
Abstract
Competition and herbivory can interact to influence the recovery of plant communities from disturbance. Previous attention has focused on the roles of dominant plant species in structuring plant communities, leaving the roles of subordinate species often overlooked. In this study, we examined how manipulating the density of a subordinate plant species, Solanum carolinense, and its insect herbivores influenced an old-field plant community in northern Florida following a disturbance. Five years following the disturbance, the initial densities of S. carolinense planted at the start of the experiment negatively influenced total plant cover and species diversity, and the cover of some grasses (e.g., Paspalum urvillei) and forbs (e.g., Rubus trivalis). Selectively removing herbivores from S. carolinense increased S. carolinense abundance (both stem densities and cover), increased the total cover of plants in the surrounding plant community, and affected plant community composition. Some plant species increased (e.g., Digitaria ciliaris, Solidago altissima) and others decreased (e.g., Paspalum notatum, Cynodon dactylon) in cover in response to herbivore removal. Herbivore effects on plant community metrics did not depend on S. carolinense density (no significant herbivory by density interaction), suggesting that even at low densities, a reduction of S. carolinense herbivores can influence the rest of the plant community. The recovery of the plant community was context dependent, depending on site- and plot-level differences in underlying environmental conditions and pre-disturbance plant community composition. We demonstrate that the density of and herbivory on a single subordinate plant species can affect the structure of an entire plant community.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The recovery of plant communities following a disturbance is influenced by an array of biotic factors (Gleason 1927; Clements 1938; Connell and Slayter 1977; Diamond 1975). For example, competition for resources has long been considered an important biotic force structuring plant communities (Hairston et al. 1960). However, herbivores can also influence plant community structure through direct plant mortality and more importantly, indirectly by altering the competitive ability of plants (Crawley 1989; Louda et al. 1990; Huntly 1991). For instance, herbivore damage to a competitively dominant plant species can weaken its competitive effect on other species, potentially increasing plant diversity by allowing subordinate (non-dominant) species to grow and persist (Carson and Root 1999). Alternatively, damage to a subordinate species can exacerbate the negative effects of dominant competitors on the subordinate plant’s growth, potentially reducing overall diversity (Kim et al. 2013).
While the separate and combined effects of competition and herbivory have been repeatedly demonstrated to be important for structuring plant communities (reviewed by Chase et al. 2002; Hambäck and Beckerman 2003), it is unclear which members of a community can influence diversity patterns. Past studies often relate changes in plant diversity to changes in the competitive ability of the dominant plant species (Crawley and Pacala 1991; Carson and Root 2000) leaving the contribution of subordinate species overlooked (but see Bach 1994). This focus on dominant species is presumably because they have the largest direct effects on other members of the plant community (Armesto and Pickett 1985; Keddy 1990). However, in these studies, the densities of both the dominant and subordinate species typically vary simultaneously, making it difficult to assess the contribution of subordinate species to overall community structure (Connell 1983; Firbank and Watkinson 1990; Inouye 2001). Similarly, in herbivore removal experiments, plant communities are compared in the presence and absence of herbivores, and any differences in community composition are thought to arise from changes in the competitive ability of dominant plants. However, these studies generally use blanket removal techniques where herbivores are removed from all plant species with the use of fences, cages, or insecticides (Crawley 1989; Crawley and Pacala 1991). Because blanket removals affect herbivores on both dominant and subordinate species, these studies cannot rule out the possibility that subordinate species are also influencing plant communities. The contribution of subordinate species to community-level processes can be assessed by selectively manipulating their densities and associated herbivores while not manipulating other plant species, and following changes in plant community structure. By ignoring the role of subordinates, we are not getting a full picture of how herbivory and competition interact to influence plant communities.
In this study, we took advantage of an existing five-year experiment designed to examine the effects of herbivores on the demography of a native weed, Solanum carolinense (Underwood and Halpern 2012), to ask how competition by a non-dominant plant, S. carolinense, and herbivory on, S. carolinense, influences the surrounding old-field plant community (i.e., other non-S. carolinense members of the local plant community). To ensure the establishment of S. carolinense, this experiment was begun by disking plots and spraying with herbicide. This major disturbance to the pre-existing plant community was followed by planting of S. carolinense in plots at a range of densities. In half of these plots, S. carolinense was sprayed with insecticide to remove insect herbivores, while other plant species were allowed to establish on their own and grow without insecticide. By assessing the composition of the plant community after 5 years of ongoing herbivory treatments, we were able to ask the following questions: (1) Can the selective removal of herbivores on individuals of a subordinate plant species influence the surrounding plant community? (2) Does the initial density manipulation of a subordinate species after a disturbance influence the rest of the plant community? (3) Do herbivore removal and density manipulations interact? That is, does the absence of S. carolinense herbivores accentuate the effects of S. carolinense density on the surrounding members of the plant community? Or does selectively removing herbivores from S. carolinense only have effects in high-density plots where the insecticide intensity (and subsequent herbivore reduction) is highest?
Methods
Study system
Our study took place in abandoned agricultural fields (hereafter “old-fields”) at the North Florida Research and Education Center in Quincy, Florida (30.54, −84.59°). These sites are characterized by annual and perennial plant species including grasses (e.g., Digitaria ciliaris and Paspalum notatum), forbs (e.g., Sida spinosa and Commelina communis), and some legumes (e.g., Cassia obtusifolia). Prior to the study, fields were mowed twice per year. Our selective herbivore removal experiment focused on S. carolinense (Carolina horsenettle), a perennial herbaceous plant native to the eastern United States. Solanum carolinense reproduces both sexually and asexually, and is commonly found in old-fields throughout the region at densities of 0.3–8.1 individuals/m2 (T. Kim unpublished data). At our study location, recruitment occurs primarily through the growth of underground horizontal rhizomes rather than by seed (N. Underwood unpublished data) and before the start of our study, S. carolinense was rare in both fields and the experimental plots. Solanum carolinense is a relatively minor component of old-field vegetation (mean 5.3–11 % cover; T. Kim unpublished data) and therefore is well suited for understanding the effects of non-dominant plants on diversity patterns and the role of herbivores in mediating those effects. In this region, S. carolinense is fed on by many specialist (e.g., Manduca sexta, Leptinotarsa juncta, and Epitrix fuscula) and generalist (e.g., Spodoptera exigua, Melanoplus and Aptenopedes spp, Kim 2012) herbivores and damage can vary in intensity (16–62 % leaf damage; Kim et al. 2013). Solanum carolinense herbivory can influence both vegetative and sexual reproduction (Wise and Sacchi 1996), but it can also influence competition between S. carolinense and other old-field plant species (Kim et al. 2013) and is a function of the local neighborhood composition (Kim and Underwood 2015).
Study design
The 40 plots in this experiment were initially designed to examine density dependence in herbivore effects on the demography of S. carolinense (Underwood and Halpern 2012). All 40 experimental plots were established in March 2007 in two different old-fields (20 plots per field). The two fields (hereafter "sites") were separated by 750 m of rural old-field land cover. Plots were established with five initial planting densities of S. carolinense (0.65, 2.77, 11.11, 22.68, and 30.86 stems/m2 (as described in Underwood and Halpern 2012), allowing us to examine the effects of the initial planting densities of S. carolinense on plant community structure (i.e., the abundance, diversity, and composition of other species within the plot). Insect herbivores were selectively removed from S. carolinense in 20 of the plots by spraying all S. carolinense stems with 0.13 % carbaryl insecticide. This allows us to examine the effect of S. carolinense herbivory on plant community structure. To avoid confounding plot area and S. carolinense density, two different plot sizes were used for each density/spray treatment combination (plots varied in area from 1.6 to 98.4 m2). However, by logistical necessity, smaller plots were associated with higher initial densities (Appendix 1). For each plot area and density combination, an insecticide-sprayed and a control plot were paired and established next to each other (to reduce environmental heterogeneity) but pairs were randomly assigned to positions within a site. This design was balanced and the design was the same at each of the two sites.
Establishment and maintenance of treatments
Before planting, plots were disturbed by disking and then sprayed with the broad-spectrum herbicide Glyphosate (Roundup, The Scotts Company LLC, Marysville, OH, USA) to remove existing plant cover and minimize surviving seeds and roots. The few “wild” S. carolinense stems that re-emerged in experimental plots after the herbicide treatment were individually treated with herbicide again during the first growing season. Insects were removed from S. carolinense in half of the plots by spraying with Carbaryl (22 ml of Sevin Concentrate per 3.785 l, GardenTech, Lexington, KY, USA), a common insecticide targeting leaf chewing insects. Carbaryl has been shown to have little to no effect on plant growth (Simms and Rausher 1987; Murthy and Raghu 1990; Lau and Strauss 2005) and binds to soil (Gunasekara et al. 2008). Solanum carolinense in control plots were sprayed with an equal amount of water. Plants were sprayed at close range (5–10 cm from each S. carolinense stem) to minimize drift to neighboring plants (mean area sprayed per plant = 28.2 cm × 26.4 cm), although it is possible that plants immediately adjacent to S. carolinense received incidental spray. Spraying was performed every 2 weeks during the growing season (April–October) for 5 years (2007–2011).
Data collection
In August 2011, we surveyed plants using 50 cm diameter hoops. The number of hoops per plot was scaled with plot size (5–13 hoops placed at fixed sampling locations; small plots were near the maximum number of non-overlapping hoops that would fit in a plot, larger plots had more hoops, Pearson r = 0.95). Within each hoop, we estimated the percent cover of each plant species and bare ground (at intervals of 0–5, 6–20, 21–40, 61–80, 81–100 %), and counted the number of S. carolinense stems, our measure of density. After assessing the community within each hoop, we inspected the entire plot area and noted any rare species that were not captured in our hoops. Such rare species were assigned a cover of 0.2 %, a cover value slightly lower than the lowest occurring species recorded from our hoop measurements (0.23 %). Plant cover for each species was averaged across hoops within each plot.
We define the surrounding old-field plant community as the non-S. carolinense members of the plant community within a plot. We thus excluded S. carolinense cover from our calculations of species richness, diversity, and composition. Species richness was the number of plant species in each plot. Total cover was used as a surrogate for plant abundance, and was estimated as the sum of each species’ percent cover, excluding S. carolinense. Shannon-Weiner diversity (hereafter “diversity”) was calculated for each plot using the mean percent cover of each species. Differences in plant community composition among plots were characterized using Bray-Curtis dissimilarity, which was based on mean cover.
Data analysis
We used four separate generalized linear models (GLMs) to examine how S. carolinense cover, species richness, total cover, and diversity of the surrounding plant community responded to the insecticide treatment, initial planting densities of S. carolinense, and their interactions. We also included three covariates. First, we included “plot size” in the models to account for area effects as we sampled larger areas in larger plots. Second, we included “site” as a covariate to account for underlying differences in environmental conditions between the two fields, such as edaphic factors or past land-use practices. Lastly, we used an index of prior plant species composition at the scale of individual plots to account for the effects of spatial variation in pre-disturbance plant species composition and/or environmental conditions within each site. Plant composition within each plot prior to disturbance was estimated from high-resolution digital orthographic imagery acquired in March 2004, 3 years before the start of the experiment. There was no other experiment performed at our sites between 2004 and 2007, our initial planting date, thus our sites remained undisturbed during this period except for occasional mowing. Three spectral bands (green, blue, and infrared) were included in a principal components analysis (PCA) in order to generate a single-band image representing the spectral radiance across multiple ranges of the electromagnetic spectrum. Ordination analysis of multispectral imagery can be used in this way to characterize plant community composition in a continuous fashion, rather than using discrete classes (Schmidtlein et al. 2007). The mean of the first-axis PCA scores corresponding to each plot area was used as an index of prior plant composition (hereafter “prior plant cover,” Appendix 2). We evaluated all potential two- and three-way interactions between the insecticide and density treatments and the covariates. To simplify our final model, we dropped all three-way interactions if none were significant and re-ran GLMs with only two-way interactions. If none of the two-way interactions were significant, we ran GLMs with only main factors. With each round of model simplification, we tested whether data met GLM assumptions (e.g., residuals normally distributed, homogeneity of variance). When necessary, we log-transformed the data.
We used a permutational MANOVA (PERMANOVA; Bray-Curtis dissimilarity) to examine how spray treatment and initial S. carolinense planting density influenced plant community composition. To determine which plant species were most influenced by the spray treatment and S. carolinense planting density, we also performed separate GLMs on the mean cover of each species within the plant communities. Again, prior plant cover, site, and plot area were included as covariates and all two- and three-way interactions were evaluated using the same model simplification procedures as the GLMs. All analyses were performed using R v3.03 (R Development Core Team 2014) and the RVAideMemoire package (Hervé 2015) for PERMANOVA.
Results
At planting, in May 2007, S. carolinense was the only plant species in the plots. However, other species quickly established and by August 2007 mean S. carolinense cover across all plots was 56.4 % and non-S. carolinense cover was 36.5 % (Underwood and Halpern, unpublished data). One year later in August 2008, mean S. carolinense cover had declined to 15.7 % (Underwood and Halpern, unpublished data) and by the end of our five-year experiment (in August 2011), S. carolinense was only a minor component of the vegetation in the plots (mean cover per plot was 3.4 %, corresponding to a mean S. carolinense density of 1.82 stems/m2). A total of 45 plant species were found in the plots (mean = 10 species per plot) and 97 % of each plot was covered with vegetation. Fourteen species, mainly grasses and particularly P. notatum, made up 95 % of the plant cover.
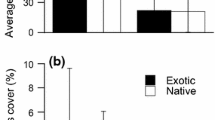
Underwood and Halpern (2012) found that insecticide reduced herbivore damage to S. carolinense in the first 4 years of the experiment (2007–2010). Not surprisingly, 5 years following disturbance, insecticide treatment positively influenced S. carolinense cover (GLM, main effect of insecticide, F 1,34 = 8.67, P < 0.01, Table 1) and the abundance rank of S. carolinense (7th in insecticide plots versus 13th in control plots, Fig. 1). The initial planting density was positively correlated with the mean densities of S. carolinense in subsequent years, including 2011 (t = 10.62, df = 38, P < 0.01, Spearman r = 0.86) and had positive effects on S. carolinense cover in 2011 (main effect of density F 1,34 = 7.71, P < 0.01, Table 1; Fig. 2).

The composition of the plant community in the absence (a) and presence (b) of herbivores on S. carolinense at the end of five-year experiment. At the start of the experiment, Solanum carolinense was the only plant species within the experimental plots. Five years later, 95 % of the cover was composed of a total of 14 species in insecticide-sprayed plots (a) and 13 species in control plots (b). Solanum carolinense (open bars) was ranked 7th overall in the absence of herbivores and 13th overall in the presence of herbivores at 4.5 % and 2.2 % cover, respectively. Error bars represent 1 ± SE

The effects of the initial densities of S. carolinense and insecticide treatment on the mean percent cover of Solanum carolinense (a), Paspalum urvillei (b), Verbena nastata (c), and Cynodon dactylon (d). Black circles and lines are plots sprayed with insecticide (“insecticide” plots); gray circles and dotted lines are plots sprayed with equivalent amounts of water (“control” plots). Densities are the number of S. carolinense ramets/m2. Lines are best fit lines from linear regression. Points represent partial residuals from linear regressions (Table 1, Appendix 2)
The plant community surrounding S. carolinense was influenced by the initial planting density of S. carolinense and insecticide treatment (Table 1). Five years following disturbance, the initial density of S. carolinense did not influence which species were present (i.e., community composition, PERMANOVA main effect F 1,24 = 1.18, P = 0.28) but increasing initial densities of S. carolinense reduced the total cover of plants (GLM, main effect F 1,22 = 6.55, P = 0.01) and decreased Shannon diversity (GLM, interaction with plot area F 1,24 = 4.72, P = 0.04). Reducing herbivory on S. carolinense sometimes influenced which species were present (i.e., community composition, PERMANOVA, interaction with site F 1,24 = 3.63, P < 0.01, interaction with prior plant cover F 1,24 = 2.96, P = 0.02) and increased the total cover of plants within the plots (GLM, main effect F 1,22 = 7.46, P = 0.01). Neither insecticide nor S. carolinense density influenced species richness (GLM, main effect of insecticide F 1,34 = 0.19, P = 0.66, main effect of density F 1,34 = 0.68, P = 0.23). Although species richness is generally a saturating function of plot area, we found that richness increased linearly with area (GLM, main effect F 1,34 = 79.25, P < 0.01). There were no effects of interactions between the initial densities of S. carolinense and herbivore reduction on any community metric.
The covers of different individual species of grasses and forbs, including both dominant and non-dominant species, were differentially affected by insecticide or the initial planting densities of S. carolinense (Figs. 2, 3; Appendix 3). For example, in the absence of S. carolinense herbivores, the cover of the most dominant grass P. notatum (mean cover 27.1 %, Fig. 3) was generally lower, but herbivore effects were context dependent (GLM, interaction with site F 1,24 = 5.29, P = 0.03; interaction with prior plant cover F 1,24 = 4.61, P = 0.04, Appendix 2). In the insecticide plots, most of the other species increased in cover, including D. ciliaris (GLM, main effect F 1,34 = 4.92, P = 0.03), Solidago altissima (GLM, interaction with site F 1,24 = 5.01, P = 0.03, interaction with prior plant cover F 1,24 = 7.16, P = 0.01) and Ipomea sp. (GLM, main effect F 1,34 = 7.91, P = 0.01). Manipulating the starting densities of S. carolinense influenced the cover of some species (e.g., Paspalum urvillei (positive effect), Verbena nastata (positive effect), Fig. 2; Appendix 3) and there was a density and insecticide interaction for Cynodon dactylon (positive effects of density in insecticide-sprayed plots only, 3-way interaction of density, insecticide and site).

Insecticide effects on the change in cover of the 10 plant species with the greatest change in cover and total cover of plants within each plot (change in mean % cover = mean % cover in insecticide-sprayed minus the mean % cover in control plots). Asterisks denote significant differences in % cover between insecticide and control plots (P < 0.05)
Underlying differences between sites and plot-level differences in pre-existing plant communities prior to the experiment influenced S. carolinense cover and community structure in 2011. Site had a significant effect on S. carolinense cover (GLM: main effect F 1,34 = 20.08 P < 0.01), total cover (GLM: interaction with insecticide F 1,22 = 4.01 P = 0.05), Shannon diversity (GLM: main effect F 1,24 = 7.99 P < 0.01), and community composition (PERMANOVA: main effect F1,24 = 4.56 P < 0.01). Prior plant cover influenced Shannon diversity (GLM: main effect F 1,24 = 7.47 P = 0.01), and community composition (PERMANOVA: main effect F 1,24 = 4.56 P < 0.01).
Discussion
The manipulation of a single, competitively subordinate plant species (S. carolinense) and its herbivores affected the surrounding plant community 5 years following a disturbance. The density manipulations affected the total cover of plants and plant diversity, while the insecticide treatment affected total plant cover and plant community composition. Richness, on the other hand, was not affected by either density or insecticide treatments. The covers of some individual species (both dominant and subordinate species) were also affected by both treatments (Figs. 2, 3; Appendix 3). Bach (1994) found similar results in sand dunes where the abundances of both dominant and subordinate species varied when herbivores were removed from one plant species (Salix cordata); however in that study richness and diversity did not change. In our study, the effects of S. carolinense herbivory reduction on other members of the plant community did not depend on the density of S. carolinense (no interaction of planting density and insecticide), so even a small reduction of S. carolinense herbivores can influence the rest of the community. If other subordinate plant species have similar effects on the rest of the plant community then selective manipulation experiments with different species could reveal a larger role for subordinate species in shaping plant communities.
Potential mechanisms of plant community response to the selective removal of herbivores
Previous studies have used blanket removal experiments to evaluate the effect of herbivores on plant community structure. However, blanket removal methods do not allow us to disentangle the mechanisms by which herbivores are affecting the plant community as herbivores feeding on both dominant and subordinate plant species are affected by the herbivore manipulation. Here, we manipulated herbivores feeding on a subordinate plant species so changes to the plant community likely arise directly or indirectly via the subordinate species, or herbivores using that species. We found that herbivore removal increased the total cover of plants, altered the cover of certain plant species, and influenced plant community composition. Reducing herbivory on S. carolinense could affect the plant community through at least two mechanisms. First, selectively spraying S. carolinense with insecticide may alter herbivore damage on other plant species. Targeted insecticide could result in mortality of generalist herbivores, reducing their ability to feed on all plants in the community. If this was the case, we would expect damage to be lower in the insecticide plots versus the control plots. On the other hand, generalist herbivores could avoid sprayed plants, and instead concentrate feeding on unsprayed plants. If this was the case, we would expect damage on neighboring plants to increase, potentially resulting in reduced cover. Our results show that total plant cover increased in plots where S. carolinense was sprayed, which is consistent with reduced overall herbivore damage. However, some species (including the dominant grass P. notatum) had decreased cover in insecticide plots, suggesting that there might be some level increased feeding on non-target plants and treated plant avoidance. Although direct effects of spraying on herbivory seem likely to contribute to cases where spraying increased cover, two lines of evidence suggest that reduction in overall damage in the community does not fully explain the effects of targeted spray on the surrounding plant community. First, our detailed observations of herbivory on two common neighboring forbs (Rubus trivalis and S. altissima), showed that spraying insecticide on S. carolinense did not influence damage to these two species (Appendix 4, GLM, R. trivalis F 1,13 = 0.001, P = 0.96, S. altissima F 1,13 = 0.308, P = 0.58). Second, most damage to S. carolinense is done by specialist herbivores on Solanaceae (Wise 2007; Underwood and Halpern 2012) rather than generalists, which suggests that another mechanism may be important.
A second mechanism for the effects of spraying on plant abundances is that releasing S. carolinense from herbivory could have changed its competitive effects on the rest of the community (e.g., Crawley 1989; Huntly 1991; Bach 1994). In the absence of herbivores, stem densities of S. carolinense were higher and overall plant size was greater (Underwood and Halpern 2012). Changes in plant size and densities could have allowed S. carolinense to compete strongly with some other plant species for resources and space. Indeed S. carolinense increased in competitive ranking (to the extent that competitive effects are reflected in percent cover) from 13th place in the presence of herbivores to 7th place in the absence of herbivores, outranking other subordinate species such as Ipomea, Carex, Hyptis mutabilis, and C. dactylon (Fig. 1). In a previous study in old-fields (Carson and Root 2000), a blanket removal of herbivores was performed and it was assumed that changes in the competitively dominant species, S. altissima, influenced plant community structure resulting in slower rates of succession and invasion by woody species. However, our results suggest that subordinate species were also likely influenced by the herbivore manipulation and could have contributed to changes in overall plant composition.
Potential mechanisms of plant community response to selectively adding S. carolinense
This experiment began with a major disturbance (disking and spraying herbicide) followed by the planting of S. carolinense. This gave S. carolinense an advantage over other plant species through preemptive competition because established S. carolinense could grow and use resources while other plant species were still in the process of re-establishment after the disturbance. Increasing the initial planting densities of S. carolinense could intensify the effect on the surrounding plant community by enhancing this advantage in establishment and thus reducing the overall cover of the surrounding plant community. There was also a negative relationship between the initial plantings of S. carolinense and diversity. It is unlikely that S. carolinense directly reduced diversity but S. carolinense could have reduced diversity indirectly by allowing other species to increase in dominance. Indeed there was a positive relationship between S. carolinense density and the cover of P. urvillei (grass) and R. trivalis (forb). It is unclear how P. urvillei and R. trivalis could benefit from increasing densities of S. carolinense but these effects could have affected plant species evenness and contributed to the overall change in diversity.
Plant community recovery was context dependent
Context dependencies can dominate many ecological processes (e.g., Foster et al. 2003; Cramer et al. 2008) and influence the recovery of plant communities following a disturbance. For example, we found that differences between sites were important factors influencing diversity and composition, and the cover of various plant species (Table 1). The experiment was conducted in two different fields that likely varied in abiotic factors (e.g., soil, topography) and land-use histories, which may have strongly influenced the recovery of the plant community following disturbance. We also observed strong effects of prior plant cover on current plant community structure. Although our experimental plots were tilled and treated with herbicide at the start of the experiment, the plant community at the end of the five-year experiment was not a result of primary succession as seeds and rhizomes survived the disturbance and this legacy of the pre-disturbance plant community influenced the contemporary plant community structure. Finally, we observed significant effects of interactions between herbivory or density and site or prior plant cover on plant community composition and the cover of various plant species (Table 1; Appendix 2), suggesting that the effects of herbivory and competition in structuring plant communities may be context dependent. Similar results have been found in other studies where herbivore effects were mediated by abiotic factors (e.g., salinity Gedan et al. 2009) or land-use history (Hahn and Orrock 2015) to influence the trajectory of secondary succession. This suggests that the plant communities might not respond to biotic interactions in predictable ways and context dependency should be considered.
Conclusions
Our study supports the idea that the density and herbivory of a subordinate species can influence plant communities. Even small changes in herbivore loads, possibly through the effects of a single subordinate plant species, can have significant effects on a plant community. Our results suggest that more attention to the effects of subordinate plant species is needed and might improve our understanding of plant community structure and dynamics.
References
Armesto JJ, Pickett STA (1985) Experiments on disturbance in old-field plant communities: impact on species richness and abundance. Ecology 66:230–240
Bach CE (1994) Effects of a specialist herbivore (Altica subplicata) on Salix cordata and sand dune succession. Ecol Monogr 64:423–445
Carson WP, Root RB (1999) Top-down effects of insect herbivores during early succession: influence on biomass and plant dominance. Oecologia 121:260–272
Carson WP, Root RB (2000) Herbivory and plant species coexistence: community regulation by an outbreaking phytophagous insect. Ecol Monogr 70:73–99
Chase JM, Abrams PA, Grover JP, Diehl S, Chesson P, Holt RD, Richards SA, Nisbet RM, Case TJ (2002) The interaction between predation and competition: a review and synthesis. Ecol Lett 5:302–315
Clements FE (1938) Nature and structure of the climax. J Ecol 24:252–282
Connell JH (1983) On the prevalence and relative importance of interspecific competition—evidence from field experiments. Am Nat 122:661–696
Connell JH, Slayter R (1977) Mechanisms of succession in natural communities and their role in community stability and organization. Am Nat 111:1119–1144
Cramer VA, Hobbs RJ, Standish RJ (2008) What’s new about old fields? Land abandonment and ecosystem assembly. Trends Ecol Evol 23:104–112
Crawley MJ (1989) Insect herbivores and plant-population dynamics. Annu Rev Entomol 34:531–564
Crawley MJ, Pacala SW (1991) Herbivores, plant parasites, and plant diversity. In: Toft CA, Aeschlimann A, Bolis L (eds) Parasite-host associations: coexistence or conflict?. Oxford University Press, Oxford, pp 157–173
Diamond JM (1975) Assembly of species communities. In: Cody ML, Diamond JM (eds) Ecology and evolution of communities. Harvard University Press, Cambridge, pp 342–444
Firbank LG, Watkinson AR (1990) On the effects of competition: from monocultures to mixtures. In: Grace JB, Tilman D (eds) Perspectives on plant competition. The Blackburn Press, Caldwell, pp 165–192
Foster DF, Swanson F, Aber J, Burke I, Brokaw N, Tilman D, Knapp A (2003) The importance of land-use legacies to ecology and conservation. Bioscience 53:77–88
Gedan KB, Crain CM, Bertness MD (2009) Small-mammal herbivore control of secondary succession in New England tidal marshes. Ecology 90:430–440
Gleason HA (1927) Further views on the succession concept. Ecology 8:299–326
Gunasekara AS, Rubin AL, Goh K, Spurlock F, Tjeerdema RS (2008) Environmental fate and toxicology of Carbaryl. Rev Environ Contam Toxicol 196:95–121
Hahn PG, Orrock JL (2015) Land-use legacies and present fire regimes interact to mediate herbivory by altering the neighboring plant community. Oikos 124:497–506
Hairston NG, Smith FE, Slobodkin LB (1960) Community structure, population control and competition. Am Nat 94:421–425
Hambäck PA, Beckerman AP (2003) Herbivory and plant resource competition: a review of two interacting interactions. Oikos 101:26–37
Hervé M (2015) RVAideMemoire: diverse basic statistical and graphical functions package. Version 0.9-45-2. http://cran.r-project.org/web/packages/RVAideMemoire/index.html
Huntly N (1991) Herbivores and the dynamics of communities and ecosystems. Annu Rev Ecol Syst 22:477–503
Inouye BD (2001) Response surface experimental designs for investigating interspecific competition. Ecology 82:2696–2706
Jin S, Yang L, Danielson P, Homer C, Fry J, Xian G (2013) A comprehensive change detection method for updating the National Land Cover Database to circa 2011. Remote Sens Environ 132:159–175
Keddy PA (1990) Competitive hierarchies and centrifugal organization in plant communities. In: Grace JB, Tilman D (eds) Perspectives on Plant Competition. The Blackburn Press, Caldwell, pp 265–290
Kim TN (2012) Community-level consequences of plant–herbivore interactions. PhD Dissertation, Florida State University
Kim TN, Underwood N (2015) A full picture of plant neighborhood effects on herbivory: damage is both density and frequency dependent. Ecology 96:1431–1437
Kim TN, Underwood N, Inouye BD (2013) Insect herbivores change the outcome of plant competition through both inter- and intraspecific processes. Ecology 94:1753–1763
Lau JA, Strauss SY (2005) Insect herbivores drive important indirect effects of exotic plants on native communities. Ecology 86:2900–2997
Louda SM, Keeler KH, Holt RD (1990) Herbivore influences on plant performance and competitive interactions. In: Grace JB, Tilman D (eds) Perspectives on plant competition. The Blackburn Press, Caldwell, pp 413–444
Murthy N, Raghu K (1990) Effect of carbaryl and 1-naphthol on seedling growth of barley, and on growth and nodulation of groundnut in two soils. Proc Plant Sci 100:11–15
R Development Core Team (2014) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Schmidtlein S, Zimmermann P, Schüpferling R, Weiß C (2007) Mapping the floristic continuum: ordination space position estimated from imaging spectroscopy. J Veg Sci 18:131–140
Simms EL, Rausher MD (1987) Costs and benefits of plant-resistance to herbivory. Am Nat 130:570–581
Underwood N, Halpern S (2012) Insect herbivores, density dependence, and the performance of the perennial herb Solanum carolinense. Ecology 93:1026–1035
Wise MJ (2007) The herbivores of Solanum carolinense (Horsenettle) in Northern Virginia: natural history and damage assessment. Southeast Nat 6:505–522
Wise MJ, Sacchi CF (1996) Impact of two specialist insect herbivores on reproduction of horse nettle, Solanum carolinense. Oecologia 108:328–337
Acknowledgments
We thank J. Simonis, J. Fort, C. Venner, J. Hines, and numerous REU students for helping to establish and maintain the project over the 5 years. We thank the staff at the University of Florida North Florida Research and Education Center for their logistical support. Comments from Joshua Grinath greatly improved this manuscript. This project was funded by NSF DEB-0717221 to N. Underwood, and NSF DEB-0716922 and NRI, CSREES, USDA Grant 2006-35320-16686 to S. Halpern.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Lauchlan Frase.
Appendices
Appendix 1
See Fig. 4.

For logistical reasons, plot size varied with density (Spearman r s = −0.948) but to avoid completely confounding density with plot area, we used two plot sizes for each density treatment. Although this does present the possibility of spurious correlations with density and plot size, there was enough variation between the two variables to decouple their effects on plant responses. However, to ensure that spurious correlations were not present, we re-ran GLMs (see “Data analysis” in Methods), replacing the significant variable (e.g., density) with the other correlated variable (e.g., plot size) and compared model results and AICc values. If the replacement did not yield the same results and the other variable did not appear significant, we conclude that it was not a spurious correlation and that the effects are real. In all cases, the density and area effects represented in the paper were real
Appendix 2
Index of prior plant composition.
Plant composition prior to disturbance was estimated from high-resolution digital orthographic quarter quad imagery acquired in March 2004, 3 years before the start of the experiment. Different plant species, but also variation in soil and vegetation water content, can reflect light of different intensities across a range of the electromagnetic (EM) spectrum, which forms a basis for all land cover classifications (e.g., Jin et al. 2013). Accordingly, we assume that the digital number values corresponding to each 1 m2 pixel represents the average spectral radiance of the individual plants within that area, and therefore the composition of plant species. Moreover, we assume underlying environmental variation potentially driving differences in prior plant cover (e.g., soil type or hydrological properties) are incorporated within radiance values. We included the three bands from the image (green, blue, and infrared) in a principal components analysis (PCA) in order to generate an image that represents spectral radiance from multiple ranges of the EM spectrum on a single band. Ordination techniques such as this can be used to characterize vegetation communities in a continuous fashion rather than forming discrete habitat classes (Schmidtlein et al. 2007). We overlaid the plot boundaries on the PCA image product and, for each plot, calculated the mean of the first-axis PCA scores (Fig. 5). The first axis explained 71.6 % of the variation in the original three-band image. The mean for each plot was used in subsequent analyses as an index of prior plant cover.
See Fig. 5.

PCA images illustrating differences in prior plant cover among plots in the west (a) and east (b) sites. Plot positions are outlined in white and ranged in sizes from 1.6 to 98.4 m2. The five initial planting densities of S. carolinense within each plot were 0.65, 2.77, 11.11, 22.68, and 30.86 stems/m2. This figure depicts all three axes as a false color composite with PCA axis 1 (71.6 %) coded as red, PCA axis 2 (25.7 %) coded as blue, and PCA axis 3 (2.7 %) coded as green, but only the mean of the first-axis scores corresponding to areas within each plot was used as a covariate in our analyses “prior plant cover”
Appendix 3
See Table 2.
Appendix 4
See Table 3.
Rights and permissions
About this article

Cite this article
Kim, T.N., Spiesman, B.J., Buchanan, A.L. et al. Selective manipulation of a non-dominant plant and its herbivores affects an old-field plant community. Plant Ecol 216, 1029–1045 (2015). https://doi.org/10.1007/s11258-015-0487-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-015-0487-3