
Abstract
In an increasingly urbanized world, some raptors successfully colonize and thrive in urban environments, leading to more frequent interactions with humans. These interactions can be either positive, such as providing ecosystem services, or negative, resulting in human-wildlife conflicts. Despite growing literature on these interactions, a comprehensive review focusing on urban environments has been lacking. This study aimed to address this gap by conducting a systematic review using Google Scholar and the Scopus bibliographic database. A total of 45 studies met the search criteria, with a predominant prevalence of the northern hemisphere. Accipitriformes was the most studied order of raptors (50%), followed by Strigiformes (37%), Falconiformes (8%), and Cathartiformes (2%). Positive interactions studied included cultural services, pest control, positive perception, carrion removal, while negative interactions involved safety damage, property damage, negative perception, disease transmission, livestock damage, nuisance and superstitions. Pest control and cultural services were supported by the evidence, although only for specific orders. Carrion removal and aggressiveness appear to decrease with urbanization, although more studies are needed to verify this premise. Both positive and negative perceptions were evident, influenced in part by the knowledge or closeness that people had towards urban raptors. We discuss how the interactions studied influence the daily lives of citizens and, in turn, how human activities shape and influence these interactions. Finally, given that cities are socio-ecological systems, we advocate for methodologies that integrate the social aspects of human-predator interactions along with ecological ones to promote coexistence.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In the modern world, the rate at which human populations and land cover are becoming urban is faster than at any other time in history (Seto et al. 2010). Indeed, urban land cover is expected to increase between 430.000 km2 and 12.568.000 km2 (Seto et al. 2011) and global population is expected to increase to 8.5 billion (from 7.7 billion in 2019) (United Nations Development Programme 2022) by 2030. Although the expansion of urban lands on rural and natural lands generally has negative consequences for biodiversity (Piano et al. 2020), some species, such as birds, are able to colonize, persist or even thrive in urban environments (Isaksson 2018). In fact, some urban areas have greater abundance of birds (or biomass) per sampling unit than nonurban habitats (Marzluff et al. 2001; Chace and Walsh 2006; van Rensburg et al. 2009; MacGregor-Fors et al. 2012; Drewitt et al. 2021). Particularly, raptor exemplify this phenomenon, with some species becoming increasingly common in urban areas due to their ability to exploit nesting and feeding opportunities available in cities (Bildstein and Therrien 2018; Mak et al. 2021) (e.g., Great Horned Owl Bubo virginianus, Peregrine Falcon Falco peregrinus, Black Sparrowhawk Accipiter melanoleucus and Black Kite Milvus migrans) (Chace and Walsh 2006; Suri et al. 2017; Mazumdar et al. 2017; Kettel et al. 2018). While cities can be ecological traps for some raptor species (Sumasgutner et al. 2014; Kettel et al. 2018), they also act as population sources for other species, such as Peregrines in the United Kingdom, which may help recolonize vacant territories (Drewitt et al. 2021). Furthermore, as previously observed for threatened species (Ives et al. 2016), cities can serve as conservation hotspots for raptors like Burrowing Owls (Athene cunicularia) (Rebolo-Ifrán et al. 2017). These species are now faring better in moderately urbanized areas than in their increasingly threatened natural habitats (Rebolo-Ifrán et al. 2017; Franco and Marçal-Junior 2018; Baladrón et al. 2020; Cavalli et al. 2023).
The increase in raptor populations in cities, coupled with the expansion of peri-urban environments (transition zones between urban and rural areas that typically exhibit relative higher levels of species diversity and abundance) (McKinney 2002; Seto et al. 2010) heightens the likelihood of human-raptor interactions (Conover 2001, p.33; Ditchkoff et al. 2006). These interactions can range from positive outcomes for humans, such as ecosystem services, to negative ones, like human-wildlife conflicts. Ecosystem services are defined as the benefits people obtain from ecosystems and the species that compose those systems (Millennium Ecosystem Assessment 2005). Ecosystem services that raptors provide might include, scavenging by vultures and other raptors, biological control of pest species (regulating services) and economic and social benefits from bird-watching and ecotourism (cultural services or nature’s material contribution to people) (Díaz et al. 2018). On the other hand, negative interactions, usually termed human–wildlife conflict, emphasize the antagonism relationships between wildlife and humans (Conover 2001, p.8; Graham et al. 2005). In this review, we adopt the definition of human-wildlife conflicts provided by Soulsbury and White (2015), which describes these conflicts as situations “… caused where the movement and activities of wildlife, such as those associated with foraging or reproduction, have an adverse impact on human interests…”. Such impacts may include aggression, property damage (both livestock and non-agricultural property), disease transmission and nuisance (Peterson et al. 2010; Soulsbury and White 2015).
While global scientific information on human-raptor interactions is increasing, current reviews primarily focus on how these interactions benefit or affect raptors at an individual or population level, such as survival or reproductive success. Most reviews detail interactions occurring in natural or rural settings (Nyhus 2016; McClure et al. 2018; Canney et al. 2022; Ballejo et al. 2022) or concentrate on specific raptor groups (Carucci et al. 2022). Although these studies are valuable and enhance our understanding of human-raptor dynamics, it is crucial to thoroughly examine the progress made in researching interactions that directly and indirectly affect humans, which we focus in this study. Raptors could improve human well-being by connecting people with nature experiences (i.e. green prescribing)(Thomas et al. 2022) and act as flagship species, but they are also more threatened than birds in general (Donázar et al. 2016; Cox et al. 2017; McClure et al. 2018). Since conservation interventions are shaped by human decisions (Mascia et al. 2003), exploring human-raptor interactions, as proposed in this review, is vital for developing more effective conservation strategies and fostering coexistence between humans and raptors. Accordingly, our objective is to review empirical studies of raptor ecosystem services and conflicts, as defined above, with human in urban environments. We quantify and synthesize existing information, identify areas of progress and pinpoint information gaps and biases. Finally, we provide recommendations for future research and management based on our scientific understanding of current human-raptor interactions.
Methods
We conducted a bibliographic search in English in Google Scholar and Scopus to find peer-reviewed articles on positive and negative human-raptor interactions in urban environments around the world until 1st of June 2023, without year restriction. We define raptor as species within orders Cathartiformes, Accipitriformes, Strigiformes and Falconiformes (Boal and Dykstra 2018). We filtered our search to studies conducted in urbanized environments, which included: cities, suburban or peri-urban areas and villages. Studies comparing natural and rural environments with urban ones were also included in the review. We focus only in those articles that studied natural or spontaneous interactions between raptors and humans; therefore, those that studied the pest control services using falconry were excluded from the analysis. To avoid repetition of information already published in the scientific literature, we excluded gray literature (theses and dissertations), books, book chapters and scientific reviews. Additionally, we discarded duplicate studies. Since we are interested in the human dimensions from interactions, we excluded studies that did not address the human aspect, specifically those did not involve raptors impacting human interests or providing ecosystem services. For example, we excluded studies that solely evaluated conservation issues such as deaths from electrocutions, pollution, intentional killings, agrochemicals, collisions with windows, or those using raptors as indicators of environmental health or as flagship species. An initial search using the keywords “urban raptor” in both digital academic search engines was conducted to cover a wide range of interactions between human and raptors. Then, we performed a second specific search using the following combination of terms: “urban” OR “city” AND “raptor” OR “bird of prey” OR “predatory birds” AND “conflict” OR “benefit” OR “ecosystem services” OR “human-raptor interaction” OR “diet” OR “control pest” OR “disease transmission” OR “aggression” OR “attack” OR “scavenger” OR “cultural service” OR “garbage”. We followed the protocol established for PRISMA Statement (Page et al. 2021) to select suitable records. We also checked the bibliographies of the articles identified in the database search for additional relevant publications that may not have appeared in Google Scholar or the Scopus bibliographic database. For more details on the methodological choices made in the selection of studies for this review, see Supplementary Materials (Fig. S1).
From each selected scientific article, we extracted the year of publication and the country of the study, the species studied, the methodology used (hereafter referred to as “Methodological approach”), the type of interaction evaluated (positive, negative or both) and whether the authors proposed any management measures to mitigate human-predator conflicts. We then classified the articles based on the type of human-raptor interaction studied and the methodology approach used (Table 1). The categories were not mutually exclusive, as a study could use more than one methodology to examine the same interaction or study multiple interactions, both positive and negative. Finally, for each species studied we extracted the conservation status from the IUCN Red List (IUCN 2024). The data were analyzed using R statistical environment (R Core Team 2023) and in SCImago Graphica (Hassan-Montero et al. 2022).
Results
Year of publication and geographical areas studied
We identified 45 studies that met our selection criteria. The earliest study was published in 1999, and there has been an increasing trend in the number of studies on this topic over time, with the highest publication rate occurring in 2018 (n = 7; Fig. 1). Scientific publications from all continents were found, but the majority were from Asia (27%; n = 12) and North America (24%; n = 11), followed by Europe and South America (each one 16%; n = 7), Africa (11%; n = 5) and Oceania (7%; n = 3; Fig. 2). In the geographical analysis of the Proportion Z-Test at the continental level (Table SM2), the following significant differences in the proportion of studies on positive interactions were evident: Europe (z = 2.11, p = 0.04), South America (z = 2.63, p = 0.01) and Oceania (z = 2.25, p = 0.02) conducted more studies on positive interactions than Asia. South America also had a higher proportion of positive studies than Africa (z = 2.16, p = 0.03). Finally, a marginal difference was observed between Oceania and Africa, likely influenced by the low sample size, in favor of Oceania (z = 1.89, p = 0.05). Regarding the proportion of negative studies, North America presented a significantly greater proportion compared to Asia (z = 2.81, p = 0.01), Africa (z = 2.25, p = 0.02) and Oceania (z = 1.94, p = 0.01).
The countries with the most registered species were slightly positively correlated with the number of published articles (Spearman test, r2 = 0.53, p = 0.02). The United States had the highest number of publications (24%; n = 11), followed by India (11%; n = 5). Spain was the country with the most species studied (43%; n = 28), followed by United States (31%; n = 20) and India (8%; n = 5). Additional data of interactions according to country are presented in Supplementary Materials, Table SM1.

Number of published studies of positive and negative human-raptor interactions in urban environments over the years

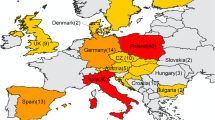
Map of the countries were the studies where performed. Bright red shows high number of articles found for a given country, ranging from 1 (dark red) to 11 (bright red). The black numbers within each country indicate the number of studies published in them
Studied taxa
Some articles investigated the same type of interaction across multiple species, families, or orders simultaneously. Consequently, the sum of studies conducted for each order or family exceeds the total number of studies reviewed (n = 45). For instance, the most studied Order was Accipitriformes (50%; n = 60), followed by Strigiformes (37%; n = 44), Falconiformes (8%; n = 10) and the least studied Cathartiformes (2%; n = 2). Adjusting for the total number of species per order reveals that Cathartiformes (29%) emerges as the most studied, followed by Accipitriformes (23%), Strigiformes (18%) and Falconiformes (15%). The total number of species considered for each order was 7 for Cathartiformes, 252 for Accipitriformes, 247 for Strigiformes and 65 for Falconiformes (Billerman et al. 2022). Three studies were not classified by Order because the identification by the authors was labeled as “Raptor or/and vulture”. The most studied families were Accipitridae (48%; n = 56) followed by Strigidae (29%; n = 34), Falconidae and Tytonidae (each one 8%; n = 10), Pandionidae (3%; n = 3) and Cathartidae (2%; n = 2). Again, in four articles it was not possible identify the family for the reason exposed above or because the authors identified the species like “Accipitriformes spp.”.
The studies record 65 species (Table 2). The most frequently studied species were the Barn Owl (Tyto alba) (8%; n = 10), followed by the Black Kite (M. migrans) and the Peregrin Falcon (F. peregrinus) (each one 3%; n = 4). Of these species, 15% (n = 8) are considered species of conservation concern by the IUCN (Table 2). Only one species (Strix mauritanica) was not evaluated for the IUCN. The rest of the species were categorized as Last concern (86%; n = 56).
Methodological approach
The identified studies employed a variety of methodological approaches to examine both positive and negative human-raptor interactions in urban environments. Interviews were the most common method, used in 26% of the articles (n = 12), followed by pellet analysis and surveys (each constituting 19%; n = 9). Field experiments and database analyses accounted for 10% each (n = 5), microbiological tests for 6% (n = 3) and camera traps and citizen science each made up 4% (n = 2). Methodologies approaches used were not mutually exclusive, as we found studies with multiple methodologies for evaluating the same or different interactions.
Human-raptors interactions
As some studies selected for this review studied more than one type of interaction, a total of 62 human-raptor interactions were found, from which 56% (n = 35) were categorized as positive and 44% (n = 27) as negative. Only 9 studies investigated positive and negative interactions at the same time. The most studied negative interaction or conflict was safety damage (11%; n = 7; Fig. 3A), while the most studied positive interaction or benefit was cultural service and pest control (each one 10%; n = 16; Fig. 3B).

Number of total published studies per each negative (A) and positive (B) interactions
Of the studies assessing positive interactions (n = 28), 79% (n = 22) confirmed their occurrence, while the remaining 21% (n = 6) found no evidence. In the case of studies examining negative interactions (n = 26), 77% (n = 20) verified the existence of conflicts, whereas the other 23% (n = 6) did not. Among the studies that confirmed the presence of negative conflicts, only six proposed management strategies to address or mitigate them.
All raptors orders had a greater number of published studies on negative interaction than on positive interaction (Fig. 4), but they stand out in different categories. For example, Strigiformes had the most studies on property damage and pest control (Fig. 4D). Accipitriformes were most studied as disease vectors and carrion removal (Fig. 4C) and were the only order studied with regards threats to human safety. The latter were only absent in pest control studies (Fig. 4C). Cathartiformes studies focused solely on disease spread and property damage (Fig. 4A). Finally, Falconiformes did not stand out in any category and were absent in studies of property damage, safety and carrion removal (Fig. 4B). Additional data of interactions discriminated by species are given in Supplementary Materials, Table S1.

Number of studies published on negative and positive interactions (highlighted in red) and their respective subcategories for each group of raptors: Cathartiformes (A), Falconiformes (B), Accipitriformes (C) and Strigiformes (D)
Discussion
Discussion section is structured as follows: First, we provide a brief overview of the current state of knowledge regarding conflicts and services of raptors in urban environments, including a discussion on geographic and taxonomic biases. We then divide the discussion into several sections. The first two sections, ‘Human-Scavenger Interactions’ and ‘Human-Predator Interactions,’ cover the generalities evidenced by the services and conflicts provided by each order of raptors. Following this, the discussion continues with the interactions defined in Table 1. To conclude, the reader will find a discussion on the management measures used or recommended in the studies.
Our global analysis revealed that raptor groups interacted differently with humans in urban environments, with certain orders dominating in specific conflicts and services. Our review also showed that raptors are underrepresented in studies on wildlife services and conflicts in urban environments, as other studies have shown (Soulsbury and White 2015; Nyhus 2016; Basak et al. 2023). The majority of species involved in services or conflicts are not classified as conservation concern, indicating an absence of study bias towards endangered species. This is unsurprising, as the traits that allow raptors to thrive in urban environments (see below in the “Human-Scavenger Interactions” section) also make them less vulnerable. Additionally, these non-threatened species are more common in cities, leading to more frequent interactions with people (Gaston 2010).
Robust geographic comparisons remain challenging due to the limited number of countries with more than three publications. Nevertheless, we conducted an exploratory analysis at the continental level, revealing a clear pattern: North America stands out for negative interactions, whereas South America and Oceania are more prominent for positive interactions. It is not surprising that North America and Oceania highlighted, as birds are frequently investigated in urban ecology studies in these regions (Magle et al. 2012). In contrast, given the increasing human-wildlife conflicts caused by expanding human populations and urban areas in the countryside of Africa and Asia, it would be logical to expect a greater proportion of studies on negative interactions between humans and raptors in these continents. However, this result was not evident. Rather than indicating a lack of conflicts, this demonstrates, as previously shown by Magle et al. (2012), the concerning lack of urban ecology studies in these regions. These results reveal a geographic bias toward the northern hemisphere, consistent with observations in studies of urban ecology (Luederitz et al. 2015; Collins et al. 2021) and raptors (Kettel et al. 2018; Boal and Dykstra 2018; Canney et al. 2022). This bias also reflects social inequalities in science, as countries in the Global South face funding restrictions and limited scholarships for education or publication (Magle et al. 2012; McClure et al. 2022). Additionally, scientists from some developed countries are nowadays forced to flee due to war, political repression and the climate crisis (Machlis and Carrero-Martinez 2024). Certainly, international funding efforts for research, publication and education should be a priority in these areas. However, our selection of English-speaking studies may have influenced our results. In the future, as knowledge accumulates across countries, it would be interesting to investigate whether conflicts or services provide by raptors are influenced by specific country characteristics such as level of development, culture, religion or geographical location.
Across the studies included in our analysis, the predominant focus was on positive interactions rather than negative ones. However, upon closer examination within each raptor order, an opposing trend emerged: negative interactions outnumbered positive ones (see Fig. 4). This discrepancy can be attributed to the inclusion of diverse species in conflict studies, such as those associated with disease transmission or property damage, which increased the prevalence of negative interactions in orders like Accipitriformes and Strigiformes.
Human-scavenger interactions
Among the raptors that consume carrion, including Accipitriformes, Cathartiformes, and Falconiformes, research on carrion removal services has primarily focused on facultative scavengers within the Accipitriformes (Table SM1). The relative lack of studies on vulture carrion removal in urban ecosystems compared to rural ones (DeVault et al. 2003; Grilli et al. 2019) may stem from their lower populations due to the high sensitivity of obligate scavengers to intense human impact, prevalent in the urban studies reviewed, although they benefit from moderate human impact as seen in some African countries (Buechley et al. 2018; Sebastián-González et al. 2019; McPherson et al. 2021a). These obligate scavengers, characterized by their large bodies and wingspans (DeVault et al. 2003), do not possess the traits typically associated with “archetypal urban raptors”, such as being small and generalist, traits beneficial to thrive in urban environments (Cooper et al. 2021; Headland et al. 2023).
While this hypothesis may explain the scarcity of studies on carrion removal by vultures, it does not account for the similar lack of research on Falconiformes. It is particularly surprising given that this order includes the clade Caracaras, most of which are facultative scavengers, Some species have high local urban abundances (Ferguson-Lees and Christie 2001; Carrete et al. 2009; Morrison and Saggese 2024) and are known to exploit carcasses resulting from fauna-vehicle collisions (Biondi et al. 2005; Lambertucci et al. 2009) or livestock carcasses (Travaini et al. 2001; Autilio et al. 2019) (e.g. Milvago chimango, Caracara plancus, Phalcoboenus australis). Like Cathartiformes, Caracaras are primarily found in South America, which suggests that the geographic bias towards the Northern Hemisphere in this review could be contributing to the underrepresentation of carrion removal studies in these orders (Morrison and Saggese 2024). The high level of study on Accipitriformes in this service, as well as in other interactions throughout our review, may be attributed not only to biological or ecological factors but also to their cosmopolitan distribution and abundance as the most prevalent order among raptors (Winkler et al. 2020a). Clearly, future research is needed to assess the ability of both obligate and facultative scavengers to provide ecosystem services in urban environments and to determine their contribution to human communities.
Human-predators interactions
Pest control facilitated by raptors stems from their foraging behavior reducing the need for costly management practices (e.g. pesticide) (Wenny et al. 2011). In our review of urban environments, Strigiformes emerge prominently in pest control studies, an expected outcome given their primarily carnivorous diet focused on small rodents (Winkler et al. 2020b, c). This role is well-established in agroecosystems (Sekercioglu et al. 2016; Mariyappan et al. 2023) and aligns with findings from Luna et al. (2021), which highlight that the most commonly observed carnivores in urban areas are those with nocturnal activity, coinciding with periods of reduced human activity (but see Gaston 2019 for an exception). This could explain the prevalence of studies focused on Strigiformes. However, there is a notable disparity when it comes to Accipitriformes, which, despite being among the most studied orders of urban predators alongside Carnivores (Luna et al. 2021), have seldom been researched for their role in pest control. This gap suggests that dietary studies of Accipitriformes in urban settings may not have explicitly targeted or tested their pest control capabilities, leaving a significant void in our understanding of their contribution to ecosystem services in these environments. A similar oversight exists for Falconiformes, even though they include urban-adapted species such as the Peregrine Falcon (Mak et al. 2021), which specializes in hunting birds. Despite this adaptation, only a limited number of studies explore their role in pest control. This indicates that natural pest control is presumed for these groups, but it has not been thoroughly studied. Lastly, it is unsurprising that there are no pest control studies involving Old World vultures (Accipitriformes) and New World vultures (Cathartiformes) given their specialized carrion-based diet (Winkler et al. 2020a, d).
Positive interactions
Carrion removal
Despite the positive impact that scavengers have on the environment, it is a topic that has been studied relatively little on a global scale (DeVault et al. 2003), particularly in urban environments (Luna et al. 2021). Our review underscores this information gap, as carrion removal is the least studied ecosystem service among all. However, align with global studies in scavenger communities (Sebastián-González et al. 2019), a pattern became evident during our review: carrion removal by raptors tends to decrease in urban areas compared to rural or pristine environments. This pattern may initially be observed because, with one exception, the studies were conducted in developed countries where waste management would be more effective (Mmereki et al. 2016; World Bank 2018). As a result, less carrion is available in these urban areas, limiting the activity of scavengers (Luna et al. 2021). In contrast, the only study that reported efficient carrion removal was conducted in a developing country, where such management practices may be less stringent (see below). Additionally, at the local level, the introduction of exotic species (Welti et al. 2020), infrastructure development and human presence (Huijbers et al. 2013, 2015; Thomson et al. 2016; Shizukuda and Saito 2023) were identified as the main factors affecting the regulatory service provided by raptors. The most alarming results were observed in Australia, where, across a gradient of urbanization, the ecosystem function of carrion removal by raptors was lost, and no other group replaced them in their absence (Huijbers et al. 2013, 2015; Thomson et al. 2016). An efficient case of carrion removal was observed in Yemen, where a high population of the Egyptian vulture (Neophron percnopterus) is supported due to the food sources, and as a consequence, they clean up 22% of the total putrescible waste matter in villages and rural areas (Gangoso et al. 2013). Considering that Egyptian vultures are globally listed as threatened (BirdLife International 2021), these findings underscore the importance of urban areas as potential conservation zones (McKinney 2002; Ives et al. 2016; Luna et al. 2021; McPherson et al. 2021a; Boakes et al. 2023). Not only do urban areas generate significant waste, but so do sites where animals are slaughtered for human consumption. This waste serves as a food subsidy for endangered species such as vultures (Buechley et al. 2022) or Black Kites (M. migrans) during their migration (Kumar 2023). Despite the critical role of human waste in supporting endangered or migratory scavengers, its presence undoubtedly enters in conflict with goals related to human health and urban development. Therefore, efforts aimed at managing and preventing zoonotic diseases in these areas will inevitably impact ecological functions (Plaza and Lambertucci 2017). ‘Vulture restaurants’ (i.e. safe feeding zones in areas inaccessible to humans) (Gilbert et al. 2007) could serve as a conservation strategy that simultaneously supports biodiversity conservation and protects human health (Adams et al. 2004).
As the decline of scavenger populations can increase human exposure to decomposing carcasses, raising the risk of pests and the spread of infectious diseases, further research is essential to understand the factors that influence the efficiency of this service and to develop effective management strategies, particularly for urban ecosystems (DeVault et al. 2016; Markandya et al. 2008; Buechley and Şekercioğlu 2016).
Pest control
While studies on pest control by birds of prey are typically conducted in agroecosystems (Sekercioglu et al. 2016; Mariyappan et al. 2023), this review reveals a diverse range of pest control activities performed by raptors in urban environments. The most frequently reported is the control of rodents (Saufi et al. 2020; Cherkaoui et al. 2021), particularly those acting as reservoirs for and transmitters of the hantavirus (Magrini and Facure 2008; Teta et al. 2012; Godoy-Guinao et al. 2017). Additionally, control of invasive alien species (Mori et al. 2020), conflictive prairie dogs (Cynomys ludovicianus) (Witmer et al. 2008), pigeons (Columbiformes) (Schneider et al. 2023) and fruit pests were also documented (Kopij and Liven-Schulman 2012). One study aimed to test the hypothesis that the high abundance of pest species was attributed to a lower abundance of raptor species; however, it found no evidence to support this hypothesis (Sorace 2002). Although all of the studies reviewed are important and contribute to the knowledge of predatory species in urban environments (Luna et al. 2021), they often do not assess whether raptor foraging behavior effectively reduces pest populations or their harmful effects. Demonstrating that raptors consume pest species is insufficient to assume a decrease in damage caused by pests. For instance, raptors may not consume pests in sufficient quantities to significantly reduce damage, or pests may alter their foraging behavior rather than decline in population due to the presence of raptors (i.e. “landscape of fear” effect) (Whelan et al. 2008; Wenny et al. 2011; Bleicher 2017). Although methodologically challenging, detailed quantification of this regulatory service would be desirable to maximize the benefits of natural pest control by raptors in urban environments (Whelan et al. 2008; Wenny et al. 2011; Donázar et al. 2016).
Cultural service
This review reveals that the cultural service of raptors was one of the most studied topics within urban environments, which may be due to the ease of collecting this type of information (i.e. through interviews). The studies reviewed demonstrate various ways in which birds of prey are integrated into urban culture, including religious ceremonies (Kumar et al. 2019; Gupta and Kumar 2021; Huang et al. 2021), recreational activities (Boal and Mannan 1999; Mayhew et al. 2016; Godoy-Guinao et al. 2017; White et al. 2018), literature about their hunting skills and myths (Molares and Gurovich 2018; Pitas 2021) and within language (Stara et al. 2016). Given the frequency of these positive interactions, and considering the daily interactions raptors have with people can generate a sense of belonging and cohesion among groups (Hunold 2017; Mak et al. 2021), researchers should undoubtedly incorporate social methodologies, such as ethnography, into their future studies. This approach would enable them to capture the cultural beliefs and values that shape interactions between humans and raptors (Stara et al. 2016; Bennett et al. 2017). Integrating social insights with ecological data will enable the development of more effective and context-specific management and conservation strategies (Mascia et al. 2003).
Negative interactions
Safety damage
Aggressive behavior, like physical attacks or other aggressive interactions or displays, is one of the most direct impacts that wildlife can have on humans (Soulsbury and White 2015). Within urban environments, our understanding of wildlife aggressiveness is limited. This may be because wildlife attacks are not frequently documented in scientific studies, although they do appear in the media (Bhatia et al. 2013; Hathaway et al. 2017). Additionally, the scarcity of information could be attributed to the absence of large predators in urban areas (Soulsbury and White 2015). Nevertheless, there is a growing concern in this topic in urban raptors, as safety damage was the most extensively studied conflict in our review. In this sense, several behavioral shifts observed between urban and rural or natural counterparts have been attributed to three independent but not necessarily exclusive mechanisms: local evolution by divergent natural selection, phenotypic plasticity, and differential colonization process (Sol et al. 2013). Regardless of the specific mechanisms, animals in urbanized environments tend to exhibit increased aggression compared to their counterparts in rural or natural areas (Miranda 2017). As seen in mammals (McCullough 1982; Thompson et al. 2003) and highlighted in some studies in our review (McPherson et al. 2016; Kumar et al. 2018b, 2019), aggression sometimes is linked with intentional and unintentional human feeding of the predators, resulting in consequent loss of fear of humans.
Although the majority of studies were conducted during the reproductive season, a period typically associated with higher levels of aggressiveness (Montgomerie and Weatherhead 1988; Redondo 1989), these investigations predominantly reported low levels of aggressiveness with varied underlying causes. For instance, in India, aggressiveness of Black Kites was low and mostly occurred in areas with poor garbage disposal and, more importantly, where religious rituals involving feeding the kites were common (Kumar et al. 2018b, 2019). In contrast, in Japan, aggressive behavior of Black Kites was common and seems to be influenced by the landscape composition rather than human activities. In this sense, areas with fewer viable habitats for nesting and foraging witness a higher occurrence of aggressive behavior in Black Kites (Galbreath et al. 2014). Urban Mississippi Kites (Ictinia mississippiensis) exhibit lower levels of aggressive responses compared to their non-urban counterparts in the United States, likely because they have become accustomed to human presence (Skipper and Boal 2019; Boal et al. 2022). In this case, these negative results can contribute to carrying out management programs based on empirical facts to counteract the misaligned perceptions that exist about the aggression of this raptor (Dickman 2010; Boal et al. 2022). Attacks by the Crowned Eagle on pets were low (less than 1% of its diet) (McPherson et al. 2016) but enough to create negative human perception that may jeopardize the conservation of this raptor (McPherson et al. 2021b).
More research is necessary to rigorously understand the factors underlying and promoting the aggressive behaviors of raptors in urban environments. However, it cannot be overlooked that human behavior towards raptors might be a contributing factor to their increased aggressiveness. Therefore, it is essential to consider not only management actions targeting the wildlife involved but also how human behaviors contribute to the likelihood of conflicts (e.g., habituating raptors through supplementary feeding) (Nyhus 2016; Lambertucci et al. 2021). By doing so, accurate actions can be implemented that modify human behavior, favoring coexistence between humans and raptors (Mascia et al. 2003).
Property damage
Basak et al. (2023) found that property damage by urban wildlife was the most common conflict. However, in our review, though well-documented, it ranks below studies on safety damage and was on par with negative perception. The documented damage primarily resulted from collisions with aircraft, with all incidents occurring in the United States. The reviewed studies involved owls (Linnell and Washburn 2018), eagles (Washburn et al. 2015), vultures (Washburn et al. 2013, 2014) and osprey (Washburn 2014). Overall, it was observed that most collisions occur inside or near airports, particularly during takeoff or landing. Another common finding is the significant increase in collisions over time for all groups of raptors studied, which researchers attribute to the rise in populations and an increase in voluntary reporting of collisions. This was the only conflict where economic costs were calculated, which ranged from $US 15,131 to $US 425,945, depending on the species involved.
Certainly, our review highlights a geographic bias and specific focus on property damage, as no other types of damage were reported, such as damage to buildings or physical structures due to nest construction, or damage to ornamental gardens (Peterson et al. 2010). More research is needed on the topic to determine if raptor cause other types of the damage to urban infrastructure.
Livestock damage
Studies on livestock damage in urban areas were limited, as this is not a common activity carried out in cities. However, studies have documented instances of raptors preying on commercially used swifts (Dhamorikar et al. 2020), livestock (McPherson et al. 2016) and domestic species (Schneider et al. 2023). In all cases, the predation pressure was low and appears to have a negligible impact on economic activities. Given these results, it does not seem that damage to livestock is a conflict in which urban raptors are protagonists.
Nevertheless, potential opportunities for this type of conflict may become more frequent as backyard farming expands globally (Pollock et al. 2012; Mok et al. 2014). Considering that urban agriculture and raising chickens can enhance food security and social well-being, particularly in developing countries (Rajkumar et al. 2021), it is desirable to raise awareness in society about methods to exclude predators, akin to practices traditionally employed in rural areas (University of New Hampshire 2019; Bosques 2023).
Disease carrier
Zoonotic diseases can increase with urbanization, especially in growing cities. The high density of humans, pets and wildlife in urban areas creates conditions that facilitate the spread of diseases through increased contacts between species (Ellwanger et al. 2022). In addition to being a threat to human health, diseases can also incur significant economic costs associated with preventive measures and health treatments for both humans and wildlife (Soulsbury and White 2015). Despite this, we found only a few studies that evaluated this conflict in urban raptors. Most studies highlighted the prevalence of bacteria posing health risks to humans, such as Salmonella spp. in American black vultures in Argentina (Plaza et al. 2019) and Escherichia coli in free-living raptors in Spain (Vidal et al. 2017). While these results underscore the potential of the studied raptors as reservoirs of pathogens affecting domestic animals and human health, the capacity for dispersal or transmission was not addressed. Opposite, one case study revealed a bacterial cellulitis infection caused by injuries inflicted by the claws of a Harris’s hawk (Parabuteo unicinctus) (Khan et al. 2011), but no follow-up studies were conducted to evaluate the prevalence of this condition in this raptor’s population.
These results suggest that little is known about the zoonotic diseases of urban raptors, specifically those caused by viruses such as Highly Pathogenic Avian Influenza (HPAI) or West Nile virus. Monitoring wild populations would be beneficial to fill these information gaps and to have early warnings of possible pathogens that could harm human health (Gray et al. 2023) and raptor populations (Pearce-Higgins et al. 2023).
Nuisance
Only two studies highlighted nuisance of raptors in urban environments. In one of them, the Hooded vulture is considered a nuisance to the citizens of Ghana due to the theft of food and defecation when they invade homes (Campbell 2009). In the other study, homeowners removed nests from Barn Owls because they were bothered by the calls they made during the night (Pande et al. 2005). Although it has not been the conflict for which urban raptors stood out, urban wildlife is usually considered a nuisance (Soulsbury and White 2015). Indeed, crows, parrots and gulls have also demonstrated similar disruptive behaviors in urban settings (Belant 1997; Menchetti and Mori 2014; Campbell 2019; Benmazouz et al. 2021). Several management strategies have been implemented to address these issues. Among the simplest, most non-lethal and effective are modifications to nesting areas and reductions in foraging opportunities (e.g., at garbage dumps) (Belant 1997; Campbell 2019; Benmazouz et al. 2021). They could easily be implemented if the substrate to be used for nesting and the trophic resources consumed by the nuisance raptor species are known (see Mak et al. 2021). However, it is also crucial to consider the cognitive capacities of these birds, such as neophobia, innovation and several forms of learning, since these traits could enable them to flexibly circumvent non-lethal control measures (Barrett et al. 2019; Biondi 2022).
Superstitions
Symbolic beliefs, such as myths, narratives and superstitions, often shape the way animals are socially perceived and could influence human attitudes toward them (Horgan et al. 2021). While there are superstitions about raptors in many aboriginal and rural communities, there is limited research on this matter in urban areas. In our review, we could only find two studies, both from South America, that identify superstitions. For instance, in Argentina, people perceive owls as diabolic creatures, mainly due to their anthropomorphic face, broad movement of the neck (about 270°), nocturnal habits, and diet based on rodents and amphibians (Molares and Gurovich 2018). Similarly, in Chile, superstitions associated birds of prey with bad ill-omen or evil (Muñoz-Pedreros et al. 2018).
Raptors have long been integral to mythical narratives and religious cultures (Macdonald 2006; MaMing et al. 2016; Sax 2021; Soni 2022,), influencing the superstitions prevalent among aboriginal and rural communities (Enriquez 2017). Notably, in urban areas like Delhi, India, religious rituals have been observed to influence Black Kites habitat selection (Kumar et al. 2018a). This raises both interesting and necessary questions about whether religious factors similarly contribute to the creation of superstitions in urban settings. Further research is essential to determine if superstitions that originate in rural areas, towns, or Aboriginal communities continue to persist in urban environments and to assess their potential role in conflicts between humans and raptors.
Positive and negative perception
Perceptions could be defined as “the way an individual observes, understands, interprets, and evaluates a referent object, action, experience, individual, policy, or outcome” (Bennett 2016; p. 585). Thus, understanding people’s perceptions about wildlife can assist in developing conservation and management plans, implementing environmental education programs to improve their public image and facilitating appropriate urban planning to conserve the ecosystem services they provide (Martínez-Abrain et al. 2008; Basak et al. 2022). People’s perceptions of predators influence how they will behave towards them (Marchini and Macdonald 2018); therefore, it is important to document these responses. For instance, in Delhi, a megacity in India, the local populace shows remarkable tolerance towards the aggressive behavior of Black Kites due to their perceived benefits, including waste removal and their role in religious rituals (Kumar et al. 2019).
A clear pattern evident in previous studies (Kansky and Knight 2014; Puri et al. 2024; Zhao et al. 2024) also emerged in the studies we reviewed: citizens who had close contact with raptors generally developed positive attitudes towards them and showed a greater interest in their protection. Conversely, indifference and negative perceptions were more prevalent among individuals who did not recognize the raptor species or had not interacted with them. For example, in both the United States and Chile, people’s perceptions of Red-tailed Hawks and Barn Owls, respectively, improved with increased interaction with the birds in their nests (Godoy-Guinao et al. 2017; White et al. 2018). Another example occurs with vultures of the genus Gyps spp. and Neophron spp., where people over 35 years of age recognized cultural and carrion removal services from these birds because they had more experiences living with them than the younger ones (Gupta and Kumar 2021). An exception was found in Chile, where urban residents demonstrated low knowledge and identification of raptor species; however, despite this, they had a greater desire to protect birds of prey due to the pest control service they provide, compared to rural citizens (Muñoz-Pedreros et al. 2018). This aligns with previous studies that document extinction of experience, less persecution and greater respect for birds of prey in cities (Miller 2005; Boal and Dykstra 2018).
Environmental education and recreational activities that bring people into contact with raptors can improve people’s perception. In Argentina, students exhibited a positive change in attitude towards owls after a school laboratory experience dissecting owl pellets, where they learned about the crucial role these birds play in controlling rodents that transmit hantavirus (Molares and Gurovich 2018).
Some studies have revealed that people may hold both positive and negative perceptions towards birds of prey, and these perceptions may vary by geographic location. For example, Hooded vultures (Necrosyrtes monachus) are recognized for their carrion and waste removal services in Guinea Bissau and Ghana. However, there are concerns in Guinea Bissau that they might transmit diseases (Henriques et al. 2018), while in Ghana, people are worried about the theft of good food and nuisance behavior (Campbell 2009). Another example is the reintroduction of the white-tailed sea eagle (Haliaeetus albicilla), where the majority of people, especially in urban areas, supported the reintroduction due to the species’ utilitarian values (e.g. economic gains from ecotourism), while farmers were concerned about potential damage to their livestock (Mayhew et al. 2016). Despite several studies investigating people’s perception of urban raptors through interviews, such studies did not constitute the majority in this review. Moreover, only a few of them combined this social methodology with the evaluation of conflicts or services through experiments or observations. Given that cities are socio-ecological systems (Liu et al. 2007; Francis and Chadwick 2013), it is crucial to incorporate the study of human dimensions when examining urban species (Horgan et al. 2021; Basak et al. 2023). Attempting to identify and manage a conflict or maximize a service requires researchers to investigate people’s opinions about the raptors species and the interaction to be evaluated. This is because people may not perceive the service or conflict or may disagree with the measures taken, potentially impacting their effectiveness in the future (Basak et al. 2023). For example, species typically considered pests, such as pigeons or parakeets, may be viewed affectionately by some members of the public due to their frequent presence and interactions with people (Francis and Chadwick 2013; Crowley et al. 2019). Combined with people’s extinction of experience with nature (Miller 2005), raptors that prey on these species might be perceived negatively (Mak et al. 2021), leading to a devaluation of their pest control services. Recognizing and addressing these dualistic perspectives is essential for providing a comprehensive understanding of public attitudes towards raptors. This knowledge is crucial for designing educational programs and conservation strategies that not only foster public support but also enhance the effectiveness of interventions, benefiting both human communities and raptor populations (Burby 2003; Coz and Young 2020; Mak et al. 2021; Jones 2024).
Management measures
Although many studies demonstrated the existence of human-raptor conflict, few suggested management measures to reduce them. A full description of all the measures used or proposed to manage conflict and increase services is beyond the scope of this review. However, one widely suggested measure is environmental education. Since many people reside in or near to urban areas, there are ample opportunities to foster an ecologically informed public that appreciates nature and values its conservation (McKinney 2002).
Environmental education programs and persuasive communication about urban raptors could inform people about the ecosystem services and benefits they offer, such as pest control, and provide methods to avoid conflicts with them (Bruskotter and Wilson 2014). Additionally, these programs could encourage greater tolerance towards minor conflicts like nuisances or superstitions, especially when the perceived risks do not align with reality (Pande et al. 2005; Dickman 2010; Barrett et al. 2019; Lambertucci et al. 2021). They can also educate the public on when certain actions, such as providing supplementary food, might have both positive effects (providing a close nature experience or a conservation action for endangered raptors, e.g., “food restaurants” for vultures) and/or negative consequences (potentially habituating raptors and increasing aggression). This knowledge can empower people to take actions that benefit both humans and birds of prey (Bruskotter and Wilson 2014; Arnulphi et al. 2017).
By applying these recommendations and considering the various human dimensions that influence tolerance towards raptors and wildlife in general, education programs could promote coexistence between humans and raptors (Kansky et al. 2016, 2021; Kansky and Kidd 2024).
Despite the effective results evidenced in this review, for environmental education to be an efficient strategy to promote coexistence between humans and raptors, it is important to highlight that researchers must be open to what people can understand and relate to in nature, even if it differs from the scientific point of view. Therefore, they must avoid imposing their “scientific vision” on the public (see examples of these situations in rewilding projects (Deary and Warren 2017; Pettersson et al. 2023). Furthermore, when planning education programs, it is crucial to consider the heterogeneity of tolerance and perceptions based on demographic factors to make education and communication strategies appropriately for the target audience (Puri et al. 2024). Finally, programs should go beyond merely providing information and should also facilitate encounters with wildlife. Promoting activities like birdwatching for urban raptors in parks allows people to learn and appreciate these interactions firsthand (Ballantyne et al. 2007; Bruskotter and Wilson 2014; Puri et al. 2024).
Conclusion
Based on our review, raptors play a crucial role in urban environments, either by providing ecosystem services or by causing conflicts with humans. However, much remains to be explored in this field, as only three types of interactions (pest control, cultural services, and safety damage) and two orders (Accipitriformes and Strigiformes) have been studied extensively. Moreover, our review underscores the lack of transdisciplinary studies that integrate social and ecological approaches. Given that urban environments function as socio-ecological systems, we noted that human dimensions significantly influence the occurrence and magnitude of certain interactions, including safety, superstition, cultural services and both positive and negative perceptions. Consequently, it is essential to understand not only the ecological dynamics of these interactions but also to integrate social perspectives (Basak et al. 2022). This approach is vital not only for uncovering the underlying causes of specific conflicts but also for understanding people’s perceptions of raptors. We support the call for a holistic research approach, as previously advocated in raptor conservation (Canney et al. 2022; Carucci et al. 2022), to minimize human-wildlife conflicts in urban settings (Dickman 2010; Basak et al. 2022, 2023) and more broadly within the field of conservation (Mascia et al. 2003; Bennett 2016; Bennett et al. 2017). Additionally, implementing experimental alongside observational studies would be beneficial to thoroughly evaluate ecosystem functions and quantify the extent of services and conflicts associated with raptors in urban environments (Whelan et al. 2008). Adopting these approaches will likely enhance the coexistence between humans and raptors in urban landscapes.
Data availability
The data presented in this study is available in Table SM1 of the supplementary material.
References
Adams WM, Aveling R, Brockington D et al (2004) Biodiversity Conservation and the eradication of poverty. Science 306:1146–1149. https://doi.org/10.1126/science.1097920
Arnulphi VBC, Lambertucci SA, Borghi CE (2017) Education can improve the negative perception of a threatened long-lived scavenging bird, the Andean condor. PLoS ONE 12:e0185278. https://doi.org/10.1371/journal.pone.0185278
Autilio AR, Bechard MJ, Bildstein KL (2019) Social scavenging by wintering striated caracaras (Phalcoboenus australis) in the Falkland Islands. Behav Ecol Sociobiol 73:27. https://doi.org/10.1007/s00265-019-2638-3
Baladrón AV, Cavalli M, Isacch JP, Bó MS (2020) Burrowing owl nest distribution and density in relation to urban development. Ethol Ecol Evol 32:237–250. https://doi.org/10.1080/03949370.2020.1711814
Ballantyne R, Packer J, Hughes K, Dierking L (2007) Conservation learning in wildlife tourism settings: lessons from research in zoos and aquariums. Environ Educ Res 13:367–383. https://doi.org/10.1080/13504620701430604
Ballejo F, Plaza P, Virgilio A, di et al (2022) Unravelling negative interactions between humans, mammalian carnivores and raptors in South America. Ecol Austral 32:620–637. https://doi.org/10.25260/EA.22.32.2.1.1843
Barrett LP, Stanton LA, Benson-Amram S (2019) The cognition of ‘nuisance’ species. Anim Behav 147:167–177. https://doi.org/10.1016/j.anbehav.2018.05.005
Basak SM, Hossain MdS, O’Mahony DT et al (2022) Public perceptions and attitudes toward urban wildlife encounters – A decade of change. Sci Total Environ 834:155603. https://doi.org/10.1016/j.scitotenv.2022.155603
Basak SM, Rostovskaya E, Birks J, Wierzbowska IA (2023) Perceptions and attitudes to understand human-wildlife conflict in an urban landscape – a systematic review. Ecol Indic 151:110319. https://doi.org/10.1016/j.ecolind.2023.110319
Belant JL (1997) Gulls in urban environments: landscape-level management to reduce conflict. Landsc Urban Plan 38:245–258. https://doi.org/10.1016/S0169-2046(97)00037-6
Benmazouz I, Jokimäki J, Lengyel S et al (2021) Corvids in Urban environments: a systematic global literature review. Animals 11:3226. https://doi.org/10.3390/ani11113226
Bennett NJ (2016) Using perceptions as evidence to improve conservation and environmental management. Conserv Biol 30:582–592. https://doi.org/10.1111/cobi.12681
Bennett NJ, Roth R, Klain SC et al (2017) Conservation social science: understanding and integrating human dimensions to improve conservation. Biol Conserv 205:93–108. https://doi.org/10.1016/j.biocon.2016.10.006
Bhatia S, Athreya V, Grenyer R, Macdonald DW (2013) Understanding the role of representations of human–Leopard conflict in Mumbai through media-content analysis. Conserv Biol 27:588–594. https://doi.org/10.1111/cobi.12037
Bildstein KL, Therrien J-F (2018) Urban birds of Prey: a Lengthy History of Human-Raptor Cohabitation. Urban Raptors: Ecology and Conservation of birds of Prey in cities. Island, Washington, DC, pp 3–17
Billerman SM, Keeney BK, Rodewald PG, Schulenberg TS (2022) Birds of the World. Cornell Laboratory of Ornithology. https://birdsoftheworld.org/bow/home. Accessed 23 Feb 2024
Biondi LM (2022) Falconiformes cognition. In: Vonk J, Shackelford TK (eds) Encyclopedia of animal cognition and behavior. Springer International Publishing, Cham, pp 2605–2613
Biondi L, María S, Bó, Favero M (2005) Dieta Del Chimango (Milvago chimango) Durante El período reproductivo en El Sudeste De La Provincia De Buenos Aires. Argentina Ornitol Neotrop 16
BirdLife I (2021) IUCN Red list of threatened species: Neophron percnopterus. https://doi.org/10.2305/IUCN.UK.2021-3.RLTS.T22695180A205187871. IUCN Red List of Threatened Species
Bleicher SS (2017) The landscape of fear conceptual framework: definition and review of current applications and misuses. PeerJ 5:e3772. https://doi.org/10.7717/peerj.3772
Boakes Z, Stafford R, Bramer I et al (2023) The importance of urban areas in supporting vulnerable and endangered mammals. https://doi.org/10.1007/s11252-023-01492-z. Urban Ecosyst
Boal CW, Dykstra CR (2018) Urban Raptors: Ecology and Conservation of birds of Prey in cities. Island
Boal CW, Mannan RW (1999) Comparative Breeding Ecology of Cooper’s Hawks in Urban and Exurban areas of Southeastern. J Wildl 63:77–84
Boal CW, Bibles BD, Pryor MM, Skipper BR (2022) Mississippi Kite Nest Defense: is there an influence of Nest phenology or human activity? J Raptor Res 56:356–361. https://doi.org/10.3356/JRR-21-74
Bosques J (2023) Ruling the roost: Showing predators who is the boss of your backyard poultry colony. In: UF/IFAS Extension Hardee County. https://blogs.ifas.ufl.edu/hardeeco/2023/09/29/ruling-the-roost-showing-predators-who-is-the-boss-of-your-backyard-poultry-colony/. Accessed 21 Jun 2024
Bruskotter JT, Wilson RS (2014) Determining where the wild things will be: using Psychological Theory to find tolerance for large carnivores. Conserv Lett 7:158–165. https://doi.org/10.1111/conl.12072
Buechley ER, Şekercioğlu ÇH (2016) The avian scavenger crisis: looming extinctions, trophic cascades, and loss of critical ecosystem functions. Biol Conserv 198:220–228. https://doi.org/10.1016/j.biocon.2016.04.001
Buechley ER, McGrady MJ, Çoban E, Şekercioğlu ÇH (2018) Satellite tracking a wide-ranging endangered vulture species to target conservation actions in the Middle East and East Africa. Biodivers Conserv 27:2293–2310. https://doi.org/10.1007/s10531-018-1538-6
Buechley ER, Murgatroyd M, Ruffo AD et al (2022) Declines in scavenging by endangered vultures in the Horn of Africa. J Wildl Manag 86:e22194. https://doi.org/10.1002/jwmg.22194
Burby RJ (2003) Making plans that Matter: Citizen involvement and government action. JAPA 69:33–49. https://doi.org/10.1080/01944360308976292
Campbell M (2009) Factors for the presence of avian scavengers in Accra and Kumasi, Ghana. Area 41:341–349. https://doi.org/10.1111/j.1475-4762.2008.00870.x
Campbell MO (2019) Links Between Human Quality of Life and Gulls in Urban Areas | SpringerLink. In: Multidimensional Approach to Quality of Life Issues: A Spatial Analysis. Springer Nature Singapore Pte Ltd, Singapore, India, p XXVII, 457
Canney AC, McGough LM, Bickford NA, Wallen KE (2022) Systematic map of human–Raptor Interaction and Coexistence Research. Animals 12:45. https://doi.org/10.3390/ani12010045
Carrete M, Tella JL, Blanco G, Bertellotti M (2009) Effects of habitat degradation on the abundance, richness and diversity of raptors across neotropical biomes. Biol Conserv 142:2002–2011. https://doi.org/10.1016/j.biocon.2009.02.012
Carucci T, Whitehouse-Tedd K, Yarnell RW et al (2022) Ecosystem services and disservices associated with vultures: a systematic review and evidence assessment. Ecosyst Serv 56:101447. https://doi.org/10.1016/j.ecoser.2022.101447
Cavalli M, Baladrón AV, Bó MS, Isacch JP (2023) Owls and the City: the breeding performance of burrowing owls Athene cunicularia is better in Urban Than in Rural Areas. Ardeola 71:81–100. https://doi.org/10.13157/arla.71.1.2024.ra5
Chace JF, Walsh JJ (2006) Urban effects on native avifauna: a review. Landsc Urban Plan 74:46–69. https://doi.org/10.1016/j.landurbplan.2004.08.007
Cherkaoui SI, Rihane A, Chergui El Hemiani B (2021) Diet of a Maghreb owl pair Strix mauritanica (Witherby, 1905) in an urban environment (Rabat City, Morocco). Ostrich 92:319–323. https://doi.org/10.2989/00306525.2021.2003884
Collins MK, Magle SB, Gallo T (2021) Global trends in urban wildlife ecology and conservation. Biol Conserv 261:109236. https://doi.org/10.1016/j.biocon.2021.109236
Conover MR (2001) Resolving Human-Wildlife conflicts: the Science of Wildlife damage management. CRC
Cooper DS, Yeh PJ, Blumstein DT (2021) Tolerance and avoidance of urban cover in a southern California suburban raptor community over five decades. Urban Ecosyst 24:291–300. https://doi.org/10.1007/s11252-020-01035-w
Cox DTC, Shanahan DF, Hudson HL et al (2017) Doses of Neighborhood Nature: the benefits for Mental Health of living with Nature. Bioscience 67:147–155. https://doi.org/10.1093/biosci/biw173
Coz DM, Young JC (2020) Conflicts over wildlife conservation: learning from the reintroduction of beavers in Scotland. People Nat 2:406–419. https://doi.org/10.1002/pan3.10076
Crowley SL, Hinchliffe S, McDonald RA (2019) The parakeet protectors: understanding opposition to introduced species management. J Environ Manag 229:120–132. https://doi.org/10.1016/j.jenvman.2017.11.036
Deary H, Warren CR (2017) Divergent visions of wildness and naturalness in a storied landscape: practices and discourses of rewilding in Scotland’s wild places. J Rural Stud 54:211–222. https://doi.org/10.1016/j.jrurstud.2017.06.019
DeVault TL, Rhodes OE Jr, Shivik JA (2003) Scavenging by vertebrates: behavioral, ecological, and evolutionary perspectives on an important energy transfer pathway in terrestrial ecosystems. Oikos 102:225–234. https://doi.org/10.1034/j.1600-0706.2003.12378.x
DeVault TL, Beasley JC, Olson ZH et al (2016) Ecosystem services provided by Avian scavengers. Why birds Matter: avian ecological function and ecosystem services. The University of Chicago, United States of America
Dhamorikar A, Kawalkar D, Gurjarpadhye P, Manchi S (2020) Crepuscular hunting of swiftlets (family: Apodidae) by Besra (Family: Accipitridae) in the urban areas of the Andaman Islands, India. J Threat Taxa 12:17323–17329. https://doi.org/10.11609/jott.5397.12.17.17323-17329
Díaz S, Pascual U, Stenseke M et al (2018) Assessing nature’s contributions to people. People Nat 359:270–272. https://doi.org/10.1126/science.aap8826
Dickman AJ (2010) Complexities of conflict: the importance of considering social factors for effectively resolving human–wildlife conflict. Anim Conserv 13:458–466. https://doi.org/10.1111/j.1469-1795.2010.00368.x
Ditchkoff SS, Saalfeld ST, Gibson CJ (2006) Animal behavior in urban ecosystems: modifications due to human-induced stress. Urban Ecosyst 9:5–12. https://doi.org/10.1007/s11252-006-3262-3
Donázar JA, Cortés-Avizanda A, Fargallo JA et al (2016) Roles of raptors in a changing world: from flagships to providers of Key Ecosystem services. Ardeola 63:181–234. https://doi.org/10.13157/arla.63.1.2016.rp8
Drewitt EJA, Cuthill IC, Sutton LJ et al (2021) Northerly dispersal trends in a lowland population of Peregrines Falco peregrinus in Southwest England. Ring Migr 36:105–115. https://doi.org/10.1080/03078698.2022.2150783
Ellwanger JH, Byrne LB, Chies JAB (2022) Examining the paradox of urban disease ecology by linking the perspectives of Urban One Health and Ecology with cities. Urban Ecosyst 25:1735–1744. https://doi.org/10.1007/s11252-022-01260-5
Enriquez PL (2017) Neotropical owls: diversity and conservation. Springer Cham
Ferguson-Lees J, Christie DA (2001) Raptors of the World. Houghton Mifflin Harcourt, Boston
Francis RA, Chadwick MA (2013) Urban ecosystems: understanding the human environment. Routledge, London
Franco FF, Marçal-Junior O (2018) Influence of urbanization on the distribution and defense strategies of the burrowing owl Athene cunicularia in the city of Uberlândia, southeastern Brazil. Rev Bras Ornitol 26:1–8. https://doi.org/10.1007/BF03544410
Galbreath DM, Ichinose T, Furutani T et al (2014) Urbanization and its implications for avian aggression: a case study of urban black kites (Milvus migrans) along Sagami Bay in Japan. Landsc Ecol 29:169–178. https://doi.org/10.1007/s10980-013-9951-4
Gangoso L, Agudo R, Anadón JD et al (2013) Reinventing mutualism between humans and wild fauna: insights from vultures as ecosystem services providers. Conserv Lett 6:172–179. https://doi.org/10.1111/j.1755-263X.2012.00289.x
Gaston KJ (2010) Valuing common species. Science 327:154–155. https://doi.org/10.1126/science.1182818
Gaston KJ (2019) Nighttime Ecology: the Nocturnal Problem. Revisit Am Nat 193:481–502. https://doi.org/10.1086/702250
Gilbert M, Watson RT, Ahmed S et al (2007) Vulture restaurants and their role in reducing diclofenac exposure in Asian vultures. Bird Conserv Int 17:63–77. https://doi.org/10.1017/S0959270906000621
Godoy-Guinao J, Díaz IA, Llanos- Pineda M, Aló D (2017) Feeding habits and people’s perception of the Barn owl (Tyto alba tuidara, J.E.Gray 1829) in urban settings of Southern Chile: implications for conservation. Gayana 81:9–16
Graham K, Beckerman AP, Thirgood S (2005) Human–predator–prey conflicts: ecological correlates, prey losses and patterns of management. Biol Conserv 122:159–171. https://doi.org/10.1016/j.biocon.2004.06.006
Gray A, Doyle S, Doyle C et al (2023) Birds and human health: pathways for a positive relationship and improved integration. Ibis n/a. https://doi.org/10.1111/ibi.13290
Grilli MG, Bildstein KL, Lambertucci SA (2019) Nature’s clean-up crew: quantifying ecosystem services offered by a migratory avian scavenger on a continental scale. Ecosyst Serv 39:100990. https://doi.org/10.1016/j.ecoser.2019.100990
Gupta U, Kumar N (2021) Vulture loss and urbanisation correlations form blind spots for conservation and development. https://doi.org/10.21203/rs.3.rs-505472/v2
Hassan-Montero Y, De-Moya-Anegón F, Guerrero-Bote VP (2022) SCImago Graphica: a new tool for exploring and visually communicating data. https://doi.org/10.3145/epi.2022.sep.02. Prof Inf 31:
Hathaway RS, Bryant A-EM, Draheim MM et al (2017) From fear to understanding: changes in media representations of leopard incidences after media awareness workshops in Mumbai, India. J Urban Ecol 3:jux009. https://doi.org/10.1093/jue/jux009.
Headland T, Colombelli-Négrel D, Callaghan CT et al (2023) Smaller Australian raptors have greater urban tolerance. Sci Rep 13:11559. https://doi.org/10.1038/s41598-023-38493-z
Henriques M, Granadeiro JP, Monteiro H et al (2018) Not in wilderness: African vulture strongholds remain in areas with high human density. PLoS ONE 13. https://doi.org/10.1371/journal.pone.0190594
Horgan FG, Mundaca EA, Crisol-Martínez E (2021) Emerging Patterns in Cultural Ecosystem Services as incentives and obstacles for Raptor Conservation. Birds 2:185–206. https://doi.org/10.3390/birds2020014
Huang Y-K, Lemaitre A, Wu H-J, Sun Y-H (2021) A Sacred Bird at the crossroads of destiny: Ethno-Ornithology of the Mountain Hawk-Eagle (Qadis) for the Paiwan people in Taiwan. J Ethnobiol 535–552
Huijbers CM, Schlacher TA, Schoeman DS et al (2013) Urbanisation alters processing of marine carrion on sandy beaches. Landsc Urban Plan 119:1–8. https://doi.org/10.1016/j.landurbplan.2013.06.004
Huijbers CM, Schlacher TA, Schoeman DS et al (2015) Limited functional redundancy in vertebrate scavenger guilds fails to compensate for the loss of raptors from urbanized sandy beaches. Divers Distrib 21:55–63. https://doi.org/10.1111/ddi.12282
Hunold C (2017) Why not the City? Urban Hawk watching and the end of Nature. Nat Cult 12:115–136. https://doi.org/10.3167/nc.2017.120202
Isaksson C (2018) Impact of Urbanization on Birds. In: Bird Species: How They Arise, Modify and Vanish, Dieter Thomas Tietze. Springer International Publishing, Cham, Switzerland, p XII, 266
IUCN (2024) The IUCN Red List of Threatened Species. In: IUCN Red List of Threatened Species. https://www.iucnredlist.org/en. Accessed 17 Jan 2024
Ives CD, Lentini PE, Threlfall CG et al (2016) Cities are hotspots for threatened species. Glob Ecol Biogeogr 25:117–126. https://doi.org/10.1111/geb.12404
Jones MS (2024) Integrating the human dimensions into fish and wildlife management depends on increasing managers’ social science fluency. Hum Dimens Wildl 0:1–8. https://doi.org/10.1080/10871209.2024.2301965
Kansky R, Kidd M (2024) Putting yourself in an animal’s shoes - empathy and intangible benefits drive tolerance towards wildlife in Namibian communal conservancies. Biol Conserv 293:110588. https://doi.org/10.1016/j.biocon.2024.110588
Kansky R, Knight A (2014) Key factors driving attitudes towards large mammals in conflict with humans. Biol Conserv 179:93–105. https://doi.org/10.1016/j.biocon.2014.09.008
Kansky R, Kidd M, Knight AT (2016) A wildlife tolerance model and case study for understanding human wildlife conflicts. Biol Conserv 201:137–145. https://doi.org/10.1016/j.biocon.2016.07.002
Kansky R, Kidd M, Fischer J (2021) Understanding drivers of human tolerance towards mammals in a mixed-use transfrontier conservation area in southern Africa. Biol Conserv 254:108947. https://doi.org/10.1016/j.biocon.2020.108947
Kettel EF, Gentle LK, Quinn JL, Yarnell RW (2018) The breeding performance of raptors in urban landscapes: a review and meta-analysis. J Ornithol 159:1–18. https://doi.org/10.1007/s10336-017-1497-9
Khan MAA, Farid M, Sobani ZA et al (2011) A case of cellulitis of the hands caused by a predatory bird attack. Asian Pac J Trop Biomed 1:166–168. https://doi.org/10.1016/S2221-1691(11)60019-1
Kopij G, Liven-Schulman I (2012) Diet of the lesser kestrel, falco naumanni, in Israel: (Aves: Falconiformes). Zool Middle East 55:27–34. https://doi.org/10.1080/09397140.2012.10648914
Kumar N (2023) Ecological impacts of poultry waste on urban raptors: conflicts, diseases, and climate change implications amidst pandemic threats. 2023.07.13.546415
Kumar N, Gupta U, Jhala YV et al (2018a) Habitat selection by an avian top predator in the tropical megacity of Delhi: human activities and socio-religious practices as prey-facilitating tools. Urban Ecosyst 21:339–349. https://doi.org/10.1007/s11252-017-0716-8
Kumar N, Qureshi Q, Jhala YV et al (2018b) Offspring defense by an urban raptor responds to human subsidies and ritual animal-feeding practices. PLoS ONE 13. https://doi.org/10.1371/journal.pone.0204549
Kumar N, Jhala YV, Qureshi Q et al (2019) Human-attacks by an urban raptor are tied to human subsidies and religious practices. Sci Rep 9. https://doi.org/10.1038/s41598-019-38662-z
Lambertucci SA, Speziale KL, Rogers TE, Morales JM (2009) How do roads affect the habitat use of an assemblage of scavenging raptors? Biodivers Conserv 18:2063–2074. https://doi.org/10.1007/s10531-008-9573-3
Lambertucci SA, Plaza P, Speziale K (2021) Minimizing fear of wildlife in urban areas. Science 374:947–947. https://doi.org/10.1126/science.abm6560
Linnell KE, Washburn BE (2018) Assessing owl collisions with us civil and US Air Force Aircraft. J Raptor Res 52:282–290. https://doi.org/10.3356/JRR-17-64.1
Liu J, Dietz T, Carpenter SR et al (2007) Complexity of coupled human and natural systems. Science 317:1513–1516. https://doi.org/10.1126/science.1144004
Luederitz C, Brink E, Gralla F et al (2015) A review of urban ecosystem services: six key challenges for future research. Ecosyst Serv 14:98–112. https://doi.org/10.1016/j.ecoser.2015.05.001
Luna Á, Romero-Vidal P, Arrondo E (2021) Predation and Scavenging in the City: a review of spatio-temporal trends in Research. Diversity 13:46. https://doi.org/10.3390/d13020046
Macdonald H (2006) Falcon. Reaktion Books, London
MacGregor-Fors I, Morales-Pérez L, Schondube JE (2012) From forests to cities: effects of Urbanization on Tropical Birds. In: Lepczyk CA, Warren PS (eds) Urban Bird Ecology and Conservation. University of California Press, pp 33–48
Machlis GE, Carrero-Martinez FA (2024) Scientists in exile. Science 384:1155–1155. https://doi.org/10.1126/science.adq9574
Magle SB, Hunt VM, Vernon M, Crooks KR (2012) Urban wildlife research: past, present, and future. Biol Conserv 155:23–32. https://doi.org/10.1016/j.biocon.2012.06.018
Magrini L, Facure KG (2008) Barn owl (Tyto alba) predation on small mammals and its role in the control of hantavirus natural reservoirs in a periurban area in southeastern Brazil. Braz J Biol 68:733–740
Mak B, Francis RA, Chadwick MA (2021) Living in the concrete jungle: a review and socio-ecological perspective of urban raptor habitat quality in Europe. Urban Ecosyst 24:1179–1199. https://doi.org/10.1007/s11252-021-01106-6
MaMing R, Lee L, Yang X, Buzzard P (2016) Vultures and sky burials on the Qinghai-Tibet Plateau. Vulture News 71:22–35. https://doi.org/10.4314/vulnew.v71i1.2
Marchini S, Macdonald DW (2018) Mind over matter: perceptions behind the impact of jaguars on human livelihoods. Biol Conserv 224:230–237. https://doi.org/10.1016/j.biocon.2018.06.001
Mariyappan M, Rajendran M, Velu S et al (2023) Ecological role and Ecosystem Services of Birds: a review. Int J Environ Clim 13:76–87. https://doi.org/10.9734/ijecc/2023/v13i61800
Markandya A, Taylor T, Longo A et al (2008) Counting the cost of vulture decline—An appraisal of the human health and other benefits of vultures in India. Ecol Econ 67:194–204. https://doi.org/10.1016/j.ecolecon.2008.04.020
Martínez-Abrain A, Crespo J, Jiménez J et al (2008) Friend or foe: societal shifts from intense persecution to active conservation of top predators. Ardeola 55:111–119
Marzluff JM, Bowman R, Donnelly R (2001) A historical perspective on urban bird research: trends, terms, and approaches. In: Marzluff JM, Bowman R, Donnelly R (eds) Avian Ecology and Conservation in an Urbanizing World. Springer US, Boston, MA, pp 1–17
Mascia MB, Brosius JP, Dobson TA et al (2003) Conservation and the Social Sciences. Conserv Biol 17:649–650. https://doi.org/10.1046/j.1523-1739.2003.01738.x
Mayhew M, Convery I, Armstrong R, Sinclair B (2016) Public perceptions of a white-tailed sea eagle (Haliaeetus albicilla L.) restoration program. Restor Ecol 24:271–279. https://doi.org/10.1111/rec.12310
Mazumdar S, Ghose D, Saha GK (2017) Offal dumping sites influence the relative abundance and roosting site selection of Black kites (Milvus migrans govinda) in urban landscape: a study from Kolkata metropolis, India. Environ Monit Assess 190:20. https://doi.org/10.1007/s10661-017-6391-7
McClure CJW, Westrip JRS, Johnson JA et al (2018) State of the world’s raptors: distributions, threats, and conservation recommendations. Biol Conserv 227:390–402. https://doi.org/10.1016/j.biocon.2018.08.012
McClure C, Vargas F, Amar A et al (2022) Conservation letter: monitoring raptor populations – A call for increased global collaboration and survey standardization. J Raptor Res 57. https://doi.org/10.3356/JRR-22-68
McCullough DR (1982) Behavior, bears, and humans. Wildl Soc Bull (1973–2006) 10:27–33
McKinney ML (2002) Urbanization, Biodiversity, and conservation: the impacts of urbanization on native species are poorly studied, but educating a highly urbanized human population about these impacts can greatly improve species conservation in all ecosystems. Bioscience 52:883–890. https://doi.org/10.1641/0006-3568(2002)052[0883:UBAC]2.0.CO;2
McPherson SC, Brown M, Downs CT (2016) Diet of the crowned eagle (Stephanoaetus coronatus) in an urban landscape: potential for human-wildlife conflict? Urban Ecosyst 19:383–396. https://doi.org/10.1007/s11252-015-0500-6
McPherson SC, Sumasgutner P, Downs CT (2021a) South African raptors in urban landscapes: a review. Ostrich 92:41–57. https://doi.org/10.2989/00306525.2021.1900942
McPherson SC, Sumasgutner P, Hoffman BH et al (2021b) Surviving the urban jungle: anthropogenic threats, Wildlife-Conflicts, and Management recommendations for African Crowned eagles. Front Ecol Evol 9. https://doi.org/10.3389/fevo.2021.662623
Menchetti M, Mori E (2014) Worldwide impact of alien parrots (Aves Psittaciformes) on native biodiversity and environment: a review. Ethol Ecol Evol 26:172–194. https://doi.org/10.1080/03949370.2014.905981
Millennium Ecosystem Assessment (2005) Ecosystems and Human Well-being: synthesis. Island, Washington, DC
Miller JR (2005) Biodiversity conservation and the extinction of experience. Trends Ecol Evol 20:430–434. https://doi.org/10.1016/j.tree.2005.05.013
Miranda AC (2017) Mechanisms of behavioural change in Urban animals: the role of Microevolution and phenotypic plasticity. In: Murgui E, Hedblom M (eds) Ecology and Conservation of birds in Urban environments. Springer International Publishing, Cham, pp 113–132
Mmereki D, Baldwin A, Li B (2016) A comparative analysis of solid waste management in developed, developing and lesser developed countries. Environ Technol Rev 5:120–141. https://doi.org/10.1080/21622515.2016.1259357
Mok H-F, Williamson VG, Grove JR et al (2014) Strawberry fields forever? Urban agriculture in developed countries: a review. Agron Sustain Dev 34:21–43. https://doi.org/10.1007/s13593-013-0156-7
Molares S, Gurovich Y (2018) Owls in urban narratives: implications for conservation and environmental education in NW Patagonia (Argentina). Neotrop Biodivers 4:164–172. https://doi.org/10.1080/23766808.2018.1545379
Montgomerie RD, Weatherhead PJ (1988) Risks and rewards of Nest Defence by parent birds. Q Rev Biol 63:167–187. https://doi.org/10.1086/415838
Mori E, Malfatti L, Le Louarn M et al (2020) Some like it alien: predation on invasive ring–necked parakeets by the long–eared owl in an urban area. Anim Biodivers Conserv 43:151–158. https://doi.org/10.32800/abc.2020.43.0151
Morrison JL, Saggese MD (2024) Assessing knowledge of the Caracaras: compiling information, identifying knowledge gaps, and recommendations for Future Research. J Raptor Res 58:1–12. https://doi.org/10.3356/JRR-23-39
Muñoz-Pedreros A, Guerrero M, Möller P (2018) Knowledge and perceptions of birds of prey among local inhabitants in Chile: implications for the biological control of rodent pests. Gayana 82:128–138
Nyhus PJ (2016) Human–Wildlife Conflict and Coexistence. Annu Rev Environ Resour 41:143–171. https://doi.org/10.1146/annurev-environ-110615-085634
Page MJ, McKenzie JE, Bossuyt PM et al (2021) The PRISMA 2020 statement: an updated guideline for reporting systematic reviews. Syst Rev 10:89. https://doi.org/10.1186/s13643-021-01626-4
Pande S, Pawashe A, Karambelkar U, Shrotri S (2005) Salvage, relocation and in-nest behaviour of Barn Owl Tyto alba chicks. Indian Birds 1:5–6
Pearce-Higgins JW, Humphreys EM, Burton NHK et al (2023) Highly pathogenic avian influenza in wild birds in the United Kingdom in 2022: impacts, planning for future outbreaks, and conservation and research priorities. Thetford, UK
Peterson MN, Birckhead JL, Leong K et al (2010) Rearticulating the myth of human–wildlife conflict. Conserv Lett 3:74–82. https://doi.org/10.1111/j.1755-263X.2010.00099.x
Pettersson HL, Holmes G, Quinn CH et al (2023) Who must adapt to whom? Contested discourses on human–wolf coexistence and their impact on policy in Spain. People Nat 5:1989–2005. https://doi.org/10.1002/pan3.10543
Piano E, Souffreau C, Merckx T et al (2020) Urbanization drives cross-taxon declines in abundance and diversity at multiple spatial scales. Glob Change Biol 26:1196–1211. https://doi.org/10.1111/gcb.14934
Pitas JH (2021) Deathly storytelling in the ecological city: how pigeons became falcon food in Baltimore. Md Soc Cult Geogr 23:29–46. https://doi.org/10.1080/14649365.2021.1950822
Plaza PI, Lambertucci SA (2017) How are garbage dumps impacting vertebrate demography, health, and conservation? Glob Ecol Conserv 12:9–20. https://doi.org/10.1016/j.gecco.2017.08.002
Plaza PI, Blanco G, Madariaga MJ et al (2019) Scavenger birds exploiting rubbish dumps: pathogens at the gates. Transbound Emerg Dis 66:873–881. https://doi.org/10.1111/tbed.13097
Pollock SL, Stephen C, Skuridina N, Kosatsky T (2012) Raising chickens in City backyards: the Public Health Role. J Community Health 37:734–742. https://doi.org/10.1007/s10900-011-9504-1
Puri M, Johannsen KL, Goode KO, Pienaar EF (2024) Addressing the challenge of wildlife conservation in urban landscapes by increasing human tolerance for wildlife. People Nat 6:1116–1129. https://doi.org/10.1002/pan3.10604
R Core Team (2023) R: a Language and. Environment for Statistical Computing
Rajkumar U, Rama Rao SV, Raju MVLN, Chatterjee RN (2021) Backyard poultry farming for sustained production and enhanced nutritional and livelihood security with special reference to India: a review. Trop Anim Health Prod 53:176. https://doi.org/10.1007/s11250-021-02621-6
Rebolo-Ifrán N, Tella JL, Carrete M (2017) Urban conservation hotspots: predation release allows the grassland-specialist burrowing owl to perform better in the city. Sci Rep 7:3527. https://doi.org/10.1038/s41598-017-03853-z
Redondo T (1989) Avian nest defence: theoretical models and evidence. Behaviour 111:161–195
Saufi S, Ravindran S, Hamid NH et al (2020) Diet composition of introduced barn owls (Tyto alba javanica) in urban area in comparison with agriculture settings. J Urban Ecol 6. https://doi.org/10.1093/jue/juz025
Sax B (2021) Avian illuminations: a Cultural history of birds. Reaktion Books, London
Schneider LM, de Oliveira Santos C, Moreira-Lima L, Hingst-Zaher E (2023) Peregrine falcon falco peregrinus in Brazil: natural history through the lens of citizen science. Ornitol Neotrop 34:29–39. https://doi.org/10.58843/ornneo.v34i1.1121
Sebastián-González E, Barbosa JM, Pérez-García JM et al (2019) Scavenging in the Anthropocene: human impact drives vertebrate scavenger species richness at a global scale. Glob Change Biol 25:3005–3017. https://doi.org/10.1111/gcb.14708
Sekercioglu ÇH, Wenny DG, Whelan CJ (2016) Why birds Matter: Avian Ecological functions and Ecosystem services. The University of Chicago Press
Seto KC, Sánchez-Rodríguez R, Fragkias M (2010) The New Geography of Contemporary Urbanization and the Environment. Annu Rev Environ Resour 35:167–194. https://doi.org/10.1146/annurev-environ-100809-125336
Seto KC, Fragkias M, Güneralp B, Reilly MK (2011) A Meta-analysis of global Urban Land Expansion. PLoS ONE 6:e23777. https://doi.org/10.1371/journal.pone.0023777
Shizukuda K, Saito MU (2023) The disappearance of small mammal carcasses in human-dominated habitats: a field experiment in Northeastern Japan. Diversity 15:339. https://doi.org/10.3390/d15030339
Skipper BR, Boal CW (2019) Nest-defense behavior of Mississippi kites in urban and exurban areas. Hum-Wildl Interact 13:142–149
Sol D, Lapiedra O, González-Lagos C (2013) Behavioural adjustments for a life in the city. Anim Behav 85:1101–1112. https://doi.org/10.1016/j.anbehav.2013.01.023
Soni HB (2022) Owl The Mysterious Bird. Pencil, India
Sorace A (2002) High density of bird and pest species in urban habitats and the role of predator abundance. Ornis Fenn 76:60–71
Soulsbury CD, White PCL (2015) Human–wildlife interactions in urban areas: a review of conflicts, benefits and opportunities. Wildl Res 42:541–553. https://doi.org/10.1071/WR14229
Stara K, Sidiropoulos L, Tsiakiris R (2016) Bound eagles, evil vultures and cuckoo horses. Preserving the Bio-cultural Diversity of Carrion Eating Birds. Hum Ecol 44:751–764. https://doi.org/10.1007/s10745-016-9864-3
Sumasgutner P, Nemeth E, Tebb G et al (2014) Hard times in the city – attractive nest sites but insufficient food supply lead to low reproduction rates in a bird of prey. Front Zool 11:48. https://doi.org/10.1186/1742-9994-11-48
Suri J, Sumasgutner P, Hellard É et al (2017) Stability in prey abundance may buffer black sparrowhawks Accipiter melanoleucus from health impacts of urbanization. Ibis 159:38–54. https://doi.org/10.1111/ibi.12422
Teta P, Hercolini C, Cueto G (2012) Variation in the diet of western barn owls (Tyto alba) along an urban-rural gradient. Wilson J Ornithol 124:589–596. https://doi.org/10.1676/11-173.1
Thomas T, Aggar C, Baker J et al (2022) Social prescribing of nature therapy for adults with mental illness living in the community: a scoping review of peer-reviewed international evidence. Front Psychol. https://doi.org/10.3389/fpsyg.2022.1041675
Thompson J, Shirreffs L, McPhail I (2003) Dingoes on Fraser Island—Tourism dream or management nightmare. Hum Dimens Wildl 8:37–47. https://doi.org/10.1080/10871200390180136
Thomson VK, Stevens T, Jones D, Huijbers C (2016) Carrion Preference in Australian Coastal raptors: effects of Urbanisation on Scavenging. Sunbird 46:16–28
Travaini A, Donázar JA, Ceballos O, Hiraldo F (2001) Food habits of the Crested Caracara (Caracara plancus) in the Andean Patagonia: the role of breeding constraints. J Arid Environ 48:211–219. https://doi.org/10.1006/jare.2000.0745
United Nations Development Programme (2022) Human Development Report 2021-22: Uncertain Times, unsettled lives: shaping our future in a transforming world. United Nations, New York
University of New Hampshire (2019) What are the best ways to protect my chickens from predators? In: Extension. https://extension.unh.edu/blog/2019/02/what-are-best-ways-protect-my-chickens-predators. Accessed 21 Jun 2024
van Rensburg BJ, Peacock DS, Robertson MP (2009) Biotic homogenization and alien bird species along an urban gradient in South Africa. Landsc Urban Plan 92:233–241. https://doi.org/10.1016/j.landurbplan.2009.05.002
Vidal A, Baldomà L, Molina-López RA et al (2017) Microbiological diagnosis and antimicrobial sensitivity profiles in diseased free-living raptors. Avian Pathol 46:442–450. https://doi.org/10.1080/03079457.2017.1304529
Washburn BE (2014) Human-Osprey conflicts: industry, utilities, communication, and transportation. J Raptor Res 48:387–395. https://doi.org/10.3356/JRR-OSPR-13-04.1
Washburn BE, Begier MJ, Wright SE (2015) Collisions between eagles and aircraft: an increasing problem in the airport environment. J Raptor Res 49:192–200. https://doi.org/10.3356/rapt-49-02-192-200.1
Welti N, Scherler P, Grüebler MU (2020) Carcass predictability but not domestic pet introduction affects functional response of scavenger assemblage in urbanized habitats. Funct Ecol 34:265–275. https://doi.org/10.1111/1365-2435.13469
Wenny DG, Devault TL, Johnson MD et al (2011) The need to quantify ecosystem services provided by birds. Auk 128:1–14
Whelan CJ, Wenny DG, Marquis RJ (2008) Ecosystem services provided by birds. Ann N Y Acad Sci 1134:25–60. https://doi.org/10.1196/annals.1439.003
White J, Kemmelmeier M, Bassett S, Smith J (2018) Human perceptions of an avian predator in an urban ecosystem: close proximity to nests increases fondness among local residents. Urban Ecosyst 21:271–280. https://doi.org/10.1007/s11252-017-0713-y
Winkler DW, Billerman SM, Lovette IJ (2020a) Hawks, Eagles, and Kites (Accipitridae), version 1.0. Birds of the World. https://doi.org/10.2173/bow.accipi1.01species_shared.bow.project_name
Winkler DW, Billerman SM, Lovette IJ (2020b) Barn-Owls (Tytonidae), version 1.0. Birds of the World. https://doi.org/10.2173/bow.tytoni1.01species_shared.bow.project_name
Winkler DW, Billerman SM, Lovette IJ (2020c) Owls (Strigidae), version 1.0. Birds of the World. https://doi.org/10.2173/bow.strigi1.01species_shared.bow.project_name
Winkler DW, Billerman SM, Lovette IJ (2020d) New World Vultures (Cathartidae), version 1.0. Birds of the World. https://doi.org/10.2173/bow.cathar2.01species_shared.bow.project_name
Witmer GW, Pipas M, Burke P et al (2008) Raptor use of artificial perches at natural areas, city raptor use of artificial perches at natural areas, city of fort collins, colorado of fort collins, colorado. Prairie Naturalist 40:38–41
World Bank (2018) What a Waste: An Updated Look into the Future of Solid Waste Management. In: World Bank. https://www.worldbank.org/en/news/immersive-story/2018/09/20/what-a-waste-an-updated-look-into-the-future-of-solid-waste-management. Accessed 10 May 2024
Zhao Q, Wang Y, Wu L et al (2024) A path to human-raccoon dog harmony: identifying factors influencing the tolerance of urban residents in Shanghai towards a neglected species. People Nat 6:1277–1287. https://doi.org/10.1002/pan3.10636
Acknowledgements
The authors would like to acknowledge to the Facultad de Ciencias Exactas y Naturales, Universidad de Mar del Plata, Argentina and Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) for the financial supported. We are grateful for all the constructive comments from the editors and reviewers.
Funding
This study was funded by the Facultad de Ciencias Exactas y Naturales of the Universidad Nacional de Mar del Plata (grant to Laboratorio Vertebrados EXA1044/21). Author Eugenia A. Bonetti has received a doctoral scholarship from Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) (EX-2022-57152941).
Author information
Authors and Affiliations
Contributions
Conception and design of the study were made by E.B, J.I and L.B. Literature search and data analysis were performed by E.B. The first draft of the manuscript was written by E.B and JJ.I, C.P, MB and L.B commented on previous versions of the final manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Bonetti, E.A., Isacch, J.P., Paterlini, C.A. et al. Global review of services and conflicts provided by raptors in urbanized habitats. Urban Ecosyst (2024). https://doi.org/10.1007/s11252-024-01579-1
Accepted:
Published:
DOI: https://doi.org/10.1007/s11252-024-01579-1