
Abstract
This study aimed to evaluate forage production and ingestive behavior of Morada Nova sheep in an intensive system with capim-tamani grass in both monoculture and silvopastoral irrigated systems in the Semi-arid region. Eighteen adult sheep, approximately 3 years old, with an average body weight of 26.8 ± 4.3 kg, were allocated to treatments with capim-tamani grass cultivated in monoculture and in silvopastoral systems with Caatinga trees. The experiment followed a completely randomized complete block design with two plots and three replications. Forage production, consumption, and behavioral activities were the assessed variables. The animals remained in the pasture daily between 6 am and 6 pm. No effects of the monoculture and silvopastoral systems were observed on the structural and productive characteristics of the capim-tamani grass pasture. There were also no observed effects on body condition score, consumption, and disappearance rate of dry matter (DM) and other nutrients in both systems. However, there was an interaction between the time animals spent under shade in monoculture and silvopastoral systems. In general, the animals spent more time under shade where there were trees, except during the period between 2 pm and 4 pm, when the times were similar. On average, the animals spent approximately 15.6% (equivalent to 1.87 h) of their total time in the artificial shade available in the monoculture system, whereas in the silvopastoral system, they remained under natural shade for approximately 40% (five and a half hours) of their time spent in the pasture during the day. The grazing frequency in both systems was approximately 70% (8.4 h) in relation to the total time spent in the pasture. The capim-tamani grass pasture managed intensively with sheep in the silvopastoral system showed similar forage production and consumption compared to the monoculture system. There was a positive influence of trees on the duration of solar radiation exposure to the animals throughout the day. The results support the need to provide shade for sheep, as well as to promote these sustainable systems in semi-arid regions.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Silvopastoral systems advocate for the integrated exploitation of forestry (native or exotic) and livestock, emerging as an alternative to mitigate the imminent effects of pasture degradation (Oliveira et al. 2021a). Simultaneously, these systems provide shaded areas, contributing to improved thermal comfort and animal well-being (Santos et al. 2021).
In Brazil, the semiarid region covers an area of 1,128,697 km2, making it the largest in the world. Additionally, it is a region that houses cattle, goats, and sheep, with these three species totaling a herd of nearly 40 million heads (IBGE 2020). The predominant vegetation in this region is the Caatinga, which exhibits a rich diversity of herbaceous, shrubby, and arboreal plants, occupying 11% of the national territory (Alves et al. 2009). Given this scenario, the adoption of integrated systems is an appropriate tool for the exploration of sustainable productive systems, in order to maintain arboreal biodiversity with minimal environmental impact.
However, in field conditions, the success of systems with natural shading has been limited by the choice of tree component spatial arrangements which provide a microclimate for the animals and promote adequate productivity and persistence of the forage plant (Santos et al. 2016). Intense levels of shading lead to a significant reduction in the quantity and quality of light, resulting in morphophysiological and productive changes that hinder the development of plants in their growth environment, affecting their efficiency of use, as well as animal performance (Lima et al. 2019; Paciullo et al. 2011). In this context, it becomes important to understand the relationship between light intensity and the choice of forage species.
The capim-tamani (Megathyrsus maximus cv. BRS Tamani) displays noteworthy attributes, such as a high rate of leaf and tiller emergence, along with high nutritional value. Additionally, it is well adapted to the soil and climatic conditions of tropical regions (Empresa 2015). Its low height allows grazing by various animal categories, including sheep. However, its use is particularly recommended in intermittent stocking systems, although continuous stocking can be considered as a management alternative, especially in conditions where there is limited availability for nitrogen fertilization at higher intensities and frequencies (Jank and Santos 2021). As there have been few studies on capim-tamani in conjunction with trees and animals, particularly considering its recent release, notable works include those by Pereira et al. (2015, 2021), evaluating a shading level of 56% (277 trees. ha− 1), during the establishment and production phases; Santos et al. (2019), who worked with 42% shading, analyzing tillering and forage production; and Oliveira et al. (2021b), who assessed two shading levels (357 and 588 trees. ha-1) on yield and nutritive value. In these studies, Tamani grass demonstrated potential for exploitation in integrated systems. However, further investigations into all production niches (i.e., animal production) are necessary.
Considering the annual seasonality of forage production as a limiting factor for ruminant production in the tropics, the development of new forage species in intensive systems subjected to fertilization, especially nitrogen, combined with irrigation techniques, has become an important tool to enhance the productive potential of pastures (Sanches et al. 2015; Vasconcelos et al. 2020).
The adoption of silvopastoral systems with sheep farming in the semi-arid region can become an important tool for the ovine industry to achieve production levels compatible with the potential the region has to offer. This approach can help meet market demands for high-quality products. In this context, it is crucial to emphasize the importance of using local breeds, which are viable alternatives for efficient and sustainable production systems (Nunes et al. 2020).
Therefore, the objective of this study was to evaluate forage production and ingestive behavior of Morada Nova sheep in an intensive system with capim-tamani grass in both monoculture and irrigated silvopastoral systems in the Semi-arid region.
Materials and methods
For the procedures conducted in this study, ethical approval was obtained and the research was carried out following the guidelines of the Ethics Committee on Animal Use at the State University of Vale do Acaraú, Ceará, Brazil (Protocol No. 003.08.021.UVA.504.02). The manuscript does not contain clinical studies or patient data.
Study location and area characterization
The experimental period took place from August 2022 to January 2023 at the Experimental Farm of the State University of Vale do Acaraú/UVA, located in Sobral, Ceará, Brazil (3° 36’ South, 40° 18’ West, and 56 m above sea level).
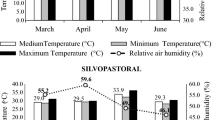
The climate of the region is classified as BSh (hot semi-arid with a monthly average temperature above 18 °C) according to the Köppen climate classification (Alvares et al. 2013). During this period, the region experienced minimum, average, and maximum temperatures of 25.8, 32.9, and 41.5 °C, respectively. The average temperatures (°C) and relative humidity (%), recorded during the experimental period, are presented in Fig. 1.

Average distribution of temperatures and relative humidity throughout the experimental period in irrigated Monoculture and Silvopastoral systems in the Semiarid. AT (ºC). ambient temperature; RH (%). relative humidity; MO. Monocultive System; SPS. Silvopastoral System
The total area of the experiment was 7,200 m², divided into 10 paddocks with field-type sheep fencing, comprising 6 experimental units and 4 reserves. The area also featured a low-pressure fixed-sprinkler irrigation system: [a fully detachable NY-25 agropolo sprinkler with a nominal diameter of ¾, spaced at 6 × 6 m, powered by a three-phase electric motor [W22 IR3 Premium 10 HP].
In 2020, manual thinning was carried out in the experimental area. In the monoculture paddocks, 100% of the vegetation was previously suppressed, while in the silvopastoral system, 8 trees per paddock were retained, corresponding to 104 trees. ha− 1, preserving them randomly in their natural condition. Before thinning, the experimental area consisted of native arboreal Caatinga. During the same period, enrichment with the grass Megathyrsus maximus cv. BRS Tamani was carried out, established in all experimental units through direct broadcasting. A phytosociological survey was conducted, identifying the following tree species: Aspidosperma pyrifolium Mart. & Zucc (pereiro, n = 2); Dipteryx odorata (cumaru, n = 6); Erythrina verna (mulungu, n = 1); Libidia ferrea (jucá or pau-ferro, n = 1); Mimosa caesalpiniaefolia (sabiá, n = 1); and Auxemma oncocalys (pau-branco, n = 13).
A soil analysis of the experimental area was performed, where composite samples (0–20 cm depth) were taken to evaluate fertility: The attributes pH; organic matter; P; K; Ca; Mg and Al were interpreted as follows (Table 1): Weak acidity; Low; Medium; Medium; High and Low, according to Alvarez et al. (1999).
Treatments and experimental design
The experimental design used was a complete randomized block design with two plots and three replications. The treatments consisted of the production of capim-tamani grass in a monoculture system and an irrigated silvopastoral system.
In the monoculture experimental area, for ethical considerations regarding animal welfare, it included a shaded area provided by a black polypropylene screen with 80% light retention, covering an area of 15 m². This corresponds to a shade offering of 2.5 m² per animal (Oliveira et al. 2013).
To calculate the coverage of the tree component, a graduated measuring tape was used to measure the radius of the tree canopies at cardinal points from the center of the trunk, with an average radius of 2.62 m. Given the formula A = πr², at the zenith, the canopy projection occupied 22.5% of the paddock, it corresponds to a shade offering of 7,2 m² per animal. The average diameter at breast height (DBH; 1.30 m) was measured at 25.62 cm, determined from the tree trunk circumference and then transformed through the relationship. \(DBH= CBH/{\pi }\), in which CBH is the circumference at breast height. The average height of the tree trunk was 7.4 m.
Animals and management
Before the start of the experiment, a uniform mowing of the paddocks was conducted, cutting at a height of 10 cm from the soil surface. After mowing, the height was monitored three times a week by measuring 30 points in each experimental unit, using the recommended height criterion for animal entry into the paddocks. A total of six growth cycles of the capim-tamani grass were assessed during the experimental period. Initially, each cycle was set at 25 days, with an average management height of 26 cm + 02 cm, in accordance with the level of nitrogen fertilization (urea, 45% N) established in 150 kg ha− 1 year− 1 in accordance with Vasconcelos et al. (2020), suitable for a system with moderate intensification (Pereira et al. 2018). The fertilization was split into 12 applications, always at the beginning and middle of each production cycle, and irrigation was carried out based on the availability of five days per week, during the nighttime. This was done to minimize water loss due to drift as well as potential nitrogen losses through volatilization, given the high temperatures during the day.
Eighteen dry Morada Nova sheep, approximately 3 years old, with an initial average body weight of 26.8 + 4.3 kg, were used in the experiment. The sheep were managed under continuous stocking with a variable stocking rate. The animals remained in the paddocks daily for a period of 12 h (morning and afternoon), starting at 6 am and ending at 6 pm. After this period, they were directed to the handling center throughout the night.
An initial average stocking rate of three animals per paddock per day was adopted, estimating an average dry matter consumption of 1.93% of body weight for adult dry ewes (NRC 2007), with a forage dry matter offering of 1.3 kg per day and considering a grazing efficiency of 40% (Hodgson 1990).
Nine test animals were randomly selected for each treatment, with three in each paddock, totaling 18 test animals. Two sheep were used as regulator animals, allocated to the reserve paddocks, to ensure pasture management goals were met using the variable stocking rate, “put and take” method (Mott and Lucas 1952), by inserting or removing regulator animals from the experimental paddocks as needed.
During the experimental period, mineral salt and water were provided ad libitum in salt feeders and water troughs, respectively, appropriately installed in the paddocks.
Structural characteristics of the pasture
Structural and productive characteristics of the pasture were assessed and quantified through collections made at the end of each growth cycle of the capim-tamani grass, totaling six evaluations throughout the research. Canopy height was measured daily by sampling 30 points from representative areas in each paddock using a retractable graduated rod in centimeters (Barthram 1981). The population density of tillers (PDT) was measured by counting the tillers within metal frames (0.25 m2) placed three times in each paddock. After measuring the PDT, a cut at ground level was performed to assess the total forage biomass (TFB). Subsequently, a subsample (300 g) was taken from each paddock to evaluate the morphological components: dead forage biomass (DFB), green forage biomass (GFB), green leaf blade biomass (GLBB), and green stem biomass (GSB), as well as the live material/dead material (LM/DM) and leaf blade/stem (LB/S) ratios. Exclusion cages (03 cages paddock-1) were employed following the methodology proposed by Klingman et al. (1943), with adaptations. The cages (height = 60 cm x width = 50 cm x length = 50 cm) were built with uniformly sized sawn wood and enclosed with green nylon mesh. Structural characteristics were recorded for each cage, parallel to the frames; both were harvested and later taken to the laboratory for weighing the total forage biomass (TFB), and a subsample (300 g) was taken from each experimental unit (i.e., paddock) for the separation of morphological components: DFB, GFB, GLBB and the ratios: LM/DM, for later chemical analyses.
Both the frames and exclusion cages in the silvopastoral system followed a methodological criterion when being deployed and fixed, respectively, in the paddocks during each growth cycle of the evaluated grass culture. This was done to ensure a balance between shaded and sunny areas. At the end of each growth cycle of the Tifton grass culture (i.e., during pasture evaluations), the cages were fixed in areas under the shade of the tree component and directly exposed to the sun. The quantity of cages fixed in the shade and sun was alternated each time, and the same procedure was applied to the deployment of frames, with three deployments directed to shade and sun in the paddocks of the silvopastoral system.
The following parameters were evaluated: forage accumulation rate, forage availability, defoliation intensity, grazing efficiency, and forage utilization efficiency. Total forage accumulation (TFA) was determined by the relationship between the cage and the frame biomasses. The green forage accumulation (GFA) was quantified by excluding the dead material from TFA. The forage accumulation rate (FAR) was obtained by dividing TFA by the number of days in the production cycle of the grass culture (i.e., 25 days).
The forage availability (FA) was estimated by the ratio.
(kg of DM.100 kg of body weight− 1. day− 1). The defoliation intensity (DI) (forage disappearance) and forage utilization efficiency (FUE) were calculated according to Combellas and Hodgson (1979) and Hodgson (1990). The grazing efficiency formula was adapted for continuous stocking, as per Meneses (2022), considering the FAR and dead material for calculation purposes:\(DI= \left[\frac{\left(TFBi - TFBf\right)}{TFBf} \right]\times 100\)
In which: DI, defoliation intensity (%); TFBi, total forage biomass in pre-grazing condition (kg of DM ha− 1); TFBf, total forage biomass in post-grazing condition (kg of DM. ha− 1).
In which: GE, grazing efficiency (%); DMI, animal dry matter intake (average kg of DM.day− 1); SSR - sheep stocking rate; OP – area occupation period (days); FAR - forrage accumulation rate (kg of DM.ha.day− 1).
In which: FUE, forrage usage efficiency (%).
The sheep stocking rate (SSR) was mantained throughout the Whole experimental period with 3 sheep (40 sheep. ha− 1 ratio) in each experimental unit (e.g., paddock) of each treatment, displaying minimal alteration following the recommended management height adopted. In other words, this sheep stocking rate (SSR) of three animals in each experimental unit during grazing days (equivalent to 40 sheep per hectare) was used for GE calculations.
Forage intake and disappearance
For the determination of fecal sample collections, collecting bags were attached to the animals, which were placed on the first day of the trial and removed on the last day. Fecal collections were performed twice a day, that is, within 24 h, collections were made before the start of the grazing period (6 am) and after the grazing period (6 pm), for five consecutive days, with a total of five trials throughout the study. After collection, the samples were placed in labeled plastic bags and weighed. The total weight of the feces was determined by summing the weights of the feces collected in the morning and afternoon, then homogenized per animal, a subsample was taken, and stored in the freezer for subsequent analyses. The disappearance of dry matter (DisDM) was calculated through the difference between the weight of the ground leaf blade samples in a mill (Wiley TE-650 TECNAL®, Piracicaba, Brazil), in a 2.5 mm sieve before incubation, and the weight obtained by drying at 105 °C for 16 h after incubation (Orskov and McDonald 1979). A fixed incubation time (96 h) was used, considering the necessary digestion time for the forage (Nocek 1988). Incubation was carried out using a fistulated bovine weighing approximately 450 kg live weight. The disappearance of other nutrients was calculated by the difference in residue and its percentage in the chemical analyses. Dry matter intake (DMI) was estimated according to Prigge et al. (1981), with adaptations. The calculation was obtained by the ratio between the fecal production obtained by total fecal collection during the 24 h of the day and the inverse of DisDM, unlike the present author who uses the inverse of in vitro dry matter digestibility of the extrusa.
From the DMI the intakes of OM, CP, NDFap, NFC and TDN, multiplying the amount of dry matter consumed by the percentage of each of the assessed nutrients. Body condition scoring (BCS) was performed weekly through palpation (scale from 1 to 5; Russel 1984) for subjective evaluation of the nutritional status of the animals. In this scale, BCS 1 represents the leanest animal, and BCS 5 represents the obese animal.
Chemical analyses
The determination of the bromatological composition of the capim-tamani (Table 2) was carried out using the material present inside the exclusion cages during each growth cycle. The samples (feces and pasture) were dried in a closed-circulation oven at 65 °C for 72 h and then ground (Wiley TE-650 TECNAL®, Piracicaba, Brasil), in a 1 mm sieve. The dry matter rates (DM; method nº 934.01), organic matter (OM) (method nº 942.05), crude protein (CP) (method nº 984.13) and ether extract (EE) (method nº 920.39) were quantified according to AOAC (2012). Neutral detergent fiber (NDF) corrected for contaminating ashes and proteins (NDFa), using a thermostable α-amylase, omitting sodium sulfate, method INCT-CA F-002/1lignin in acid detergent (method INCT-CA F-005/1) and insoluble nitrogen in neutral detergent (INND) was determined following the standard analytical methods of the National Institute of Science and Technology in Animal Science (INCT-CA, Detmann et al. 2012). Non-fibrous carbohydrates (NFC) were calculated according to Detmann and Valadares Filho (2010), using the NFDap in the calculation The total digestible nutrients (TDN) was calculated according to the method proposed by Weiss et al. (1992). The NRC was estimated according to the NRC (2001): TDN = dCP + dNDF+(dEE×2.25) + dCNF, in which dCP is the digestible crude protein, dNDF is the digestible fiber in neutral detergent, dEE is the digestible ether extract and dNFC are the digestible non-fiber carbohydrates.
Behavioral responses
For behavioral assessments, there were two trials on consecutive days in each growth cycle of the capim-tamani culture, totaling twelve trials during the research. On the eve of the trials, the sheep from each treatment were numbered on the flank with non-toxic, visible spray paint. Each behavior trial lasted for 12 uninterrupted hours, corresponding to the time the animals spent in the experimental area (i.e., the period the animals were in the paddocks), starting at 6 a.m. and ending at 6 p.m. Subsequently, the following activities were recorded on spreadsheets at 10-minute intervals, resulting in six observations per hour, 72 per day, and 864 per animal during the experimental period: time spent in shade (tree or shade net projection), grazing, rumination, idle time (standing or lying down), and other random activities. Additionally, continuous activities were recorded, i.e., those occurring independently of the observation moment, including urination, drinking water, defecation, and mineral supplement ingestion. Simultaneously, the bite rate (bites min-1) was recorded using a stopwatch. The management procedures and data collection were performed by twelve properly trained professionals, with rotation between the morning and afternoon periods of each trial. For data tabulation, the decision was made to divide the data into two-hour intervals (6AM to 8AM, 8AM to 10AM, 10AM to 12PM, 12PM to 2PM, 2PM to 4PM, and 4PM to 6PM), allowing for the assessment and characterization of animal behavior during both the morning and afternoon periods spent in the paddocks.
Statistical analysis
For the structural variables of the pasture, the assumption of data normality for each system was checked using the Shapiro-Wilk at a 5% significance level. Averages were compared using the Student’s t-test at a 5% significance level, employing the Pooled or Satterthwaite method considering equal or different variances, respectively, through the TTEST procedure in SAS software (SAS Institute Inc. 2018).
For the variables of consumption and DesMS, only the effect of different systems was tested as a fixed effect, considering animals as repetitions and the paddock as a random effect (block effect). Data significance was analyzed using the F-test through the MIXED procedure in SAS software (SAS Institute Inc. 2015) with the following statistical model:
In which:
\({Y}_{ijk}\): value of the i-th experimental unit (animal) subjected to the j-th system in the k-th block (paddock); \(\mu\): fixed effect of the overall mean; \({S}_{j}\): fixed effect level of the system; \({\alpha }_{k}\): 12pplie effect of the k-th block; \({\epsilon }_{ijk}\): random effect associated with the i-th experimental unit (animal) subjected to the j-th system in the k-th block (paddock), where \({\epsilon }_{ijk}\) assumes iid N(0, σA2).
For the behavioral variables, the effect of different systems (silvopastoral and monoculture), evaluation period, and system × period interaction were analyzed for significance using the F-test through the MIXED procedure of the SAS software (SAS Institute Inc. 2015). The evaluation period factor was considered a repeated measure over time. In the selection of the covariance matrix structure for repeated measures, the corrected Akaike information criterion and the Bayesian information criterion were used. The following statistical model was used for these variables:
In which:
\({Y}_{ijk}\): value of the i-th experimental unit (paddock) subjected to the j-th system in the k-th evaluation period; \(\mu\): fixed effect of the overall mean; \({S}_{j}\): fixed effect of the system level; \({\alpha }_{ij}\): random effect of the i-th repetition in the j-th system, 12ppli α_ij assumes iid N(0, σA2); \({P}_{k}\): fixed effect of the evaluation period; \({\left(SP\right)}_{jk}\): fixed effect of the interaction between system and evaluation period; \({\epsilon }_{ijk}\): random effect associated with the i-th repetition in the j-th system and k-th evaluation period, where \({\epsilon }_{ijk}\) assumes ~ N(0, σB2), considering that σB2 is the variance and covariance matrix, as it assumes error dependence.
The residuals from the models above were tested for fit to the normal distribution using the Shapiro-Wilk test applied at a significance level of 5% (p < 0.05). In the presence of a difference between fixed effect factors by the F-test at a significance level of 5% (p < 0.05), Tukey’s mean comparison tests were applied, also considered significant at 5% (p < 0.05).
Results
Pasture structural traits
No effects (p > 0.05) of monoculture and silvopastoral systems were observed on all components of capim-tamani biomass (Table 3). The same occurred for theLM/DM and LB/S.
Regarding the productive responses of the capim-tamani grass, they were not affected (p > 0.05) by the shading of the silvopastoral system (Table 4). Green Forage Accumulation (GFA) accounted for 78.4% of the average Total Forage Accumulation (TFA = 1,085 kg DM ha− 1). Neither system affected Forage Availability, with an approximate average value of 5.0 kg DM per 100 kg of BW− 1 day− 1, as well as Defoliation Intensity, Grazing Efficiency, and Forage Use Efficiency showed no effect, with averages close to 20%, 58%, and 47%, respectively. BCS, Dry Matter Intake, and Disappearance. Average BCS was 2,65 in ewes, not differing between monoculture and silvopastoral systems (Table 5). Neither system affected (p > 0.05) DM consumption expressed in g/day and %BW. A similar pattern was observed for other nutrient consumptions (OM, CP, NDFcp, NFC, and TDN), which did not differ between them (Table 5). There was no effect of the natural shading of trees on the percentages of dry matter disappearance and other nutrients from capim-tamani (Table 6).
Behavioral responses
There was interaction (Treatment x Period) between the time the animals spent in the shade in monoculture and silvopastoral systems. In the periods evaluated, the animals spent more time in the shade in the system with natural tree shading compared to monoculture, except for the period between 2 and 4 PM, where the times were equal. In the monoculture system with only artificial shading (i.e., shade cloth), the animals remained in the shade on average about 15.6% throughout the evaluated period, equivalent to just under two hours under the shade cloth of the twelve daily hours they were in the paddocks, with no influence from the evaluated two-hour periods. In the silvopastoral system, on the other hand, animals remained approximately 40% of the entire evaluated period, which corresponded to approximately five and a half hours under natural shading; with a longer time spent between 4 and 6 pm with 72.8% during this period. In the other hours, there were similar percentages, with an average time close to 33% (40 min) in each two-hour interval. There was no effect of the systems on behavioral activities (grazing, ruminating, standing and lying idle, and other activities). However, there was an effect (p < 0.05) of the periods, except for lying idle, which showed similarity throughout the entire period of animals’ stay in the paddocks (Table 7). On average, the animals grazed for approximately 70% (8.4 h) of the time spent in the paddocks. The highest grazing frequencies occurred throughout the morning period (between 6 and 8 am and 10–12 pm), with 4.8 h of grazing by the animals. This behavioral pattern with a high frequency of grazing persisted until 12 − 2 pm (80.6%) of the afternoon period. Overall, for other activities such as ruminating, standing idle, lying idle, and other activities, there was a significant increase in the last period of stay in the paddocks (4–6 pm), where the most common activities were standing idle (35%) and ruminating (30.5%).
The average bite rate was 45 bites per minute; however, there was an effect of the period on this behavior. The highest rates (48.6 bites per minute) were observed between 12 − 2 pm, while the lowest occurred between 4 and 6 pm with an average of 35.9 bites per minute.
In terms of absolute frequency presented for activities recorded punctually during the periods of animal stay in the paddocks (Fig. 2), a similar pattern between the systems was observed for both defecation and urination frequencies, with a total of 17 and 18 occurrences, respectively, and an average frequency of about three times in each two-hour period.

Absolute frequency of specific activities of Morada Nova sheep managed in capim-tamani grass pasture under irrigated monoculture and silvopastoral systems in the Brazilian semiarid
There were a total of eight records of animals drinking water in both systems. For the activity of mineral supplement intake, the silvopastoral system showed a slightly higher frequency (ten times) compared to the monoculture system, which was observed only six times.
Discussion
In silvopastoral systems, microclimatic changes can affect productivity, pasture persistence, and animal behavior. However, with appropriate spatial arrangements of the tree component, maintaining balance among pasture, trees, and animals, in similar productive responses to traditional systems with no trees can be achieved (Paciullo et al. 2017).
The level of shading used (22.65%) did not lead to an increase or reduction in the structure of the capim-tamani pasture compared to the monoculture system. This suggests that the arrangement and density of trees used within the pasture provided conditions for moderate shading, suitable for the development of the forage and its resistance to imposed shading.
However, it is important to emphasize the attention that should be given during the establishment of woody cover levels in integrated systems, highlighting the main characteristics and attributes for selecting tree species, such as tree density, canopy architecture, and trunk height (Caron et al. 2012; Oliveira et al. 2018). Shading can limit the quantity and quality of photosynthetically active radiation in the understory, triggering a series of changes in plants, such as a decrease in tiller population (Gastal and Lemaire 2015), reducing biomass production (Lima et al. 2019), which was not observed in the present study.
Linked to the tree arrangement, maintaining the productivity of a silvopastoral system also relies on the correct selection of the forage species. The chosen species should not only be shade-tolerant but also exhibit suitable productivity, adaptability to the management employed, and compatibility with the soil and climatic conditions of the region (Andrade et al. 2003). In the Semi-Arid region, capim-tamani demonstrated favorable structural and production characteristics in the silvopastoral system, highlighting phenotypic plasticity in response to the continuous grazing method recommended for sheep. The stability in tiller population (i.e., PDT) observed during the experimental period for both evaluated systems reflects effective and efficient intensive pasture management (nitrogen fertilization and irrigation). This is particularly important as tiller population is considered a crucial variable regarding pasture longevity and formation (Araújo et al. 2020).
Nevertheless, the PDT quantified in this study was 27.35% lower when compared to the same fertilization conditions studied by Vasconcelos et al. (2020), who assessed the morphogenic and structural traits of capim-tamani under crescent doses of nitrogen. This reduction is attributed to the different planting methods. The authors mentioned above carried out manual planting in rows with a spacing of 0.25 m between rows, compared to the broadcast sowing technique. Additionally, the influence of grazing and trampling by animals on the grass could contribute to these differences. The LB/S ratio, which was not affected by the shading, is an important indicator of forage nutritive value, directly linked to protein content (Mochel Filho et al. 2016) and digestibility, consequently influencing the improvement of animal ingestive behavior (Euclides et al. 1999). A critical limit for this ratio is considered to be 1,0 (Pinto et al. 1994), and values below this threshold would imply a decrease in both the quantity and quality of forage produced. The values found in this study, 3,31 for monoculture and 2,97 for silvopastoral, are well above this recommended threshold. From the perspective of animal nutrition, the production of stems is not desirable (Freitas et al. 2012), having a direct effect on grazing time, bite rate, and digestibility. During the periods of higher grazing frequency observed in the animals, the average bite rate was close to 48 bites per minute, which is quite similar to that reported by Forbes and Hodgson (1985) in a study on the behavior of sheep in densely sown perennial ryegrass pasture, where they observed a bite rate of 48 bites per minute. In comparison to the study by Meneses (2022), which evaluated irrigated capim-tamani in monoculture, fertilized with urea (450 kg ha− 1 N year-1) and castor cake, under a similar continuous grazing method with sheep, higher average values were observed for the common structural characteristics (PDT, TFB, GFB, GLBB and LB/S ratio). However, the recommended height in their study was 22 cm with a crop growth cycle of 24 days. Regardless, integrated systems can meet agroecosystem premises with economically sustainable goals (Motta-Delgado et al. 2019) when compared to more intensive traditional systems, but fertilization constitutes a significant portion of the production cost (Avelino Cabral et al. 2021). Both the system and, especially, the nitrogen fertilization dose show a positive correlation with forage yield (Martuscello et al. 2015). Once nutritional requirements are met, nitrogen determines the speed of growth and forage production (Braz et al. 2011). Additionally, it is worth noting the positive responsiveness of capim-tamani to increasing doses up to1.200 kg ha− 1 N per year (Martuscello et al. 2019). In the present study, this farming under natural shading displayed acclimatization and flexibility under continuous grazing management and the recommended height for sheep, particularly with the support of irrigation and fertilization at150 kg ha− 1 year-1.
Similar or different responses obtained in the pasture can be conditioned by shading. As it is dynamic, shading is related to the distribution of trees, trunk height, and varies according to the inclination of the earth’s axis relative to the sun, which can vary the amount of light over time (Geremia et al. 2018). Therefore, the dynamics of forage accumulation and supply followed the same trend as the components of capim-tamani biomass, not being influenced by the production systems.
Despite the lower number of studies on forage management in silvopastoral systems compared to monoculture systems (Oliveira et al. 2021a), especially using capim-tamani in continuous grazing, many cultivars of Megathyrsus maximus have already shown moderate levels of shade tolerance (Andrade et al. 2003; Santiago-Hernandez et al. 2016). This indicates that they have potential in these integrated systems. The results obtained in this study further reinforce this idea, as the productivity of capim-tamani did not differ from the monoculture system. Considering the estimated average FA of 5,0 kg of DM/100 kg of BW day− 1, in this condition the average DI was 20%, with GE close to 58%. Gonçalves et al. (2018) report that the continuous grazing method allows for greater forage selection by the animal. Under these conditions, the low average defoliation intensity (DI) we found prioritizes a higher level of selectivity, favoring greater animal performance, accompanied by adequate Forage Use Efficiency (EUF), which averaged 45% efficiency. However, understanding the relationships between pasture characteristics and the ingestive behavior of ruminants is crucial for devising pasture management strategies. The fact that the animals did not receive supplementation is due to logical nutritional reasons. Morada Nova breed females have a low adult weight and lower nutritional requirements during the maintenance phase; thus, feeding costs for the ewes are reduced (Facó et al. 2008). In this case, the animals depended solely on forage, which could lead to increased competition and pressure regarding selectivity and consumption (Jochims et al. 2010). Therefore, observing the nutritional status of the animals through Body Condition Score (BCS) became crucial to assess the pasture’s potential in both systems and to make informed decisions regarding future reproductive management. In general, a BCS of 3 in ewes is characteristic of a more balanced metabolic state, indicating an adequate nutritional balance (Caldeira et al. 2007). This classification aligns with the average value observed in our study. Considering the nutritional requirements in animal production systems, a minimum of 7% crude protein in the diet is recommended, necessary for maintaining the growth of bacteria aimed at microbial protein production, which is crucial for the nutritional efficiency of ruminants (NRC 2001). In the conditions of the present study, average values above the expected were observed (monoculture: 8.28% and silvopastoral: 8.27%; Table 2), highlighting the importance of capim-tamani grass for intensive production systems in the Semi-Arid region. Associated with the availability of forage and the observed nutritional value of capim-tamani grass, the consumption did not differ between the evaluated systems. However, variations in consumption can occur in similar environments, as shown in the study by Sousa et al. (2015), who investigated the effect of the silvopastoral environment on forage (dry matter) consumption by sheep compared to the monoculture environment of capim-marandu grass. The authors found that animals had higher consumption in the shaded environment compared to the monoculture system. This result was attributed to the better microclimatic conditions provided by this system, as the improved thermal comfort provided lower metabolic and thermogenic dietary restriction (Forbes 1995). For the percentages of DisDM and other nutrients, there was no difference between the evaluated systems, which may be due to the observed similarity in the structural, nutritional, and productive characteristics of the capim-tamani. According to Orskov and McDonald (1979), disappearance values of 99.9%, 64.6%, and 53.7% are considered high, medium, and low degradability, respectively. In this sense, our study found that capim-tamani grass had a medium disappearance rate, providing efficiency in nutrient utilization. However, tropical forages typically exhibit low degradability (Gerdes et al. 2000), mainly due to the high content of cell wall components such as hemicellulose, cellulose, and lignin, and the low content of potentially digestible compounds, such as non-fibrous carbohydrates, protein, ether extract, vitamins, and minerals (Reis et al. 2005). Therefore, this forage exhibited suitable characteristics that did not compromise ruminal degradation, which may have been positively influenced by the established height management goals, coupled with the effect of irrigation and nitrogen fertilization during the experimental period. However, it is emphasized that cultivars of Megathyrsus maximus are a viable alternative in irrigated intensive systems for use in semi-arid environments (Veras et al. 2020), as well as their productive potential and nutritional quality (Pereira et al. 2022), and promotion of adequate digestibility (Stabile et al. 2010) In the analysis of behavioral variables, the predominantly longer time spent in the shade by animals in the silvopastoral system throughout the entire period of their stay in the paddocks further reinforces the dissemination of these livestock systems in the current scenario. Because, in addition to being sustainable, they promote animal production based on well-being (Lima et al. 2019; Pinheiro and Nair 2018), especially in the semi-arid region, where the impact of climate on thermal stress is even more critical. The peak of animal permanence in the shade being observed between 16 and 18 h, reaching over 70% during this period, is naturally common, as it approaches sunset. Moreover, there is variation in the shadow formation process according to the sun’s position throughout the day, and the configuration of the shadow is crucial and influenced by the angle of incidence of solar radiation (Silva 2006). In other words, the area of the shadow tends to increase from noon, promoting a higher percentage of shaded area at the end of the day, especially where there were tree components in the pasture. Therefore, there are possibilities that behavioral responses may not have been directly conditioned by shading since, being a locally adapted breed like Morada Nova, it portrays adaptability to the semi-arid climate (McManus et al. 2014). This is different from the behavior of exotic breeds (specialized) or also called commercial breeds, which, when observed in these integrated systems, constantly seek shaded areas for thermolysis (Dada et al. 2021).
However, the natural shading of the tree component improves the microclimate and thermal comfort indices due to the reduction of direct solar radiation (Santos Neto et al. 2022). Thus, it improves feeding behavior conditions and, consequently, animal performance. The highest grazing frequencies by the animals occurred throughout the morning period (between 6 and 8 am and 10–12 am), extending until 12 − 2 pm. This behavior may reveal one of the ways the animals dealt with heat stress because, in addition to the milder temperatures recorded during this period (Image 1), it is also inferred that the pasture is moist due to nighttime irrigation. In the evening, the animals reduce grazing, seeking to dissipate heat in areas with high temperatures (Façanha et al. 2020; Ferreira et al. 2020). This behavior may also be associated with the management of the animals, which spent 12 h (from 6 am to 6 pm) in the paddocks grazing and the entire night in enclosures without feeding. As they entered the paddocks in the morning, regardless of the system (monoculture or silvopastoral), the first thing they did was intense grazing to satisfy their hunger. This situation aligns with what was observed by Gregorini (2012), where animals tend to wait for the pasture to dry before initiating their behavioral habits, but this occurs under specific conditions where animals either remain in the pasture for 24 h or receive supplementation after the period they leave the pasture. As the grazing frequency decreased from 2 pm to 4 pm until 4 pm to 6 pm, there was a parallel increase in behavioral activities such as rumination (2 pm to 4 pm: 19.3%, 4 pm to 6 pm: 30.5%), standing idle (2 pm to 4 pm: 16.8%, 4 pm to 6 pm: 35%), and other activities (2 pm to 4 pm: 1.6%, 4 pm to 6 pm: 4.7%). Possibly, during these hours, the animals were already satiated, and as the animals become satiated, a significant portion of their time is spent in other activities (Hower et al. 1998). This is also evident and explained by the average reduction in the bite rate, which dropped to only 35.9% during the period between 4 pm and 6 pm. The inverse relationship between grazing frequency and activities such as rumination, idleness (lying down and standing), and other activities, particularly in the late hours of the animals’ stay in the paddocks (i.e., 4 pm to 6 pm), can be explained in this study by the animals’ habitual behavior of anticipating their return to the pens during the nighttime, positioning themselves near the gates. Consequently, the grazing frequency decreased, and activities involving interaction, rumination, and idleness increased. Sheep exposed to higher direct sunlight radiation tend to increase water intake compared to those kept in thermally comfortable environments (De et al. 2020). Sousa et al. (2015) observed a total water consumption percentage 11% higher in sheep that were in full sun monoculture systems compared to those in silvopastoral systems. According to Baumer (1991), animals protected from heat can reduce water intake by up to 20%. However, in our study, in absolute terms (Fig. 2), a similar frequency pattern was observed throughout the day for both systems. This observed similarity may be directly associated with the innate behavior of locally adapted breeds, even reflecting in urinary frequency, which was also similar between them.
The defecation frequency was similar between systems, occurring three times every two hours throughout the period when the sheep were in the paddocks. This could be due to the exclusivity of their diet, which consisted only of capim-tamani pasture, and also because of similar consumption patterns. Fecal excretion in an animal is inversely proportional to digestibility but directly related to the amount of feed ingested (Cardoso et al. 2014).
Conclusion
The intensively managed capim-tamani pasture under continuous grazing in sheep in the silvopastoral system showed similar forage production and consumption compared to the monoculture system. In addition, the silvopastoral system reduces solar radiation, directly influencing the ingestive behavior of animals throughout the day. Finally, to demonstrate the feasibility of using these integrated systems with the native Caatinga tree component, there is a need to continue research in other fields and thus provide further guidance. However, the present study serves as a dynamic and appropriate starting point, with promising results for these complex integrated systems and sustainable, managed intensively for the boosting the sheep industry in semi-arid regions worldwide.
Data availability
Data will be made available on request.
References
Alvares CA, Stape JL, Sentelhas PC, De Moraes Gonçalves JL, Sparovek G (2013) Köppen’s climate classification map for Brazil. Meteorol. Zeitschrift 22:711–728. https://doi.org/10.1127/0941-2948/2013/0507
Alvarez VVH, Novaes RF, Barros NF, Cantarutti RB, Lopes AS (1999) Interpretação dos resultados das análises de solos. In: Ribeiro AC, Guimaraes PTG (eds) Recomendação para o uso de corretivos e fertilizantes em Minas Gerais: 5ª Aproximação. Comissão de Fertilidade do Solo do Estado de Minas Gerais, Viçosa, pp 25–32
Alves JJA, Araujo MA de, Nascimento SS do (2009) DegradaUção da Caatinga: Uma Investigação Ecogeográfica. Revista Caatinga, Mossoró 22(3):126–135
Andrade CD, Garcia R, Couto L, Pereira OG, Souza AD (2003) Desempenho de seis gramíneas solteiras ou consorciadas com o Stylosanthes guianensis cv. Mineirão e eucalipto em sistema silvipastoril. Rev Bras Zootec 32(6):1845–1850
AOAC (2012). Ofcial Method of Analysis. 19 th ed. Association of Ofcial Analytical Chemists; (Gaithersburg, MD)
Araújo RA, Rodrigues RC, Costa CS, Santos FNS, Lima AJT, Rodrigues MM (2020) Dynamics and stability of marandu grass tillers in monocrop systems and babassu palm silvopastoral systems. Acta Sci - Agron 42. https://doi.org/10.4025/actasciagron.v42i1.42445
Avelino Cabral CE, Cabral CHA, Santos ARM, Motta AM, Mota LG (2021) Impactos Técnico-Econômicos Da Adubação De Pastos. Nativa 9 173–181. https://doi.org/10.31413/nativa.v9i2.12047
Barthram GT (1981) Sward structure and the depth of the grazed horizon. Grass Forage Sci 36:130–131
Baumer M (1991) Animal production, agroforestry and similar techniques. Agrofor Abstracts 4:179–198
Braz TGS, Fonseca DM, Freitas FP, Martuscello JA, Santos ME, Santos MV (2011) Morphogenesis of Tanzania Guinea grass under nitrogen doses and plant densities. Revista Brasileira De Zootecnia 1420–1427 Viçosa, v. 40, n. 7. https://doi.org/10.1590/S1516-35982011000700004
Caldeira RM, Belo AT, Santos CC, Vazques MI, Portugal AV (2007) The effect of body condition score on blood metabolites and hormonal profiles in ewes. Small Rumin Res 68:233–241. https://doi.org/10.1016/j.smallrumres.2005.08.027
Cardoso ES, Santana Junior HA, Santana EOC, Ferreira AHC, Maciel MS, Oliveira ZF, Figueiredo CB, Brito JM (2014) Avanços no estudo de consumo em bovinos a pasto – revisão. Rev Eletr Nutr 11(5):3648–3659
Caron BO, Lamego FP, Souza VQ, Costa EC, Eloy E, Behling A, Trevisan R (2012) Interceptação Da radiação luminosa pelo dossel de espécies florestais e sua relação com o manejo das plantas daninhas. Ciência Rural 42(1):75–82. https://doi.org/10.1590/S0103-84782012000100013
Combellas J, Hodgson J (1979) Herbage intake and milk production by grazing dairy cows 1. The effects of variation in herbage mass and daily herbage allowance in a short-term trial. Grass Forage Sci 34:209–214. https://doi.org/10.1111/j.1365-2494.1979.tb01469.x
Dada JMV, Santos MLP dos, Muniz PC, Nunes-Zotti MLA, de Barros FRO, Vieira FM (2021) Postpartum behavioural response of Santa inês x dorper ewes and lambs in a silvopastoral system. Small Rumin. Res. 203. https://doi.org/10.1016/j.smallrumres.2021.106495
De K, Kumar D, Sharma S, Kumawat P, Mohapatra A, Sahoo (2020) Effect of drinking earthen pot water on physiological response and behavior of sheep under heat stress. J Therm Biol 87:102476. https://doi.org/10.1016/j.jtherbio.2019.102476
de Oliveira FA, Turco SHN, de Araújo GGL, Clemente CAA, Voltolini TV, Garrido MS (2013) Comportamento De Ovinos Da raça Santa Inês em ambientes com e sem disponibilidade de sombra. Rev Bras Eng Agrícola E Ambient 17:346–351. https://doi.org/10.1590/s1415-43662013000300015
de Pereira M, Almeida RG, Macedo MCM, dos Santos VAC, Gamarra ÉL, Castro-Montoya J, Lempp B, Morais MDG (2021) Anatomical and nutritional characteristics of Megathyrsus maximus genotypes under a silvopastoral system. Trop Grasslands-Forrajes Trop 9:159–170. https://doi.org/10.17138/TGFT(9)159-170
Detmann E, Valadares Filho SC (2010) On the estimation of non-fibrous carbohydrates in feeds and diets. Arq Bras Med Veterinária E Zootec 62:980–984. https://doi.org/10.1590/s0102-09352010000400030
Detmann E, Souza MA, Valadares Filho SC, Queiroz AC, Berchielli TT, Saliba EOS, Cabral LS, Pina DS, Ladeira MM, Azevedo JAG (2012) Métodos para análise de alimentos - INCT - Ciência Animal. 1st edn. Suprema, Visconde do Rio Branco
dos Santos MLP, Dada JMV, Muniz PC, Nunes-Zotti MLA, de Barros FRO, Vieira FMC (2021) Physiological responses of Santa Inês x Dorper ewes and lambs to thermal environment of silvopasture and open pasture systems. 205:0–5. https://doi.org/10.1016/j.smallrumres.2021.106565
Empresa Brasileira de (2015) Pesquisa Agropecuária – EMBRAPA - BRS Tamani, Forrageira híbrida De Panicum maximum. Embrapa Gado de Corte
Euclides VPB, Luiz Roberto LRL, Macedo MCM, De Oliveira MP (1999) Consumo Voluntário De Forragem De Três Cultivares De Panicum maximum sob Pastejo. Rev Bras Zootec 28:1177–1185. https://doi.org/10.1590/s1516-35981999000600002
Façanha DAE, Ferreira J, Silveira RMF, Nunes TL, de Oliveira MG, de Sousa JER, de Paula VV (2020) Are locally adapted goats able to recover homeothermy, acid-base and electrolyte equilibrium in a semi-arid region? J Therm Biol 90:102593. https://doi.org/10.1016/j.jtherbio.2020.102593
Facó O, Paiva SR, Alves LRN, Lôb RNB, Villela LCV (2008) Raça morada nova: origem, características E perspectivas. Sobral: Embrapa Caprinos. Embrapa Caprinos. Documentos, 75.
Ferreira J, Silveira RMF, de Sousa JER, Façanha DAE (2020) Locally adapted goats efficiently gain and lose heat in an equatorial semi-arid environment. Int J Biometeorol 64:1777–1782. https://doi.org/10.1007/s00484-020-01959-0
Forbes JM (1995) Voluntary food intake and diet selection in farm animals. CABI Publishing. Wallingford, UK
Forbes TDA, Hodgson J (1985) Comparative studies of the influence of sward conditions on the ingestive behaviour of cows and sheep. Grass Forage Sci 40:69–77. https://doi.org/10.1111/j.1365-2494.1985.tb01721.x
Freitas FP, da Fonseca DM, dos Santos Braz TG, Martuscello JA, Santos MER (2012) Forage yield and nutritive value of Tanzania grass under nitrogen supplies and plant densities. Rev Bras Zootec 41:864–872. https://doi.org/10.1590/S1516-35982012000400006
Gastal F, Lemaire G (2015) Defoliation, shoot plasticity, sward structure and herbage utilization in pasture: review of the underlying ecophysiological processes. Agriculture 5:1146–1171. https://doi.org/10.3390/agriculture5041146
Gerdes L, Werner JC, Colozza MT, Possenti RA, Schammass EA (2000) Avaliação De Características De Valor Nutritivo das Gramíneas Forrageiras Marandu, Setária E Tanzânia Nas Estações do Ano. Rev Bras Zootec 29:955–963. https://doi.org/10.1590/s1516-35982000000400003
Geremia EV, Crestani S, Mascheroni JDC, Carnevalli RA, Mourão GB, Silva SC (2018) Sward structure and herbage intake of Brachiaria brizantha Cv. Piatã in a crop-livestock-forestry integration area. Livest Sci 212:83–92. https://doi.org/10.1016/j.livsci.2018.03.020
Gonçalves RP, Bremm C, Moojen FG, Marchi D, Zubricki G, Caetano LAM, Neto AB, de Carvalho PC (2018) Grazing down process: the implications of sheep’s ingestive behavior for sward management. Livest Sci 214:202–208. https://doi.org/10.1016/j.livsci.2018.06.005
Gregorini P (2012) Diurnal grazing pattern: its physiological basis and strategic management. Anim Prod Sci 52:416–430. https://doi.org/10.1071/AN11250
Hodgson J (1990) Grazing management: Science in to pratice. [s.l]. Longman Scientific & Technical, Essex, p 203p
Howery LD, Provenza FD, Ruyle GB (1998) How do domestic herbivores select nutritious diets on rangelands? Behavioral Education for Human, Animal, Vegetation, and Ecosystem Management (BEHAVE). p 52
IBGE (2020) Censo Agropecuário. Rio de Janeiro: IBGE. Disponível em: https://biblioteca.ibge.gov.br/visualizacao/periodicos/3093/agro_2017_resultados_preliminares.pdf. Acesso em setembro de 2023
Jank L, Santos MF (2021) Capim - BRS Tamani (Panicum maximum Jacq). Comun Técnico 161:1–48
Jochims F, Pires CC, Griebler L, Bolzan AMS, Dias FD, Galvani DB (2010) Comportamento ingestivo e consumo de forragem por cordeiras em pastagem de milheto recebendo ou não suplemento. Rev Bras Zootec 39:572–581. https://doi.org/10.1590/s1516-35982010000300017
Klingman DL, Miles SR, Mott GO (1943) The cage method for determining consumption and yield of pasture herbage 1. Agron J 35:739–746. https://doi.org/10.2134/agronj1943.00021962003500090001x
Lima MA, Paciullo DSC, Morenz MJF, Gomide CAM, Rodrigues RAR, Chizzotti FHM (2019) Productivity and nutritive value of Brachiaria decumbens and performance of dairy heifers in a long-term silvopastoral system. Grass Forage Sci 74:160–170. https://doi.org/10.1111/gfs.12395
Martuscello JA, da Silva LP, da Cunha D, de NFV, Batista AC do, Braz S, Ferreira TG dos (2015) PS. Adubação nitrogenada em capim-massai: Morfogênese e produção. Cienc Anim Bras 16:1–13. https://doi.org/10.1590/1089-68916i118730
Martuscello JA, Rios JF, Ferreira MR, Assis JA, Braz TG, Cunha DNFV (2019) Produção e morfogênese de capim BRS Tamani sob diferentes doses de nitrogênio e intensidades de desfolhação. Bol Indústria Anim 76:1–10. https://doi.org/10.17523/bia.2019.v76.e1441
McManus C, Hermuche P, Paiva S, Ferrugem Moraes J, de Mendes MC (2014) Geographical distribution of sheep breeds in Brazil and their relationship with climatic and environmental factors as risk classification for conservation. Brazilian J Sci Technol 1:3. https://doi.org/10.1186/2196-288x-1-3
Meneses AJG (2022) Desempenho bioeconômico e controle parasitário em ovinos terminados intensivamente a pasto utilizando a torta de mamona como insumo alternativo. 133 f. Tese (Doutorado em Zootecnia) – Universidade Federal do Ceará, Fortaleza
Mochel Filho W, Carneiro MAAP, Andrade ACM, Magalhães J, Rodrigues B, Santos F, Costa N (2016) Produtividade E composição bromatológica de Panicum maximum cv. Mombaça sob irrigação e adubação azotada. Rev Ciências Agrárias 39:81–88. https://doi.org/10.19084/rca14154
Mott GO, Lucas HL (1952) The design conduct and interpretation of grazing trials on cultivated and improved pastures. In: Proceedings of the international grassland congress. pp 1380–1395
Motta-Delgado PA, Martínez HEO, Rojas-Vargas EP (2019) Indicators associated to pastures sustainability: a review. Cienc Tecnol Agropecu 20:409–430. https://doi.org/10.21930/rcta.vol20_num2_art1464
National Research Council (2007) Nutrient requirements of small ruminants: sheep, goats, cervids, and new world camelids, 1st edn. National Academy Press, Washington DC, p 384
National Research Council (2001) Nutrient requirements of small ruminants: sheep, goats, cervids, and new world camelids, 7th edn. National Academy Press, Washington DC, p 362
Nocek JE (1988) Situ and other methods to Estimate Ruminal protein and energy digestibility: a review. J Dairy Sci 71:2051–2069. https://doi.org/10.3168/jds.S0022-0302(88)79781-7
Nunes SF, Ferreira J, Silveira RMF, Sales DC, de Sousa JER, Paiva SR, Façanha DAE (2020) Morphometric characterization and zoometric indices of white Morada Nova breed: the first step for conservation. Small Rumin Res 192. https://doi.org/10.1016/j.smallrumres.2020.106178
Oliveira CC, Alves FV, Almeida RG, Gamarra ÉL, Villela SDJ, Almeida MPGM (2018) Thermal comfort indices assessed in integrated production systems in the Brazilian savannah. Agrof Syst 92:1659–1672. https://doi.org/10.1007/s104
Oliveira VAV, Braz TGS, Silva RB, Veloso ACR, Rufino LDA, Martuscello JA, Mourthé MHF, Barros LV (2021a) Defoliation heights for palisade grass cv. Marandu in silvopastoral system. Arq Bras Med Vet e Zootec 73:665–674. https://doi.org/10.1590/1678-4162-12253
Oliveira CC, Pereira M, Santos VAC, Romagnoli GT, Serra AP, Macedo MCM, Almeida RG (2021b) Establishment and production of forage grasses under different levels of shading in an integrated system. In: World congress on integrated crop-livestock-forestry systems. 2., Campo Grande. Proceedings… Brasília: Embrapa, pp 258–263
Orskov ER, Mcdonald I (1979) The estimation of protein degradability in the rumen from incubation measurements weighted according to rate of passage. J Agric Sci 92:499–503. https://doi.org/10.1017/S0021859600063048
Paciullo DSC, Gomide CAM, Castro CRT, Fernandes PB, Muller MD, Pires MDFA, Xavier DF (2011) Características produtivas e nutricionais do pasto em sistema agrossilvipastoril, conforme a distância das árvores. Pesquisa Agropecuária Brasileira 46(10):1176–1183
Paciullo DSC, Gomide CAM, Castro CRT, Mauricício RM, Fernandes PB, Morenz MJF (2017) Morphogenesis, biomass and nutritive value of Panicum maximum under different shade levels and fertilizer nitrogen rates. Grass Forage Sci 72(3):590–600. https://doi.org/10.1111/gfs.12264
Pereira M, Almeida RG, Bungenstab DJ, Morais MG, Lempp B, Macedo MC (2015) Panicum maximum genotypes yield under integrated 46 crop-livestock-forest system in the Brazilian Cerrado. In: World congress on integrated crop-livestock-forest systems. Brasília. Proceedings… Brasília: Embrapa, 1 p
Pereira LET, Nishida NT, Carvalho LR, Herling VR (2018) Recomendações para correção e adubação de pastagens tropicais. Faculdade de Zootecnia e Engenharia de Alimentos da Universidade de São Paulo, Pirassununga. https://doi.org/10.11606/9788566404227
Pereira M, de Difante G, dos G, Ítavo S, Rodrigues LCV, Gurgel JG, Dias ALC, Ítavo AM, de Veras CCBF (2022) Costa ABG Da, Monteiro GO De A. Production potential and quality of Panicum maximum cultivars established in a semi-arid environment. Trop Anim Sci J 45:308–318. https://doi.org/10.5398/tasj.2022.45.3.308
Pinheiro FM, Nair PKR (2018) Silvopasture in the Caatinga biome of Brazil: a review of its ecology, management, and development opportunities. For Syst 27:1–16
Pinto JC, Gomide JA, Maestri M (1994) Produção De matéria seca e relação folha:caule de gramíneas forrageiras tropicais, cultivadas em vasos, com duas doses de nitrogênio. Rev Bras Zootec 23(3):313–326
Prigge EC, Varga GA, Vicini JL, Reid RL (1981) Comparison of ytterbium chloride and chromium sesquioxide as fecal indicators. J Anim Sci 53:1629–1633. https://doi.org/10.2527/jas1982.5361629x
Reis RA, Melo GMP, Bertipaglia LMA, Oliveira AP (2005) Otimização Da utilização Da forragem disponível através da suplementação estratégica. In:. Volumosos na produção de ruminantes. Funep, Jaboticabal, pp 187–238
Russel A (1984) Body condition scoring of sheep. Farm Pract. https://doi.org/10.1136/inpract.6.3.91
Sanches AC, Gomes EP, Rickli ME, Fasolin JP, Soares MRC, Goes RHTB (2015) Produtividade e valor nutritivo do capim Tifton 85 irrigado e sobressemeado com aveia. Revista brasileira de engenharia agrícola e ambiental 19(2):126–133. https://doi.org/10.1590/1807-1929/agriambi.v19n2p126-133
Santiago-Hernández F, López-Ortiz S, Ávila-Reséndiz C, Jarillo-Rodríguez J, de Pérez-Hernández P (2016) Dios Guerrero-Rodríguez J. Physiological and production responses of four grasses from the genera Urochloa and Megathyrsus to shade from Melia azedarach L. Agrofor Syst 90:339–349. https://doi.org/10.1007/s10457-015-9858-y
Santos D, de C, Guimarães Júnior R, Vilela L, Pulrolnik K, Bufon VB, de França AF (2016) Forage dry mass accumulation and structural characteristics of Piatã grass in silvopastoral systems in the Brazilian savannah. Agric Ecosyst Environ 233:16–24. https://doi.org/10.1016/j.agee.2016.08.026
Santos VAC, Almeida RG, Laura VA, Alves FV (2019) Growth and forage yield of tropical grasses in silvopastoral systems in Brazil. Pesquisa Florestal Brasileira 39:294–294
Santos Neto CF dos, da Silva RG, Maranhão SR, Torres AFF, Barbosa Filho JAD, Macedo VHM, Cândido MJD (2022) Microclimate and animal thermal comfort indexes in different silvopastoral system arrangements in Caatinga. Int J Biometeorol 66:449–456. https://doi.org/10.1007/s00484-021-02182-1
SAS Institute Inc (2015) SAS/STAT® 14.1 User’s Guide. SAS Institute Inc, Cary, NC
SAS Institute Inc (2018) SAS/STAT® 15.1 User’s Guide. SAS Institute Inc, Cary, NC
Silva RG (2006) Tree shade evaluation and prediction in cattle feedlots. Eng Agric 26:268–281. https://doi.org/10.1590/s0100-69162006000100029
Sousa LF, Maurício RM, Paciullo DSC, Silveira SR, Ribeiro RS, Calsavara LH, Moreira GR (2015) Forage intake, feeding behavior and bioclimatological indices of pasture grass, under the influence of trees, in a silvopastoral system. Trop Grasslands 3:129–141.
Stabile S, Salazar dos Santos DR, Jank L, Rennó FP, Silva LFP (2010) Características De Producao E qualidade nutricional de genótipos de capimcoloniao colhidos em três estádios de maturidade. Rev Bras Zootec 39:1418–1428. https://doi.org/10.1590/S1516-35982010000700004
Vasconcelos ECG, Cândido MJD, Pompeu RCFF, Cavalcante ACR, Lopes MN (2020) Morphogenesis and biomass production of BRS Tamani guinea grass under increasing nitrogen doses. Pesqui Agropecu Bras 55. https://doi.org/10.1590/S1678-3921.PAB2020.V55.01235
Veras EL, de Santos Difante L, Gurgel G, da Costa ALC, Rodrigues ABG, Costa JG, Neto CM, de Gusmão Pereira JVE, Costa M (2020) Tillering and structural characteristics of panicum cultivars in the Brazilian semiarid region. Sustain 12. https://doi.org/10.3390/su12093849
Weiss WP, Conrad HR (1992) St. Pierre NR. A theoretically-based model for predicting total digestible nutrient values of forages and concentrates. Anim Feed Sci Technol 39:95–110. https://doi.org/10.1016/0377-8401(92)90034-4
Acknowledgements
To the Ceará Foundation for Scientific and Technological Development (FUNCAP) for funding the project and granting a scholarship to the first author. The authors express their gratitude to the State University of Vale do Acaraú for providing the necessary infrastructure for the completion of this research.
Author information
Authors and Affiliations
Contributions
Genilson Cesar Alves- Writing – review and editing; Statistical analysis. Robson Mateus Freitas Silveira, Vitor Hugo Maués Macedo, Elayne Cristina Gadelha Vasconcelos, Roberto Cláudio Fernandes Franco Pompeu, Hélio Henrique Araújo Costa, Ederson Gomes Eufrásio, Concepta McManus, Clemente Fernandes dos Santos Neto, Concepta McManus- review and editing. Aline Vieira Landim – Conceptualization, Supervision.
Corresponding author
Ethics declarations
Animal rights
This research was submitted to Animal Ethics Committee and was approved by process number 2017.01-014.
Consent to publish
The authors state that all human participants in the research provided consent for the publication of the data.
Financial support
The National Council for Scientific and Technological Development - CNPq.
Conflict of interest
Not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Alves, G.C., Neto, C.F.S., Silveira, R.M.F. et al. Do forage production and ingestive behavior of locally adapted sheep differ from an irrigated silvopastoral system to an irrigated monoculture in the semi-arid region?. Trop Anim Health Prod 56, 232 (2024). https://doi.org/10.1007/s11250-024-04073-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11250-024-04073-0