
Abstract
Prescribed burning has been a successful ecosystem restoration tool worldwide. In this study, we burned two different degraded Mongolian grasslands in spring: sparsely vegetated Argalant (ARG) and Hustai National Park (HNP), dominated by the unpalatable forb Artemisia adamsii. We investigated soil properties, seed bank, seedling density, vegetation, biomass, and forage quality before and/or after burning in the grasslands. Burning weakly affected soil properties and significantly reduced seed bank density. Seedling density of Chenopodium aristatum was greater and that of Cleistogenes squarrosa was lower in burned than in unburned plots of HNP, although C. squarrosa showed greater vegetative growth in burned plots. The floristic composition showed small changes after burning, depending on the pre-burning vegetation, but cover of A. adamsii decreased after burning in both ARG and HNP. Thus, the reduced seed bank density by burning led to weak responses of vegetation composition to burning due to strong vegetative growth. However, burning suppressed the unpalatable forb (A. adamsii) and hastened palatable grass (C. squarrosa) productivity, which would be attractive for livestock production, especially for overgrazed areas with low forage palatability. Our findings illustrate the possibility of using spring prescribed burning as a restoration tool in degraded Mongolian steppe, as well as the importance of site-specific conditions for understanding postfire recovery.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The degradation of Mongolian steppe by overgrazing is of global concern (Yoshihara et al. 2008). Although grazing exclusion with fencing is the usual management option for restoration of degraded steppe, the recovery of vegetation within enclosures depends on rainfall events, and a long time is required to achieve recovery (Sasaki et al. 2009). In the face of the current rapid degradation of steppe ecosystems in Mongolia, there is an increasing need for the development of prompt management prescription for restoration.
Prescribed or controlled burning is widely regarded as an effective vegetation restoration option for degraded grassland (Moyes et al. 2005). Burning has been shown to enhance grass production by improving light conditions due to reduced self-shading, accelerating nitrogen mineralization, and increasing soil temperature and soil mineral elements due to deposition of dark ash (Hobbs and Schimel 1984; Gibson 2009). In the second postfire year, the diversity and seed density increase due to the contribution of fugitive species (Gonzalez and Ghermandi 2008). Specifically in rangelands, prescribed burning is expected to promote livestock production because restored plants after fire generally have high nutrient levels and palatability (Allen et al. 1976; Moe and Wegge 1997). However, arid environments have unique plant succession after disturbance, such that the general principles developed in more mesic environments cannot be applied. For example, an early study by Launchbaugh (1964) showed that production was markedly decreased in an arid environment following fire, and three growing seasons were needed to recover from the fire. In Mongolia, there was an average of 160 wild fires per year between 1981 and 1999, which increased to 188 between 2000 and 2008, with an average of 2,933,659 to 3253,000 ha being burned each year (Darren et al. 2009). Despite this trend, fire management on the steppe of Mongolia has not yet been investigated.
Here, we carried out field experiments and investigated changes caused by burning and its effects on vegetation, soil, and seed bank dynamics in semiarid Mongolian steppe. Because species-specific responses to burning have led to inconstant conclusions depending on local conditions (Anderson and Menges 1997; Todd et al. 2000; Rau et al. 2008), we burned two different types of degraded rangelands and compared their responses to burning. The characteristic vegetation responses to overgrazing in Mongolian grassland have been a reduction of plant density, seen mainly in more arid environments (Sasaki et al. 2009), or replacement by unpalatable species, seen mainly in more mesic environments (Yoshihara et al. 2008). Argalant (ARG) is a village in Mongolia’s steppe region and is characterized by sparse vegetation due to heavy grazing. It therefore corresponds to the first type of degraded rangeland. Hustai National Park (HNP) has been protected from livestock grazing for conservation purposes for the past 15 years. In areas that experienced severe grazing before the protection, however, the unpalatable perennial rhizomatous forb Artemisia adamsii still remains, and thus HNP represents the latter type of overgrazed rangeland. Artemisia adamsii is a strong competitor under severe grazing and is thus recognized as an indicator of pasture degradation in Mongolian steppe (Yoshihara et al. 2010). The aim of prescribed burning in Mongolia is to reduce the coverage of A. adamsii and accelerate the production of palatable grasses for livestock production.
Methods
Study area
The study areas are located 100 km west of Ulaanbaatar (47°50.0′N, 106°00.0′E), in Mongolia’s steppe region. The ARG and HNP areas are located about 30 km apart. The sites received 232 mm of annual precipitation, averaged over the past decade. The annual average temperature is 0.2 °C, and average monthly temperatures vary greatly: between −20.6 °C in January and 19.0 °C in July. The soils are Haplic Kastanozems as identified by the world reference base for soil resources (FAO 1988) based on soil profile morphology and physico-chemical properties. Stipa krylovii is a palatable perennial bunch grass that dominates less-grazed areas of Mongolian steppe; Carex spp. and Cleistogenes squarrosa are rhizomatous and bunch palatable perennial graminoids growing under medium grazing, and A. adamsii and Chenopodium spp. are degradation-indicator ruderal perennial and annual forbs (Hoshino et al. 2009; Yoshihara et al. 2010).The study areas have a long history (centuries) of being grazed by domestic livestock under nomadic patterns of land use. The main livestock in this province are sheep (50 %), goats (36 %), horses (7.5 %), and cattle (6.5 %) (National Statistical Office of Mongolia 2011), and wild horses also inhabit the area.
We applied prescribed burning to four 20 m × 20 m sites (blocks) in each of the ARG and HNP study areas in April 2012. The soil surface temperature around burning is shown in Fig. S1. The burned areas were fenced soon after the burning to prevent disturbance from livestock and wildlife.
Soil survey
We randomly established four plots (each 1 m2) in the experimental sites. To investigate the soil properties, we extracted soil samples (10 cm depth) from each plot in October 2011 (pre-burning), April 2012 (immediate after burning), and August 2012 (4 months after burning). At each site, we extracted 12 samples in total and homogenized the samples before analysis. The soil samples were air-dried and then analyzed. Soil pH was measured with a glass electrode, and electric conductivity (EC) was measured with an EC meter. Soil carbon contents were analyzed using dry combustion (Matejovic 1993). Exchangeable soil Ca and Mg and available K2O were measured using atomic absorption spectrophotometry (Gallaher et al. 1975). Available soil phosphorus (P2O5) was measured using the Truog–Bray method.
Seed bank survey
To investigate soil seed bank abundance and composition, soil sampling was conducted prior to and immediately after burning at each site, except for one ARG site that was added after the pre-burning survey was conducted. At each site, 16 sampling points were set on a lattice grid with 4-m intervals, and two soil samples were collected at each point using a core sampler at two depths: upper layer (0–1 cm deep with 20-cm2 surface area) and lower layer (1–6 cm deep with 20-cm2 surface area). Plants and litter were removed before soil sampling.
We then conducted a germination experiment in incubators. For pre-burning soil samples, which were collected before winter, cold stratification was conducted in plastic bags with wet conditions for 8 weeks at 4 °C in a dark incubator, to enhance release from seed dormancy (Ronnenberg et al. 2008). The soil samples were then spread thinly (<1 cm thick) on plastic trays filled with seed-free sand, and the germination test was performed at 10/20 °C (12/12 h) with dark/light (12/12 h) conditions in incubators. All soils were moistened arbitrarily and were stirred after 1 month to enhance germination from the bottom soil layer. Germinated seedlings were counted and species were identified every 2–7 days for 2 months.
Seedling recruitment and vegetative growth
To examine the effects of burning on seedling recruitment, a seedling survey was conducted in August 2012. This survey was not conducted in ARG because of less seedling emergence. We established four plots (1 m × 1 m) inside the four HNP sites (burned plots) and another four outside the site fence (unburned plots) and recorded current-year seedlings. We counted two predominant seedling species: the annual forb Chenopodium aristatum and the perennial grass C. squarrosa. The number of seedlings of the other species was low (<30 in total).
To examine the response of vegetative growth to burning, we investigated the plant size distribution of C. squarrosa in August 2013. Cleistogenes squarrosa is a C4 bunchgrass that forms short and dense tussocks. C4 grasses are expected to increase under burning (Bond et al. 2005). Because the basal area of C. squarrosa tussocks is related to the number of current-year tillers (Liang et al. 2002), we can evaluate vegetative growth of this species based on basal area. We established four plots (50 cm × 50 cm) inside and outside fences of the four HNP sites, as burned and unburned plots, respectively. We measured the long and short diameters of all C. squarrosa tussocks within the plots and then calculated the tussock basal area.
Plant cover and biomass
We conducted vegetation surveys in August of 2011, 2012, and 2013 (pre-burning and 4 and 16 months after burning). We established four quadrats (1 m × 1 m) inside the fence at all ARG and HNP sites, and measured the percent cover of each plant species.
We took a sample of aboveground plant biomass from four established sub-quadrats (50 cm × 50 cm) at each site at the same time that we conducted the vegetation survey. Because the study plots were excluded from grazing pressure after burning by fencing, we could not examine the true effects of burning on plant biomass in rangeland. Therefore, we established four control plots without burning treatment within each fenced site and collected plant biomass in August 2013. The collected samples were oven-dried to constant weight at 75 °C for 48 h. The samples collected in 2011 and 2012 were chemically analyzed in the laboratory to determine the crude protein, total fat, crude fiber, and mineral content (Ca, Mg, K, Na). Crude protein concentrations in the plant samples were measured by determining nitrogen using the Kjeldahl method, then multiplying the result by 6.25 to convert the nitrogen content into crude protein. Total fat was measured using the method of AOAC (1990). The concentration of crude fiber was determined according to the method described by Van Soest (1994). Atomic absorption spectroscopy was used to determine other mineral element concentrations.
Statistical analysis
We analyzed the soil properties using repeated-measures ANOVA to examine the main effects of period at each sampling location. The seed bank density and species richness were analyzed for each location using a generalized linear mixed-effect model (GLMM) with a Poisson distribution and site ID as a random intercept. The number of seeds for all species and three functional types (annual forb, perennial forb, and perennial graminoid) and species richness were calculated using the sum of the upper and lower soil samples for each sampling point. The number of C. aristatum and C. squarrosa seedlings was analyzed by ANOVA with a random intercept of experimental sites. The cumulative basal area of C. squarrosa per each plot was compared between unburned and burned plots using ANOVA with a random intercept of experimental sites. Detrended correspondence analysis (DCA) was applied to assess the effect of burning and temporal patterns of changes in floristic composition, based on the cover of each species. To downweight rare species, we removed species occurring in fewer than a threshold number of species (i.e., 0.2 % of mean plant cover). To compare plant biomass between treatments (pre-burning and 4 and 16 months after burning and fenced plots), we used ANOVA with a random intercept of experimental sites. When necessary, data were log-transformed to meet the assumptions of normality and similarities of variance.
Results
Soil properties
Soil pH was decreased and soil K concentration was increased by burning in ARG. Mineral concentrations in each plot showed different responses to burning, and thus there was no significant trend in other soil properties (EC, TC, Ca, Mg, and P) in ARG or in all soil properties in HNP after burning.
Seed bank
The germinable seed bank comprised 315 seeds of 19 species before burning (Table S1). The three plant functional types (i.e., annual forbs, perennial forbs, and perennial graminoids) accounted for more than 80 % of these, and perennial forbs dominated. Most seeds (40–50 %) were found in the upper layer (0–1 cm, 1.2 seeds/20 cm3) in comparison with those in the lower layer (1–6 cm, 1.7 seeds/100 cm3) (Table S1). The seed density of all species and species richness were lower after burning than before burning in both areas (Fig. 1). The seed densities of annual forbs and perennial forbs were also lower after burning in both areas, but that of perennial graminoids did not differ before and after burning in HNP (Fig. 1). The seeds of A. adamsii, Carex spp., and C. squarrosa did not germinate after burning, whereas Stipa sp. seeds were found both before and after burning, particularly in HNP (Table S1).

a Seed bank density and b species richness of all species, and (c–e) seed density of three functional types within each soil core (100 cm−3) in ARG and HNP before and after burning treatment. Boxes indicate 75, 50, and 25th percentiles, and whiskers indicate 90 and 10th percentiles. Circles show outliers
Seedling emergence and vegetative growth
The seedling density of C. aristatum was higher and that of C. squarrosa was lower in burned than in unburned plots of HNP (Table 1).
The cumulative basal area of C. squarrosa was higher in burned than in unburned plots (Fig. 2a, F 1,27 = 12.24, P < 0.01). The size distribution of C. squarrosa tended to differ between the treatments (Fig. 2b, c). The most frequent size of C. squarrosa tussocks was <10 cm2 in basal area in both unburned and burned plots, whereas the tussocks larger than 40 cm2 were recorded only in burned plots.

a Cumulative basal area of Cleistogenes squarrosa tussocks in unburned and burned plots and (b, c) the size class distribution of basal area
Vegetation
Before burning, perennial species were dominant, and there were fewer annual species in both ARG and HNP (Table S2). Artemisia adamsii showed the greatest coverage in HNP, and Carex spp. and C. squarrosa were also dominant (Table S2). Artemisia adamsii also appeared in all the ARG sites, and Achnatherum splendens and Artemisia frigida dominated. The cumulative plant cover was about 54–63 % in ARG and about 70–80 % in HNP both before and after burning (Table 2).
DCA results showed that the changes in floristic composition by burning differed among sites, although the temporal changes did not override the floristic differences among sites in the original vegetation before burning (Fig. 4). In ARG, the trajectory of two of four sites fluctuated along axis I; one site moved to and another site moved away from zero of axis I, where Carex spp. was arrayed (Fig. 3a, b). The trajectory of three of four sites moved to the upper side of axis II, and the species score of A. adamsii was arrayed on the lower side of axis II. In HNP, the DCA scores of four sites departed from the coordinate origin, where A. adamsii was arrayed (Fig. 3c, d). The site score of one site moved far to the left side of axis I, where Carex spp. was arrayed. These results indicated that the cover of A. adamsii decreased after burning in both ARG and HNP, whereas the cover of Carex spp. increased or decreased site-specifically after burning. No trajectories of any sites in either ARG or HNP reached the coordinate for S. krylovii.

DCA ordination diagrams of site and species scores in (a, b) ARG and (c, d) HNP. Trends in floristic composition from 2011 to 2013 are indicated by arrows. The centroids of DCA values for four quadrats of each site are shown. Species codes: Achnatherum splendens, A spl; Allium bidentatum, A bid; Artemisia adamsii, A ada; Artemisia frigida, A fri; Artemisia palustris, A pal; Astragalus brevifolius, A bre; Caragana leucophloea, C leu; Carex spp., Carex; Chenopodium aristatum, C ari; Cleistogenes squarrosa, C squ; Convolvulus ammannii, C amm; Cymbaria dahurica, C dah; Elymus chinensis, E chi; Heteropappus hispidus, H his; Potentilla bifurca, P bif; Stipa krylovii, S kry
Plant biomass
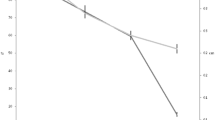
Plant biomass differed significantly among treatments in both ARG (F = 9.148, P = 0.002) and HNP (F = 8.340, P = 0.003; Fig. 4). Average plant biomasses at 4 and 16 months after burning were 48 and 53 g/m2 in ARG and 32 and 19 g/m2 in HNP greater than those in pre-burned plots in both cases.

Changes in plant biomass (mean ± standard error) after burning treatments and at the control sites (without burning and grazing) in ARG and HNP
Forage qualities
We found no significant differences in pasture forage qualities before and after burning, except for a difference in crude fiber content in HNP, judged at P = 0.05 (Fig. 5). In HNP, mineral concentrations of grass after burning were increased in the plots where the concentration was low before burning (Fig. 5). The average Mg, K, and Na concentrations of grass after burning were more than 2, 5, and 1.5 times those in pre-burned plots, whereas Ca was decreased. For ARG, there were no significant mineral concentration changes after the burning.

Plant nutritive values and mineral element concentrations at each site before and after burning treatment. CP crude protein, CF crude fiber. Solid and dashed lines indicate ARG and HNP, respectively
Discussion
The responses of seed bank survival and germination to burning differed depending on climate, vegetation, and fire type (Laterra et al. 2006; Gonzalez and Ghermandi 2008). In fire-prone chaparral, fire is needed to release seeds from dormancy and induce seed germination (Keeley and Fotheringham 1998; Odion and Davis 2000; Auld and Denham 2006). In contrast, for fire-intolerant species, burning often reduces seed bank survival due to the effects of smoke, heat, and physical scarification (Odion and Davis 2000; Mamede and de Araújo 2008). In this study, seed bank density and species richness decreased remarkably after burning. Seeds would have no tolerance to burning damage (e.g., heat) in Mongolian steppe, which has not historically suffered from burning disturbance (Darren et al. 2009).
In addition, the response of seed bank survival to burning was dependent on plant functional type. The numbers of seeds of annual and perennial forbs were reduced after burning, whereas the seeds of desirable perennial graminoids, particularly Stipa spp., remained viable even after burning in HNP. In a semiarid Patagonian grassland, the seed bank of annual species was not susceptible to burning because the small seeds are buried deeply in the soil (Gonzalez and Ghermandi 2008). In contrast, in Mongolian steppe, seeds suffer burning damage because most seeds exist within the thin surface soil (Auld and Denham 2006). For perennial graminoids, however, the lemma covering Stipa seeds would protect them from burning damage (Franzese and Ghermandi 2012).
In the perennial-dominated Mongolian steppe, seedling recruitment was originally rare, but the pattern was altered by burning. The number of C. aristatum seedlings increased, and those of C. squarrosa decreased after burning. The lower seedling emergence of C. squarrosa after burning was consistent with the few seeds of this species in the seed bank after burning, although we cannot explain the higher number of C. aristatum seedlings after burning.
In contrast to seedling recruitment, burning enhanced the vegetative growth of the desirable perennial grass C. squarrosa. C. squarrosa had a larger tussock-size distribution in burned sites. Burning does not influence the photosynthesis response of C4 C. squarrosa (Zhang et al. 2008), but tussock litter removal by burning appears to enhance vegetative growth of the tussock (Gao et al. 2007; Wang et al. 2011). Therefore, burning would change the plant recruitment strategy and the population size distribution via changes in the balance of seed and vegetative recruitment for perennial species.
In Mongolian steppe, total plant cover did not seem to be altered by burning and had recovered by 4 months after burning. Plant species composition also did not show a large change after burning and depended strongly on the pre-burning vegetation. These weak responses of vegetation to burning are likely due to the predominantly vegetative regrowth of perennial species with less seed recruitment. Although burning reduced the seed bank and altered the subsequent seedling response, seed recruitment would contribute less to postfire revegetation.
Nonetheless, burning tended to decrease the cover of the undesirable perennial forb A. adamsii. Previous studies have shown that burning suppresses unpalatable plant species and enhances the production of palatable plants (e.g., Augustine and Milchunas 2009). In a shortgrass steppe, prescribed burning reduced an unpalatable subshrub species (Compositae) by increasing mortality (McDaniel et al. 1997; Augustine and Milchunas 2009). The vegetative growth of A. adamsii, which forms a woody stem, should be susceptible to damage by burning, unlike perennial herbaceous species, which have little aboveground biomass in spring. Furthermore, the decrease of A. adamsii cover enhances the postfire growth of C. squarrosa, which is a weak competitor (Gao et al. 2005, 2007; Wang et al. 2008).
In ARG, although plant biomass was increased by burning, the degree of change was comparable to that of the control plots, indicating that this increase was achieved not due to the burning effect but rather from the absence of grazing. In HNP, however, plant biomass after burning was greater than control plots, which would be attributed to the burning effect. Compared to ARG, HNP had greater coverage of A. adamsii, which is a short plant, and thus plant biomass per unit area is not large. The relative increase of grass instead of A. adamsii is the reason why biomass increased after burning in HNP. Moreover, burning removed plant litter on the ground in HNP (Augustine and Milchunas 2009), improved light conditions, and facilitated plant growth (Gittins et al. 2011). Indeed, the plant biomass of C. squarrosa was larger after burning (Fig. 2).
Implications
In ARG, the seed bank density and productivity were decreased by burning. In HNP, burning caused the composition of the seed bank to shift from forbs to perennial graminoids (e.g., Stipa sp.) and the plant species composition to shift from A. adamsii to Carex spp. Because the recovery pattern after burning differed markedly depending on the pre-burning vegetation communities, the existing community must be considered when using burning as a restoration tool in Mongolian overgrazed grassland. Where vegetation is sparse due to overgrazing, as in ARG, burning would worsen the situation. However, where sufficient vegetation exists but there has been a reduction in forage palatability by overgrazing, as in HNP, burning would be desirable because it would suppress the unpalatable forbs (A. adamsii) and hasten palatable graminoid production (Carex spp., Stipa sp.).
Historically, Mongolian pastoralists did not use fire, and fire was perceived negatively because they were unaware of any benefits resulting from either wild or controlled fire (Darren et al. 2009). The limitation of our results is not a complete factorial design with burning and grazing factors. However, this study is the first to examine the utility of spring prescribed burning as a restoration tool of degraded Mongolian steppe. Our findings also reveal the importance of considering site-specific conditions for understanding postfire recovery.
Spring burning would accelerate the decline of plant production in sparse vegetated grasslands. Spring burning may enhance the vegetation shift from the dominance of unpalatable forbs to palatable graminoids in degraded steppe formerly dominated by unpalatable forbs. In dryland rangelands, the effectiveness of burning for improving productivity is site dependent.
References
Allen LJ, Harbers LH, Schalles RR, Owensby CE, Smith EF (1976) Range burning and fertilizing related to nutritive value of bluestem grass. J Range Manage 29:306–308
Anderson R, Menges E (1997) Effects of fire on sandhill herbs: nutrients, mycorrhizae, and biomass allocation. Am J Bot 84:938–948
AOAC (1990) Official methods of analysis. Association Official Analytical Chemists
Augustine DJ, Milchunas DG (2009) Vegetation responses to prescribed burning of grazed shortgrass steppe. Range Ecol Manage 62:89–97
Auld T, Denham A (2006) How much seed remains in the soil after a fire? Plant Ecol 187:15–24
Bond WJ, Woodward FI, Midgley GF (2005) The global distribution of ecosystems in a world without fire. New Phytol 165:525–538
Darren J, Byambasuren O, Myers RL, Babler M (2009) Fire management assessment of the eastern steppe, Mongolia. GFI technical report (2009)-1a. The Nature Conservancy, Arlington
FAO (1988) FAO/Unesco soil map of the world. Revised legend, with corrections and updates. World Soil Resources Report 60. FAO, Rome
Franzese J, Ghermandi L (2012) Effect of fire on recruitment of two dominant perennial grasses with different palatability from semi-arid grasslands of NW Patagonia (Argentina). Plant Ecol 213:471–481
Gallaher RN, Weldon CO, Futral JG (1975) An aluminum block digester for plant and soil analysis. Soil Sci Soc Am J 39:803–806
Gao YZ, Wang SP, Han XG, Patton BD, Nyren PE (2005) Competition between Artemisia frigida and Cleistogenes squarrosa under different clipping intensities in replacement series mixtures at different nitrogen levels. Grass Forage Sci 60:119–127
Gao YZ, Wang SP, Han XG, Chen QS, Zhou ZY, Patton BD (2007) Defoliation, nitrogen, and competition: effects on plant growth and resource allocation of Cleistogenes squarrosa and Artemisia frigida. J Plant Nut Soil Sci 170:115–122
Gibson DJ (2009) Grasses and grassland ecology. Oxford University Press, New York, p 305
Gittins C, Ghermandi L, Bran D (2011) Studying the post-fire performance of tussock grasses in Patagonia: survival, biomass production and early competition. J Arid Environ 75:986–990
Gonzalez S, Ghermandi L (2008) Postfire seed bank dynamics in semiarid grasslands. Plant Ecol 199:175–185
Hobbs NT, Schimel DS (1984) Fire effects on nitrogen mineralization and fixation in mountain shrub and grassland communities. J Range Manage 37:402–405
Hoshino A, Yoshihara Y, Sasaki T, Okayasu T, Jamsran U, Okuro T, Takeuchi K (2009) Comparison of vegetation changes along grazing gradients with different numbers of livestock. J Arid Environ 73:687–690
Keeley JE, Fotheringham CJ (1998) Smoke-induced seed germination in California chaparral. Ecology 79:2320–2336
Laterra P, Ortega EZ, Ochoa MC, Vignolio OR, Fernández ON (2006) Interactive influences of fire intensity and vertical distribution of seed banks on post-fire recolonization of a tall-tussock grassland in Argentina. Aust Ecol 31:608–622
Launchbaugh JL (1964) Effects of early spring burning on yields of native vegetation. J Range Manage 17:5–6
Liang C, Michalk DL, Millar GD (2002) The ecology and growth patterns of Cleistogenes species in degraded grasslands of eastern Inner Mongolia, China. J Appl Ecol 39:584–594
Mamede MD, de Araújo FS (2008) Effects of slash and burn practices on a soil seed bank of caatinga vegetation in northeastern Brazil. J Arid Environ 72:458–470
Matejovic I (1993) Determination of carbon, hydrogen, and nitrogen in soils by automated elemental analysis (dry combustion method). Commun Soil Sci Plant Anal 24:2213–2222
McDaniel KC, Hart CR, Carroll DB (1997) Broom snakeweed control with fire on New Mexico blue grama rangeland. J Range Manage 50:652–659
Moe SR, Wegge P (1997) The effects of cutting and burning on grass quality and axis deer (Axis axis) use of grassland in lowland Nepal. J Tropical Ecol 13:279–292
Moyes AB, Witter MS, Gamon JA (2005) Restoration of native perennials in a California annual grassland after prescribed spring burning and solarization. Rest Ecol 13:659–666
National Statistical Office of Mongolia (2011) Mongolian statistical yearbook 2010. National Statistical Office of Mongolia, Ulaanbaatar
Odion DC, Davis FW (2000) Fire, soil heating and the formation of vegetation patterns in chaparral. Ecol Monog 70:149–169
Rau BM, Chambers JC, Blank RR, Johnson DW (2008) Prescribed fire, soil, and plants: burn effects and interactions in the central great basin. Rangeland Ecol Manage 61:169–181.
Ronnenberg K, Wesche K, Hensen I (2008) Germination ecology of Central Asian Stipa spp.: differences among species, seed provenances, and the importance of field studies. Plant Ecol 196:269–280
Sasaki T, Okayasu T, Ohkuro T, Shirato Y, Jamsran U, Takeuchi K (2009) Rainfall variability may modify the effects of long-term exclosure on vegetation in Mandalgobi, Mongolia. J Arid Environ 73:949–954
Todd PA, Phillips JDP, Putwain PD, Marrs RH (2000) Control of Molinia caerulea on moorland. Grass Forage Sci 55:181–191
Van Soest PJ (1994) Nutritional ecology of the ruminant. Cornell University Press
Wang JH, Wang SP, Schnug E, Haneklaus S, Patton B, Nyren P (2008) Competition between Stipa grandis and Cleistogenes squarrosa. J Arid Environ 72:63–72
Wang S, Zhao X, Qu H, Zuo X, Lian J, Tang X, Powers R (2011) Effects of shrub litter addition on dune soil microbial community in Horqin sandy land, northern China. Arid Land Res Manage 25:203–216
Yoshihara Y, Chimeddorj B, Buuveibaatar B Lhagvasuren, Takatsuki S (2008) Effects of livestock grazing on pollination on a steppe in eastern Mongolia. Biol Conserv 141:2376–2386
Yoshihara Y, Okuro T, Buuveibaatar B, Undarmaa J, Takeuchi K (2010) Clustered animal burrows yield higher spatial heterogeneity. Plant Ecol 206:211–224
Zhang Y, Niu S, Xu W, Han Y (2008) Species-specific response of photosynthesis to burning and nitrogen fertilization. J Integr Plant Biol 50:565–574
Acknowledgments
We are grateful to T. Otoda and D. Kubo for their help in the field survey. This work was financially supported by the Global Environmental Fund of the Ministry of the Environment of Japan (D-1105) and by the Mitsui & Co. Environment Fund (R10-B196).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Jennifer Lee Firn, PhD.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Yoshihara, Y., Koyama, A., Undarmaa, J. et al. Prescribed burning experiments for restoration of degraded semiarid Mongolian steppe. Plant Ecol 216, 1649–1658 (2015). https://doi.org/10.1007/s11258-015-0548-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-015-0548-7