
Abstract
Given that rates of urbanization are rising globally, the need to maintain and enhance urban biodiversity has become important. This study examines the relative influence of different land-use types as well as environmental resources (trees, flowering trees, fruiting trees, shrubs, telecommunication mast, pylons, electric poles, buildings) on bird diversity indicators in a rapidly developing urban settlement in Ghana. Remote sensing was used to estimate the extent of conversion of natural habitats into urban settlements. Using point count surveys, bird species were recorded and compared in randomly selected plots of four land-use types of farmlands, remnant forest, residential and commercial areas. We further evaluated the relative influence of habitat resources on bird diversity indicators. We found a significant expansion of built-up areas into natural habitats in the study area. Also, there has been a significant increase in sparse vegetation coupled with a drop in the area covered by dense vegetation over the last three decades. Avifauna diversity indicators differ significantly across the four land-use types with urban farmlands being the most species diverse, followed by urban remnant forests, then residential and finally commercial areas. Our findings suggest that avian species diversity indicators decreased significantly with increasing land-use intensity and revealed that the study area still possesses significant conservation potentials for urban birds and by extension biological diversity as long as vegetation fragments are maintained within a sustained urban expansion framework. We propose increasing the quantity and complexity of the vegetation cover in residential areas by supporting citizens to establish private yards to increase the city’s green network.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The global human population currently stands at about 7.9 billion people and is predicted to increase by 1.05% per annum (United Nations, 2021). In relation to this, it has been projected that by 2030, more than 60% of the human population will live in urban areas and that most of this increase will occur in developing nations (United Nations, 2021). Consequently, human population distributions in time and space will become more unevenly distributed with a concentration in the urban areas (Leveau and Leveau 2012; United Nations, 2021). This increase in human populations and distribution could present several challenges to biodiversity as there would be a proportional increase in demand for natural resources to support this population (Ganaie et al. 2020; Guan et al. 2020). This suggests that habitats for biodiversity will be destroyed as urban and its suburbs continue to expand into natural habitats (Leveau and Leveau 2016). Therefore, understanding how biodiversity within urban areas would respond to the changes in their environment is crucial in the conservation planning and management of urban land.
There is evidence that the degree of urban development and type of urban land use can change the composition and distribution of animal communities including birds (Clergeau et al. 2006; Stratford and Robinson 2005). Generally, studies have shown that urbanization causes a reduction in the richness and relative abundance of species for most biodiversity (Callaghan et al. 2021; Guenat et al., 2019; Leveau 2021; Piano et al. 2020). Many studies on birds in urban settlements reported declining trends in the population of species and an increase in the risks of extinction as urbanization increases (Chamberlain et al. 2017; Dri et al. 2021; Leveau 2019). This is so because urban expansion results in the conversion of forests, grasslands, pastures, sometimes wetlands, and water bodies into built environments (Solecka et al. 2017; Vasenev et al. 2019). These converted habitats consequently become hostile and uninhabitable to most original native species (Xu et al. 2018). However, the biodiversity that does occur in urban areas contribute significantly to the quality of life of the human population such as the ecosystem services they provide (Wu 2014), the psychological well-being of people (Cheke and Tratalos 2007; Tratalos et al. 2007), educational (Dearborn and Kark 2010), and economic values [e.g., property prices are increased in higher biodiversity areas (Hope et al. 2008)].
Given that rates of urbanization are rising globally, the need to maintain and enhance urban biodiversity has become increasingly important. Although land conversion pressure and species response to development have been well studied in the developed world (Schwarz et al. 2017), little is known about the dynamics of populations and decline trends, especially for birds in tropical Africa (Leveau et al. 2021).
Studies on land conversion pressure, urbanization trends, and urbanization influences on urban bird communities in developing economies particularly sub-Saharan Africa are uncommon (Lepczyk et al. 2017; Leveau et al. 2021). Although one of the most important changes on global environmental change is population growth that leads to land cover/land use changes which in turn leads to the conversion of natural environment into built environment, there are no or few established guidelines for the acquisition and development of lands in many parts of Africa. In Ghana, like many countries in sub-Saharan Africa, ownership of land is vested in the custody of heads of clans and traditional chiefs and not the central government; hence, laws governing land acquisition and development are rather weak or not implemented. Not many people acquire the relevant permit before they start construction of building of any form (Agbosu 2000; Mersha et al. 2021). As a result, vast areas of forests are lost daily and unexpectedly to unregulated farming, mining, and infrastructural development (Boon et al. 2009). In these circumstances, wildlife occupying such habitats lose their original habitats overnight, and either they leave or adapt (Lowry et al. 2013). The impact of this overnight loss of habitat becomes severer on habitat-sensitive species which may become vulnerable (Morante-Filho et al. 2015) to higher risks of population declines and eventually local extinctions (Brewster et al. 2018). It is therefore imperative that conservation and urban management efforts are directed towards understanding faunal diversity and assemblage structure within highly unpredictable urban land-use types. This knowledge, if available can assist in finding ways to mitigate the negative impacts of fluctuations in the urban landscape for animals whiles providing the advantage of harnessing potential opportunities for the conservation of biodiversity in urban settlements in Africa.
This study examines land-use change on urban bird diversity indicators as well as evaluates how environmental resources influence avifauna in a rapidly developing urban settlement in the Cape Coast Metropolitan Assembly (CCMA) in Ghana. To achieve this, avifauna abundance, diversity, and richness (diversity indicators) were surveyed within four different land-use types (as major predictors) and variations observed in these diversity indicators (response variables) were modelled as a function of variation in environmental resources (explanatory covariates) in the study area. We hypothesized that natural land-use types (urban remnant forests and farmlands) would have higher bird diversity estimates than built-up environments and that natural native vegetation covariates would positively influence avifauna assemblage structure in the study area.
Methodology
Study area
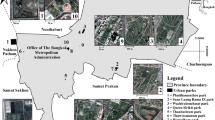
Cape Coast Metropolitan Assembly (5°6′23.4″N 1°14′29.04″W) in the Central Region of Ghana (one of the 16 administrative regions in Ghana) (Fig. 1.), covers a land area of approximately 122 km2, with a settlement population of about 189,925 people and an annual population growth rate of 1.0% (Ghana Statistical Service 2021).

Map of the study area (Cape Coast Metropolitan Assembly) showing the distribution of study sites and land use types
Cape Coast Metropolitan Assembly is predominantly urban with three-quarters of the population residing in urban areas. Located within the Guinea-Congo vegetation zone of West Africa, CCMA has double maxima rainfall pattern ranging from 750 to 1000 mm, with the major wet season from May to July (Ghana Meteorological Agency, 2021). It is a humid area with monthly relative humidity varying from 85 to 99%. The present vegetation of the metropolis consists of shrubs, grasslands, native forest fragments, and coastal thickets. The common trees in the forest fragments and coastal thickets include African mahogany (Khaya ivorensis), silk cotton tree (Ceiba pentandra), oil palm trees (Elaeis guineensis), and other species of the Family Arecaceae. The original vegetation of dense shrubs, which the rainfall supported, has been replaced by sparse secondary vegetation because of clearing for farming, charcoal burning, and other human activities. Elephant grass (Pennisetum purpureum), Guinea grass (Panicum maxima), Centro (Centrosema pubescens), the invasive weed siam weed (Chromolaena odorata) and many more weedy species dominate the secondary vegetation. The core of the metropolis is built-up and is a mosaic of residential areas, car parks, open markets and offices of corporate bodies. Interspersed among these is vegetation which consists of shrubs and, grasses, native forest fragments and coastal thickets. Other areas such as farmlands are restricted to the periphery with active farmlands dominated by maize (Zea mays), plantain (Musa paradisiaca), cocoyam (Xanthosoma sagittifolium), tomatoes (Solanum lycopersicum) and cassava (Manihot esculenta). Abandoned farmlands are dominated by shrubs and small trees that are occasionally cut down for charcoal production. Green spaces, usually remnant forests are restricted within the campuses of two universities and a number of second-cycle schools. These remnant forests also retain some of the original vegetation of the metropolis (Deikumah and Kudom 2010).
Satellite data, image processing, classification, and analysis
Landsat satellite data (20% Cloud Free) of four dates in the past three decades were downloaded from United States Geological Survey (USGS) Earth Explorer website. All the data were pre-processed and projected to the Universal Transverse Mercator (UTM) projection system. The satellite data collected are presented in Appendix A.
ENVI (Excelis) version 5.3 and ArcGIS version 10.6 were used for the image processing. Image classification involves the assignment of pixels to different classes of land use and land cover identified within the CCMA. Five classes of land cover were pre-determined with consideration to area falling within the following; dense vegetation (forest reserves and urban remnant forests), sparse vegetation (shrub, grassland, farmland), built-up (including residential and commercial), and water (rivers, streams, lakes, lagoons, wetlands).
A supervised classification method with Support Vector Machine algorithm was adopted for the land use/cover classification using the ENVI 5. 3 software. Support Vector Machine makes use of a user-defined kernel function to plot a set of non-linear decision boundaries in the original dataset into linear boundaries of a higher-dimension construct. The algorithm used by the Support Vector Machine Classification tool is based on statistical learning theory, which determines a hyperplane that optimally separates two classes. Training data is used to determine the optimum hyperplane and generalize its ability to verify using validation data (Han et al. 2007).
Experimental design and land-use classification
Based on the dominant vegetation, land use, and intensity, three each of the four main land-use types within the study area were selected for this study. These include commercial areas (marketplaces, lorry parks, and areas with a high concentration of shops); residential areas (organized communities and settlements), farmlands (areas dominated by croplands and farm bushes), and urban remnant forest (thickets with less evidence of active human activities) in CCMA. The size of the land-use types ranges from 1.88 to 2.01 km2 while the study sites were located at least 1.5 km apart. Forty-five grid points, 200 m apart were generated using QGIS version 3.14.1 and distributed among the three replicates of each land-use type. This was to ensure the independence of sample points within each site and reduce the chance of double-counting of birds. The points were exported onto Global Positioning System (GPS) device, Garmin eTrex 10® and located on the field.
Avifauna survey
To document the avifaunal composition of CCMA, a 50 m fixed-radius point count survey was conducted twice from January to February and June to July. These two periods were chosen for bird surveys for the understanding of the influence of seasons on bird assemblage structure and also seasonal resource variations that support birds in the study area.
Bird surveys were conducted twice daily from 06:00 h to 10:00 h and from 15:00 h to 18:00 h each day to coincide with periods when birds are most active. During each survey, 10 min was spent at the point count station to count birds. Using a pair of binoculars (Nature-Trek 8 × 42) and Borrow and Demey (2014) bird species and the number of individuals seen or heard were recorded.
In addition, all birds encountered opportunistically during other field research within the survey were documented. Data from these encounters were not used in the final analyses but were used to build the species list for the CCMA.
Vegetation survey
Natural vegetation for this study is defined as photosynthetic plants that developed with little or no human interference (Ramankutty et al. 2010). It includes artificially created green plants and exotic plant species. Details of natural vegetation that potentially influence bird assemblage structure quantified in this study are presented in Table 1.
A 30 × 30 m quadrat was placed randomly at each sampling point and the number of small trees, flowering trees, and large trees as well as estimated percentage vegetation cover was recorded. Shrubs were counted within a 10 × 10 m quadrat within the area. Percentage vegetation cover representing the total area within the 30 × 30 m quadrat that is covered by vegetation be it lawns, grasses, herbs, or any natural vegetation was estimated by eye to the nearest 5% (Manu 2003). Trees with a diameter at breast height (DBH) greater than 40 cm were classified as large trees, and tress with DBH less than 40 cm were classified as small tress while shrubs were classified as plants having multiple stems emanating from the root level (Hawthorne and Lawrence 2013). The presence or absence of crops or gardens within each sampling point was also recorded.
Artificial resource surveys
Artificial resources for this study are resources that are not natural but are potential resources for birds in the absence of natural vegetation in the urban setting. Within each sampling point (50 m radius), the number of telecom masts, electric poles, and buildings were recorded. The presence or absence of paved roads, and refuse dumps were also recorded and used for further analyses.
Data and statistical analysis
Land use/cover changes detection and analysis
Post-classification comparison change detection was used to compare changes in land use/cover within the CCMA. The post-classification approach was used to compare land classification results of the four images from the comparative years on a pixel-by-pixel basis. Using this approach, a resultant change detection matrix was used to extract the areas of change and the magnitude in each land cover type. We further used linear regression to test for significant changes in the observed land cover changes.
Avifauna data analysis
Overall species richness (i.e., the number of individual species seen during the survey) and the total number of individuals of all species representing bird abundance were computed for each study site and land-use type. Avian species diversity was calculated using the Shannon-Wiener index from the vegan package (Oksanen et al. 2007). Species accumulation curve was used to illustrate the rate of species accumulation among land-use types and within CCMA. Shapiro Wilks test was used to check for normality of the response variables (i.e., abundance, richness, and diversity). The response variables at the site level were found to be normally distributed (Shapiro-Wilk normality test W = 0.91, p-value = 0.06) hence Analysis of Variance (ANOVA) was used to test for variations in mean abundance, richness, and diversity of birds among the four land-use types and between seasons. This was followed by a post hoc test using a Tukey-HSD test at 95% confidence level for multiple comparisons of means of the avian diversity indicators across land-use types. Similarly, ANOVA was used to test for variations in quantified environmental variables (both natural and artificial) across land-use types. A multi-collinearity test was conducted on all explanatory variables to eliminate strongly correlated variables. Explanatory variables were taken as strongly correlated when Pearson’s correlation coefficient exceeds 50% (i.e., r > ± 0.50). Only one of the two correlated variables was maintained in the final model based on its ecological importance and potential influence on the relevant response variable.
Over-dispersion was detected in the data hence, a General Linear Model (GLM) with the quasi-Poisson family was used to model the relationship between the response variables and the explanatory variables. The main predictors were land-use type and season while other covariates were small trees, large trees, flowering trees, shrubs, percentage vegetation cover, telecommunication mast/pylons, electric poles, number of buildings, presence or absence of a paved road, crops, and refuse. Using stepwise deletion, explanatory variables that had no significant relationship with the response variables were dropped from the model. Model residual plots from regression models were used to check whether the model assumptions were met using the function check_model(model) in the “performance” package. We then ranked the explanatory variables according to their relative influence on each of the response variables using their parameter estimates. Spatial autocorrelation was tested on the sampling points using a spline correlogram. The sampling points were found not to be spatially correlated. All analyses were done in R statistical software 4.2.1 (R Core Team, 2022).
Results
Trends in land use/cover change in CCMA over the last three decades
Land cover within CCMA has changed significant (F4,12= 35.01, p < 0.001). Built-up areas have increased significantly from 1990 to 2020 in CCMA (Table 2). Dense vegetation cover within CCMA decreased significantly by 38% as a result of the increase in areas now covered with buildings. The land area covered by water in 1990 has also reduced by 0.39% while areas with sparse vegetation have increased significantly by 24% over the last three decades (Table 2; Fig. 2.)

Land Use and Land Cover change in Cape Coast Metropolitan Assembly between 1990 to 2020
Overview of bird surveys
The species accumulation curves approached an asymptote for the entire study area (Appendix B, Fig. 1a.), as well as for individual land-use types (Appendix B, Fig. 1b) suggesting that each study landscape was adequately surveyed and most species in the study area were detected. Overall, 8,583 individual birds comprising 154 species from 52 families were recorded within the four land-use types in Cape Coast Metropolitan Assembly (Supplementary information). These include 127 common residents, 18 intra-African migrants, and nine Palearctic migrants. The highest number of individuals, 1160 representing 13.5% were recorded from the family Ploceidae, 1016 (11.8%) from the Estrildidae, and 838 (9.8%) were from the Pycnonotidae family. Family Scolopacidae and Sylviidae had the lowest record of only one individual each.
Variations in bird diversity indices within land-use types
Species diversity
Mean (Standard Error) diversity for the entire survey was estimated as 3.28 (0.11). Species diversity differed significantly across land-use types (F3,19= 32.01, p < 0.001) but not between seasons (F1,19 = 2.87, p = 0.10). Comparatively, farmland was the most species diverse with mean species diversity of 3.69 (0.13) while commercial land-use type had the lowest with a mean of 2.82 (0.08) (Fig. 3a). However, a Tukey HSD test shows that commercial centres and residential areas differ significantly from urban remnant forests while commercial and residential land use types differ significantly from farmlands.

Variation in species diversity indicators within land-use types in the Cape Coast Metropolitan Assembly. Similar alphabets on standard error bars do not differ significantly
Species abundance
Mean (SE) abundance in the CCMA was estimated at 715 (91). Comparatively, more birds were encountered on farmlands 1065 (243) with commercial areas having the least number of individuals 506 (82) (Fig. 3b). The results of analyses of variance revealed significant variations in avifaunal abundance across land-use types (F3,19 = 5.35, p < 0.01) but not between seasons (F1,19 = 2.8, p = 0.11). However, a Tukey-HSD test shows that the significant variation lies between farmland-commercial and residential-farmland.
Species richness
Overall mean (SE) species richness was estimated as 58 (6) and varied significantly across land-use types in the CCMA (F3,19 = 21.96, p < 0.01) but not between season (F1,19 = 1.58, p = 0.22), with farmlands being the most species-rich land use type with a mean value of 82 (6) and commercial centres the least with a mean value of 37 (4) (Fig. 3c). A Tukey-HSD test shows that commercial and residential land use differ significantly with farmlands while urban remnant forests differ significantly with commercial and residential land use.
Landscape and site scale variations in environmental variables in the CCMA
Estimated environmental resources comprising both natural stand vegetation covariates and artificial support for birds in the study landscape differed significantly across land-use types (Appendix C). Number of small trees, large trees, and flowering plants were higher in urban remnant forests as predicted with mean SE values of 5.33 (0.32), 8.62 (0.33), and 1.16 (0.12) respectively, while shrub density and percentage vegetation cover were highest in farmlands with mean values of 8.92 (0.33) and 77.61% (2.14) respectively. Number of buildings, electric poles, and telecommunication masts were more in commercial centres with mean values of 6.06 (0.17), 5.55 (0.32), and 0.22 (0.05) respectively. Only number of flowering trees (t =-5.96, df = 287.04, p < 0.001) and fruiting trees (t =-4.04, df = 287.37, p < 0.001) were found to vary significantly between seasons.
The results of a multi-collinearity test suggested little collinearity among variables (Appendix D). Although percentage vegetation cover and number of buildings were negatively correlated (r = -0.86), they were both used in the final model because the presence of buildings was treated as an artificial resource while vegetation cover as a natural variable, and also, they influenced species diversity indicators differently.
Relationship between environmental variables and species diversity indicators in CCMA
Generally, the main predictors (i.e., land use type and season) and environmental covariates (i.e., presence of crops, flowering trees, number of buildings, and the interaction between buildings and vegetation cover) showed a significant relationship with species diversity (F10, 349 = 29.92, p < 0.001). The presence of crops and flowering trees showed a significantly positive relationship with species diversity. Although buildings showed a significant negative relationship with species diversity, an interaction between buildings and vegetation cover showed a significant positive relationship with species diversity (Appendix E).
Similarly, the main predictors, presence of crops, large trees, flowering trees, and the interaction between buildings and vegetation cover showed a significant relationship with species abundance (F12, 347 = 13,489, R2 = 0.99, p < 0.001). Species abundance was found to have a positive significant relationship with flowering trees and the presence of crop or garden within the landscape as well as the interaction between buildings and vegetation cover (Appendix E). Large trees, number of buildings, and percentage vegetation cover had a significant negative relationship with species abundance. An increase in flowering trees and the presence of crops or gardens increases bird species abundance, while an increase in large trees, buildings, and percentage vegetation cover results in a decrease in species abundance.
Furthermore, the main predictors and presence of crops, large trees, flowering trees, and the interaction between the number of buildings and vegetation cover showed a significant but varying relationship with species richness (F10, 349 = 3874, R2 = 0.99, p < 0.001). Flowering trees and the presence of crops or gardens in an area as well as the interaction between buildings and vegetation cover were found to have a positive significant relationship with species richness (Appendix E). An increase in these covariates increases bird species richness. While large trees, number of buildings, and percentage vegetation cover had a significant negative relationship with species richness.
Generally, from the parameter estimates of all final models, crops had the highest positive significant effect on all species diversity indicators while the interaction between buildings and percentage vegetation cover had the least positive influence within the CCMA (Fig. 4.). Number of buildings and large trees had the highest and lowest significant negative influence on species diversity indicators respectively.

Rank of final model parameter estimates, effect size, and direction of significant predictors (covariates) on avian diversity indicator (response variables). *CP = crop, BD = buildings, VC = percentage vegetation cover, FT = flowering trees, LT = large trees
Relationship between environmental variables and species diversity indicators within land-use types
In commercial areas, farmlands, and residential areas, only percentage vegetation cover, presence of crops, and flowering trees were found to have a significant positive influence on species diversity respectively (Table 3). Within remnant forests, flowering trees were found to have a significant positive relationship with species diversity while electric poles were found to have a significant negative relationship with species diversity. Only flowering trees had a positive significant relationship with species diversity within residential areas (Table 3).
Similarly, vegetation cover and the presence of crops had a significant positive relationship with species abundance within commercial areas while large trees had a significant negative relationship with species abundance within farmlands. Within urban remnant forests, flowering trees were found to have a significant positive effect on bird abundance while vegetation cover had a negative effect on bird abundance. Flowering trees and vegetation cover were all found to have a positive influence on bird abundance in residential areas (Table 3).
Percentage vegetation cover had a significant positive effect on species richness within commercial areas. Within farmlands number of flowering plants and the presence of crops had a significant positive influence on species richness while an increase in large trees caused a significant decrease in the species richness. Flowering trees also positively influenced bird species richness while percentage vegetation cover within urban remnant forests significantly influenced species richness negatively. Within residential areas, flowering trees showed a significant positive effect on species richness (Table 3).
Discussion
Landuse/land cover changes within CCMA
Our results from the land cover change analysis provide concrete evidence of extensive land cover temporal changes within CCMA. Built-up areas and sparse vegetation have increased, resulting in a decrease in areas with dense vegetation and water cover. This finding supports findings by Asabere et al. (2020) and Ashiagbor et al. (2019) who found similar trends in other two urban areas in the country. The observed increase in built-up area and sparse vegetation could be attributed to a growing human population. CCMA had a settlement population of 170,000 people and a growth rate of 3.5% per annum in 2010 according to the 2010 population and housing census (Ghana Statistical Service, 2013). However, our physical observation and remote sensing data as demonstrated in the current study revealed extensive expansion of human settlements as new lush infrastructure is located deeper into areas that were previously covered by forests and natural vegetation. Also, the growing demand for agricultural products for this increasing human population has intensified the rate of natural terrestrial land being shifted to agricultural lands mostly at the periphery of the CCMA which may have contributed to the increase in sparse vegetation as revealed in earlier studies in other regions (Pérez-Hernández and Gavilán 2021).
CCMA is characterized by a mosaic landscape with urban remnant forests found within the enclave of senior high schools and university campuses with a minimal level of disturbance to wildlife. This landscape supports a considerable diversity of avifauna including a globally threatened species the Hooded vulture Necrosyrtes monachus that were recorded in all the land use types. These findings confirm earlier studies (e.g., Demeyrier et al. 2016; Mason 2000; Stratford and Robinson 2005), that revealed that urban landscapes support biodiversity particularly avifauna populations including species of conservation concern.
Variations in bird species diversity measures within land-use types
Although not surprising, this study revealed that diversity indicators decreased with increasing land-use intensity offering an insight into the distribution of habitat resources available to birds within the study landscape while suggesting that farmlands and urban forest reserves offer the widest spectrum of resources for birds compared to residential and commercial areas. This finding is consistent with those of Millard et at., (2021) who found that increasing land-use intensity from minimal to intense use was associated with a significant change in pollinators (i.e., reductions in overall pollinator biodiversity at high land-use intensity).
Our results show that farmlands recorded the highest number of species and individuals which confirms the intermediate disturbance hypothesis (IDH). IDS states that moderate disturbances in a habitat can create a mosaic of microhabitats that support more species than the extremes of the disturbances (Shea et al. 2004; Wilkinson 1999). Consequently, fewer species were recorded in residential and commercial areas which are next to farmlands in terms of disturbance intensity. This result corroborates that of earlier studies by Newbold et al. (2013) and de Lima et al. (2013) who found a substantial global impact of land-use intensity on the local abundance of bird species and with the results of several previous studies on a wide range of species (Fontana et al. 2011; Hof et al. 2011; MacGregor-Fors et al. 2010).
Surprisingly, the forest habitat recorded the second-highest species diversity indicators which deviated from the general known trend that an area with more diverse trees will have more resources for birds, hence more bird species (Jakobsson and Lindborg 2017). However, the remnant forest within the CCMA has received high levels of degradation from hunters, firewood collectors, illegal chain saw activities, and it also serves as an illegal refuse dumping site for some people. These activities may have depleted the resources for birds, hence habitat-sensitive species may have found the place uninhabitable. Also, the remnant forest has received further encroachment rendering the sizes too small to support biodiversity. This could be the reason for relatively few species recorded in this habitat. The farmlands retain a high proportion of shrubs, crops, and few large and small trees. These vegetation parameters have been found to contribute positively to the abundance, richness, and diversity of birds (MacGregor-Fors 2008).
Relationships between environmental resources and variation in avian diversity indicators
Generally, the presence of crops and flowering trees showed a significant positive relationship with species diversity within CCMA. An increase in the flowering trees and the presence of crops could translate into the availability of nectar, pollen, and other foraging materials for farmland birds and insects. This result is not surprising, since vegetation variables have been positively related to bird species diversity in the past by MacGregor-Fors (2008). These vegetation resources offer various benefits to birds, such as foraging resources, cover, and nesting sites (Melles et al. 2003). Percentage vegetation cover and large trees had a significant negative relationship with species diversity indicators. This finding, however, contradicts earlier research by Kebrle et al. (2021) who found large trees as a key factor for bird diversity as well as a study by Rico-Silva et al. (2021) who found native bird richness and abundance to be positively influenced by woody vegetation cover. However, our finding can be attributed to the fact that vegetation cover within the study area was mostly lawns that are managed regularly through weeding and occasionally spraying with herbicides. This may have reduced available food for specialist species and hence provided resources for only a few categories of birds like the Cattle Egret Bubulcus ibis that was constantly found foraging for arthropods in the lawns. Also, the present vegetation of CCMA is a coastal thicket, which supports mostly savanna birds and few forest specialists that uses large trees more often for roosting and nesting.
The number of buildings had a significant negative relationship with species diversity indicators. This result confirms findings in previous studies by Rodrigues et al. (2018) who found a negative effect of buildings on both species richness and abundance. In CCMA, the density of buildings appears to be related to urbanization and is a good indicator of the level of urbanization. However, the interaction between buildings and vegetation cover showed a significant positive relationship with all three species diversity indicators. This findings agrees with findings by Chamberlain et al. (2019), whose model suggested that maintaining green space in at least an equal proportion to the built environment is likely enhance urban biodiversity in developing countries. This suggests that bird diversity would be enhanced within the study area as long as vegetation fragments are maintained within areas with buildings.
Implications for bird conservation, urban planning, and management
Land use/land cover changes have been widely discussed in landscape and ecology studies, and the implementation of these research findings in landscape planning and management would contribute to decreasing biodiversity loss and maintaining ecosystem function in increasingly fragmented landscapes (Bregman et al. 2014). The CCMA is expanding into habitats for wildlife with inadequate consideration of urban biodiversity management. Land acquisition and development are poorly regulated. As a result, vast areas of forest are being lost daily to unregulated farming and development (Boon et al. 2009), leading to loss of biodiversity overnight. By considering this we propose different biodiversity conservation strategies with this study. First, we propose that the CCMA urban planning policies should adopt an environmental sustainability framework that focuses on regulating land clearing within the Metropolis. This policy should also focus on increasing the quantity and complexity of the vegetation cover in residential areas by establishing urban green networks. This could be in the form of supporting citizens in maintaining residential vegetation (e.g., private yards), this would increase the city’s green areas and promote biodiversity conservation. Also, urban area management and planning activities should continuously be evaluated to measure their effectiveness and amended where necessary. We recommend monitoring the environmental quality of cities using birds as bio-indicators. Such monitoring programs should use a volunteer-based project, such as the bird atlas project. These urban management and planning activities could assist in the maintenance and promotion of bird species richness in urban areas, as well as increasing the environmental quality of the urban systems we live in.
Code availability
The datasets generated during and/or analysed during the current study are not publicly available but are available from the corresponding author on reasonable request.
References
Agbosu LK (2000) Land law in Ghana: Contradiction between Anglo-American and customary conceptions of tenure and practices. University of Wisconsin, Madison
Asabere SB, Acheampong RA, Ashiagbor G, Beckers SC, Keck M, Erasmi S, Schanze J, Sauer D (2020) Urbanization, land use transformation and spatio-environmental impacts: Analyses of trends and implications in major metropolitan regions of Ghana. Land Use Policy 96:104707
Ashiagbor G, Amoako C, Asabere SB, Quaye-Ballard JA (2019) Landscape transformations in rapidly developing peri-urban areas of Accra, Ghana: Results of 30 years. Open Geosci 11(1):172–182
Boon E, Ahenkan A, Baduon BN (2009) An assessment of forest resources policy and management in Ghana. Proceedings of the 29th Annual Conference of the International Association for Impact Assessment, Accra, Ghana, 16–22
Borrow N, Demey R (2014) Birds of western Africa, Second edn. Christopher Helm
Bregman TP, Sekercioglu CH, Tobias JA (2014) Global patterns and predictors of bird species responses to forest fragmentation: implications for ecosystem function and conservation. Biol Conserv 169:372–383
Brewster CL, Beaupre SJ, Willson JD (2018) Habitat loss and local extinction: linking population declines of eastern collared lizards (Crotaphytus collaris) to habitat degradation in Ozark glades. J Herpetology 52(3):352–360
Callaghan CT, Liu G, Mitchell BA, Poore AGB, Rowley JJL (2021) Urbanization negatively impacts frog diversity at continental, regional, and local scales. Basic Appl Ecol 54:64–74
Chamberlain DE, Henry DAW, Reynolds C, Caprio E, Amar A (2019) The relationship between wealth and biodiversity: A test of the Luxury Effect on bird species richness in the developing world. Glob Change Biol 25(9):3045–3055
Chamberlain DE, Kibuule M, Skeen R, Pomeroy D (2017) Trends in bird species richness, abundance and biomass along a tropical urbanization gradient. Urban Ecosyst 20(3):629–638
Cheke RA, Tratalos JA (2007) Migration, patchiness, and population processes illustrated by two migrant pests. Bioscience 57(2):145–154
Clergeau P, Jokimaki J, Snep R (2006) Using hierarchical levels for urban ecology. Trends in Ecology and Evolution 21:660–661
de Lima RF, Dallimer M, Atkinson PW, Barlow J (2013) Biodiversity and land-use change: Understanding the complex responses of an endemic‐rich bird assemblage. Divers Distrib 19(4):411–422
Dearborn DC, Kark S (2010) Motivations for conserving urban biodiversity. Conserv Biol 24(2):432–440
Deikumah JP, Kudom AA(2010) Biodiversity status of urban remnant forests in Cape Coast, Ghana.Journal of Science and Technology (Ghana), 30(3)
Demeyrier V, Lambrechts MM, Perret P, Grégoire A (2016) Experimental demonstration of an ecological trap for a wild bird in a human-transformed environment. Anim Behav 118:181–190
Dri GF, Fontana CS, de Sales Dambros C (2021) Estimating the impacts of habitat loss induced by urbanization on bird local extinctions. Biol Conserv 256:109064
Fontana S, Sattler T, Bontadina F, Moretti M (2011) How to manage the urban green to improve bird diversity and community structure. Landsc Urban Plann 101(3):278–285
Ganaie TA, Jamal S, Ahmad WS (2020) Changing land use/land cover patterns and growing human population in Wular catchment of Kashmir Valley, India. GeoJournal, 1–18
Ghana Meteorological Agency (2021) Ghana Meteorological Agency’s Seasonal Forecast for the Northern Sector and Seasonal Forecast Update for Southern Sector of Ghana
Ghana Statistical Service (2021) Ghana Population and housing census, General report, Cape Coast municipality. Ghana Statistical Service, Accra, Ghana
Ghana Statistical Service (2013) 2010 population and housing census district analytical report, Cape Coast municipality
Guan J, Kirikkaleli D, Bibi A, Zhang W (2020) Natural resources rents nexus with financial development in the presence of globalization: is the “resource curse” exist or myth? Resour Policy 66:101641
Guenat S, Kunin WE, Dougill AJ, Dallimer M (2019) Effects of urbanisation and management practices on pollinators in tropical Africa. Journal of Applied Ecology, 56(1), 214–224.
Han D, Chan L, Zhu N (2007) Flood forecasting using support vector machines. J Hydroinformatics 9(4):267–276
Hawthorne W, Lawrence A(2013) Plant identification: creating user-friendly field guides for biodiversity management. Routledge
Hof C, Araújo MB, Jetz W, Rahbek C (2011) Additive threats from pathogens, climate, and land-use change for global amphibian diversity. Nature 480(7378):516–519
Hope D, Gries C, Zhu W, Fagan WF, Redman CL, Grimm NB, Nelson AL, Martin C, Kinzig A(2008) Socioeconomics drives urban plant diversity. Urban Ecology (pp. 339–347)
Jakobsson S, Lindborg R (2017) The importance of trees for woody pasture bird diversity and effects of the European Union’s tree density policy. J Appl Ecol 54(6):1638–1647
Kebrle D, Zasadil P, Hošek J, Barták V, Šťastný K (2021) Large trees as a key factor for bird diversity in spruce-dominated production forests: Implications for conservation management. For Ecol Manag 496:119460
Lepczyk CA, La Sorte FA, Aronson MFJ, Goddard MA, MacGregor-Fors I, Nilon CH, Warren PS(2017) Global patterns and drivers of urban bird diversity. In Ecology and conservation of birds in urban environments (pp. 13–33)
Leveau LM (2019) Urbanization induces bird color homogenization. Landsc Urban Plann 192:103645
Leveau LM (2021) Big cities with small green areas hold a lower species richness and proportion of migrant birds: A global analysis. Urban Forestry and Urban Greening 57:126953
Leveau LM, Jokimäki J, Kaisanlahti-Jokimäki ML (2021) Urbanization buffers seasonal change in composition of bird communities: A multi-continental meta-analysis. J Biogeogr 48(10):2391–2401
Leveau LM, Leveau CM (2012) The role of urbanization and seasonality on the temporal variability of bird communities. Landsc Urban Plann 106(3):271–276
Leveau LM, Leveau CM (2016) Does urbanization affect the seasonal dynamics of bird communities in urban parks? Urban Ecosyst 19(2):631–647
Lowry H, Lill A, Wong BBM (2013) Behavioural responses of wildlife to urban environments. Biol Rev 88(3):537–549
MacGregor-Fors I (2008) Relation between habitat attributes and bird richness in a western Mexico suburb. Landsc Urban Plann 84(1):92–98
MacGregor-Fors I, Morales-Pérez L, Quesada J, Schondube JE (2010) Relationship between the presence of House Sparrows (Passer domesticus) and Neotropical bird community structure and diversity. Biol Invasions 12(1):87–96
Manu SA (2003) Effects of habitat fragmentation on the distribution of forest birds in South Western Nigeria with particular reference to Ibadan Malimbe and other malimbes. University of Oxford
Mason CF (2000) Thrushes are now largely restricted to the built environment in eastern England. Divers Distrib 6(4):189–194
Melles S, Glenn S, Martin K(2003) Urban bird diversity and landscape complexity: species-environment associations along a multiscale habitat gradient.Conservation Ecology, 7(1)
Mersha S, Gebremariam E, Gebretsadik D(2021) Drivers of informal land transformation: a perspective from the peri-urban area of Addis Ababa. GeoJournal, 1–14
Millard J, Outhwaite CL, Kinnersley R, Freeman R, Gregory RD, Adedoja O, Gavini S, Kioko E, Kuhlmann M, Ollerton J (2021) Global effects of land-use intensity on local pollinator biodiversity. Nat Commun 12(1):1–11
Morante-Filho JC, Faria D, Mariano-Neto E, Rhodes J(2015) Birds in anthropogenic landscapes: the responses of ecological groups to forest loss in the Brazilian Atlantic Forest.PLoS One, 10(6)
Newbold T, Scharlemann JPW, Butchart SHM, Şekercioğlu ÇH, Alkemade R, Booth H, Purves DW(2013) Ecological traits affect the response of tropical forest bird species to land-use intensity. Proceedings of the Royal Society B: Biological Sciences, 280(1750), 20122131
Oksanen J, Kindt R, Legendre P, O’Hara B, Stevens MHH, Oksanen MJ et al (2007) The vegan package. Community Ecol Package 10(631–637):719
Pérez-Hernández J, Gavilán RG(2021) Impacts of Land-Use Changes on Vegetation and Ecosystem Functioning: Old-Field Secondary Succession. Plants 2021, 10, 990
Piano E, Souffreau C, Merckx T, Baardsen LF, Backeljau T, Bonte D, Brans KI, Cours M, Dahirel M, Debortoli N (2020) Urbanization drives cross-taxon declines in abundance and diversity at multiple spatial scales. Glob Change Biol 26(3):1196–1211
R Core Team (2022) A Language and Environment for Statistical Computing, R Foundation for Statistical Computing, Vienna Austria. https://www.r-project.org/
Ramankutty N, Foley JA, Hall FG, Collatz GJ, Meeson BW, Los SO, De Brown E, Landis DR (2010) ISLSCP II potential natural vegetation cover. ORNL DAAC
Rico-Silva JF, Cruz-Trujillo EJ, Colorado ZGJ (2021) Influence of environmental factors on bird diversity in greenspaces in an Amazonian city. Urban Ecosyst 24(2):365–374
Rodrigues AG, Borges-Martins M, Zilio F(2018) Bird diversity in an urban ecosystem: the role of local habitats in understanding the effects of urbanization.Iheringia. Série Zoologia,108
Schwarz N, Moretti M, Bugalho MN, Davies ZG, Haase D, Hack J, Hof A, Melero Y, Pett TJ, Knapp S (2017) Understanding biodiversity-ecosystem service relationships in urban areas: A comprehensive literature review. Ecosyst Serv 27:161–171
Shea K, Roxburgh SH, Rauschert ESJ (2004) Moving from pattern to process: coexistence mechanisms under intermediate disturbance regimes. Ecol Lett 7(6):491–508
Solecka I, Sylla M, Świąder M(2017) Urban sprawl impact on farmland conversion in a suburban area of Wroclaw, Poland. IOP Conference Series: Materials Science and Engineering, 245(7), 72002
Stratford JA, Robinson WD (2005) Distribution of neotropical migratory bird species across an urbanizing landscape. Urban Ecosyst 8(1):59–77
Tratalos J, Fuller RA, Evans KL, Davies RG, Newson SE, Greenwood JJD, Gaston KJ (2007) Bird densities are associated with household densities. Glob Change Biol 13(8):1685–1695
United Nations Department of Economics and Social Affairs; Population Division (2021) World Population Clock: 7.9 Billion People (2021) Worldometer. https://www.worldometers.info/world-population
Vasenev VI, Yaroslavtsev AM, Vasenev II, Demina SA, Dovltetyarova EA (2019) Land-use change in New Moscow: First outcomes after five years of urbanization. Geogr Environ Sustain 12(4):24–34
Wilkinson DM(1999) The disturbing history of intermediate disturbance.Oikos,145–147
Wu J (2014) Urban ecology and sustainability: The state-of-the-science and future directions. Landsc Urban Plann 125:209–221
Xu X, Xie Y, Qi K, Luo Z, Wang X (2018) Detecting the response of bird communities and biodiversity to habitat loss and fragmentation due to urbanization. Sci Total Environ 624:1561–1576
Acknowledgements
We are grateful to A.P. Leventis Ornithological Research Institute (APLORI), Jos, Nigeria, and the Department of Conservation Biology and Entomology, University of Cape Coast, Ghana for the financial support.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Statements and declarations
The authors have no relevant financial or non-financial interests to disclose.
Conflicts of interest/Competing interests
The authors declare no competing interest.
Data transparency
The datasets generated during and/or analysed during the current study are not publicly available but are available from the corresponding author on reasonable request.
Consent for publication
The authors do consent to the publication of this work in the above Journal.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Appendices
Appendices

Overall species accumulation curve showing the rate of species accumulation per sample visit in the Cape Coast Metropolitan Assembly
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Afrifa, J.K., Monney, K.A. & Deikumah, J.P. Effects of urban land-use types on avifauna assemblage in a rapidly developing urban settlement in Ghana. Urban Ecosyst 26, 67–79 (2023). https://doi.org/10.1007/s11252-022-01281-0
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-022-01281-0