István Mazál, Ákos Molnár, Péter Szűcs and Balázs Németh for their help in the field survey, and Anna Sáfár for careful language editing. The project was funded by the Hungarian Science Foundation (OTKA 79158), Őrség National Park Directorate and the TÁMOP-4.2.2.D-15/1/KONV-2015-0023 project of the European Union and European Social Found. Péter Ódor was supported by the Bolyai János Research Scholarship of the Hungarian Academy of Sciences.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
The authors declare that they have no conflicts of interest.
Additional information
Communicated by Thomas A. Nagel.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Márialigeti, S., Tinya, F., Bidló, A. et al. Environmental drivers of the composition and diversity of the herb layer in mixed temperate forests in Hungary. Plant Ecol 217, 549–563 (2016). https://doi.org/10.1007/s11258-016-0599-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-016-0599-4