
Abstract
Concentrated human activities such as the burning of fossil fuels have resulted in chronic nitrogen (N) additions to urban ecosystems. We predicted that urban development in North Texas (NTX; the largest “megapolitan” region in the Great Plains) would be positively correlated with atmospheric concentrations of nitrogen oxides (NOx) and with the leaf tissue quality (lower C:N ratio) and herbivory of the long-lived native tree, post oak (Quercus stellata). Data from air monitoring stations were used to calculate distance-weighted estimates of atmospheric NOx for 11 sites of differing urban development across NTX. Soil samples were collected at each site along with post oak leaves, estimates of herbivory, and measurements of tree size. Percent urban development was strongly positively correlated with atmospheric NOx concentrations, though there was no correlation between atmospheric NOx and soil N. There was a positive relationship between soil N and leaf tissue quality, but only where atmospheric NOx was relatively low, possibly due to factors that covary with urban development. Herbivory was not significantly correlated with leaf tissue quality, but leaves from the two most urban sites had the greatest amount of insect herbivory. The NOx concentrations in NTX were lower than other industrialized cities, which may be due to climate and topography differences or the relatively young age of this urban area. This study adds to the expanding body of literature examining how urban ecosystems are affected (or not) by N deposition and suggests that interactions among NOx, soils, and plants are complex and sometimes, counterintuitive.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
With the world population growing and urban areas expanding, more native landscape is being transformed to suit human needs. If current trends in population growth and density continue, urban land cover will increase 1.2 million km2 by 2030, which is triple the urban land area in 2000 (Seto et al. 2012). In urban areas, concentrated human activities result in habitat fragmentation, altered hydrology, elevated temperatures, greater concentrations of carbon dioxide (CO2), higher regional ozone (O3), and altered nutrient cycles (Pickett et al. 2011). All of these alterations to the landscape can impact native flora and fauna, though one of the most impactful is the addition of nutrients to a nitrogen (N) -limited landscape (Bobbink et al. 2010; Pan et al. 2010).
Biologically available N is produced naturally (via lightning and biological nitrogen fixation) at a rate of about 125 Tg N per year. Through various activities, humans introduce an additional 300 Tg N per year (Galloway et al. 2008). In urban areas, biologically available nitrogen is added primarily through the combustion of fossil fuels. The combustion of fossil fuels in automobiles and industry result in the creation of nitrogen oxide byproducts (NO and NO2, collectively NOx). These NOx compounds are released into the atmosphere and may return to the ground in gaseous form, as dry particles, or most commonly as nitrate (NO3 −) ions in precipitation - a process known as N deposition. Thus, atmospheric NOx concentrations are directly positively correlated with NO3-N deposition in the U.S. (Butler et al. 2003; Butler et al. 2005; Lawrence et al. 2000). Atmospheric NOx inputs to urban areas enter an already complex N cycle, further complicated by human activities and the interactions between human activities and climate. This may result in an urban N cycle drastically different from the surrounding rural area.
In N-limited terrestrial ecosystems, plant communities that receive additional N inputs generally exhibit greater overall productivity (Lebauer and Treseder 2008; Vitousek et al. 1997). At the organismal level, increases in N availability increase plant growth rate, reproductive output, tissue quality (lower C:N ratio), and palatability to herbivores (Chapin 1980; Yang et al. 2011b). However, greater nutrient availability can also change competitive relationships among species, resulting in a loss of biodiversity - particularly in native flora (Aronson et al. 2014; Bobbink and Roelofs 1995). Thus, nitrogen additions may affect plant species and communities in complex and conflicting ways.
Our understanding of plant responses to anthropogenic N additions is further complicated by the fact that responses are also mediated (or co-limited) by aspects of climate such as temperature and precipitation (Lebauer and Treseder 2008; Shen et al. 2008; Xia and Wan 2008; Yang et al. 2011a). Aspects of climate influence N mobility and the ability of ecosystems to act as sources or sinks for N additions (Bai et al. 2015).
Across the U.S., there is a growing body of research contributing to our understanding of the interaction between anthropogenic N and climate (Bedison and McNeil 2009; Boggs et al. 2005; Kahan et al. 2014; McNeil et al. 2007). This body of research encompasses many ecoregions, however, there is a distinct paucity of research for the southern Great Plains. North Texas, the metropolitan area encompassing the cities of Dallas and Fort Worth (henceforth NTX), is located within the sub-humid, sub-tropical southern Great Plains ecoregion and has a population of over 7 million (U.S. Census Bureau 2016). The NTX area is rapidly expanding and has been identified as an area of urban development ripe for exploring the impacts of urbanization (Grimm et al. 2008). Despite this, there has been very little investigation into the effects of concentrated human activities on this biome; such research could help clarify the role of climate variables in urban ecosystem properties and function by providing an important comparison to existing research sites.
Here, we explored the effects of chronic atmospheric NOx exposure on a long-lived native tree, post oak (Quercus stellata), across a gradient of urban development in NTX. By sampling soil and trees located in areas of differing urban development and comparing them to annual atmospheric NOx concentrations, we were able to test the following hypotheses:
-
1.
Atmospheric NOx will increase along a gradient of increasing urban development
-
2.
Increased atmospheric NOx will be positively correlated with:
-
a.
elevated soil nitrogen (via N deposition)
-
b.
increased tissue quality (lower leaf C:N ratio via greater soil N availability)
-
c.
increased herbivory of leaf tissue (via increased palatability)
-
a.
Methods
Study area
The area surrounding the Dallas-Fort Worth metropolitan area belongs to the “Cross Timbers Climate Division,” a sub-tropical and sub-humid mixed savanna and woodland (National Oceanic and Atmospheric Administration (NOAA) 2016). Post oak (Quercus stellata) and blackjack oak (Quercus marilandica) dominate the overstory of the Cross Timbers, with an understory of shrubs and grasses. Where there are stream bottoms, trees such as bur oak (Quercus macrocarpa), Shumard oak (Quercus shumardii), and sugar hackberry (Celtis laevigata) dominate. The sites in our study have a fifty-year annual precipitation average of 92 cm with site averages ranging from 84 to 107 cm and a fifty-year mean temperature of 18.3 °C with site averages ranging from 17.8–18.8 °C. Though the region is categorized as humid subtropical, cyclical oscillations in Pacific Ocean surface temperatures and air pressure known as El Niño and La Niña have long-term impacts on Texas precipitation, leading to periods of moderate to severe drought (Texas Water Deveopment Board 2012). The region is drained by the Trinity River and its forks. Very little of the natural ecoregions remain due to cattle grazing, conversions to cropland, and urbanization (Griffith et al. 2007).
The first major period of population growth in NTX occurred in the late 1880s and a second rapid development period began in the 1960s (Vision North Texas 2008). The pattern of development in NTX is such that the city-centers of Dallas and Fort Worth were the earliest settled and are now the most densely populated. The percentage of land dedicated to urban development decreases in all directions with increasing distance from the city-centers.
Study species and experimental design
Post oaks were chosen as the subject of this study because of their ubiquity and status as the dominant tree in the region. Post oaks are a slow-growing deciduous species, and remnant forests in NTX have an age range of 200–300 years (Diggs et al. 1999). The post oaks of the NTX region are well adapted to the poor soil and periods of extreme drought.
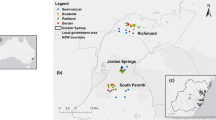
Survey sites were chosen a priori from publicly accessible parks along and near the Trinity River. Nineteen sites were identified as possible survey sites, and eleven were chosen based on the presence of post oak trees. The sites represented a range of urban development (see section below) with less urban sites on both the eastern and western sides of the metropolitan region (Fig. 1).

Map of the NTX area with urban development in light gray, surface water in dark gray, survey sites (indicated by a plus), and NOx monitoring sites (indicated by a point); inset denotes locations of counties in TX
Between May 28, 2014 and June 13, 2014, the survey sites were explored on foot to locate the natural areas – those not maintained by watering, mowing, or fertilizing – within each park. The first five post oak trees identified within natural areas were selected for the survey. For each tree, diameter at breast height (DBH) was measured, and loose litter (Oi layer) was removed from an area near the base of the tree so that a 10 cm long × 5 cm diameter PVC pipe could be used to obtain a soil sample. The lowest main branch of the tree was identified and followed laterally to the point at which the branch was approximately 3.5 cm. There, the terminal branch was separated from the tree using a 3.5 m pole pruner. To estimate herbivory, five leaves were randomly selected from the branch and photographed on grid paper to be analyzed later. An additional five leaves (with no or minor herbivory) were haphazardly selected from the same branch for use in quantifying tissue quality (C:N ratio).
To calculate our estimate of herbivory, the leaf photographs were opened in Adobe Photoshop V 14.1.2. The outline of the leaf as it was in the field was traced and the area converted from pixels to cm2. The remaining leaf tissue was used to hand draw a full leaf shape based on a generalized post oak leaf template scaled to appropriate size. The total area of the estimated original leaf was divided by the actual leaf area and subtracted from one hundred to estimate percent herbivory.
Soil samples were refrigerated at 3 °C before being sent to Texas A&M Soil, Water, and Forage Testing Lab (College Station, TX) for analysis of pH, KCl-extractable nitrate-N (NO3-N) and ammonium-N (NH4-N), phosphorous (P), potassium (K), calcium (Ca), magnesium (Mg), sulfur (S), and sodium (Na). Leaf samples were dried in a 50 °C oven for one week before grinding and analysis of percent C and percent N using a Perkin Elmer CHN series 2400 analyzer.
Measures of atmospheric nitrogen and urban development
The Texas Commission on Environmental Quality (TCEQ) has established atmospheric NOx monitoring sites across North Texas (Fig. 1). Because NOx concentrations (ppbv) are positively correlated with N deposition (Butler et al. 2003; Elliott et al. 2007; Likens et al. 2005; Redling et al. 2013), we used atmospheric NOx as a proxy for N deposition in this study. Hourly NOx concentration data were obtained from the TCEQ’s on-line database (TCEQ 2015a) for the period of 01 January 2013 to 31 December 2013 (the most recent full year of data prior to the study). Atmospheric NOx data from 2011 and 2012 were also examined, and matched very closely with 2013, thus the 2013 data are considered to be representative for the region and are the only data presented here. An inverse distance weighting formula was used to calculate the estimated hourly NOx values in parts per billion by volume (ppbv) for the survey sites based on all available TCEQ air monitoring sites within a 40 km radius. The formula was as follows:
where (P) represents estimated NOx values for each survey site (i), G is the number of monitoring stations, Pj is the NOx values of monitoring station at location j, and Dij is the distance between the monitoring station j, and survey site, i. In this equation n = 2 - a constant used by NOAA for determining missing rainfall measurements. For the KSP survey site, only one air monitoring station was within 40 km (3 km away) and consequently was the only value used. For the MWP site, there were no data from the closest TCEQ monitoring site, therefore, the next closest site (56 km away) was used as the sole data source. The 2013 hourly values were averaged into monthly values to examine seasonal trends, and each hour of the day was averaged across 2013 to examine diurnal trends.
To calculate “percent urban development,” we used the U.S. Department of the Interior’s 2011 National Land Cover Database (NLCD; Homer et al. 2015). NLCD urban land-use categories are calculated based on percent impervious surface area (ISA). The NLCD categories “Developed High Intensity,” “Developed Medium Intensity,” and “Developed Low Intensity” represent ISA values of 80–100%, 50–79%, and 20–49% respectively. For a set radius around each site, the area of these categories were summed into one value and divided by the total area of that radius to calculate “percent urban development.” This was done for each survey site at a 1 k, 3 k, 5 k, 10 k, 15 k, and 30 k radius to find the best determination of urban development. The same calculations for percent urban development were made for each of the TCEQ monitoring sites.
Statistics
All statistical analyses were conducted using R 3.1.2 (R Core Team 2014).
The percent urban development value for each site was regressed against the estimated atmospheric NOx. This was repeated for each of the radii listed to evaluate which radius was the best indicator of urban development, based on R2 value. This analysis was also conducted at all radii for the measured atmospheric NOx at the TCEQ monitoring sites to determine the accuracy of our estimated NOx values.
Site averages were calculated for each variable examined, and percent herbivory was arcsine square-root transformed. A Pearson’s R correlation matrix was constructed using R package psych (Revelle 2014) to examine relationships between relevant soil characteristics (NO3-N, NH4-N, and pH), site characteristics (percent urban development and hourly NOx) and plant characteristics (DBH, leaf C:N, and percent herbivory). Soil NO3-N and soil NH4-N were summed to create the variable “soil N” as an indicator of total available soil N and used in the remainder of the analyses.
For the response variables leaf C:N and leaf herbivory, we wanted to examine the effects of multiple predictor variables and their potential interactions. The multiple linear regression was conducted in R and the significance of terms was determined using Type III sums of square from function “Anova” in R package car (Fox et al. 2009). For leaf C:N ratio, we started with a model that included NOx, soil N, and the interaction between NOx and soil N, as well as the covariates DBH and soil pH. Soil pH and DBH were included as covariates because of their influence on nutrient availability and primary productivity, respectively (Aber and Melillo 2001). We then used automated stepwise model section using exact AIC as the model selection criterion (function “step AIC” in R package MASS; Venables and Ripley 2002). This eliminated nonsignificant terms DBH and soil pH from the model, leaving us with a final model that included only soil N, NOx, and their interaction.
For leaf herbivory, we used a similar multiple linear regression process as with leaf C:N using the predictor variables leaf C:N, percent urban, and their interaction, along with tree DBH as a covariate. Using the same stepwise process, tree DBH was eliminated. The final model included leaf C:N, percent urban, and their interaction.
Results
NOx: Spatial and temporal patterns
Overall, the relationship between urban development and estimated atmospheric NOx was positive and linear, though the strength of the relationship varied depending on the radius of the area calculated. At radii of 3 k, 5 k, 10 k, 15 k, and 30 k, the relationship between NOx and urban development was highly significant (p < 0.001), with R2 values of 0.35, 0.58, 0.69, 0.66, and 0.62 respectively. The strongest relationship was found at the 10 k radius (y = 8.9x + 6.1, adjusted R2 = 0.66, F(1,9) = 20.74, p = 0.001; Fig. 2). The measured atmospheric NOx at the TCEQ monitoring sites was also highly correlated with the percentage of urban development at the 10 k radius (adjusted R2 = 0.61, F(1,12) = 18, p < 0.001), confirming the relationships documented for the survey sites. Thus, we used the percent urban development at the 10 k radius as our metric of urban development for the remainder of the analyses.

Plot of average hourly atmospheric NOx values in 2013 for each of the sites against the calculated percent urban development of that site, y = 8.9x + 6.1, adjusted R2 = 0.66, F(1,9) = 20.74, p = 0.001
The overall hourly average of atmospheric NOx in 2013 for each of the sites ranged from 4 to 12.9 ppbv (Table 1). As sites increased in percent urban development, atmospheric NOx also increased with the exception of two sites: the median atmospheric NOx value belonged to RLP - the site with the highest urban development - and the fourth highest atmospheric NOx value belonged to the fourth most rural site – FWN (Table 1, Fig. 2). There were no characteristics of these sites that would immediately explain why they are outliers.
From an hourly perspective, atmospheric NOx peaked at hour 7, coinciding with morning rush hour traffic (Fig. 3). A second longer lasting, but lower magnitude increase began at 17 h (coinciding with evening rush hour), peaked around 21 h, and decreased to the morning low by hour 1. The lowest hourly NOx values were found during the hottest part of the day from 13 to 16 h. The intensity of the daily peaks was for the most part dependent on the percentage of urban development for each site. For example, the diurnal range in hourly atmospheric NOx averages for EMP, the site at the 1st quartile of urban development (henceforth used as an example of a low urban development site), was 9.4 ppbv while the site at the 3rd quartile of urban development, MCP (henceforth used as an example of a high urban development site), had an hourly atmospheric NOx range of 14.2 ppbv. Throughout the day, atmospheric NOx values at the less urban sites fell below the value of the median urban development site (LLA = 41.8% urban development), and the more urban sites had values above the median site. However, as noted above, FWN, the eighth most urban site, did not follow this trend, and tended to have one of the highest hourly atmospheric NOx values (Fig. 3).

Average hourly NOx values in 2013 for each survey site. The dotted line with triangles represents the site with the median percent urban development. Solid lines represent the most urban sites with the darkest being the most urban. Hashed lines represent sites with less urban development with the darkest being the lowest percent urban
Seasonally, the highest atmospheric NOx values occur during the colder winter months, while the lowest occur during the warmer summer months (Fig. 4). Regardless of season, rural sites generally had lower atmospheric NOx values compared to urban sites. The one exception again is the FWN site, which had a lower urban development value, but atmospheric NOx values above those of the median site (Fig. 4). The magnitude of seasonal changes in average hourly atmospheric NOx is again closely related to the percentage of urban development for each site. EMP had an average hourly low of 4.7 ppbv in June and a high of 14.0 ppbv in December, while MCP had an average hourly low of 5.4 ppbv in June and 21.4 ppbv in December (Fig. 4).

Average monthly NOx values in 2013 for each survey site. The dotted line with triangles represents the site with the median percent urban development. Solid lines represent the most urban sites with the darkest being the most urban. Hashed lines represent sites with less urban development with the darkest being the lowest percent urban
Soil characteristics
The average soil pH for all sites was 7.6 ± 0.1, and soil pH was not correlated with atmospheric NOx, soil NO3-N, or soil NH4-N. The averages for soil NO3-N and soil NH4-N for all sites were 13.2 ± 0.4 and 9.3 ± 1.4 ppm, respectively. For combined soil N, EMP (low urban) fell below the mean and MCP (high urban) above the mean, however, there were no clear trends across the sites (Table 1). Soil Ca, Mg, S, and Na were also measured at each site in addition to pH, but there were no significant relationships between them or with the other measured variables (data not shown). There was no relationship between soil N and atmospheric NOx across the sites (Table 1; Fig. 5).

Correlation matrix and plot for percent urban development, NOx (ppm), leaf C:N ratio, soil nitrate (ppm), soil ammonium (ppm), soil ammonium (ppm), total available soil N (ppm), tree DBH (cm), soil pH, and percent leaf herbivory with Holm-adjusted p values. Only p values less than 0.05 are shown and bolded
Plant characteristics
Site averages for DBH ranged from 31 to 71 cm (Table 1). The westernmost sites tended to have the smallest diameter trees (Fig. 1; Table 1), but there was no statistical relationship between longitude and DBH. There was also no relationship between DBH and leaf tissue quality.
Leaf C:N ratio did not vary widely across the study sites. The average for all sites was 26.0 ± 0.4 with EMP (low urban) and MCP (high urban) both at a C:N ratio of 27 (Table 1). There were no significant correlations between leaf C:N and other measured variables. However, the multiple linear regression revealed a significant interaction between the effects of atmospheric NOx and soil N on leaf C:N (Table 2). In order to visualize this relationship, the sites were divided at the median atmospheric NOx level (9.8 ppbv) into low- and high-NOx categories containing 6 and 5 sites, respectively. At low levels of atmospheric NOx, leaf C:N and soil N were negatively correlated, indicating that tissue quality increased with increasing soil N while at high levels of atmospheric NOx, leaf C:N and soil N were positively correlated, indicating that tissue quality decreased with increasing soil N (Fig. 6). This relationship was particularly clear for the sites with the three lowest and three highest values of NOx, (Fig. 6). We ran the same linear regression model with percent urban development in place of NOx, and found no relationship.

Plot of leaf C:N as a function of soil N at two levels of NOx based on the median concentration. Sites were divided into high-NOx (greater than the median of 9.8 ppbv, represented in black) and low-NOx (less than the median of 9.8 ppbv, represented in gray). Black squares represent the three highest NOx sites and gray squares represent the three lowest NOx sites
Herbivory
The average leaf herbivory was 7 ± 0%, and the majority of sites had herbivory levels of less than 10%. The two exceptions to this were MTC (13%) and RLP (20%), the two most urban sites (Table 1). The representative low urban site had lower percent herbivory than the representative urban site (EMP = 5% and MCP = 8%, respectively), however, there were no clear trends in herbivory across the entire gradient. We did not see a relationship between herbivory and atmospheric NOx or with any of the other measured variables (Fig. 5).
Discussion
Atmospheric NOx increased along a gradient of increasing urban development
The strong positive relationship between urban development and atmospheric NOx concentrations seen in cities across the globe (Gao 2007; Hagemann et al. 2014; Xie et al. 2016) was also confirmed for NTX. However, atmospheric NOx levels in NTX were lower than those documented in other urban areas. In Nanjing, China, hourly NOx concentrations during the summer months of 2008 averaged 19 ppbv and hourly NOx concentrations during the winter months averaged 35 ppbv (Xie et al. 2016). In contrast, the hourly NOx concentrations for NTX site MTC averaged 7 ppbv during the summer months of 2013 and 20 ppbv during the winter months of 2013. In the eastern U.S. (New Jersey), the hourly NOx average at 7 am (during morning rush hour) in the spring of 2007 was 60 ppbv (Song et al. 2011), whereas the NOx concentration at 7 am in the spring of 2013 at MTC was 13 ppbv. Though urban atmospheric NOx concentrations in the U.S. have been on the decline in the past decade (Lu et al. 2015), this decline does not account for the magnitude of the differences between NTX NOx concentrations in 2013 and other cities in 2007–2008. Differences in topography and wind patterns could be contributing to the lower values of NOx in NTX relative to other cities.
As expected, the daily rise and fall of atmospheric NOx closely followed morning and evening rush hour traffic (Fig. 3), as approximately 50% of NOx emissions in NTX were attributed to vehicular sources in 2015 (TCEQ 2015b). As in other US cities, the morning peak in atmospheric NOx is likely strengthened by the atmospheric inversion that occurs overnight (Song et al. 2011). As the evening rush hour occurs, NOx concentrations again increase, though the peak is smaller in the evening due to greater solar radiation and photochemical reactions in the afternoon (Fig. 3).
The seasonal variation in atmospheric NOx in NTX also followed the patterns of NOx in US and other global cities (Vellingiri et al. 2015a; Xie et al. 2016; Zhang et al. 2003) with lower hourly averages in the summer and the highest hourly averages in the winter (Fig. 4). Higher winter atmospheric NOx relative to summer levels may be attributed to a combination of factors including lower solar radiation and less photochemical activity, increased consumption of fuels, and poorer dispersion conditions.
The diurnal and seasonal patterns of NOx can be seen at each site, though the magnitude of the peaks was much greater at the sites with greater urban development. MTC, the second most urban site, showed the greatest variation in average hourly (Fig. 3) and seasonal (Fig. 4) atmospheric NOx, while the range of NOx values for KSP (the least urban) was 45% smaller for the peak hourly averages and 25% smaller for the peak seasonal average. This suggests that plants at the more urban sites experience a “pulse” of N input during peak atmospheric NOx seasons rather than a relatively constant low-level input as found at more rural sites.
Soil nitrogen did not increase with greater NOx
We hypothesized that the effects of cumulative atmospheric NOx additions would result in elevated soil N via chronic low-level N deposition. However, our study revealed no correlations between atmospheric NOx, soil NO3-N, or soil NH4-N (Fig. 5). Our findings may be the result of the relatively small increase in atmospheric NOx in NTX urban areas (relative to other major metropolitan areas, discussed above), or it may be that the signal has been obscured by other processes or transformations that occur in the urban N cycle. We discuss some of these possibilities below.
If the atmospheric NOx does reach the ground in the form of N deposition, it may not be captured in a springtime measurement of extractable soil N. Various soil processes such as immobilization and denitrification of N by bacteria may mean that very little of the added anthropogenic N is available to plants. There is also evidence from other US cities that N can move directly from deposition to leaching without any intermediate biological processes (Rao et al. 2013). The fact that our sites are in close proximity to the Trinity River and the sampling occurred just after the annual spring rains raises the possibility that these NTX sites experience rapid NO3-N leaching. Although we measured soil N during the period of active growth in post oak, the seasonal nature of atmospheric NOx levels may mean that annual N mineralization is better correlated with NOx.
Effect of soil N on leaf C:N depends on NOx
We did not see the same positive relationship between soil N and leaf tissue quality that has been documented in reviews of fertilization experiments (Phoenix et al. 2012; Yang et al. 2011b). Most fertilization experiments use much higher levels of simulated N deposition than are realistic for many areas, thus the amount of N deposited across our gradient was likely much lower than the amounts used in fertilization experiments. In our results, however, there was an effect of the interaction between soil N and atmospheric NOx on leaf C:N. At low levels of NOx, the positive correlation between greater soil N availability and greater tissue quality occurred as expected. At levels of NOx above the median site value, the relationship reversed. This suggests that the usual positive correlation between soil and plant N concentrations is decoupled when NOx and associated influences of the urban environment are greater.
One possible explanation is that that the elevated temperatures associated with urban areas are confounding the relationship between soil N and foliar C:N ratios. Red oak (Quercus rubra) grown along an urban to rural transect in New York, U.S., and in growth chambers showed differential responses to N availability based on night-time and inter-annual differences in temperature (Searle et al. 2012). While the range in annual average temperature of our sites is only 1 °C, it is possible that the elevated night-time temperatures associated with urban areas (George et al. 2007) are influencing either plant nitrogen uptake or allocation.
Herbivory not correlated with greater NOx
We did not see a relationship between atmospheric NOx and herbivory, nor did we see the expected intermediary relationship between leaf C:N and herbivory. However, there was a trend toward greater herbivory with increasing urban development, with the two most urban sites having the greatest levels of herbivory. Raupp et al. (2010) offer several thoughts on the possible mechanisms behind elevated herbivory in urban areas: the reduction of native plant diversity in urban environments may contribute to the reduction of natural enemies; elevated temperatures in developed areas may favor an increased population of herbivores; and the (non-native) vegetative complexity may provide more opportunities to hide from predators. These mechanisms may be at play among the sites we studied, but only significant in the most urban locations.
In NTX, chewing insects commonly found on post oak trees include leaf-cutting bees (Megachilidae), katydids (Tettigoniidae), luna moths (Actias luna), and other Lepidoptera larvae, some of which have the potential to cause significant damage (Texas A&M Forest Service 2015). Insects can affect N cycling by consuming leaf tissues and depositing frass, although the magnitude of this effect is not well known in urban ecosystems (Raupp et al. 2010). Gaining a greater understanding of the interactions between the urban environment in NTX and insect herbivores will be important for establishing a baseline as development and climate change persist.
Conclusions
This study adds to the recently expanding body of literature examining how urban ecosystems are affected (or not) by N deposition and suggests that interactions among NOx, soils, and plants are complex and sometimes, counterintuitive. We found that atmospheric NOx was highly correlated with percentage of urban development in NTX, as in other cities. The temporal patterns affirm that in this region, atmospheric NOx were driven by combustion of fossil fuels in automobiles. However, the atmospheric NOx concentrations were somewhat lower than those reported in other major metropolitan areas. Despite the positive correlation between urban development and NOx across our sites, we found no relationship between atmospheric NOx and soil N, suggesting a minimal effect of NOx on these soils. Leaf tissue quality was positively related to soil N as expected, but only at low NOx values. This suggests that at higher NOx values, the expected correlation between soil and plant N concentration is influenced by other factors that covary with urbanization. Herbivory was not correlated with leaf tissue quality, but there was a trend toward greater herbivory with greater urban development.
This research also highlights the need for further examination of N deposition components and quantities in NTX. There is a growing body of evidence suggesting a shift from primarily NO3-N deposition in urban areas to a greater proportion of NH4-N (Du et al. 2014; Li et al. 2016; Rao et al. 2013). Thus, it is possible that the elevated atmospheric NOx concentrations associated with urban development are not a good indicator of overall N deposition in large, sprawling urban areas.
Studies such as this one, done on a local scale (50-100 km), are vitally important in linking ecosystem processes to urbanization patterns (Brazel et al. 2000). The rapid growth and sprawling and heterogeneous nature of the urban development in NTX provides an opportunity to expand our understanding of urban areas still being actively developed and those that are relatively young. Additionally, the location of NTX in the sub-humid, sub-tropical southern Great Plains also provides the opportunity to further explore the dynamic feedback between climate and urbanization as Texas is predicted to be one of the hardest hit U.S. states in future climate change scenarios (Melillo et al. 2014). The higher temperatures, irregular rainfall, and more dry-days predicted for NTX may further inhibit urban forest C and N retention and lead to accelerations of C and N losses from the terrestrial ecosystem (Bai et al. 2015). Future examinations of urban ecosystems in NTX will be particularly useful in expanding general patterns that can be used to predict and potentially mitigate ecological responses to the urban environment.
References
Aber JD, Melillo JM (2001) Terrestrial ecosystems, Second edn. Academic Press, San Diego
Aronson MFJ, Handel SN, Puma IP, Clemants SE (2014) Urbanization promotes non-native woody species and diverse plant assemblages in the New York metropolitan region. Urban Ecosystems 18:31–45. doi:10.1007/s11252-014-0382-z
Bai SH, Xu Z, Blumfield TJ, Reverchon F (2015) Human footprints in urban forests: implication of nitrogen deposition for nitrogen and carbon storage. J Soils Sediments 15:1927–1936. doi:10.1007/s11368-015-1205-4
Bedison JE, McNeil BE (2009) Is the growth of temperate Forest trees enhanced along an Abient nitrogen deposition gradient? Ecology 90:1736–1742
Bobbink R, Roelofs JGM (1995) Ecological effects of atmospheric deposition on non-forest ecosystems in Western Europe. In: Heij GJ, Erisman JW (eds) Studies in Environmental Science, vol 64. Elsevier, pp 279–292. https://www.elsevier.com/books/acid-rain-research-do-we-have-enoughanswers/erisman/978-0-444-82038-9
Bobbink R et al (2010) Global assessment of nitrogen deposition effects on terrestrial plant diversity: a synthesis. Ecol Appl 20:30–59. doi:10.1890/08-1140.1
Boggs JL, McNulty SG, Gavazzi MJ, Myers JM (2005) Tree growth, foliar chemistry, and nitrogen cycling across a nitrogen deposition gradient in southern Appalachian deciduous forests. Can J For Res 35:1901–1913. doi:10.1139/x05-128
Brazel A, Selover N, Vose R, Heisler G (2000) The tale of two climates - Baltimore and phoenix urban LTER sites. Clim Res 15:123–135
Butler TJ, Likens GE, Vermeylen FM, Stunder BJB (2003) The relation between NOx emissions and precipitation NO3− in the eastern USA. Atmos Environ 37:2093–2104. doi:10.1016/s1352-2310(03)00103-1
Butler TJ, Likens GE, Vermeylen FM, Stunder BJB (2005) The impact of changing nitrogen oxide emissions on wet and dry nitrogen deposition in the northeastern USA. Atmos Environ 39:4851–4862. doi:10.1016/j.atmosenv.2005.04.031
Chapin FS III (1980) The mineral nutrition of wild plants. Annu Rev Ecol Syst 11:233–260
Diggs GM, Lipscomb BL, O'Kennon RJ, Mahler WF, Shinners LH (1999) Shinners' and Mahler's illustrated flora of north Central Texas. Botanical Research Institute of Texas, Fort Worth
Du E, de Vries W, Galloway JN, Hu X, Fang J (2014) Changes in wet nitrogen deposition in the United States between 1985 and 2012. Environ Res Lett 9:095004. doi:10.1088/1748-9326/9/9/095004
Elliott EM et al (2007) Nitrogen isotopes as indicators of NOx source contributions to atmospheric nitrate deposition across the Midwestern and northeastern United States. Environ Sci Technol 41:7661–7667. doi:10.1021/es070898t
Fox J et al. (2009) CAR: Companion to applied regression, R Package version 1.2–16. http://cran.r-project.org/web/packages/car/index.html. Accessed August 2012
Galloway JN et al (2008) Transformation of the nitrogen cycle: recent trends, questions, and potential solutions. Science 320:889–892. doi:10.1126/science.1136674
Gao HO (2007) Day of week effects on diurnal ozone/NOx cycles and transportation emissions in Southern California. Transp Res Part D: Transp Environ 12:292–305. doi:10.1016/j.trd.2007.03.004
George K, Ziska LH, Bunce JA, Quebedeaux B (2007) Elevated atmospheric CO2 concentration and temperature across an urban–rural transect. Atmos Environ 41:7654–7665. doi:10.1016/j.atmosenv.2007.08.018
Griffith G, Bryce S, Omernik J, Rogers A (2007) Ecoregions of Texas. Texas Commission on Environmental Quality, Austin
Grimm NB et al (2008) The changing landscape: ecosystem responses to urbanization and pollution across climatic and societal gradients. Front Ecol Environ 6:264–272. doi:10.1890/070147
Hagemann R, Corsmeier U, Kottmeier C, Rinke R, Wieser A, Vogel B (2014) Spatial variability of particle number concentrations and NOx in the Karlsruhe (Germany) area obtained with the mobile laboratory ‘AERO-TRAM’. Atmos Environ 94:341–352. doi:10.1016/j.atmosenv.2014.05.051
Homer CG et al (2015) Completion of the 2011 National Land Cover Database for the conterminous United States - representing a decade of land cover change information. Photogramm Eng Remote Sens 81:345–354
Kahan A, Currie W, Brown D (2014) Nitrogen and carbon biogeochemistry in Forest sites along an indirect urban–rural gradient in southeastern Michigan. Forests 5:643–665
Lawrence GB, Goolsby DA, Battaglin WA, Stensland GJ (2000) Atmospheric nitrogen in the Mississippi River basin — emissions, deposition and transport. Sci Total Environ 248:87–100. doi:10.1016/S0048-9697(99)00533-1
Lebauer DS, Treseder KK (2008) Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology 89:371–379
Li Y, Schichtel BA, Walker JT, Schwede DB, Chen X, Lehmann CMB, Puchalski MA, Gay DA, Collett JL (2016) Increasing importance of deposition of reduced nitrogen in the United States. Proc Natl Acad Sci 113(21):5874–5879. doi:10.1073/pnas.1525736113
Likens GE, Buso DC, Butler TJ (2005) Long-term relationships between SO2 and NOx emissions and SO42- and NO3- concentration in bulk deposition at the Hubbard brook experimental Forest, NH. J Environ Monit 7:964–968. doi:10.1039/B506370A
Lu Z, Streets DG, de Foy B, Lamsal LN, Duncan BN, Xing J (2015) Emissions of nitrogen oxides from US urban areas: estimation from ozone monitoring instrument retrievals for 2005–2014. Atmos Chem Phys Discuss 15:14961–15003. doi:10.5194/acpd-15-14961-2015
McNeil BE, Read JM, Driscoll CT (2007) Foliar nitrogen responses to elevated atmospheric nitrogen deposition in nine temperate Forest canopy species. Environ Sci Technol 41:5191–5197. doi:10.1021/es062901z
Melillo JM, Richmond TT, Yohe G (2014) Climate Change Impacts in the United States: The Third National Climate Assessment. doi:10.7930/J0Z31WJ2
National Oceanic and Atmospheric Administration (2016) U.S. Climate Divisions. http://www.ncdc.noaa.gov/monitoring-references/maps/us-climate-divisions.php. Accessed August 2015
Pan J, Widner B, Ammerman D, Drenovsky R (2010) Plant community and tissue chemistry responses to fertilizer and litter nutrient manipulations in a temperate grassland. Plant Ecol 206:139–150. doi:10.1007/s11258-009-9630-3
Phoenix GK et al (2012) Impacts of atmospheric nitrogen deposition: responses of multiple plant and soil parameters across contrasting ecosystems in long-term field experiments. Glob Chang Biol 18:1197–1215. doi:10.1111/j.1365-2486.2011.02590.x
Pickett STA et al (2011) Urban ecological systems: scientific foundations and a decade of progress. J Environ Manag 92:331–362. doi:10.1016/j.jenvman.2010.08.022
R Core Team (2014) R: A language and environment for statistical computing. R Foundation for Statistical Computing. http://www.R-project.org/
Rao P, Hutyra L, Raciti S, Templer P (2013) Atmospheric nitrogen inputs and losses along an urbanization gradient from Boston to Harvard Forest, MA. Biogeochemistry:1–17. doi:10.1007/s10533-013-9861-1
Raupp MJ, Shrewsbury PM, Herms DA (2010) Ecology of herbivorous arthropods in urban landscapes. Annu Rev Entomol 55:19–38. doi:10.1146/annurev-ento-112408-085351
Redling K, Elliott E, Bain D, Sherwell J (2013) Highway contributions to reactive nitrogen deposition: tracing the fate of vehicular NOx using stable isotopes and plant biomonitors. Biogeochemistry 116:261–274. doi:10.1007/s10533-013-9857-x
Revelle W (2014) psych: Procedures for personality and psychological research. https://cran.r-project.org/web/packages/psych/index.html. Accessed April 2016
Searle SY, Turnbull MH, Boelman NT, Schuster WSF, Yakir D, Griffin KL (2012) Urban environment of New York City promotes growth in northern red oak seedlings. Tree Physiol 32:389–400. doi:10.1093/treephys/tps027
Seto KC, Guneralp B, Hutyra LR (2012) Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools. Proc Natl Acad Sci U S A 109:16083–16088. doi:10.1073/pnas.1211658109
Shen W, Wu J, Grimm N, Hope D (2008) Effects of urbanization-induced environmental changes on ecosystem functioning in the phoenix metropolitan region, USA. Ecosystems 11:138–155. doi:10.1007/s10021-007-9085-0
Song F, Young Shin J, Jusino-Atresino R, Gao Y (2011) Relationships among the springtime ground–level NOx, O3 and NO3 in the vicinity of highways in the US East Coast. Atmos Pollut Res 2:374–383. doi:10.5094/APR.2011.042
TCEQ (2015a) Air Monitoring Sites. http://www17.tceq.texas.gov/tamis/index.cfm?fuseaction=home.welcome. Accessed April 2014
TCEQ (2015b) Texas Five-Year Ambient Monitoring Network Assessment
Texas A&M Forest Service (2015) Forest Health: Insects. http://texasforestservice.tamu.edu/foresthealth/insects/. Accessed February 2016
Texas Water Deveopment Board (2012) Water for Texas 2012 State Water Plan. Texas
U.S. Census Bureau (2016) Annual Estimates of the Resident Population: April 1, 2010 to July 1, 2015
Vellingiri K, Kim K-H, Jeon JY, Brown RJC, Jung M-C (2015a) Changes in NOx and O3 concentrations over a decade at a central urban area of Seoul, Korea. Atmos Environ 112:116–125. doi:10.1016/j.atmosenv.2015.04.032
Venables W, Ripley B (2002) Modern applied statistics using S. Springer, New York
Vision North Texas (2008) Regional Choices Report. http://www.visionnorthtexas.org/regionalchoices/regionalchoices.html. Accessed April 2016
Vitousek PM et al (1997) Human alteration of the global nitrogen cycle- sources and consequences. Ecol Appl 7:737–750
Xia J, Wan S (2008) Global response patterns of terrestrial plant species to nitrogen addition. New Phytol 179:428–439. doi:10.1111/j.1469-8137.2008.02488.x
Xie M et al (2016) Temporal characterization and regional contribution to O3 and NOx at an urban and a suburban site in Nanjing, China. Sci Total Environ 551–552:533–545. doi:10.1016/j.scitotenv.2016.02.047
Yang H, Li Y, Wu M, Zhang ZHE, Li L, Wan S (2011a) Plant community responses to nitrogen addition and increased precipitation: the importance of water availability and species traits. Glob Chang Biol 17:2936–2944. doi:10.1111/j.1365-2486.2011.02423.x
Yang Y, Luo Y, Lu M, Schädel C, Han W (2011b) Terrestrial C:N stoichiometry in response to elevated CO2 and N addition: a synthesis of two meta-analyses. Plant Soil 343:393–400. doi:10.1007/s11104-011-0736-8
Zhang R, Tie X, Bond DW (2003) Impacts of anthropogenic and natural NOx sources over the US on tropospheric chemistry. Proc Natl Acad Sci 100:1505–1509
Acknowledgements
This research was supported by a grant from the Beta Phi Chapter of the Phi Sigma Biological Honors Society at UTA. Thank you to Dr. Thomas Chrzanowski of UTA, Dr. Melanie Sattler of UTA, Gautam Raghavendra of UTA and Jayme Walton of SWCA Environmental Consultants.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Green, M.L., Foster, K. & Gough, L. Urban development in the southern Great Plains: effects of atmospheric NOx on the long-lived post oak tree (Quercus stellata). Urban Ecosyst 20, 651–661 (2017). https://doi.org/10.1007/s11252-016-0622-5
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11252-016-0622-5