
Abstract
Across the globe there is an increasing number of initiatives promoting biodiversity in urban areas – both for the benefit of native wildlife and the people who live in cities. In these situations, the role that companion animals, such as cats and dogs, play as predators of wildlife becomes increasingly important. The objectives of this case study were two-fold. Firstly, to investigate activity patterns of domestic cats inside a 75 ha urban reserve; and secondly, to survey the attitudes and beliefs of the community neighbouring the reserve about pet ownership and the threat that domestic cats and dogs may pose to native wildlife. Twelve motion-activated camera traps were triggered by cats 83 times during the 32-day study period. Distance from reserve edge was found to affect the rate of cat detection, with almost six times as many cat-related triggers 25 m from the reserve edge than at 100 m. Distance from the nearest walking track within the reserve had no significant effect on detection rate. The online survey found that the urban reserve is highly valued by the local community for its provision of habitat for native wildlife and, in general, there is strong agreement that the threat cats pose to native wildlife is a problem. Attitudes of cat owners, however, did differ from those of non-owners, especially in their degree of support for suggested solutions. Advocacy and education about the effects of cats on native wildlife may alter the behaviour of cat owners who value native biodiversity, however, these strategies alone are unlikely to persuade pet owners who are not motivated by conservation goals.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Urban biodiversity and cats
As evidence for the importance of human-nature relationships grows (Dearborn and Kark 2010; Aaron and Witt 2011; Cox et al. 2017), there is an increasing desire internationally to encourage the proliferation of biodiversity in urban environments where the majority of people now live (Dunn et al. 2006; United Nations 2014). In some cities, this is resulting in changes to urban ecosystems with restoration and occasionally reintroduction of native flora and fauna (Miskelly and Powlesland 2013; van Heezik and Seddon 2018). Many such initiatives have been successful at achieving conservation goals (Hannon and Hafernik 2007; Miskelly and Powlesland 2013; Vieira et al. 2015) and at engaging the public (Silvertown 2009; Peters et al. 2015). However, sometimes these changes to the urban ecosystem can create social tension, either when the restored biodiversity negatively impacts human interests (Charles 2012; Soulsbury and White 2015), or where the existing behaviours of people adversely affect the survival of species being targeted by conservation. One source of such tension is wildlife predation by free-roaming, owned domestic cats (van Heezik 2010).
Cats have lived as commensals with humans for over ten thousand years and are now one of the most common pets worldwide (Driscoll et al. 2007). As pets, cats increase awareness and lay-person understanding about animals and have helped shape human values about nature (Twardek et al. 2017), however, the predatory nature of felines, in combination with traditions around cat ownership that permit them to roam freely, makes predation of wildlife by cats a highly complex, global conservation issue (Loss and Marra 2017). Cats have been implicated in 26% of bird, reptile and mammal extinctions worldwide since 1500 AD (Doherty et al. 2016). While their effects are most severe on islands with prey that are naïve to mammalian predators, they also have damaging effects in continental systems where other carnivores are present because cat populations are inflated above the carrying capacity of the ecosystem by humans subsidising cats with food, shelter and vaccination (Beutel et al. 2017). Exacerbating the threat of a high abundance of cats in residential areas is evidence that cats can travel great distances. GPS tracking studies conducted in suburban and periurban environments show that cat home ranges vary considerably by individual, but can be as large as 20 ha (Barratt 1997; Kays and DeWan 2004; Metsers et al. 2010; Hall et al. 2016a). This means that parks, reserves and natural environments that are close to residential areas are likely to be entered by cats.
Debate around management of cats is often polarising (Peterson et al. 2012) and despite the body of evidence showing that cats kill large quantities of wildlife (e.g., 100–350 million birds per year in Canada, Blancher 2013; > 20,000 silvereyes (Zosterops lateralis) per year in Dunedin, New Zealand, van Heezik et al. 2010; also Loss et al. 2013; Krauze-Gryz et al. 2017), cat advocacy groups have often pointed at the lack of conclusive evidence that cats can affect species at the population level (RSPB 2018). However, a synthesis of the available research on this topic by Loss and Marra (2017) argues that evidence of such impacts is compelling and stresses the importance of exercising precaution in the face of uncertainty when it comes to management of cats (sensu Calver et al. 2011).
Cats in New Zealand
The threat of feline predation upon native wildlife is well known in New Zealand (Kikillus et al. 2017). Like many introduced mammals, cats are successful hunters of the country’s naïve native fauna which are evolutionarily ill-adapted to cope with mammalian predators. There are many examples of predation by cats (Flux 2007; van Heezik et al. 2010) including at least one extinction in New Zealand: the last known Stephens Island wren (Traversia lyalli) was purportedly killed by the lighthouse keeper’s cat (Galbreath and Brown 2004).
In many cities worldwide, parks and reserves provide important habitat for native wildlife (Lerman et al. 2014; Fernández-juricic and Jokimäki 2001; Wellington City Council 2015). However, given the high densities of cats in New Zealand cities (220–250 km−2, van Heezik et al. 2010) and what is known about their home range sizes, biodiversity in urban reserves is likely to be at risk from cat predation. Cat ownership is deeply entrenched in New Zealand’s culture, as it is in much of the western world, and historically there have been few restrictions on cat ownership compared with dog ownership (van Heezik 2010). While other introduced mammalian predators such as rodents, mustelids and possums can be controlled in reserves by trapping or poisoning, and free-roaming dogs prohibited, fewer regulations exist for cat ownership. In Wellington, New Zealand, proposed changes to an animal bylaw requiring microchipping and registration of cats were met with much public controversy (Radio New Zealand 2016). While many people agreed with the proposal, presumably due to perceived benefits for endemic wildlife, others felt that the proposed changes were an encroachment upon their rights and threatened the safety of cats (Feline Rights New Zealand 2015). It is likely that people with companion (pet) cats may feel differently about the policy than those who do not own cats. In an international study, Hall et al. (2016b) found that while the percentage of owners and non-owners who believed it important to have wildlife in cities was approximately equal, 78% of non-owners believed that cats killing wildlife in urban areas was a serious problem compared with only 51% of cat owners. These differences of opinions probably stem from either differences in perceived knowledge about the ecological impacts of cats as predators, or differences in values for either cats or urban biodiversity. Beliefs and perceived impacts vary widely among different groups of stakeholders (Wald et al. 2013), including cat owners (McLeod et al. 2017). For example, while most cat owners are very concerned about the welfare of their pets, others have little attachment and are largely indifferent to their welfare (McLeod et al. 2017).
Aims
The issue of cat predation of wildlife is complex and multifaceted. While there is a growing body of literature on single aspects of the issue (e.g., prey collected by cats, roaming behaviour or public attitudes) very few studies have attempted to integrate multiple factors. Here we integrate two key facets of the issue in one study system: one ecological (cat activity) and the other social (the attitudes and practices of citizens). The present case study had two key objectives. Firstly, to investigate the distribution of free-roaming domestic cat activity within an urban reserve in Wellington; and secondly, to investigate the views and beliefs of the community neighbouring the reserve regarding pet ownership and the threat that domestic animals pose to native species. It was hoped that examining the attitudes of residents living immediately adjacent to the reserve would provide information about a subset of the public whose actions regarding cat ownership and management could make a difference to biodiversity in the reserve. It also serves to illustrate what level of support might be expected for a variety of potential policies and what opportunities exist in the community for improved education and advocacy. Specifically, the study addressed the following questions: 1. how does the activity of free-roaming domestic cats vary across the reserve, and does distance from the reserve edge and distance from nearest walking track make a difference? 2. what are the attitudes and beliefs of the residents neighbouring the reserve regarding ownership of domestic cats and the threats pets can pose to native wildlife, and how do these attitudes vary with proximity to reserve and pet-ownership status?
Methods
Site description and species
Wellington, the capital city of New Zealand, has seen a recent resurgence in endemic avifauna aided by the creation of the Zealandia ecosanctuary and city-wide control of introduced possums (McArthur et al. 2018; Miskelly 2018; Brockie and Duncan 2012). Nestled in suburban Wellington, Zealandia is a 225 ha wildlife sanctuary enclosed by a specially designed fence that keeps it free of all mammalian pest species except mice (Fig. 1). Since the completion of the fence in 1999 the sanctuary has been the recipient of a number of successful bird translocations including kākā (Nestor meridionalis septentrionalis), toutouwai (Petroica longipes), tieke (Philesturnus rufusater), korimako (Anthornis melanura) and hihi (Notiomystis cincta) (Miskelly and Powlesland 2013). The sanctuary, along with widespread pest control by Wellington City Council (WCC), has also facilitated the growth of existing native bird populations such as kererū (Hemiphaga novaeseelandiae) and tūī (Prosthemadera novaeseelandiae) which are now prolific in the surrounding suburbs (McArthur et al. 2018; Miskelly 2018; Brockie and Duncan 2012).

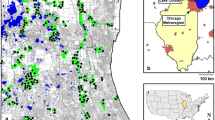
a Locations of Zealandia (outlined in blue with hatching) and Polhill Reserve (outlined in yellow) relative to Wellington CBD (outlined in red). b Location of Wellington City, New Zealand. Satellite image: LINZ 2016
Adjacent to the Zealandia sanctuary on its north-eastern boundary, is the 75 ha Polhill Reserve, ten minutes from central Wellington and surrounded by the suburbs of Aro Valley, Highbury and Brooklyn (Fig. 1). Popular with mountain bikers, runners and birdwatchers, the reserve is maintained through the collaboration of the city council and a number of community groups: Waimapihi Trust, Aro Valley Restoration Project, Brooklyn Trail Builders, and Polhill Restoration Project (PRP) (Prebble 2015, unpublished report). Due to its proximity to Zealandia, Polhill Reserve receives many birds that ‘spill over’ the fence. While bird species such as kererū, tūī and kākā thrive in the reserve, other species such as korimako, tieke and toutouwai are restricted to low numbers and breeding is rare (P. Stanley Ward, PRP, pers. comm. 2016).
Rats (Rattus spp.) and mustelids (Mustela spp.), are controlled by PRP and possums (Trichosurus vulpecula) are managed every 3 months by WCC (Prebble 2015, unpublished report). Residual abundances of these three taxa are monitored by PRP and WCC, however there are currently no methods used to quantify the abundance or distribution of domestic animals in the reserve, despite cats being frequently seen and many people using the reserve to exercise dogs (P. Stanley Ward, PRP, pers. comm. 2016).
Measuring cat activity in the urban reserve
Cat activity was measured using motion-detecting cameras (Bushnell Trophy Cam, Model No. 119677c/119736c). Using buffer functions in ArcGIS software (v. 10.3.1; ESRI 2015) locations were identified in Polhill Reserve that were either 25 m or 100 m from the reserve edge and at the same time either 10 m or 50 m from the nearest track. This provided a set of 56 potential locations comprising four different distance combinations (see legend of Fig. 2). Cameras were installed at 12 of these locations: three at each of the four distance combinations (Fig. 2). These locations were selected based on: availability of trees for camera attachment and having a clear field of view. Cameras were installed on trees at heights of approximately 1 m from the ground. Using the methods of Anton et al. (2018a, b), they were positioned horizontally facing uphill or angled towards the ground 1.5–2 m in front, operated 24 h a day and were set to take three photos over a 3 second period when triggered. The cameras were in place from August 30 to September 30, 2016.

Map of Polhill Reserve showing tracks (black lines), reserve edge (yellow line) and location of camera traps. Marker colour indicates distance from nearest track (blue = 10 m, white = 50 m) and marker shape indicates distance from reserve edge (squares = 25 m, triangles = 100 m). Numbers within markers indicate total number of cat triggers at each camera. Addresses within 100 m of reserve are shown as black dots within semi-transparent yellow polygon. Satellite image: LINZ 2016
On September 3, corflute chew cards baited with cat food were installed on trees within the field of view of each camera to test their efficacy as a lure and detection method for mammalian predators. These were made, installed and inspected using the methods of Sweetapple and Nugent (2011). The chew cards remained in place until September 30 when the cameras were retrieved.
Evaluating community beliefs about pet ownership
All street addresses within 100 m of the reserve were selected to receive a questionnaire. Addresses were identified by creating a 100 m buffer around the outside of the reserve using ArcGIS software (ESRI 2015). This buffer was used to select street addresses from the ‘NZ street address (electoral)’ layer available from the Land Information New Zealand database (https://data.linz.govt.nz/layer/ 50779-nz-street-address-electoral-deprecated/ Accessed 26 Sept 2016). This resulted in the selection of 465 residential addresses in the suburbs of Aro Valley, Brooklyn and Highbury (Fig. 2). On September 27, flyers informing residents of an online survey (and directing them towards its URL) were delivered at all addresses where letterboxes were present (n = 447) between the hours of 10 am and 6:30 pm. That evening the survey was publicised on social media via the Polhill Restoration Group Facebook page. The survey was closed after 21 days, on November 18.
Questions in the survey were designed firstly, to categorise respondents by pet ownership status, proximity of their property to the reserve and their degree of ‘environmental investment’, and secondly, to record their level of agreement with 14 statements reflecting attitudes and beliefs about pet ownership and the threat that cats and dogs may pose to native wildlife (see list of statements in Table 1). Participants recorded their responses to statements on a 5-point Likert scale where 1 indicated strongly disagree and 5 indicated strongly agree.
Statistical analysis
Images from all cameras were viewed and the number capturing each animal species recorded. Images containing cats were used to create a metric of ‘cat triggers’: the total number of independent trigger events capturing photos of cats per camera for the duration of the study period. Attempts to determine a natural interval for independence based on the clustering of observations, sensu Brook et al. (2012), did not reveal a clear cut pattern. We therefore used a threshold of 30 mins as in Brook et al. (2012), assuming that the occurrence of repeat ‘captures’ in this study would be similar given the same species were involved. For every image capturing a cat, the cat’s fur length, colour and any other potentially identifying features (e.g., collar) were recorded in the hope that these might allow identification of individuals.
Due to the study’s small scale (within a single small reserve, where cats regularly migrate across its boundary), and the fact that individual cats could not always be distinguished, we were unable to make estimates of cat abundance. We instead focussed on cat activity and reason that, given the high variability of hunting behaviour and range size of domestic cats (Morgan et al. 2009), a single, very active cat could interact with wildlife as much as several less active cats.
To assess the effect of distance from reserve edge and distance to nearest track on cat activity, a generalised linear model (GLM) was used. Number of cat triggers over the 32-day study period was the dependent variable and distance of camera from both the reserve edge and the nearest track were fixed factors. A Poisson distribution of errors was assumed.
The effect of chew cards on the rate of cat triggers was assessed using a generalised linear mixed model with a Poisson distribution of errors. Number of cat triggers was used as the dependent variable, with time grouped into three 5-day periods as a fixed factor and individual camera as a random factor. Cat triggers recorded between August 30 and September 13 were divided into three periods for analysis: 5 days prior to chew card installation, 5 days immediately after installation and 5 days after this. The assumption being that the potential lure of cat food would be most effective when fresh (i.e., within 5 days of deployment).
It was expected that the different modes of recruitment for the survey (information flyer in mailboxes vs. social media) could introduce bias, as respondents living more than 100 m from the reserve (who learnt of the survey via social media) might be more involved in conservation or have a greater connection with nature on average than those who received physical flyers in their mailboxes. To test for this, a chi-squared test for homogeneity was performed using an ‘environmental investment index’ (EII) to compare respondents who received a flyer (and therefore lived within 100 m of the reserve) with those who reported living within 500 m but did not receive a flyer. The EII was a binary variable, where 1 coded for participants who responded yes for either “Are you involved in any community ecological restoration groups?” or “Do you regularly donate or volunteer with a conservation, environmental or otherwise nature-focused organisation?” and 0 coded for neither. This same test was used to assess whether cat owners and non-cat owners within the 500 m ‘neighbourhood’ differed significantly in their investment in the environment.
To simplify the analysis, responses to the 14 statements in the survey were coded to create a binary measure of agree-not agree (as suggested by Hall et al. 2016b). This was achieved by coding responses of 4 and 5 (agree and strongly agree) as 1 (agree) and 1, 2 and 3 (strongly disagree, disagree and not sure/neutral) as 0 (not agree).
For residents living within 500 m of the reserve, GLMs were used to assess the relationships between the predictor variable: pet ownership status, and dependent variable: agreement with statements. These used a binomial distribution with a logit link. All GLMs were fitted in R v.3.3.2 (R Core Team 2013) using the GLM function from the stats package.
Results
Distribution of cats in the urban reserve
The 12 camera traps were triggered 4,396 times in total and together collected 13,188 images. A large proportion of these were false triggers resulting either from movement of the tree to which the camera was tethered, or from the movement of vegetation in front of the camera. The number of false triggers varied widely between cameras depending on what vegetation type was in its line of sight. A wide variety of animals triggered the camera. Birds were frequently captured, especially introduced species such as blackbirds (Turdus merula), chaffinches (Fringilla coelebs) and sparrows (Passer spp.), but also occasionally native birds such as kākā (n = 5), tieke (n = 1) and kārearea (Falco novaeseelandiae) (n = 1). Images of mammals were rarer. Thirty trigger events captured rabbits or hares (Family: Leporidae), 14 rats and 11 mice (Mus musculus). People were also captured by cameras: most unwittingly, although a few did interact with the chew cards and cameras. No dogs were detected at any of the camera sites. The total number of ‘independent’ cat triggers across all cameras was 83, with cats being detected at 11 of the 12 camera locations and on 31 of the 32 days that the cameras were operational. Many cat triggers appeared to involve the same individuals (possibly at multiple cameras), however, accurate identification of all individuals was not possible. Eight individuals were distinguished based on differences in colour, pelage pattern, hair length and presence of a collar, however it is likely that this number underestimates the total number of different cats detected.
Cats were present in the reserve at all times of day, although rates did vary: the highest number of cat triggers was detected between 6 pm and 10 pm, and the lowest between midnight and 8 am (Fig. 3).

Probability density function of cat triggers at different times of day. Grey rectangles represent night. Dashed lines represent the earliest and latest sunrise and sunset times during the 32-day study period (from www.timeanddate.com; accessed 19/5/19). All times given in New Zealand Standard Time. Vertical lines below plot indicates the distribution of observations (n = 83)
Cats were over five times more likely to be detected close to the reserve edge (25 m in) compared to further in at 100 m (z = −3.629; p < 0.001) with an average of 11.83 ± 5.19 triggers per 32 days at 25 m from the reserve edge compared with 2.00 ± 0.58 at 100 m. Distance from track did not have a significant effect on the number of detections (z = −1.007; p = 0.314) and there was no significant interaction between these two variables (z = 0.845; p = 0.398) (Fig. 4).

Box plot showing the significant effect of distance from reserve edge on number of cat trigger events over a 32 day period (p < 0.001). Colour of overlaid data points indicates camera’s distance from nearest track
Of the 12 chew cards installed, none showed any bite marks or indentations. This was surprising given that photographs showed rats and cats licking or sniffing cards on 22 independent occasions (e.g., Fig. 5). The 5-day period immediately following installation of chew cards saw an 80% increase in cat triggers compared with both the 5 days prior to installation and the period 6–10 days after installation (Fig. 6). The results of the generalised linear mixed model, however, found no significant effect of time period (z = −1.490, p = 0.136) on the number of cat trigger events.

Photograph from motion-detecting camera showing a domestic cat interacting with a corflute chew card

Number of cat triggers during 5-day periods before and after the installation of chew card lures (N = 12 cameras). Camera-level data values are integers, but lines have been jittered for clarity. Error bars around averages are standard errors
Community beliefs about pet ownership
In total, 166 people responded to the survey and of these 61 received information flyers in mail boxes (37%; response rate of mail drop = 14%). The remaining respondents reported finding out about the survey via social media (51%) and word of mouth or email (12%), particularly through community organisations. The majority of responses (60%) came from the three suburbs bordering the reserve; Aro Valley, Brooklyn and Highbury, but others came from more distant Wellington suburbs, the farthest from 18 km away.
Eighty-nine respondents were classified as living within 500 m of Polhill Reserve and of these 32 owned cats, 14 owned dogs, 3 owned both cats and dogs, and 42 did not own a pet. Other pets listed included rabbits (n = 1), chickens (n = 3) and other birds (n = 1). Over half of these respondents (50%) reported visiting the reserve at least once a week and only five (5%) had never visited the reserve (Fig. 7a). The community uses the reserve for a wide range of activities, the most popular being walking (89% of people), running (31%) and bird watching (31%) (Fig. 7b). Other uses suggested by participants included clearing traps, weeding and teaching children about nature. The most frequent response to the question “What attributes do you most value about the reserve?” was that it provides habitat for native wildlife (93%). This was closely followed by the provision of ‘green-space’ (85%) and that the reserve offers an escape from the city (58%) (Fig. 7c).

Summary of survey responses: a. frequency of reserve visitation (n = 81); b. number of responses indicating particular reserve uses (n = 83); and c. number of responses indicating particular values of the reserve (n = 87). Total height of each bar indicates number of responses. The proportion of the two colours in each bar indicate the ownership status of the respondents weighted by the proportion of all respondents who were cat owners (light grey) or non-cat owners (dark grey)
Results of the chi-square homogeneity test showed no significant differences in the Environmental Investment Index (EII) among the respondents living <100 m from the reserve and those living between 100 m and 500 m away (χ2 = 0.538, df = 1, n = 89, p = 0.463). There was also no significant difference in EII among cat owners (CO) and non-cat owners (NO), (χ2 = 0.096, df = 1, n = 88, p = 0.757), however these groups did differ in their responses to other statements.
Perceptions of cats as predators differed by ownership status. A significantly higher proportion of non-cat owners agreed that cats posed a threat to native wildlife in urban reserves (Statement 1, NO = 95% cf. CO = 72%) and that this threat was a problem (Statement 3, NO = 95% cf. CO = 69%) (Table 1, Fig. 8).

Percentage of responses to each level on the Likert scale for the 14 statements (upper = cat owners (CO) and below = non-cat owners (NO)). Asterisks indicate significance (p < 0.05). n = 88 for all statements except 2, 10 and 14 for which n = 87)
Attitudes towards the proposed actions that could reduce cat predation on wildlife also differed. Almost double the proportion of non-cat owners compared with cat owners agreed that the local government should enforce stricter regulations on ownership (Statement 6, NO = 82% cf. CO = 44%) and that cat-free zones were a good solution to the issue of cat predation (Statement 10, NO = 68% cf. CO = 39%). More than double the proportion of non-cat owners over cat owners agreed that keeping all cats inside or on an owner’s property would be a good solution (Statement 7, NO = 61% cf. CO = 25%) (Table 1, Fig. 8).
The reverse was true for the statement that making a cat live indoors is unethical and unfair on the cat (Statement 8, NO = 38% cf. CO = 81% agreement). Ownership status groups also differed significantly in their agreement with the statement that “Free roaming domestic cats are beneficial to native wildlife as they reduce populations of rats and mice” with a significantly higher proportion of cat owners agreeing (Statement 11, NO = 13% cf. CO = 59%) (Table 1, Fig. 8). All other differences in the proportional agreement with statements were non-significant.
Of the 72% of owners who agreed with the statement “Cats pose a threat to native wildlife in urban reserves” 13% did not agree with the statement “The threat of predation by cats on native wildlife is a problem”.
Discussion
Cat distribution in Polhill reserve
Cats were detected at all but one camera location in Polhill Reserve and it is therefore likely that cats are not only frequent visitors to the reserve, but also may be present (albeit at varying densities) across much of its total area. The finding that cats were more frequently detected close to the reserve edge is consistent with findings from a number of other studies (Barratt 1997; Kays and DeWan 2004). A study at another urban reserve in Wellington (Greater Wellington Regional Council 2016, unpublished report) reported a pronounced decrease in cat activity between 50 and 100 m from the urban fringe but also found that one cat was detected at the camera farthest from the urban fringe (600 m). This indicates that even though the activity of cats may initially drop rapidly, some individual cats may roam many hundreds of metres into reserves from the urban fringe. Previous research shows that home ranges of cats in suburban areas vary widely depending on numerous factors (Hall et al. 2016a). Two New Zealand studies report ranges between 0.2 ha and 19 ha but averaging about 3 ha (Barratt 1997; Metsers et al. 2010). Using circular home ranges and assuming these range sizes are representative for the Polhill neighbourhood, cats might be expected to penetrate on average 98 m into the reserve with some cats penetrating up to 246 m. This is consistent with the results of the present study. Given that most of the reserve is within 150 m of the nearest house (the maximum distance to nearest house, close to the Zealandia fence, is around 400 m) it is likely that a large proportion of the reserve falls within the home ranges of cats resident near the reserve boundary. Although the majority of our camera locations were closest to the eastern edge of the reserve, it seems likely that cat activity should be similar at equivalent distances from the north-western edge.
It is possible that some of the cats captured were feral or more likely stray (feral cats are considered those whose populations are not dependent upon human activity, while stray cats receive some provisioning from humans, even if only through scavenging (Farnworth et al. 2010)). However, given that collars were present on some animals, nearly 40% of survey respondents within 500m own cats, and WCC has no records of feral populations in the area, (H. Louw, WCC, pers. comm., 2019) it seems likely that the majority should be owned.
While direct predation upon native species is of primary concern, free-roaming domestic cats have also been shown to have important indirect effects on avian nest success. The presence of a domestic cat around blackbird nests, even for a short period of time, can significantly reduce fecundity by changing parental provisioning rates and increasing rates of nest predation by other species (Bonnington et al. 2013). In addition to occasions of direct predation, it is possible that cat presence may change the foraging behaviours of birds dispersing from the Zealandia sanctuary, interfering with reproductive processes and reducing the likelihood of successful establishment within Polhill Reserve.
Distance from track was not found to have an effect on the frequency of cat detection. Morgan et al. (2009) reported that cats found in an urban reserve most frequently used walking tracks, however, the use of tracks in this reserve for activities such as running, dog walking and mountain biking may also deter cats from these areas. Cameras were not placed directly on tracks in the hope of minimising human interference and it is possible that our cameras missed cats that accessed the reserve primarily along tracks. As a result, our results may underestimate cat activity in the reserve.
This is likewise true for dogs. Although no dogs were detected by the cameras, 15% of survey respondents reported using the reserve for dog exercise, and in the course of the field work for this study dogs were frequently seen both on and off-leash. It seems likely that off-leash dogs remain primarily on tracks and were therefore not detected. There have been a number of reports of dogs off-leash attacking and killing birds in Wellington including fledgling kākā and a tieke in Polhill (Radio New Zealand 2017), however, further research is required to quantify this issue.
Chew cards baited with cat food were found to be ineffective as a tool for detecting cats, although this study does not rule them out as a lure to be used in combination with cameras. Anton et al. (2018b) found that peanut butter lures increased the efficiency of cameras for detection of rats and mice. Given that the third 5-day period received interaction rates the same as before the installation of chew cards it seems likely that bait freshness is an important factor determining rate of interaction. A longer-lasting and easier to refresh alternative lure is rabbit meat in a perforated plastic container as used by Greater Wellington Regional Council (Greater Wellington Regional Council 2016, unpublished report). Additionally, Bengsen and Butler (2011) stress the importance of using visual as well as food-based lures when estimating cat abundance as cats rely heavily on visual cues for hunting and the use of food-based lures alone may appeal only to a subset of the cat population.
Community attitudes towards the reserve and pet ownership
The majority of residents agree that cats pose a threat to native wildlife (86%, Table 1) and that this threat is a problem (85%). This is similar to another New Zealand study that found that 69% of the 1011 participants were concerned about predation on native wildlife by companion cats (Walker et al. 2017). The higher proportion identified in the present study is possibly due to the demography of the neighbourhood which has been found to relate to degree of concern (Grayson et al. 2002; Walker et al. 2017). The high level of concern is reinforced by the finding that Polhill Reserve is highly valued by the community for the habitat it provides for native wildlife. There is also a high level of agreement that pet owners should take steps to reduce the threat posed by their animal to wildlife (88%).
There is generally less support, however, for the enforcement of stricter regulations by local government (68%) and many of the suggested solutions to the problem of wildlife predation by cats, such as confining cats to indoors or the owner’s property (48%), or cat exclusion zones (58%). These results mirror those of a similar Australian study which found mixed support for 24 h confinement (Travaglia and Miller 2018). For the statements proposing solutions, views were particularly divergent among cat owners and non-cat owners. Very few owners of cats agreed that keeping cats indoors or contained on the owner’s property was a good solution to the problem (25%), with a large proportion agreeing that this is unethical (81%). Views and practices about keeping cats indoors differ strongly among cultures. The U.K. is similar to New Zealand with 6% of cats living exclusively indoors, while in the U.S. and Japan confinement is very common with more than 70% of all cats being kept indoors (Hall et al. 2016b). These differences may be attributed to threats to cats that exist in these countries (e.g., predation of cats from other wildlife or traffic accidents) or the lifestyles of owners (Hall et al. 2016b). In Australia, a number of councils have implemented cat-confinement zones which require cat owners to keep cats inside between sunrise and sunset (van Heezik 2010). Councils say that confinement of cats offers benefits to both the cat and wildlife safety and has no negative implication for cat wellbeing if environmental enrichment is provided (City of Kingston 2016).
For cities in which councils or managers wish to reduce the number of cats in urban reserves for the sake of biodiversity, it is especially important to consider the values and beliefs of cat owners as it is the behaviours of this group that have the largest impact (Linklater et al. 2019). By looking at the responses of cat owners to statements 1 and 3 (Table 1) it is possible to categorise owners into three groups that might be approached in different ways: 1. those that do not agree that cats pose a threat to native wildlife; 2. those that agree that cats pose a threat to wildlife and that this threat is a problem; and 3. those that agree that cats pose a threat but do not agree that this is a problem (Fig. 9).

Venn diagram illustrating public perceptions about wildlife predation by cats and strategies for affecting change in public knowledge (arrows)
More than one quarter (28%) of cat owners surveyed fell into the first category, indicating that further education about the ecology of cats as predators in urban ecosystems (van Heezik et al. 2010) as well as the complexity of community dynamics may alter their understanding. This latter point could be particularly important as the greatest disparity between owners and non-owners was over the statement “Free-roaming domestic cats are beneficial to native wildlife as they reduce populations of rats and mice” (NO = 12% cf. CO = 59%). Although there is substantial theoretical and practical evidence for meso-predator release (the increased abundance of a medium-sized predator, such as rodents, after removal of the apex predator that was supressing it) in systems involving cats (Courchamp et al. 1999; Rayner et al. 2007), the assumption that this will occur in all contexts is challenged (Elmhagen and Rushton 2007). Further research on this topic is needed. The significant trapping effort that is maintained in Polhill Reserve means that cats are unlikely to play as large a role in controlling rat and mustelid populations here as they might in other ecosystems. Providing information about the effectiveness of trapping may eliminate a key justification for allowing cats to roam freely in the reserve.
Over half (66%) of cat owners agree that cats pose a threat to native wildlife and that this threat is a problem, similar to the results of other studies (e.g., 70% of respondents agreed cats in wildlife reserves are harmful to wildlife, Lilith et al. 2006) This group is likely to be willing to consider steps that will reduce the threat of their animal to wildlife (e.g., use of belled collars, confinement) and perhaps may decide not to replace their cat. Many approaches that are likely to be acceptable to cat owners, are effective at reducing predation rates. Research has shown that the use of belled collars, for example, reduces the catch rate of cats by around 50% (Ruxton et al. 2002; Gordon et al. 2010) and a number of other products developed have been tested with varying results (Calver et al. 2007; Calver and Thomas 2011; Hall et al. 2015).
A small number of the cat owners surveyed (9%) agree that cats pose a threat to native wildlife but do not agree that this is a problem. This group presumably value ownership of a cat above the proliferation of native wildlife, at least in an urban context, thus education about the detrimental effects of free-roaming cats is less likely to change the behaviour of this group. Recent studies have addressed the challenge of changing owner behaviour (McLeod et al. 2015, 2017; MacDonald et al. 2015; Linklater et al. 2019). In particular, it appears that owners are often less persuaded by advocacy to protect wildlife, but more often by a desire to improve their pet’s welfare and to act for their own convenience.
Our study was only conducted at a single reserve and therefore conclusions about cat behaviour should not be overgeneralised from our results alone. Nonetheless, it provides valuable data of the local community’s attitudes and beliefs at a time when national and international debate about the interaction between cats and wildlife is gaining a heightened profile (Linklater et al. 2019).
Conclusions
Initiatives that encourage the proliferation of native wildlife in urban environments have benefits for wildlife as well as for the lives of people who live in urban areas, however, these initiatives face numerous challenges. This research has found that free-roaming domestic cats are present in the studied urban reserve in Wellington, New Zealand and although cat activity declines with distance from the reserve edge, cats are likely to be active through a large proportion of the reserve’s area. A high proportion of residents in this neighbourhood believe that free-roaming cats within the reserve are a problem for wildlife, although this view is more prevalent among non-cat owners than among owners of cats. While advocacy and education about the effects of cats on native wildlife may alter the behaviour of some cat owners these strategies alone are unlikely to persuade pet owners who are not motivated by conservation goals.
References
Aaron RF, Witt PA (2011) Urban students’ definitions and perceptions of nature. Child Youth Environ 21:145–167
Anton V, Hartley S, Geldenhuis A, Wittmer HU (2018a) Monitoring the mammalian fauna of urban areas using remote cameras and citizen science. J Urban Eco 4:1–9
Anton V, Hartley S, Wittmer H (2018b) Evaluation of remote cameras for monitoring multiple invasive mammals in New Zealand. N Z J Ecol 42:74–79
Barratt DG (1997) Predation by house cats, Felis catus (L.), in Canberra, Australia. I. Prey composition and preference. Wildl Res 24:263
Bengsen A, Butler J (2011) Wildlife research estimating and indexing feral cat population abundances using camera traps. Wildl Res 38:732–739
Beutel T, Reineking B, Tiesmeyer A, Nowak C, Heurich M (2017) Spatial patterns of co-occurrence of the European wildcat Felis silvestris silvestris and domestic cats Felis silvestris catus in the Bavarian Forest National Park. Wildl Biol 1:1–8
Blancher P (2013) Estimated number of birds killed by house cats (Felis catus) in Canada. Avian Conservation and Ecology 8(2):3
Bonnington C, Gaston KJ, Evans KL (2013) Fearing the feline: domestic cats reduce avian fecundity through trait-mediated indirect effects that increase nest predation by other species. J Appl Ecol 50:15–24
Brockie RE, Duncan C (2012) Long term trends in Wellington City bird counts: 1969-2006. Notornis 59:1–6
Brook LA, Johnson CN, Ritchie EG (2012) Effects of predator control on behaviour of an apex predator and indirect consequences for mesopredator suppression. J Appl Ecol 49:1278–1286
Calver M, Thomas S (2011) Effectiveness of the liberator in reducing predation on wildlife by domestic cats. Pac Conserv Biol 16:244–250
Calver M, Thomas S, Bradley S, McCutcheon H (2007) Reducing the rate of predation on wildlife by pet cats: the efficacy and practicability of collar-mounted pounce protectors. Biol Conserv 137:341–348
Calver MC, Grayson J, Lilith M, Dickman CR (2011) Applying the precautionary principle to the issue of impacts by pet cats on urban wildlife. Biol Conserv 144:1895–1901
Charles KE (2012) Tree damage in Wellington as a result of foraging for sap and bark-dwelling invertebrates by the North Island kaka (Nestor meridionalis septentrionalis). Notornis 59:171–175
City of Kingston. (2016). Cat Confinement. Available from http://www.kingston.vic.gov.au/ Community/Pets/Cat-Ownership/Cat-Confinement. (Accessed July 16, 2017)
Courchamp F, Langlais M, Sugihara G (1999) Control of rabbits to protect island birds from cat predation. Biol Conserv 89:219–225
Cox DTC, Shanahan DF, Hudson HL, Plummer KE, Siriwardena GM, Fuller RA, Anderson K, Hancock S, Gaston KJ (2017) Doses of neighbourhood nature: the benefits for mental health of living with nature. BioScience 67:147–155
Dearborn DC, Kark S (2010) Motivations for conservation of urban biodiversity. Conserv Biol 24:432–440
Doherty TS, Glen AS, Nimmo DG, Ritchie EG, Dickman CR (2016) Invasive predators and global biodiversity loss. Proc Natl Acad Sci 113:11261–11265
Driscoll CA, Menotti-Raymond M, Roca AL, Hupe K, Johnson WE, Geffen E, Harley EH, Delibes M, Pontier D, Kitchener AC, Yamaguchi N, O'Brien SJ, Macdonald DW (2007) The near eastern origins of cat domestication. Science 317:519–523
Dunn RR, Gavin MC, Sanchez MC, Solomon JN (2006) The pigeon paradox: dependence of global conservation on urban nature. Conserv Biol 20:1814–1816
Elmhagen B, Rushton SP (2007) Trophic control of mesopredators in terrestrial ecosystems: top-down or bottom-up? Ecol Lett 10:197–206
ESRI (2015) ArcGIS Desktop: Release 10.3.1. Environmental Systems Research Institute, Redlands
Farnworth MJ, Dye NJ, Keown N (2010) The legal status of cats in New Zealand: a perspective on the welfare of companion, stray, and feral domestic cats (). J Appl Anim Welf Sci 13(2):180–188
Feline Rights New Zealand (2015) News. Available from http://www.felinerights.org/ (Accessed August 18, 2016)
Fernández-juricic E, Jokimäki J (2001) A habitat island approach to conserving birds in urban landscapes: case studies from southern and northern Europe. Biodivers Conserv 10:2023–2043
Flux JEC (2007) Seventeen years of predation by one suburban cat in New Zealand. New Zealand Journal of Zoology 34:289–296
Galbreath R, Brown D (2004) The tale of the lighthouse-keeper’s cat: discovery and extinction of the Stephens Island wren (Traversia Iyalli). Notornis 51:193–200
Gordon JK, Matthaei C, Van Heezik Y (2010) Belled collars reduce catch of domestic cats in New Zealand by half. Wildl Res 37:372–378
Grayson J, Calver M, Styles I (2002) Attitudes of suburban Western Australians to proposed cat control legislation. Aust Vet J 80:536–543
Greater Wellington Regional Council (2016) Cat monitoring: Otari-Wilton’s Bush case study Unpublished report:9p
Hall CM, Fontaine JB, Bryant KA, Calver MC (2015) Assessing the effectiveness of the Birdsbesafe antipredation collar cover in reducing predation on wildlife by pet cats in Western Australia. Appl Anim Behav Sci 173:40–51
Hall CM, Bryant KA, Haskard K, Major T, Bruce S, Calver MC (2016a) Factors determining the home range of pet cats: a meta-analysis. Biol Conserv 203:313–320
Hall CM et al (2016b) Community attitudes and practices of urban residents regarding predation by pet cats on wildlife: an international comparison. PLoS One 11:1–31
Hannon ER, Hafernik JE (2007) Reintroduction of the rare damselfly Ischnura gemina (Odonata: Coenagrionidae) into an urban California park. J Insect Conserv 11:141–149
Kays RW, DeWan AA (2004) Ecological impacts of inside/outside house cats around a suburban nature preserve. Anim Conserv 7:1–11
Kikillus KH, Chambers GK, Farnworth MJ, Hare KM (2017) Research challenges and conservation implications for urban cat management in New Zealand. Pac Conserv Biol 23:15–24
Krauze-Gryz D, Zmihorski M, Gryz J (2017) Annual variation in prey composition of domestic cats in rural and urban environment. Urban Ecosyst 20:945–952
Lerman SB, Nislow KH, Nowak DJ, DeStefano S, King DI, Jones-Farrand DT (2014) Using urban forest assessment tools to model bird habitat potential. Landsc Urban Plan 122:29–40
Lilith M, Calver M, Styles I, Garkaklis M (2006) Protecting wildlife from predation by owned domestic cats: application of a precautionary approach to the acceptability of proposed cat regulations. Austral Ecology 31:176–189
Linklater WL, Farnworth MJ, van Heezik Y, Stafford KJ, MacDonald EA (2019) Prioritizing cat-owner behaviors for a campaign to reduce wildlife depredation. Conservation Science and Practice 1:e29
Loss SR, Marra PP (2017) Population impacts of free-ranging domestic cats on mainland vertebrates. Front Ecol Environ 15:502–509
Loss SR, Will T, Marra PP (2013) The impact of free-ranging domestic cats on wildlife of the United States. Nat Commun 4:1–7
Macdonald E, Milfont T, Gavin M (2015) What drives cat-owner behaviour? First steps towards limiting domestic-cat impacts on native wildlife. Wildl Res 42:257–265
McArthur N, Flux I, Harvey A, Ray, S (2018) State and trends in the diversity, abundance and distribution of birds in Wellington City. Client report prepared for Greater Wellington Regional Council. Wildlife Management International Ltd, Blenheim
McLeod LJ, Hine DW, Please PM, Driver AB (2015) Applying behavioural theories to invasive animal management: towards an integrated framework. J Environ Manag 161:63–71
McLeod LJ, Hine DW, Bengsen AJ, Driver AB (2017) Assessing the impact of different persuasive messages on the intentions and behaviour of cat owners: a randomised control trial. Prev Vet Med 146:136–142
Metsers EM, Seddon PJ, Van Heezik YM (2010) Cat-exclusion zones in rural and urban-fringe landscapes: how large would they have to be? Wildl Res 37:47–56
Miskelly CM (2018) Changes in the forest bird community of an urban sanctuary in response to pest mammal eradications and endemic bird reintroductions. Notornis 65:132–151
Miskelly CM, Powlesland RG (2013) Conservation translocations of New Zealand birds, 1863-2012. Notornis 60:3–28
Morgan SA, Hanson CM, Ross JG, Hickling GJ, Ogilvie SC, Paterson AM (2009) Urban cat (Felis catus) movement and predation activity associated with a wetland reserve in New Zealand. Wildl Res 36:574–580
Peters MA, Hamilton D, Eames C (2015) Action on the ground: a review of community environmental groups’ restoration objectives, activities and partnerships in New Zealand. N Z J Ecol 39:179–189
Peterson MN, Hartis B, Rodriguez S, Green M, Lepczyk CA (2012) Opinions from the front lines of cat colony management conflict. PLoS One 7:1–7
Prebble J (2015) Polhill Management Plan Unpublished report:37p
R Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Radio New Zealand (2016) Wgtn councillors vote for cat microchip proposal. Available from http://www.radionz.co.nz/news/national/310118/wgtn-councillors-vote-for-cat-microchip-proposal (Accessed July 16, 2017)
Radio New Zealand (2017) Predator-free in the city. Available from https://www.radionz.co.nz/national/programmes/ourchangingworld/audio/201832286/predator-free-in-the-city (Accessed March 21, 2019)
Rayner MJ, Hauber ME, Imber MJ, Stamp RK, Clout MN (2007) Spatial heterogeneity of mesopredator release within an oceanic island system. Proc Natl Acad Sci 104:20862–20865
RSPB (2018) Are cats causing bird declines? Available from: https://www.rspb.org.uk/birds-and-wildlife/advice/gardening-for-wildlife/animal-deterrents/cats-and-garden-birds/are-cats-causing-bird-declines/. Accessed 11/11/18
Ruxton GD, Thomas S, Wright JW (2002) Bells reduce predation of wildlife by domestic cats (Felis catus). J Zool 256:81–83
Silvertown J (2009) A new dawn for citizen science. Trends Ecol Evol 24:467–471
Soulsbury CD, White PCL (2015) Human-wildlife interactions in urban areas: a review of conflicts, benefits and opportunities. Wildl Res 42:541–553
Sweetapple P, Nugent G (2011) Chew-track-cards: a multiple-species small mammal detection device. N Z J Ecol 35:153–162
Travaglia M, Miller KK (2018) Cats in the Australian environment: what’s your purr-spective? Aus J Environ Manag 255:153–173
Twardek WM, Peiman KS, Gallagher AJ, Cooke SJ (2017) Fido, fluffy and wildlife conservation: the environmental consequences of domesticated animals. Environ Rev 25:381–395
United Nations (2014) World Urbanization Prospects: The 2014 Revision, Highlights (ST/ESA/SER.A/352). United Nations
van Heezik Y (2010) Pussyfooting around the issue of cat predation in urban areas. Oryx 44:153–154
van Heezik Y, Seddon P (2018) Animal reintroductions in peopled landscapes: moving towards urban-based species restorations in New Zealand. Pac Conserv Biol, Special Issue
van Heezik Y, Smyth A, Adams A, Gordon J (2010) Do domestic cats impose an unsustainable harvest on urban bird populations? Biol Conserv 143:121–130
Vieira BP, Fonseca C, Rocha RG (2015) Critical steps to ensure the successful reintroduction of the Eurasian red squirrel. Anim Biodivers Conserv 38:49–58
Wald DM, Jacobson SK, Levy JK (2013) Outdoor cats: identifying differences between stakeholder beliefs, perceived impacts, risk and management. Biol Conserv 167:414–424
Walker JK, Bruce SJ, Dale AR (2017) A survey of public opinion on cat (Felis catus) predation and the future direction of cat management in New Zealand. Animals 7:1–20
Wellington City Council (2015) Our Natural Capital. Wellington’s biodiversity strategy and action plan 2015
Acknowledgements
The authors wish to acknowledge Paul Stanley Ward from the Polhill Restoration Project for his support and knowledge of the study site. We also thank Xandra Carroll and S. Vishnu Vardhan for their assistance setting up cameras in the field, Jennifer Vaughan for her work distributing flyers and three anonymous reviewers for their suggestions on this manuscript. Stephen Hartley acknowledges support from MBIE grant UOWX1601, People, Cities and Nature.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
This research was carried out with the approval of the Victoria University of Wellington Human Ethics Committee (Approval 23591).
Rights and permissions
About this article

Cite this article
Woolley, C.K., Hartley, S. Activity of free-roaming domestic cats in an urban reserve and public perception of pet-related threats to wildlife in New Zealand. Urban Ecosyst 22, 1123–1137 (2019). https://doi.org/10.1007/s11252-019-00886-2
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-019-00886-2