
Abstract
The southern Appalachian Mountains have experienced rapid human population growth rates since the 1980s. Land used practices are shifting from rural to residential. The majority of development has been low density, and is often near biologically diverse areas such as National Forests and National Parks. The long-term effects of urbanization in the southeastern Appalachian Mountains are not clearly understood and even less is known with respect to stream salamander response to urbanization. In order to determine the temporal influence of exurban housing on southern Appalachian streams we sampled 27 first- and second-order streams in watersheds containing exurban developments ranging in age from 4 to 44 years, along with eight forested streams, over the course of two summers. We sought to determine if the relative age of an exurban development related to occupancy and abundance of southern Appalachian stream salamanders. Age of exurban development and other watershed scale variables were not top predictors of salamander assemblages, while local site variables such as salinity and undercut banks predicted the abundance of several species of salamander. Our results suggest local habitat improvements can be used to better conserve salamanders and stream ecosystems in an increasingly urbanized region.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
As the global population increases, more people are moving into urban areas and the size of these areas is increasing (Mackun and Wilson 2011). The population of the United States increased by 9.7% from 2000 to 2010, and 83.7% of the population currently resides in urban areas (Mackun and Wilson 2011). Due to land use change, previously forested or agricultural area, especially forested hillsides, are being converted to residential land (Weir and Bolstad 1998). The majority of this development has been at low housing densities, particularly in the Southeastern U.S. (Mcdonald et al. 2010), where population growth was 16.6% between 2000 and 2010 (Pollard and Jacobsen 2011). This form of residential development is termed exurban development, because the proportion of impervious surface within the watershed and housing density are typically lower than thresholds associated with urban environments, yet higher than rural regions (Theobald 2004). Exurban development is projected to increase in the southern Appalachian Mountains in future decades (Weir and Bolstad 1998, Theobald 2010), which may further jeopardize many species and ecosystems. Even moderate levels of development (<5% impervious surface) can lead to significant alterations in stream ecosystems (Price and Leigh 2006, Walsh et al. 2005).
The southern Appalachian Mountains represent an area of the United States where exurban development is increasing rapidly and the long-term effects are poorly understood (Kirk et al. 2012). Low intensity urbanization can yield levels of degradation similar to acute, major urbanization (Weaver and Garman 1994), so simple assessments of impervious surface may not always yield information on biological response. Gagne and Fahrid (2010) found that as a low-density development aged, the diversity and abundance of five wetland frog species declined. In the same study only the gray tree frog was found to increase in abundance after approximately 40 years (Gagne and Fahrid 2010). A study of bird diversity in suburban areas found that as residential developments aged, the diversity of birds decreased as well; the newest housing developments typically had the highest bird diversity (Loss et al. 2001). As exurban developments age, the influences they exert on stream systems may decrease or increase depending upon the specific mechanisms influencing biota. For example, a potential stressor such as sedimentation is likely to decrease as neighborhoods age; however, losses of riparian vegetation or changes in stream chemistry may be maintained at a high level or increase over time because of mowing, lawn care, or road salt (Kelly et al. 2008). In turn, these changes can impact stream-dwelling organisms in predictable ways (Walsh et al. 2005).
Lungless stream-dwelling salamanders (family Plethodontidae) are an excellent candidate for monitoring stream disturbances in the Southeastern U.S. Stream salamanders are of high importance to Appalachian low-order, fishless streams because they are the top predator in these systems (Davic and Welsh 2004, Keitzer and Goforth 2013, Johnson and Wallace 2011) and contribute to stream nutrient dynamics via nutrient recycling, standing crop, and retention (Keitzer and Goforth 2013, Milanovich et al. 2015, Milanovich and Hopton 2016). Stream salamanders can be useful indicator species because populations are relatively stable and have high abundance (Peterman et al. 2007), but respond quickly to environmental change (Hairston 1986, Price et al. 2011, 2012). Additionally, stream salamanders undergo long-term exposure to the negative influences of urbanization at various life stages both on land and in the water, and as a result salamander densities and species richness are known to be reduced by urbanization (Willson and Dorcas 2003, Barrett and Guyer 2008, Price et al. 2011, 2012). Previous studies indicate salamander abundance decreases with the amount of impervious surface in the watershed (Willson and Dorcas 2003), due to increased flooding in urban streams (Barrett et al. 2010a). Most knowledge on salamander response to development has been derived from the Piedmont ecoregion of the U.S. (Barrett and Price 2014), and very few studies have focused on the Appalachian Mountains, which represent the global center of biodiversity for this taxon (Surasinghe and Baldwin 2015, Frisch et al. 2016).
The recovery trajectories of headwater streams and resident salamander populations following exurban development are unknown. Can such sites return to conditions similar to undisturbed sites over the long-term? Which key environmental variables are most altered by residential development, and do such alterations relate to the size or age of the development? Following rapid growth and expansion of suburban housing developments in the Appalachian Mountains (McDonald et al. 2010), there is a need to answer these questions to better inform habitat- and species-specific conservation plans in a biodiversity hotspot. In this study we assessed the influence of several watershed-scale variables (age of exurban development) and a suite of in-stream measures on the occupancy and abundance of five salamander species. Based on previous research (reviewed in Barrett and Price 2014), we predicted watershed-scale measures of disturbance would be most important in predicting salamander presence or abundance, and that the negative influences of development would be exacerbated in older neighborhoods (Gagne and Fahrid 2010).
Methods
Study area
This study took place within the Southern Blue Ridge Ecoregion of North Carolina and Tennessee. Elevation in this montane area is between 500 and 2000 m and the climate ranges from temperate to boreal. Some areas of this ecoregion exhibit the highest level of rainfall in the United States east of the Cascades. This ecoregion contains more than 400 endemic species of plants and animals, more than any other North American ecoregion (The Nature Conservancy and Southern Appalachian Forest Coalition 2000).
Site selection and landscape variables
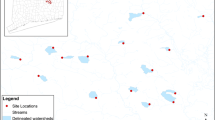
Our goal was to identify as many sites as possible with similar amounts of impervious surface that varied in the age of development present within the watershed. Aerial imagery from 2014 was used to identify watersheds with exurban development. After locating watersheds containing potential exurban developments, a high resolution streams layer from the National Hydrography Dataset was overlain with tax parcel data from Sevier County, TN and Macon County/Jackson County, NC. In ARCGIS 10.1 (ESRI 2011, Redlands, CA). From this overlay, we were able to identify 80 potential fishless streams in watersheds containing only exurban development (i.e. no agriculture, golf course, etc.) Attempts were made to contact the property owners by phone or in person to gain access to the sites, where we then investigated further to determine stream size and development. After ground validations we identified 27 first- or second-order exurban streams. Eight additional streams were selected that contained no impervious surface within their watershed, four in Tennessee within the Great Smoky Mountains National Park and four in North Carolina and the Coweeta Hydrologic Laboratory (Fig. 1). All forested and exurban sites were previously logged, but with harvests that occurred more than 75 years ago.

Sample locations (salamander icons) were in Sevier County, Tennessee (dashed) and Macon County, North Carolina (solid). Inset map shows the sample design for each site
We used tax parcel information to identify the age of each individual structure within the watershed for each stream. These ages were then averaged to assign an age for the exurban development, with the assumption that roads would have been installed shortly before housing construction. Exurban housing ranged in age from 4 to 42 years (mean = 25.99 yrs) across the 27 watersheds with development. We calculated the percentage of impervious surface coverage for each watershed by obtaining 2014 leaf off aerial imagery (0.65 m resolution) from the counties containing our study areas. We hand-delineated polygons around all impervious surfaces and calculated the percent of the watershed they covered, which ranged between 1 and 17% across our sites. We calculated distance to impervious surface using the “near” tool in ARCGIS 10.2 (ESRI 2011, Redlands, CA) by measuring the distance from stream sample plots to the nearest edge of an impervious surface polygon.
Field methods
From May–June, we made three visits to 35 sites in 2014 and again in 2015. Upon our first visit to each study stream we established a 45-m transect along the length of the stream, which was then divided into three 10-m sections, each separated by a 5-m section (Fig. 1). We sampled salamanders by dip-netting in the entire wetted area of the 5-m sections in one pass by a field team lined up perpendicular to the stream (Quin et al. 2007). We also placed two leaf litter bags haphazardly in each of the three 10-m sections to capture larvae (Pauley and Little 1998, Nowakowski and Maerz 2009). Dip-netting was performed on the first and second site visit, and leaf litter bags were placed on the first visit and then checked on the second and third visits during each year. Leaf litter bags were constructed using 2.54 cm (1 in. plastic mesh bags that are 0.09 m2 in size and filled with leaf litter from the riparian area. During sampling occasions we recorded the number of captures and species for all salamanders. Analyses were performed using the sum total of individual captures from a stream via leaf litter bags and the total from dip netting during a given visit. This approach led to four distinct counts in each of the two seasons (two counts from leaf litter bags and two counts from dip netting). The variation in counts allowed for estimates of detection probability (see Analysis below for details).
At each stream, we measured wetted width, maximum depth, bank height, percent undercut banks, and composition of streambed material within each 5-m section. We measured streambed composition as percentage of sedimentation, pebbles, gravel, rock, and bedrock (to the nearest 5%). A YSI Sonde 600R (YSI Ohio, USA) was used to measure water pH, temperature, salinity, conductivity, and dissolved oxygen (DO) in each of the 5-m sections once for each field season. Each of the above in-stream measures were averaged to use in the analysis.
We measured percent canopy cover three times in the middle of the stream using a densiometer on the first site visit during the first field season. Along each of the 5-m sections, we measured 10 m from the bank of the stream to establish a 50-m2 vegetation plot (Fig. 1). The plot was measured on the right side of the first section, the left side of the second section, and the right side of the third section. We estimated percentage of ground covered by course woody debris, vegetation, and bare ground (defined as rock or soil not covered by vegetation) within each plot to the nearest 5%. We considered any fallen limb or tree larger than 10 cm to be course woody debris. We estimated basal area within 0.40 ha (one acre) of each section using a 10 BAF basal area prism. Basal area is an estimate of average tree stem coverage within an area. These values were averaged for the analysis.
Analyses
We used multivariate multiple regression to examine the influence of three watershed-scale variables (percent impervious surface, age of exurban development, and distance to impervious surface) on a suite of uncorrelated (R < 0.7) local scale response variables. We used Type I sum of squares for model evaluation. When a watershed–scale variable was found to significantly predict local factors (α < 0.10), we used least–squares linear regression to identify specific bivariate relationships that were statistically significant (α ≤ 0.05). We used a more liberal α for the multivariate test because we considered this portion of the analysis exploratory in nature. All linear models were run in Program R (R Development Core Team 2005).
Differences in salamander assemblage structure across sites were measured in terms of species-level occupancy and abundance. We began the analysis by first standardizing all covariates by calculating z-scores. Before evaluating the factors influencing salamander occupancy or abundance, the influence of three detection covariates were evaluated against a null model of equal detection: the maximum air temperature for the date of sampling, Julian date of sampling, and sampling method (dip-netting or leaf litter traps). Once the best covariate for detection was identified, we applied that variable to subsequent models where occupancy was evaluated as a function of one or more covariates. We modeled occupancy and abundance as a function of three watershed scale and eleven uncorrelated site scale covariates (site elevation plus those variables listed in Table 1). Occupancy and abundance covariates were initially evaluated as univariate models, and then variables from top models (ΔAIC < 2.0) were combined to test for additive and multiplicative interactions between variables. We constructed each model such that it contained four sampling occasions: two dip net and two leaf litter trap samples. Because age of exurban development was irrelevant for our eight forested sites, we first evaluated the influence of age of exurban development by constructing candidate model sets among the 27 sites with exurban development. For those model sets where age of exurban development did not emerge as a strong predictor of occupancy or abundance (ΔAIC < 4), we then used all 35 sites for further evaluation of habitat factors influencing salamanders.
We ran single-season occupancy models using program Presence (Hines 2006) for mud salamanders (Pseudotriton montanus) because they had adequate detections to fit an occupancy model, but insufficient capture numbers to estimate abundance. Due to concerns over accuracy of species identification for mud salamanders, we used only 2015 data in the analysis. Occupancy is an instantaneous measure of the distribution of a population within a focal region. Occupancy models allow for simultaneous estimates of covariates related to species detection probability (p, a nuisance parameter) and species occupancy (ψ, the parameter of primary interest). At a site a species may be present and detected, present but not detected, or absent. Because the final two scenarios cannot be distinguished, detection probability must be estimated. This is done by recording detection and non-detection data across multiple site visits during aperiod of time short enough to assume no colonization or extinction across sites (Mackenzie et al. 2006).
For species with sufficient captures to generate parameter estimates for abundance, we applied N-mixture abundance models (Royle 2004) using the unmarked package (pcount function; Fiske and Chandler 2011) in Program R (R Development Core Team 2005). We analyzed the two field seasons separately and report here on the results of two single-season models for black-bellied salamanders (Desmognathus quadramaculatus) and Blue Ridge two-lined salamanders (Eurycea wilderae) because identification of these species was certain, while only 2015 data were used for seal salamanders (D. monticola). We applied abundance models using count data for four sampling occasions each season. N-mixture abundance models estimate detection probability (p) and site-level abundance (λ), and users are able to evaluate the models where both of these parameters vary as a function of covariates. Both occupancy and abundance models assume that the population is closed during a season and that counts between sites (streams) are independent.
Results
Local habitat response to landscape factors
Multivariate multiple regression revealed that local response variables were not strongly predicted by landscape-scale factors; however, age of exurban development and impervious surface had a marginally significant relationship to local-scale measures (P = 0.06; Pillai Trace = 0.64 and P = 0.08; Pillai Trace = 0.62 respectively). Subsequent bivariate linear models evaluating the effect of age of exurban development on local environmental measures revealed only one significant relationship with age of development and two with impervious surface (P < 0.05); both older developments and areas with more impervious surface tended to have higher stream banks, and increasing impervious surface increased salinity.
Occupancy and abundance models
The detection probability of mud salamanders was heavily influenced by date of sampling, with detections increasing as the season progressed. As a result of this effect, all models exploring occupancy covariates for this species included Julian date as a detection covariate. Several local-scale variables offered competitive explanations of mud salamander occupancy probability (Table 2). The models with both low ΔAIC (< 2) and high model weight (> 0.20) included DO, the percent of undercut bank, and site elevation. Mud salamanders occupied streams at lower elevation sites that had lower levels of dissolved oxygen (Fig. 2). They were also associated with sites that had a higher percentage of undercut banks.

Plot of the top model for occupancy of mud salamanders (Pseudotriton montanus) in western NC and eastern TN, USA, which included both elevation (plotted here at the mean, 1st, and 3rd quartiles) and dissolved oxygen (standardized values are shown). X-axis values are standardized, actual values ranged between 85 and 100% dissolved oxygen
Detection probability of black-bellied salamanders was highest when the dip-net method was used, so sampling method was incorporated into all subsequent models of abundance. The only well-supported model for black-bellied salamanders was one in which abundance decreased with increasing salinity. This model had the most support in both 2014 and 2015 (Table 3; Fig 3). Blue Ridge two-lined salamander detection probability was also highest when animals were sampled with a dip-net. Subsequent models of abundance that included this detection covariate revealed different top explanatory variables between years. In 2014 DO best predicted Blue Ridge two-lined salamander abundance (negative relationship, Fig. 3c), and in 2015 a model in which abundance increased with percentage of impervious surface was the best supported (Table 4, Fig. 3d). Seal salamander detection probability was a function of sampling method. None of the covariates we evaluated in the 2015 data emerged as better predictors of abundance than a null model in which abundance was assumed to be equal across all sites.

Plots of the top abundance model for black-bellied salamanders for a 2014 and b 2015, and Blue Ridge two-lined salamanders for c 2014 and d 2015. The black midline on each plot is the mean abundance value, while the outer gray lines represent the 95% confidence intervals. X-axis values are standardized, actual values were 0–0.16 g/L, 86.9–98.8%, and 0–17.7% for salinity, percent dissolved oxygen, and percent impervious surface respectively
Discussion
While abundance and occupancy of the salamander assemblages we sampled were not directly predicted by watershed-scale variables, two of these larger-scale measures, age of exurban development (across a range of 4 to 42 years) and distance to impervious surface, marginally predicted local-scale environmental variables. Nevertheless, further evaluation of this relationship indicated only bank height increased significantly with increasing amounts of impervious surface and in older neighborhoods, possibly from long term changes in hydrology, and that more impervious surface led to higher salinity levels. Overall, these results indicate that the mere presence of an exurban development (defined as decentralized urban development with the level of impervious surface in our study being below 20%) in Southern Appalachia may not dramatically alter stream characteristics. Our results show that certain actions like reduction in the use of road salt can be taken to reduce the influence of an exurban development on streams and salamander populations.
Although stream salamander abundance returns to pre-disturbance conditions in 20–60 years after a timber harvest (Demaynadier and Hunter 1995, Crawford and Semlitsch 2008, Homyack and Haas 2009), we did not find a similar salamander recovery with time in exurban neighborhoods. Timber harvest and housing construction both entail removal of large quantities of vegetation and considerable erosion. Changes in stream salamander assemblage following forest harvest can be mitigated by forested buffers (Demaynadier and Hunter 1995, Peterman and Semlitsch 2009); however, buffers in urban areas have not been shown to limit the influence of riparian forest loss on stream salamanders (Willson and Dorcas 2003). Age of exurban development never significantly predicted salamander abundance or occupancy, or local-site habitat variables. This is likely related to key differences exhibited by exurban developments in the presence of impervious surface. These surfaces represent a press disturbance, which leads to a cascading complex of stressors (Burcher et al. 2007, Barrett and Price 2014). The ways in which these stressors interact likely varies among watersheds and species responses are not likely to be uniform. For example, increasing salinity (likely from road salting) had negative impacts on one salamander species, but did not appear as an important predictor for others (Table 3; Fig. 3). Salinity, among other stressors, would not exist in forested watersheds managed for timber, so the ecological inferences that can be transferred between these disturbance types is limited.
Variation in environments differentially influenced occupancy or abundance of our focal species. On average, mud salamanders had a higher occupancy probability at lower elevation sites. In addition to this factor, the species was more commonly found at sites with a high percentage of undercut banks. In the Blue-Ridge ecoregion mud salamanders have been shown to tolerate disturbance, but they appear to be less tolerant in the piedmont ecoregion (Surasinghe and Baldwin 2015). In contrast, black-bellied salamanders are thought to be indicators of less disturbed habitat (Surasinghe and Baldwin 2015). Our top-ranked models support this idea, and implicate salinity as the driver of black-bellied salamander decline in exurban watersheds. Stream salinity in mountainous areas almost certainly increases due to the use of road salt in the winter. Kelly et al. (2008) showed that up to 91% of salinity in rural streams could be attributed to road deicing, and road salt not only persisted beyond the application period, but water salinity increased over time. Howard and Haynes (1993) showed that only 45% of the salt applied to roads each year escaped the watershed; remaining salt was retained and slowly leaked out with the ground water. Abundance relationships for Blue Ridge two-lined salamanders differed by year; however, in both years species abundance increased with typical indictors of higher disturbance (high % impervious surface and low DO). Black-bellied salamanders may reduce abundance of Blue Ridge two-lined salamanders in less disturbed habitats (Crawford 2016); thereby explaining high abundance of Blue Ridge two-lined salamanders in disturbed areas where black-bellied salamanders are reduced.
Other studies have shown impervious surface to be a strong negative influence on most salamander populations (Gagne and Fahrid 2010, Barrett and Price 2014). Our results are largely inconsistent with these studies. Most species we surveyed did not show a strong response to impervious surface, and Blue Ridge two-lined salamanders had higher occupancy in areas with more impervious surface. Our data reveal the prominent role local-scale measures of the environment can play in setting occupancy and abundance for stream organisms. It is possible that in steep–slope, low–order streams, watershed-scale variables become less important and local-scale habitat becomes the driving influence (Cecala et al. 2014; Frisch et al. 2016). Montane exurban developments are typically found along ridge-lines and higher elevations, and are more associated with streams. Because the influences of land-use change become more important at larger spatial scales (Roth et al. 1996), land managers and future researchers should consider the size of the watershed when making management decisions. It is likely that there is a threshold for percent impervious surface influence on salamander assemblages in streams surrounded by exurban developments, but that that value is above our sample range (>17% impervious surface). Management of larger streams, especially large enough for fish, will most likely need to consider impervious surface at thresholds much lower than 17% (Helms and Feminella 2005). It is also important to acknowledge that there is variation in occupancy and abundance even in forested sites and that previous disturbance history may also be playing a role in urbanized watersheds.
Conclusion
Our data align with other studies in that drivers to changes in salamander assemblages are complex and non-singular (Burcher et al. 2007, Barrett et al. 2010b, Barrett and Price 2014). Our data show that local habitat has a much stronger influence on stream salamander populations than watershed-scale variables such as age of exurban development and impervious surface in montane regions. Land owners and developers who aim to maintain stream communities similar those found in nearby forested sites should consider forested riparian buffers, maintaining heterogeneous stream substrate, and reducing water salinity. The amount of impervious surface within the watershed and the distance between impervious surfaces and streams may indirectly influence salinity and stream substrate (Howard and Haynes 1993; Kelly et al. 2008). We were only able to find a single study that evaluated the ability of forested buffers to decrease stream salinity. Results from this research suggested that buffers in the streams evaluated did not protect streams from elevated ion concentrations (Madden et al. 2007). Salinity could be reduced by limiting road salting to times of high use, such as the holiday season in late November to early January, or in areas with permanent residents. Working with homeowners’ associations and county governments may provide the easiest way to develop management strategies for exurban neighborhoods, as it would allow for multi-stakeholder development of solutions over broad areas.
References
Barrett K, Guyer C (2008) Differential responses of amphibians and reptiles in riparian and stream habitats to land use disturbances in Western Georgia, USA. Biol Conserv 141:2290–2300
Barrett K, Helms BS, Guyer C, Schoonover JE (2010a) Linking process to pattern: Causes of stream-breeding amphibian decline in urbanized watersheds. Biol Conserv 143:1998–2005
Barrett K, Helms BS, Samoray S, Schoonover JE (2010b) Growth patterns of a stream vertebrate differ between urban and forested catchments. Freshw Biol 55:1628–1635
Barrett K, Price SJ (2014) Urbanization and stream salamanders: a review, conservation options, and research needs. Freshw Sci 33:927–940
Burcher CL, Valett HM, Benfield EF (2007) The landcover cascade: Relationships coupling land and water. Ecology 88:228–242
Cecala KK, Lowe WH, Maerz JC (2014) Riparian disturbance restricts in-stream movement of salamanders. Freshw Biol 59:2354–2364
Crawford JA (2016) Trade-off between dessication and predation risk in the Blue Ridge two-lined salamander (Eurycea wilderae). Copeia 104:21–25
Crawford JA, Semlitsch RD (2008) Post-disturbance effects of even-aged timber harvest on stream salamanders in Southern Appalachian forests. Anim Conserv 11:369–376
Davic RD, Welsh HH (2004) On the ecological roles of salamanders. Annu Rev Ecol Evol Syst 35:405–434
deMaynadier PG, Hunter ML (1995) The relationship between forest management and amphibian ecology: A review of the North American literature. Environ Rev 3:230–261
ESRI (2011) ArcGIS Desktop: Release 10. Environmental Systems Research Institute, Redlands
Fiske, I. and Chandler, R. (2011) Unmarked: An R package for fitting hierarchical models of wildlife occurrence and abundance. J Stat Softw 43(10), 1–23. URL http://www.jstatsoft.org/v43/i10
Frisch JR, Peterson JT, Cecala KK, Maerz JC, Jackson CR, Gragson TL, Pringle CM (2016) Patch occupancy of stream fauna across a land cover gradient in the southern Appalachians, USA. Hydrobiogia 773:163–175
Gagne SA, Fahrid L (2010) Effects of Time Since Urbanization on Anuran Community Composition in Remnant Urban Ponds. Environ Conserv 37:128–135
Hairston NG (1986) Species packing in Desmognathus salamanders: Experimental demonstration of predation and competition. Amer Nat 127:266–291
Helms B, Feminella J (2005) Detection of biotic responses to urbanization using fish assemblages from small streams of western Georgia, USA. Urban Ecosyst 8:39–57
Hines JE (2006) PRESENCE2 - Software to estimate patch occupancy and related parameters. USGS-PWRC. http://www.mbr-pwrc.gov/software/presence.html
Homyack JS, Haas CA (2009) Long-term effects of experimental forest harvesting on abundance and reproductive demography of terrestrial salamanders. Biol Conserv 142:110–121
Howard KWF, Haynes J (1993) Groundwater contamination due to road de-icing chemicals – salt balance implications. Geosci Can 20:1–8
Johnson BR, Wallace JB (2011) Bottom-up limitation of a stream salamander in a detritus-based food web. Can J Fish Aquat Sci 62:301–311
Keitzer SC, Goforth RR (2013) Spatial and seasonal variation in the ecological significance of nutrient recycling by larval salamanders in Appalachian headwater streams. Freshwater Sci 32:1136–1147
Kelly VR, Lovett GM, Weathers KC, Findlay SEG, Strayer DL, Burns DJ, Likens GE (2008) Long-term sodium chloride retention in a rural watershed: Legacy effects of road salt on streamwater concentration. Environ Sci Technol 42:410–415
Kirk RW, Bolstad PV, Manson SM (2012) Spatio-temporal trend analysis of long-term development patterns (1900-2030) in a Southern Appalachian County. Landscape Urban Plan 104:47–58
Loss SR, Ruiz MO, Brawn JD (2001) Relationships between avian diversity, neighborhood age, income, and environmental characteristics of an urban landscape. Biol Conserv 142:2578–2585
Mackenzie DI, Nichols JD, Royle JA, Pollock KH, Bailey LL, Hines JE (2006) Occupancy estimation and modeling: inferring patterns and dynamics of species occurrence. Academic Press, San Diego
Mackun P, Wilson S (2011) Population Distribution and Change: 2000 to 2010. 2010 Census Briefs https://www.census.gov/prod/cen2010/briefs/c2010br-01.pdf Accessed 20 August 2014
Madden S, Robinson G, Arnason J (2007) Spatial variation in stream water quality in relation to riparian buffer dimensions in a rural watershed of Eastern New York State. Northeast Nat 14:605–618
Mcdonald RI, Forman RTT, Kareiva P (2010) Open Space Loss and Land Inequality in United States’ Cities, 1990-2000. PLoS One 5(3):e9509
Milanovich JR, Hopton ME (2016) Stoichiometry of excreta and excretion rates of a stream-dwelling plethodontid salamander. Copeia 104:26–34
Milanovich JR, Maerz JC, Rosemond AD (2015) Stoichiometry and estimates of nutrient standing stocks of larval salamanders in Appalachian headwater streams. Freshw Biol 60:1340–1353
Nowakowski AJ, Maerz JC (2009) Estimation of larval stream salamander densities in three proximate streams in the Georgia Piedmont. J Herp 43:503–509
Pauley T, Little M (1998) A new technique to monitor larval and juvenile salamanders in stream habitats. Banisteria 12:32–36
Peterman WE, Craword JA, Semlitsch RD (2007) Productivity and significance of headwater streams: Population structure and biomass of the black-bellied salamander (Desmognathus quadramaculatus). Freshw Biol 53:347–357
Peterman WE, Semlitsch RD (2009) Efficacy of riparian buffers in mitigating local population declines and the effects of even-aged timber harvest on larval salamanders. Forest Ecol Manag 257:8–14
Pollard K, Jacobsen L (2011) The Appalachian Region in 2010: A census data overview chart book. URL http://www.arc.gov/research/researchreportdetails.asp?REPORT_ID=94 Accessed August 2015
Price K, Leigh DS (2006) Morphological and sedimentological responses of streams to human impact in the Southern Blue Ridge Mountains, USA. Geomorphology 78:142–160
Price SJ, Cecala KK, Ra B, Dorcas ME (2011) Effects of urbanization on occupancy of stream salamanders. Conserv Biol 27:547–555
Price SJ, Browne RA, Dorcas ME (2012) Evaluating the effects of urbanization on salamander abundances using a before-after control-impact design. Freshw Biol 57:193–203
Quin T, Hayes MP, Dugger DJ, Hicks L, Hoffmann A (2007) Comparison of two techniques for surveying headwater stream amphibians. J Wildlife Manage 71(1):282–288
R Development Core Team (2005) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. ISBN 3–900051–07-0, URL http://www.R-project.org
Roth NE, Allan JD, Erickson DL (1996) Landscape influences on stream biotic integrity assessed at multiple spatial scales. Landsc Ecol 11:141–156
Royle JA (2004) N-mixture models for estimating population size from spatially replicated counts. Biometrics 60:108–115
Surasinghe TD, Baldwin RF (2015) Importance of riparian forest buffers in conservation of stream biodiversity: response to land uses by stream-associated salamanders across two southeastern temperate ecoregions. J Herpetol 49:83–94
The Nature Conservancy and Southern Appalachian Forest Coalition (2000) Southern Blue Ridge Ecoregional Conservation Plan: Summary and Implementation Document. The Nature Conservancy: Durham, North Carolina. URL https://www.conservationgateway.org/ConservationPlanning/SettingPriorities/EcoregionalReports/Documents/SBR-V1.pdf Accessed August 2015
Theobald DM (2004) Placing exurban land-use change in a human modification framework. Front Ecol Environ 2:139–144
Theobald DM (2010) Estimating natural landscape change from 1992 to 2030 in the conterminous US. Landsc Ecol 25:999–1011
Walsh CJ, Roy AH, Feminella PD, Cottinham PD, Groffman PM, Morgan RP (2005) The urban stream syndrome: Current knowledge and the search for a cure. J N Am Benthol Soc 24:706–723
Weaver LA, Garman GC (1994) Urbanization of a watershed and historical changes in a stream fish assemblage. T Am Fish Soc 123:162–172
Weir DN, Bolstad P (1998) Land-Use Changes in Southern Appalachian Landscapes: Spatial Analysis and Forecast Evaluation. Ecosystems 1(6):575–594
Willson JD, Dorcas ME (2003) Effects of Habitat Disturbance on Stream Salamanders: Implications for Buffer Zones and Watershed Management. Conserv Biol 17:763–771
Acknowledgements
We thank B. Cairco, M. McDevitt, B. Miller, N. Pappas, undergraduate researchers participating over several semesters in a Clemson University Creative Inquiry course, and many volunteers for assistance in the field and laboratory. We sincerely appreciate the numerous landowners who provided access to their property during the course of this project along with the Great Smoky Mountains National Park and Coweeta Hydrologic Laboratory for granting us access to forested reference sites. This work was supported by the Creative Inquiry program and Department of Forestry and Environmental Conservation at Clemson University. We are also grateful for the additional funding provided by the City of Greenville Zoo and University of North Carolina Highlands Biological Station. This research was approved by Clemson University’s IACUC under AUP 2014-021.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Weaver, N., Barrett, K. In-stream habitat predicts salamander occupancy and abundance better than landscape-scale factors within exurban watersheds in a global diversity hotspot. Urban Ecosyst 21, 97–105 (2018). https://doi.org/10.1007/s11252-017-0694-x
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-017-0694-x