
Abstract
Grazing in small-scale dairy systems reduces costs and enhances sustainability. However, binary pastures (BP) have low persistency, which may be increased by including more species given their flexibility to withstand different agroecological and management situations. The objective was to assess a multi-species (MS) pasture of perennial ryegrass (RG), tall fescue (TF), bromegrass (BG), red clover (RC) and white clover (WC) in comparison to a BP of perennial ryegrass (RG) – WC grazed by six Holstein dairy cows during the dry season in an on-farm double cross-over experiment, with three 14-day each experimental periods for animal variables, and a split-plot design for pasture variables at 3 cow/ha. There were differences (P < 0.05) between pastures for sward height (MS 5.0 vs. BP 4.0 ± 0.10 cm, both MS and BP) and net herbage accumulation (MS 902 vs BP 228 ± 172.4 kg DM/ha, both MS and BP) and differences (P < 0.05) for chemical composition among periods. There were differences (P < 0.01) among periods for estimated DM intake, but no differences (P > 0.05) for milk yield (mean 16.8 kg/cow/day), milk fat or protein content (mean 31.8 and 28.8 g/kg). Multi-species pastures are a viable alternative for SSDS in the highlands of central Mexico, being more resistant to harsh environment and to weed and kikuyu grass invasion.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In Mexico, over 85% of farms with cattle have small herds of up to 30 cows (INEGI 2018), with around 150,000 dairy farms of which over 78% are small-scale with a herd size between 3 and 35 milking cows plus replacements (Prospero-Bernal et al. 2017).
Small-scale dairy systems (SSDS) are an option to ameliorate poverty and contribute to rural development (Espinoza-Ortega et al. 2007; Mc Dermott et al. 2010); but high feeding costs limit their sustainability in the highlands of central Mexico (Fadul-Pacheco et al. 2013; Prospero-Bernal et al. 2017). The inclusion of grazing temperate irrigated pastures and complementary maize silage for the dry season reduce feeding costs, increase profitability and enhance sustainability, as farms rely more on their home-grown pasture and forage production and less in purchased inputs (Prospero-Bernal et al. 2017).
Cut and carry pastures for SSDS were introduced in the highlands of central Mexico since the early 1970s in areas with available irrigation during the dry season. Grace et al. (2018) mentioned that grazing is the most economical way of feeding ruminants, and Pincay-Figueroa et al. (2016) and Prospero-Bernal et al. (2017) showed that in SSDS in the highlands of Mexico, although there were no differences in milk yields and milk composition, grazing reduced feeding costs between 20 and 25%.
Grass and legume pastures are an important feature of animal systems in temperate areas (Novy et al. 1995; Castro-Rivera et al. 2012; Burbano-Muñoz et al. 2018). Binary pastures of perennial ryegrass (Lolium perenne) and white clover (Trifolium repens) are common in ruminant production systems in Western Europe, Australia, New Zealand and other temperate regions of the world given their high yields, resistance to grazing and their high nutritional value (Bryant et al. 2016; Dodd et al. 2019).
More recently, MS pastures (mixtures of more than one plant functional group) have been evaluated against conventional RG/WC pastures. Multi-species pastures may offer improved dry matter (DM) yields and the reduction in the excretion of nitrogen (Scott and Pennell 2006; Dodd et al. 2019).
Grace et al. (2018) mentioned that besides reducing production costs from a higher DM yield, MS pastures may reduce seasonal fluctuations due to the complementarity of different growth patterns of the species included enhancing pasture persistence and a more uniform supply of herbage even in difficult conditions.
Multi-species pastures besides enhancing a beneficial botanical diversity may be more persistent and with higher and stable yields due to a larger number of plants per unit surface, which may be more resistant to weed and pest invasions, with a particular advantage in dry years (Sanderson et al. 2005).
Grazing of conventional binary RG/WC pastures in small-scale dairy farms in the highlands of central Mexico has proven successful in reducing feeding costs while keeping similar or slightly higher yields than under cut and carry management (Pincay-Figueroa et al. 2016; Prospero-Bernal et al. 2017). However, the sub-humid temperate climate with a long very dry season, limited irrigation and high stocking rates mean that sown temperate species do not persist beyond 2 or 3 years and pastures are invaded by the subtropical kikuyu grass (Cenchrus clandestinus) (Plata-Reyes et al. 2018).
Multi-species pastures may show an advantage under these agroecological (Sanderson et al. 2005) and management conditions. The objective was then to assess the productive performance of grazing dairy cows, pastures and herbage chemical composition of a multi-species pasture (three temperate grasses and two legumes) in comparison with a conventional binary RG/WC pasture in a small-scale dairy system on-farm experiment.
Materials and methods
Study area
An on-farm experiment following guidelines for participatory livestock technology development (Conroy 2005) was undertaken in the highlands of central Mexico, in the municipality of Aculco, located between 20 ° 06′ and 20 ° 17’ North and between 99 ° 40′ and 100 ° 00’ West. Mean altitude is 2440 m, and the climate is sub-humid temperate, a mean temperature of 13.2 °C and frosts from October to February, and annual rainfall is above 700 mm with a distinct rainy season (May to October) and dry season (November to April) (Celis-Alvarez et al. 2016).
The collaborating farmer owns 10 ha and a herd of nine Holstein cows plus replacements. The family runs the farm, as is usual in SSDS (Prospero-Bernal et al. 2017). Eight hectares are sown to maize for silage, and 2 ha have access to limited irrigation which are used for pastures.
Animals
Six multiparous cows were selected, with a mean milk yield prior to the experiment of 18 ± 3 (kg/cow/day), 71 ± 43 days in milk, 459 ± 41 kg mean live weight and mean body condition score of 2.4 ± 0.2. Cows were randomly assigned to treatment sequence with three cows per sequence.
The experiment took place between 2nd of March and 12th of April 2019, in three 14-day each experimental periods (EP) (Pérez-Ramírez et al. 2012), with 11 days for adaptation to diets and 3 days for measurements and recordings. Short-term EP are acceptable in grazing studies (Miguel et al. 2014).
Treatments were grazing MS pasture or BP. In accordance with the collaborating farmer’s tradition and decision, cows also received 4.6 kg DM/cow/day of 19% CP commercial concentrate divided in two allocations provided at each of two milkings (6:00 and 18:00 h).
It is a common practice in these small-scale dairy systems to supplement large amounts of concentrate (Fadul-Pacheco et al. 2013) in the belief they are essential to sustain milk yields. The collaborating farmer’s decision to supplement this amount of concentrate was respected as a premise of participatory research. The amount of concentrate is the same for all cows regardless of body weight or milk yield.
Cows continuously grazed pastures for 8 h/day, from 8:00 to 16:00 h, and kept indoors in an open pen overnight. Drinking water was freely available both in pastures and in the pen.
Pasture establishment
The experiment took place in two 1 ha pastures at a stocking rate of 3 cows/ha. The MS pasture was sown on 4 June 2018 to 15 kg/ha of perennial ryegrass (Lolium perenne cv. Tetragrain), 15 kg/ha of tall fescue (Lolium arundinaceus cv. Cajun II), 10 kg/ha of bromegrass (Bromus catharticus cv. Matua), 10 kg/ha of red clover (Trifolium pratense cv. Kenland) and 3 kg/ha of white clover (Trifolium repens cv. Fiona). The BP was sown on 30 August 2018 to 40 kg/ha of perennial ryegrass (Lolium perenne cv. Mathilde) and 3 kg/ha of white clover (Trifolium repens cv. Fiona).
Fertilization at sowing was 80 N – 80 P2O5–60 K. A second fertilization on the binary pasture only was on 30 March 2019 with 40 kg N/ha. Pastures had limited flood irrigation every month.
Pasture variables
Each pasture was nominally divided in two sub-pastures, as pasture replicates. Rising plate grass metre sward height (SH) was recorded at the beginning of the experiment and at the end of each EP taking 15 measurements per pasture division. Net herbage accumulation (NHA) was estimated following Burbano-Muñoz et al. (2018), from three 0.5 × 0.5 × 0.6 m exclusion cages per pasture division (a total of six exclusion cages per pasture treatment), cutting to ground level with shears inside a 0.4 × 0.4 quadrat outside the cage in day 0 and inside the exclusion cage on day 14 to estimate NHA by difference and expressed as kg DM/ha and kg DM/ha/day, placing the cages at random thereafter.
Hand-plucked herbage simulating grazing (Wayne 1964) was collected from each pasture at the end of each period and a composite sample for chemical analyses by triplicate using AOAC (1995) Official methods. Dry matter (DM), organic matter (OM), crude protein (CP) (method 990.03), neutral detergent fibre (NDF) (method 2002.04), acid detergent fibre (ADF) (method 973.18) and in vitro organic matter digestibility (IVOMD), from which metabolizable energy content was estimated (eME), following procedures described by Celis-Alvarez et al. (2016) and Burbano-Muñoz et al. (2018).
Botanical composition of herbage from both pastures was determined from five random 50 g samples per period cut to ground level; manually separated in grass and clover species, green and dead and other plants (forbs); and expressed as percentage in dry weight after drying in a draught oven for 48 h at 55 °C (Sanderson et al. 2005).
Animal variables
Cows were milked twice daily at 5:00 h and 17:00 h. Daily milk yields were recorded with a clock spring balance, and the mean daily yield for the last 3 days was used for statistical analyses (Plata-Reyes et al. 2018). Aliquot samples of milk were taken during the last 3 days to determine milk fat and protein content by the Gerber and Kjeldahl methods approved by Mexican standards for analyses of milk composition (COFOCALEC-Consejo para el Fomento de la Calidad de Leche y Sus Derivados A.C. 2004), and mean results were used for statistical analyses.
Milk urea nitrogen (MUN) was also determined by the enzymatic colorimetric method described by Chaney and Marback (1962), as has been reported from the previous work (Celis-Alvarez et al. 2016; Plata-Reyes et al. 2018).
Cows were weighed for two consecutive days at the end of each experimental period with an electronic portable weighbridge and mean live weight (kg/cow) used for analyses. Body condition score was recorded at the last weighing on a 1 to 5 scale (Wildman et al. 1982).
Herbage dry matter intake (DMI) was estimated by the utilized metabolizable energy method (Baker 1982, pp. 77--93), as in the previous work (Plata-Reyes et al. 2018), from metabolizable energy requirements minus that provided by the concentrate and the estimated eME content of herbage to estimate herbage and total DMI (kg DM/cow/day).
Statistical analyses
Pasture variables were analysed with a split-plot design where treatments (MS or BP pastures) were main plots as fixed effects and EP as split-plots (random effects) (Kaps and Lamberson 2004, pp. 294--312), with the following model:
where μ is the general mean; B is the effect of blocks (pasture divisions) (i = 1, 2, 3); T is the effect of pasture treatment (main plot) (j = 1, 2); E is the error term for main plots [r(T)ij]; p is the effect of experimental periods (split - plot) (k = 1, ..., 3); Tp is the interaction term between treatments and measurement periods; and e is the error term for split plots.
Animal variables were analysed as a double cross-over design with six cows during three EP following the treatment sequence MS – BP – MS and BP – MS – BP (Kaps and Lamberson 2004, pp. 294--312), analysed with a mixed model procedure of SAS Institute (2012) where treatment, period, sequence and the interaction treatment*period were fixed effects, whereas cow within treatment was considered the random effect in the following model:
where μ is the general mean, S is the effect due to treatment sequence (i= 1, 2), Cj (i) is the effect due to cows within treatment sequence (j = 1, 2, 3), Ph (i) is the effect due to experimental period within treatment sequence (h = 1, 2, 3), tk is the effect due to treatment (k = 1, 2), and eijkh is the residual variation.
Cross-over experimental designs are useful for on-farm experiments with small-holders, as they maximize limited experimental units (cows) (Kaps and Lamberson 2004, pp. 294--312) and have proven their value in the previous work with a limited number of cows (Miguel et al. 2014; Granados-Rivera et al. 2017; Plata-Reyes et al. 2018).
Results
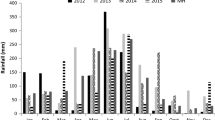
Figure 1 shows the meteorological conditions during the experiment, with basically no rain (only 20.5 mm), but with very high maximal temperatures above 25 °C which increased evapotranspiration. Mean minimal temperatures were 5.7 °C.

Rainfall and mean maximal and minimal temperatures during the experiment
Pasture variables
Table 1 shows results for SH and NHA. There was a highly significant (P < 0.01) difference with compressed SH being 1 cm higher in the MS (5 cm) than in the BP (4 cm). There were also highly significant differences among EP, declining in EP2 to recover slightly in EP3 for both pastures.
MS had a significantly higher (P = 0.03) NHA than BP (902 and 228, kg DM/ha, respectively). NHA for BP dropped dramatically in EP3, while in MS, there was a slight increase compared to EP2. There were no differences (P > 0.05) among EP for NHA.
Table 2 shows results for chemical composition of herbage and the concentrate. There were significant interactions (P < 0.05) for OM, CP and both fibre fractions, NDF and ADF.
There were no differences among pastures on CP content (P = 0.09). But, there were differences for EP in MS where a sharp decline in EP2 was observed to increase again in EP3, while CP content in BP remained constant in the first two periods to increase in EP3.
NDF and ADF showed larger variation in MS than in BP, with a slight increase in both NDF and ADF in MS from EP1 to EP2 to drop 90 g NDF/kg DM and over 50 g ADF/kg DM in EP3.
However, there were no differences (P > 0.05) between pastures or among EP for IVOMD with good quality herbage in both pastures, with a mean IVOMD of 797 and eME of 10.6 MJ/kg DM.
In Table 3 can be observed the botanical composition of pastures. Ryegrass was in low proportion in MS pastures (5.8%). It was interesting to note how the greater diversity of MS resulted in 67% of DM in green plants, with only 4.6% of invasion by spontaneous vegetation (forbs).
The BP in contrast showed that 37.4% of available live herbage was from spontaneous vegetation, and 5.9% was kikuyu grass (Cenchrus clandestinus) and invasive subtropical grass. Therefore, in BP, only 44.2% of green forage was from the preferred sown RG and WC.
Animal variables
Table 4 shows results for animal variables. There were no statistical differences (P > 0.05) in herbage dry matter intake (HDMI) between pasture treatments, but there was a highly significant difference (P < 0.01) among periods, with an increase in HDMI in EP2 and EP3 compared to EP1, with the same situation for total dry matter intake (TDMI).
There were no statistical differences (P > 0.05) in any of the other animal variables, between pastures or among periods. Milk yields were 17.2 and 16.3 (kg/cow/day) for MS and BP, respectively (P = 0.66), whereas fat and protein content were 31.6 and 32.1 (g/kg) for MS and BP, respectively (P = 0.93 and 0.37). Milk urea nitrogen for cows on MS was 13% higher (19.4 mg/dL) than cows on BP (16.8 mg/dL).
Discussion
Pasture variables
Adverse environmental conditions like drought and high temperatures (Fig. 1) affected pastures productive variables (height and NHA); however, BP was more sensible to these factors resulting in 75% lower NHA and lower sward height (1 cm) than MS pasture.
These sward heights were higher than that reported by Pincay-Figueroa et al. (2016), working also with binary pastures of RG and WC under continuous grazing by dairy cows in the dry season, recording a mean grass metre sward height of 3.1 cm and milk yields over 17 kg/cow/day, although in that experiment irrigation was regularly available.
In the work herein reported, besides low rain, limited irrigation and high maximal temperatures, there were mean minimal temperatures under 6 °C, the threshold below which temperate pastures stop growing. Those low temperatures might have hampered pasture growth, particularly white clover which is more sensible to low temperatures (Parsons and Chapman 2000).
The combination of environmental factors and grazing pressure (high stocking rate) affected the botanical composition of BP, where white clover and ryegrass represented 42%, whereas dead material, forbs and kikuyu grass represented 58%. As EP progressed, RG and forbs increased representing 42.7 and 43.1%, respectively, of the botanical composition at the end of the experiment.
On the contrary, MS pasture was less affected by the environmental and grazing pressure. White clover, BG and RG proportions increased from EP1 through EP3, whereas TF increased from EP 1 to EP2, to decrease in EP3. In this pasture, invasive species were kept at minimum agreeing with Dodd et al. (2019).
Despite the harsh environmental conditions, the nutritional characteristics of both pastures were not affected and were not different among them (Table 2). Nutritional quality indicators like CP, NDF, ADF and IVOMD are considered adequate and within the range of values reported in the literature (Miguel et al. 2014; Grace et al. 2018).
In contrast, the variety of species sown in MS created favourable conditions for clovers, such that white and red clover represented over 46% of available herbage DM, and although conditions were also unfavourable for RG in MS such that it represented only 5.8% of available herbage DM, companion TF and BG were able to grow more vigorously such that the three grasses comprised 29% of available green herbage DM, resulting in significantly higher NHA (Table 3).
Woodward et al. (2020) reported low persistency in RG pastures due to water deficit, as well as weed invasion, situation like that observed in the BP treatment, which is in line with the findings in our study.
Sanderson et al. (2005) reported higher production of multi-species pastures compared to a BP pasture and noted less presence of weeds. In the study herein reported, weeds represented 37% of available green herbage DM in BP while only 4.8% in the MS pasture (Table 3). When kikuyu grass is considered, invasive species represented almost a third of available green herbage DM in the BP pasture.
Kikuyu is a subtropical grass, originally from East Africa but naturalized in the highlands of central Mexico that invades RG/WC pastures leading to very low persistency of the sown pastures and that cannot survive the stress of the dry season, limited irrigation and high stocking rates in SSDS (Plata-Reyes et al. 2018). Even at an early stage after sowing, as in the BP pasture, kikuyu grass already comprised nearly 6% of available green herbage DM.
Cows prefer herbage that is easier to graze, those with less NDF like RG (Horadagoda et al. 2009), which may have had an influence on the low percentage of RG of the botanical composition in both pastures.
Animal variables
There are sward factors that affect grazing cows performance (height, herbage mass availability, tiller density, etc.) and animal factors that affect pasture productivity (stocking rate, age, milk yield, and live weight) (Mayne et al. 2000).
Net herbage accumulation in the MS pastures (64 kg DM/ha/day) provided ample availability for grazing cows, while it was a limiting factor for DMI in the BP with just 16 kg DM/ha/day, with a severe shortage during EP3. Despite the higher NAH in MS pasture, sward height still was under the ideal height (6 to 8 cm) acting as a limiting factor for cows to increase DMI and milk yields (Leaver 1985; Mayne et al. 2000; Parsons and Chapman 2000).
Despite the reduced availability of herbage in BP, milk yields in both treatments were not different and within the range (14–16 kg/cow/day) of those reported in other works on feeding strategies based on continuous grazing of BP in SSDS (Pincay-Figueroa et al. 2016; Plata-Reyes et al. 2018).
Concentrates represented 37 and 41% of TDMI (MS and BP, respectively). Grazing supplementation is intended to increase total dry matter or energy intakes particularly when there are restrictions on swards characteristics (herbage availability and grazing height). Under this situation (as in BP), supplements act as a buffer avoiding reductions on DMI, maintaining milk yields, avoiding body weight and body condition losses (Leaver 1985). If concentrates were not offered to cows, the low availability of herbage and high stoking rate would have further deteriorated grazing conditions putting at risk pasture persistency.
Roca-Fernández et al. (2016) in France evaluated pastures under rotational grazing comparing RG/WC to MS pastures (grasses – legumes – forb). Mean milk yield was 17.3 kg/cow/day; but with significantly higher yields in the pastures with four and five species (mean of 18.0 kg/cow/day) than RG/WC (mean of 16.6 kg/cow/day).
These results were due to the combined effects of better quality of herbage and a higher DM intake, similar to observations in the work herein reported for MS (a significant difference of 0.8 kg HDMI/cow/day more in MS than BP), reflected in numerical but not statistical higher milk yields in MS.
McCarthy et al. (2020) reported form a meta-analysis that grazing multi-species pastures compared to BP pastures increased milk yields on average by 1.2 kg/cow/day (in this study, the difference was 0.9 kg milk/cow/day higher in MS, although not significant).
Milk composition, with mean milk fat and protein content (31.8 ± 0.4 and 28.8 ± 1.1 g/kg), is within Mexican standards for raw milk. MUN values were higher than other reports from small-scale dairy systems in Mexico (Celis-Alvarez et al. 2016; Plata-Reyes et al. 2018) but within normal values mentioned by Powell et al. (2011) for cows in permanent confinement in the USA and by Ariyarathne et al. (2019) and Lopez-Villalobos et al. (2018) for grazing dairy cows.
Multi-species pastures are a viable option for SSDS in the highlands of central Mexico, with a higher proportion of sown species that make them more resistant to weed and kikuyu grass invasion. The plasticity and complementarity of different temperate grass species enabled better clover development and higher net herbage accumulation, of similar quality to herbage from binary perennial ryegrass-clover pastures and better grazing conditions that increased dry matter intake.
References
AOAC. 1995. Official Methods of Analysis. 15th ed., Association of Official Analytical Chemists, Arlington, VA.
Ariyarathne, H.B.P.C., Correa-Luna, M., Blair, H.T., Garrick, D.J., and Lopez-Villalobos, N., 2019. Estimation of heritabilities for milk urea and efficiency of crude protein utilization by day of lactation in a New Zealand dairy cow herd. New Zealand Journal of Animal Science and Production, 79, 183--187
Baker, R.D., 1982., Estimating herbage intake from animal performance In: Leaver JD, editor. Herbage Intake Handbook. Maidenhead (UK): (British Grassland Society), 77--93
Bryant, R.H., Edwards, G.R., and Robinson, B., 2016. Comparing response of ryegrass-white clover pasture to gibberellic acid and nitrogen fertiliser applied in late winter and spring. New Zealand Journal of Agricultural Research, 59, 18—31. DOI: https://doi.org/10.1080/00288233.2015.1119164
Burbano-Muñoz, V.A., López-González, F., Estrada-Flores, J.G., Sainz-Sánchez, P.A., and Arriaga-Jordán, C.M., 2018. Oat silage for grazing dairy cows in small scale dairy systems in the highlands of central Mexico. African Journal of Range and Forage Science, 35, 63--70. DOI: https://doi.org/10.2989/10220119.2018.1473493
Castro-Rivera, R., Hernández-Garay, A., Vaquera-Huerta, H., Hernández-Girón, J.P., Quello-Carrillo, A.R., Enríquez-Quiroz, J.F., and Martínez-Hernández, P.A., 2012. Productive performance of grass-legume associations under grazing conditions. Revista Fitotecnia Mexicana, 35, 87--95.
Celis-Alvarez, M.D., López-González, F., Martínez-García, C.G., Estrada-Flores, J.G., and Arriaga-Jordán, C.M., 2016. Oat and ryegrass silage for small-scale dairy system in the highlands of central Mexico. Tropical Animal Health and Production, 48, 1129--1134. DOI https://doi.org/10.1007/s11250-016-1063-0
Chaney, A.L., Marback, E.P., 1962. Modified reagents for determination of urea and ammonia. Clinical Chemistry, 8,130--132
COFOCALEC-Consejo para el Fomento de la Calidad de Leche y Sus Derivados A.C, 2004. NMX-700-COFOCALEC-2004. Sistema producto leche, alimento lácteo, leche cruda de vaca, especificaciones fisicoquímicas, sanitarias y métodos de prueba. México, D.F.
Conroy, C., 2005. Participatory livestock research. Bourton-on-Dunsmore (UK): ITDG Publishing.
Dodd, M., Dalley, D., Wims, C., Elliott, D., and Griffin, A. 2019. A comparison of temperate pasture species mixtures selected to increase dairy cow production and reduce urinary nitrogen excretion. New Zealand Journal of Agricultural Research, 62, 504--527. DOI:https://doi.org/10.1080/00288233.2018.1518246
Espinoza-Ortega, A., Espinosa-Ayala, E., Bastida-López, J., Castañeda-Martínez, T., and Arriaga-Jordán, C.M., 2007. Small-scale dairy farming in the highlands of central Mexico: Technical, economic and social aspects and their impact on poverty. Experimental Agriculture, 43, 241--256. DOI:https://doi.org/10.1017/S0014479706004613.
Fadul-Pacheco, L., Wattiaux, M.A., Espinoza-Ortega, A., Sánchez-Vera, E., and Arriaga-Jordán, C.M., 2013. Evaluation of sustainability of smallholder dairy production system in the highlands of Mexico during the rainy season. Agroecology and Sustainable Food Systems, 37, 882--901. DOI: https://doi.org/10.1080/21683565.2013.775990
Grace, C., Boland, T.M., Sheridan, H., Lott, S., Brennan, E., Fritch, R., and Bridget-Lynch, M., 2018. The effect of increasing pasture species on herbage production, chemical composition and utilization under intensive sheep grazing. Grass and Forage Science, 73, 852--864. DOI: https://doi.org/10.1111/gfs.12379.
Granados-Rivera, L.D., Hernández-Mendo, O., González-Muñoz, S.S., Burgueño-Ferreira, J.A., Mendoza-Martínez, G.D., and Arriaga-Jordán, C.M., 2017. Effect of palmitic acid on the mitigation of milk fat depression syndrome caused by trans-10, cis-12-conjugated linoleic acid in grazing dairy cows. Archives of Animal Nutrition, 71, 428--440. DOI: https://doi.org/10.1080/1745039X.2017.1379165
Horadagoda, A., Fulkerson, W.J., Nandra, K.S., and Barchia, I.M., 2009. Grazing preferences by dairy cows for 14 forage species. Animal Production Science, 49, 586--594. https://doi.org/10.1071/EA08299
INEGI – Instituto Nacional de Estadística y Geografía, 2018. Encuesta Nacional Agropecuaria (ENA) 2017: Conociendo el campo de México. Resultados. https://www.inegi.org.mx/contenidos/programas/ena/2017/doc/ena2017_pres.pdf. Accessed 14 Nov 2019
Kaps, M., and Lamberson, W.R., 2004. Biostatistics for Animal Science. Trowbridge: Cromwell Press. Change-over designs, 294—312.
Leaver, J.D., Effects of supplements on herbage intake and performance. In: Frame J, Editor. Grazing. Occasional Symposium No. 19, British Grassland Society, 1985: 79--88
Lopez-Villalobos, N., Correa-Luna, M., Burke, J.L., Sneddon, N.W., Schutz, M.M., Donaghy, D.J., and Kemp, P.D., 2018. Genetic parameters for milk urea concentration and milk traits in New Zealand grazing dairy cattle. New Zealand Journal of Animal Science and Production, 78, 56--61
Mayne, C.S., Wright, I. and Fisher G.E.J., 2000. Grassland management under grazing and animal response. In: Hopkins, (ed) Grass: its production and utilization, Chapter 10, (Oxford, UK, Blackwell Science), 247-291.
Mc Dermott, J.J., Stall, S.J., Freeman, H., and Van de Steeg, J.A., 2010. Sustaining intensification of smallholder livestock systems in the tropics. Livestock Science, 130, 95--109. DOI:https://doi.org/10.1016/j.livsci.2010.02.014
McCarthy, K.M., McAloon, C.G., Lynch, M.B., Pierce, K.M., and Mulligan, F.J., 2020. Herb species inclusion in grazing swards for dairy cows — A systematic review and meta-analysis. Journal of Dairy Science, 103, 1416--1430. https://doi.org/10.3168/jds.2019-17078
Miguel, M.F., Ribeiro-Filho, H.M.N., de Andrade, E.A., Moraes Genro, M.T., and Delagarde, R., 2014. Pasture intake and milk production of dairy cows grazing annual ryegrass with or without corn silage supplementation, Animal Production Science, 54, 1810-1816.
Novy, E.M., Casler, M.D., and Hill, M.R. Jr., 1995. Selection for persistence of tetraploid ryegrasses and festulolium in mixture with perennial legumes. Crop Science, 35, 1046--1051.
Parsons, A.J., and Chapman, G.F., 2000. The principles of pasture growth and utilization. In: Hopkins A., editor. Grass: Its production and utilization (3rd. Edition), Oxford (U.K): British Grassland Society and Blackwell Science, 31--89
Pérez-Ramírez, E., Peyorad, J.L., and Delagarde, R., 2012. N-Alkanes v. ytterbium/faecal index as two methods for estimating herbage intake of dairy cows fed on diets differing in the herbage: maize silage ratio and feeding level. Animal, 6, 232--244. DOI: https://doi.org/10.1017/S1751731111001480.
Pincay-Figueroa, P.F., López-González, F., Velarde-Guillén, J., Heredia-Nava, D., Martínez-Castañeda, F.E., Vicente, F., Martínez-Fernández, A., and Arriaga-Jordán, C.M., 2016. Cut-and-carry vs. grazing of cultivated pastures in small-scale dairy systems in the central highlands of Mexico. Journal of Agriculture and Environment for International Development, 110, 349--363. DOI: https://doi.org/10.12895/jaeid.20162.496
Plata-Reyes, D.A., Morales-Almaraz, E., Martínez-García, C.G., Flores-Calvete, G., López-González, F., Prospero-Bernal, F., Valdez-Ruiz, C.L., Zamora-Juárez, Y.G., and Arriaga-Jordán, C.M., 2018. Milk production and fatty acid profile of dairy cows grazing four grass species pastures during the rainy season in small-scale dairy systems in the highlands of Mexico. Tropical Animal Health and Production, 50, 1797--1805. https://doi.org/10.1007/s11250-018-1621-8
Powell, J.M., Wattiaux, M.A., and Broderick, G.A., 2011. Short communication: Evaluation of milk urea nitrogen as a management tool to reduce ammonia emissions from dairy far DM, Journal of Dairy Science, 94, 4690--4695. doi: https://doi.org/10.3168/jds.2011-4476
Prospero-Bernal, F., Martínez-García, C.G., Olea-Pérez, R., López-González, F., and Arriaga-Jordán, C.M., 2017. Intensive grazing and maize silage to enhance the sustainability of small-scale dairy system in the highlands of Mexico. Tropical Animal Health and Production, 49, 1537--1544. DOI https://doi.org/10.1007/s11250-017-1360-2
Roca-Fernández, A.I., Peyraud, J.L., Delaby, L., and Delagarde, R., 2016. Pasture intake and milk production of dairy cows rotationally grazing on multi-species swards. Animal, 10, 1448--1456. doi:https://doi.org/10.1017/S1751731116000331
Sanderson, M.A., Soder, K.J., Muller, L.D., Klement, K.D., Skinner, R.H., and Goslee, S.C., 2005. Forage mixture productivity and botanical composition in pastures grazed by dairy cattle. Agronomy Journal, 97, 1465--1471. doi:https://doi.org/10.2134/agronj2005.0032
SAS Institute. 2012. SAS/STAT 9.3. User’s Guide. Version 9.4 ed. Cary (NC): SAS Institute Inc.
Scott, D., and Pennell, C.G.L., 2006. Agronomic methods: Evaluation by multiple-species pasture mixtures. New Zealand Journal of Agricultural Research, 49, 191--200. https://doi.org/10.1080/00288233.2006.9513709
Wayne, C.C., 1964. Symposium on nutrition of forages and pastures: Collecting samples representative of ingested material of grazing animals for nutritional studies. Journal Animal Science, 23, 265--270
Wildman, E.E., Jones, G.M., Wagner, P.E., Boman, R.L., Troutt, H.F. Jr., and Lesch, T.N., 1982. A dairy cow body condition scoring system and its relationship to selected production characteristics, Journal of Dairy Science, 65, 495--501
Woodward, J.R., Van Oijen, M., Griffiths, W.M., Beukes, C.P., and Chapman, D.F., 2020. Identifying causes of low persistence of perennial ryegrass (Lolium perenne) dairy pasture using the Basic Grassland model (BASGRA). Grass and Forage Science, 75, 45--63. DOI: https://doi.org/10.1111/gfs.12464
Acknowledgements
Authors express gratitude to the farmer and his family who participated in this experiment, whose privacy is respected by not disclosing their names. This work was undertaken thanks to funding the by the Autonomous University of the State of Mexico (Universidad Autónoma del Estado de México) through grant UAEM 4487/2018/CI and the Mexican National Council for Science and Technology (CONACYT – Consejo Nacional de Ciencia y Tecnología) for the postgraduate grant for Mónica Muciño-Álvarez. Our appreciation also to Ms. Laura Edith Martínez-Contreras for her assistance in laboratory analyses.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interests.
Statement on ethical standards and animal rights
The paper reports an on-farm experiment undertaken with the active participation of a collaborating farmer who had knowledge of the objectives of the work and was duly informed at all times, and his privacy and that of his family is respected by not disclosing their names. On-farm research with collaborating farmers and experimental methods with dairy cows followed accepted research procedures by Universidad Autónoma del Estado de México.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Muciño-Álvarez, M., Albarrán-Portillo, B., López-González, F. et al. Multi-species pastures for grazing dairy cows in small-scale dairy systems in the highlands of Mexico. Trop Anim Health Prod 53, 113 (2021). https://doi.org/10.1007/s11250-021-02564-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s11250-021-02564-y